
Microbial and Plant-Based Compounds as Alternatives for the Control of Phytopathogenic Bacteria

1
Laboratory for Plant Protection, Department of Agriculture and Nutrition, Institute of Agriculture and Tourism, Carlo Hugues 8, 52440 Poreč, Croatia
2
Agricultural Institute of Slovenia, Hacquetova Ulica 17, 1000 Ljubljana, Slovenia
3
Department of Plant Pathology, Division of Phytomedicine, Faculty of Agriculture, University of Zagreb, Svetošimunska Cesta 25, 10000 Zagreb, Croatia
4
Skink d.o.o., Cesta Za Valaltu-Lim 20/c, 52210 Rovinj, Croatia
5
CITAB–Centre for the Research and Technology of Agro-Environmental and Biological Sciences, University of Trás-os-Montes and Alto Douro (UTAD), Quinta de Prados, 5000-801 Vila Real, Portugal
*
Author to whom correspondence should be addressed.
Horticulturae 2023, 9(10), 1124; https://doi.org/10.3390/horticulturae9101124
Submission received: 4 August 2023
/
Revised: 5 October 2023
/
Accepted: 10 October 2023
/
Published: 12 October 2023
(This article belongs to the Special Issue Advanced Research of Horticultural Plants Interactions with Bacteria and Fungi)
Abstract
:Plant pathogens pose a significant threat to agricultural productivity and food security worldwide. The use of traditional chemical pesticides for plant disease management raises concerns due to the emergence of pesticide resistance and their potential adverse effects on human health and the environment. As a result, there is a growing interest in exploring alternative approaches for plant disease control. This review provides an overview of the antimicrobial potential of some plant-derived compounds, including essential oils, plant extracts, wastes and their major constituents, against plant pathogenic bacteria. The antimicrobial activity is attributed to the diverse chemical composition of these plant-derived compounds and their ability to target multiple cellular processes in pathogens’ cells. Furthermore, the review explores the use of some antagonistic bacteria and fungi as control tools. These beneficial microorganisms have shown promising results in suppressing the growth of plant pathogens through various mechanisms such as competition, antibiosis and induced systemic resistance. This review discusses the advantages and limitations of using plant-derived compounds and antagonistic microorganisms for plant disease management. Moreover, it highlights the need for further research to optimize their efficacy, develop sustainable formulations and evaluate their performance under field conditions.
1. Introduction
In the vast realm of bacterial species, more than a hundred have been identified as plant pathogens, accounting for a fraction of the tens of thousands to billions of bacterial species known [1,2]. Among them, the most troublesome and widely distributed plant pathogenic bacteria primarily belong to the Gram-negative taxa Xanthomonas, Agrobacterium, Pseudomonas, Xylella and Erwinia. Gram-positive phytopathogenic bacteria are represented by genera such as Arthrobacter, Clavibacter, Curtobacterium and Rhodococcus. These bacteria exhibit a broad host range, infecting a variety of economically important herbaceous and woody plant species [2], resulting in reduced crop yields and diminished product quality. Although quantifying the exact magnitude of yield losses caused by bacterial infections is challenging, it is estimated that plant diseases account for 10–16% of total global plant production losses [2,3] and economical losses of over one billion dollars annually [4]. Notably, the genus Xanthomonas alone has been implicated in approximately 350 diseases affecting around 400 different plant species [5,6,7]. The relative importance of numerous phytopathogenic bacterial species with descriptions of specific hosts and diseases is discussed by Mansfield et al. [8]. Specifically, the most significant harmful bacteria from both economic and scientific perspectives are those belonging to the Pseudomonas syringae complex, Ralstonia solanacearum, Rhizobium radiobacter, different strains of Xanthomonas bacteria and Clavibacter michiganensis subsp. michiganensis.
The extent of damage that these bacterial species can inflict on agriculture largely depends on their epidemiology and the biological factors that determine their virulence. Bacteria can reside as epiphytes or endophytes in plants, leading to infections and manifesting in symptoms such as chlorosis, necrosis, knots, lesions and overall decline in plant health. These infections can sometimes remain latent or asymptomatic [9,10,11,12]. Predominantly Gram-negative plant pathogenic bacteria possess specific virulence factors that enable them to overcome plant defense mechanisms. These factors include toxin production, quorum sensing, two-component signal transduction (TCST) systems and other biological mechanisms [13,14,15,16]. To ensure their survival and successful infection, pathogenic bacteria have evolved strategies to manipulate the host defense responses by interfering with hormone signalling pathways. For instance, bacteria such as R. radiobacter and Pseudomonas savastanoi pv. savastanoi synthesize cytokinins and auxins, which serve as carbon and nitrogen sources, while also inducing infection symptoms. These hormones can disrupt normal plant growth and development [13]. Another example is P. syringae, which produces a phytotoxin called coronatine. Coronatine enhances bacterial growth and facilitates systemic spread within the host by mimicking the plant hormone jasmonic acid (JA). Interestingly, JA typically functions in plant defense responses against herbivores and necrotrophic pathogens. However, by producing coronatine, bacteria suppress the synthesis of salicylic acid (SA), a plant hormone that is effective against biotrophic and hemibiotrophic pathogens such as bacteria [17,18]. Plant pathogenic bacteria are predominantly classified as hemibiotrophic, displaying a lifestyle in which they initially infect and colonize living host tissue and subsequently transition to surviving in dead plant tissue. However, exceptions to this pattern exist, such as the tumor-inducing bacterium of the genus Agrobacterium/Rhizobium, which exclusively thrives in living plant tissue [14]. From an epidemiological perspective, successful plant pathogenic bacteria possess a range of essential traits for their survival and establishment within their hosts. These traits also include motility, signalling mechanisms, adhesion capabilities and the ability to degrade plant-derived compounds.
Phytopathogenic bacteria coexist with beneficial bacterial communities within and on plant hosts that could interfere with the virulence level of pathogenic bacterial species and the severity of diseases. The interaction between pathogenic and beneficial bacteria has been suggested to influence the overall condition and health of plants. However, the precise association and interaction between the plant microbiota and plant health are still not fully understood [19,20]. The emerging field of socio-microbiology has been developed to study these interactions and associations among microorganisms, recognizing their importance as a valuable tool in biodiversity preservation [21]. Considering the coexistence of diverse microbial populations, the application of environmentally friendly preparations for plant protection has become a necessity. By exploring and understanding these complex microbial interactions, eco-friendly approaches can be developed to mitigate the impact of pathogenic agents on plants.
Bacterial disease management is faced with many limitations, one being the lack of effective registered bactericide preparations. Copper-based products can be used to control bacteria that are not resistant to copper, but they can have negative effects on beneficial bacteria and plants [22]. The major concern with the use of antibiotics in plant protection is the occurrence of multidrug resistance of bacteria through horizontal gene transfer or mutations, which have been reported in the literature [23]. To address these challenges, the cultivation of genetically modified plants is regarded as a potential strategy for disease control. Nevertheless, plant modifications, including genetic modifications, are subject to rigorous public scrutiny because of concerns about their perceived unnaturalness and possible hazards, analogous to the employment of synthetic pesticides in agriculture [3]. In response to the stringent modern demands and the environmental consequences associated with the use of synthetic pesticides and excessive nutrients, the development of alternative and environmentally friendly preparations is essential for sustainable agricultural production, one of the most important sectors of the Sustainable Development Goals [24]. Green preparations, which are environmentally and economically advantageous, offer a solution for agriculture and the control of plant diseases [25]. These preparations aim to provide effective disease control while minimizing environmental impact and promoting sustainable farming practices. Biopesticides are natural products that are categorized as microbial and biochemical biopesticides used to control pests and diseases. These preparations occupy a total of 5% of the global pesticide market with environmentally favourable properties such as biodegradation and selective action against targeted organisms. The largest share among biopesticides is those based on microorganisms, with approximately 90% of available preparations based on entomopathogenic bacterial species and few preparations against fungal and bacterial pathogens of vegetables, fruits and ornamental plants, e.g., Botrytis, Sclerotinia, Fusarium, Alternaria, Xanthomonas, etc. [26]. However, biochemical biopesticides may eventually prevail in the market, owing to their wider spectrum and stability of preparations in contrast to living microorganisms, which often encounter problems in practice because of their limited availability, slow action and high cost. Biochemical biopesticides could be formulated using essential oils (EOs), plant secondary metabolites, polyphenols, etc. [24]. Although plants employ diverse defense mechanisms to combat pathogens, polyphenols play a crucial role as constituents of the plant cell wall structure and act as barriers in the plant defense system [15,27]. Specifically, polyphenols exhibit toxicity towards bacteria and can inflict damage to microbial cell membranes [28], and thus are promising as active molecules for the development of biochemical biopesticidal products. Consequently, numerous studies highlight the potential application of EOs and plant extracts in plant disease control. According to a review by Wińska et al. [29], EOs have well-established antibacterial activity. Other studies demonstrate the antibacterial potential of plant waste products that are produced in large quantities in agricultural and food production industries, due to the high content of phenolic compounds with antimicrobial properties. Some of these are olive and grape wastes, and fruit and vegetable peels [30]. This review examines the antimicrobial potential of plant-derived compounds, such as EOs, plant extracts and their main constituents, against bacterial plant pathogens. It evaluates their effectiveness, explores their mechanisms of action, and assesses their suitability as eco-friendly alternatives for plant disease management. It also investigates the use of some antagonistic bacteria and fungi as control measures. The review aims to summarize the current knowledge, identify the most promising compounds and microorganisms and their sources, discuss their mode of action, address any limitations or challenges, and suggest future research directions in this field.
2. Antagonistic Microorganisms and Microbial Biopesticides
2.1. Potential of Some Antagonistic Fungal Strains against Phytopathogenic Bacteria
In recent studies, several filamentous fungi have been investigated as potential biological control agents against pathogenic microorganisms. These studies, comprehensively discussed by Poveda and Baptista [31], have highlighted the antibacterial effects of tested fungi. From the perspective of socio-microbiology, it has been observed that fungi compete with economically important bacteria such as Xanthomonas, Pseudomonas, Xylella fastidiosa and Rhizobium radiobacter through various mechanisms including parasitism, antibiosis and competition for space and nutrients. The presence of certain fungal species, mainly belonging to the genera Trichoderma, Epiccocum, Drechslera and Alternaria, has been found to positively impact the plant’s defense as they establish beneficial interactions with the plant. These fungi have the potential for the biological control of certain bacterial pathogens. In one study, the fungal strain of Fusarium tricinctum and its metabolites demonstrated antibacterial potential against P. syringae pv. actinidiae (bacterial canker in kiwifruit). The fungus alone suppressed bacterial growth by 59.5% in vitro, while its metabolites (imidazole alkaloids and enniatins) were active at MIC range from 25 to 50 µg/mL [32]. This finding highlights the potential of fungal-derived compounds as effective agents against bacterial pathogens. Another study [33] investigated the antibacterial activity of 17 morphotypes of orange endophytic fungi, where several strains belonging to the genera Fusarium, Diaporthe, Colletotrichum, Geotrichum, Penicilium and Polyporus exhibited significant antibacterial activity against X. citri subsp. citri, the causative agent of citrus canker. Notably, Geotrichum sp. (gc-1-127-30) and Diaporthe biconispora (gc-1-128-79) strains showed the strongest efficacy for inhibiting bacterial growth with MIC of 62.5 and 31.3 µg/mL. Interestingly, individual compounds of D. biconispora extracts were significantly variable as antimicrobials, showing a MIC range from 12.5–>1000 µg/mL. The lowest MIC was determined for an unidentified compound (compound 2), suggesting the need for its further identification and evaluation as a control agent of pathogenic bacteria. Trichoderma is one of the most commonly encountered genera of biocontrol fungi on the market.
While it exhibits some level of antibacterial activity in vitro, T. longibrachiatum is effective against various foodborne bacteria. However, it also suppresses the growth of beneficial bacteria such as P. fluorescens and Bacillus subtilis, which benefit the host by inducing stress resistance, antibiosis and metabolite synthesis [34]. Moreover, the use of trichokonins derived from T. koningii SMF2 has been reported to inhibit the growth of the Gram-positive bacterium C. michiganensis subsp. michiganensis, the causal agent of bacterial wilt in tomato. However, its efficacy has been limited against Gram-negative plant pathogens such as Ralstonia solanacearum and Erwinia carotovora [35]. These studies demonstrate the diverse antimicrobial potential of various fungal species and their metabolites against bacterial pathogens. Notably, some fungal strains showed a selective action against targeted bacterial strains, and thus could be implemented as control agents without interfering with natural populations of beneficial microorganisms. Further research is needed to explore and evaluate their efficacy in controlling bacterial diseases in agricultural and horticultural settings.
2.2. Potential of Some Antagonistic Bacterial Strains and Bacteriophages against Phytopathogenic Bacteria
Plant growth-promoting bacteria (PGPB), such as certain strains of Streptomyces, Bacillus and Pseudomonas, have been extensively studied for their ability to promote plant growth and act as biocontrol agents against plant pathogens. Among the antagonistic bacteria studied for their suppressive effects on phytopathogenic organisms, the genera Pseudomonas, Bacillus, Azospirillum and Streptomyces have received significant attention. P. fluorescens and Streptomyces spp. are among the most extensively studied biopesticide bacteria. Some Streptomyces strains produce antibiotics that can suppress the growth of plant pathogens, while Bacillus and Pseudomonas strains produce other secondary metabolites such as phytoalexins to inhibit the growth of microorganisms [36]. These bacteria have been found to also stimulate the natural resistance of host plants while exerting antibiosis and competing for nutrients and space with pathogens [37,38,39,40,41,42,43,44,45]. Soil-dwelling bacteria of Streptomyces spp. are known for their production of diverse antibiotics, including streptomycin. Streptomycin has been widely used to control bacterial diseases, particularly for managing the fire blight of apples and pears in antibiotic-approved agricultural regions [46]. The bacterium P. fluorescens produces metabolites and enzymes that reduce the growth of pathogenic bacteria and fungi in the soil and on the surface of plant organs, which is a characteristic trait of fluorescing Pseudomonas species. In a recent study [47], the researchers investigated the antagonistic properties of bacterial isolates obtained from healthy olive leaves against a Pseudomonas savastanoi pv. savastanoi. Out of the 46 isolates tested, five exhibited antagonistic activity in vitro. Notably, Bacillus sp. Og2 and P. fluorescens Oq5 showed the highest inhibition of P. savastanoi growth. Furthermore, the secondary metabolites of these isolates demonstrated inhibitory effects after approximately 6 days of incubation. These findings align with numerous studies on the antagonistic activity of bacteria against various plant pathogenic bacterial species, including R. solanacearum, X. axonopodis pv. malvacearum, E. amylovora, R. radiobacter, P. syringae, X. arboricola, Pectobacterium carotovorum and Clavibacter michiganensis, on different plant hosts, e.g., apple, pear, cotton and apricot [48,49,50,51]. Previous studies have suggested that the antagonistic activity of P. fluorescens may be attributed to the secretion of minor phenolic compounds [49].
In recent discussions, the effective use of antagonistic bacteria, such as B. subtilis 2515-1 against P. savastanoi pv. savastanoi, the causal agent of olive knot disease, has gained attention [52]. These antagonistic bacteria have been found to produce various enzymes and secondary metabolites, resulting in a greater reduction in knot weight compared with standard copper-based preparations. Specifically, B. subtilis 2515-1 reduced the knot weight by 50%, whilst a reduction of 37% was achieved using 0.5% copper sulphate preparation. Notably, the antagonistic bacterial strain B. subtilis F1 has shown relative heat resistance, which is advantageous in regions with high temperatures (up to 40 °C) during the vegetation period [53], suggesting the further examination of promising Bacillus species to be implemented as control agents in arid regions. Additionally, other antagonistic strains, including B. megaterium, P. koreensis and B. pumilus, have been examined recently for their inhibitory effects on the causal agent of olive knot disease, with inhibition of knot formation ranging up to 65% [54]. However, the efficacy of bacterial inhibition is strongly dependent on the specific bacterial strain tested in vitro, in planta and in vivo [20,52]. In a study by Mina et al. [20], B. amyloliquefaciens exhibited the highest antagonistic potential among 27 bacterial strains, inhibiting the growth of P. savastanoi pv. savastanoi by 26.8% and reducing overall disease severity by 43.7% in in planta experiments. Furthermore, B. amyloliquefaciens demonstrated beneficial effects on the host, promoting dry shoot weight (+55%) and root water content (+39.6%). Thus, B. amyloliquefaciens shows promise as an efficient ecological solution for the management of olive knot disease, which is prevalent in olive-growing arid regions worldwide. Moreover, in a recent study by Broniarek-Niemiec et al. [55], the application of commercial preparations containing bacterial strains, specifically B. amyloliquefaciens, demonstrated inhibitory effects on the incidence of bacterial canker caused by P. syringae in cherry orchards in Poland over two growing seasons. Although the efficacy of tested microbial-based preparations was lower than conventional copper and aluminium-fosetyl-based treatments, their effectiveness was still observed.
Bacteriophages, also known as phages, are viruses that specifically target and infect bacteria. They can be employed as a biocontrol agent to selectively eliminate pathogenic bacteria while preserving beneficial microbial communities. The use of bacteriophages shows promise in the field of bacterial disease management due to their high specificity and ability to evolve alongside bacterial populations. Phage-based preparations are used in some agricultural regions against X. campestris pv. vesicatoria and P. syringe pv. tomato, C. michiganensis subsp. michiganensis, E. amylovora and X. citri subsp. citri, as well as soft-root bacteria [56]. According to the review by Buttimer et al. [57], experiments conducted using bacteriophages are focused on various phytopathogenic bacterial species listed among the top 10 pathogenic bacteria in molecular plant pathology [8], such as Pseudomonas syringae pv. porri causing bacterial blight of leek, resulting in a reduction of disease severity in field trials. Moreover, phages were studied against Xanthomonas species, which are infectious to tomatoes, onions, grapefruit and oranges. Most of these studies were in planta or in vivo, and in the case of X. campestris pv. vesicatoria and X. euvesicatoria pv. allii, which cause bacterial spot and leaf blight on tomatoes and onions, respectively, field experiments show that the efficiency was comparable to the application of copper-mancozeb treatments. Efficient control of bacterial pathogens and decrease in disease severity was also determined against soft-root bacteria, Dickeya solani and Streptomyces scabiei, on potatoes and radishes, respectively, X. fastidiosa on grapevines and olives, R. solanacearum on tomatoes, P. carotovorum subsp. carotovorum on lettuce, X. axonopodis pv. citri on grapefruit, X. axonopodis pv. citrumelo on oranges, Pseudomonas tolaasii on mushrooms and E. amylovora on pear and apple trees [57,58].
2.3. Advantages and Disadvantages of Antagonistic Microorganisms in Plant Protection
The utilization of antagonistic bacteria as biopesticides offers promising prospects for sustainable plant protection strategies. By incorporating these bacteria into pesticide formulations such as sprays, dusts and granules, effective pest management can be achieved while simultaneously promoting plant health and natural resistance mechanisms. Ongoing research endeavours to explore the full potential of biopesticide bacteria in agricultural practices. However, it is worth noting that the production of preparations based on Pseudomonas species is limited by a lack of spore production, which challenges formulation preparation and long-term storage [36]. Overcoming this problem and developing suitable formulations for Pseudomonas species is crucial to ensure their viability and efficacy as biopesticides over extended periods of time. It is also important to consider the diversity of species and how they influence certain properties of the host plant. Using multiple species in preparations broadens its action spectrum [45], but it also lengthens the registration process that entails seven steps, namely: isolation of microorganisms, characterization; determination of efficacy under controlled conditions; determination of efficacy under semi-controlled conditions; determination of efficacy in the field; standardization of the product and production; and, ultimately, production of a commercial biopreparation and launching the preparation on the market. On the other hand, biopesticides also can be less effective than chemical pesticides, and their effectiveness can be strongly affected by environmental factors such as temperature, humidity and soil pH [36]. The application of beneficial bacteria in field conditions is also one of the limiting factors, in terms of the complexity of research which entails interaction studies between various environmental factors and different genotypes of bacteria and other organisms [59]. For example, in contrast to the study by Krid et al. [53], the study by Ahmed and Hasnain [60] determined the negative impact of high temperatures on the survival of beneficial bacteria of the genus Bacillus. Furthermore, it is important to investigate the competitiveness of biopesticidal strains such as Bacillus spp. against the non-pathogenic strain of Pantoea agglomerans, which showed a suppressive effect against P. syringae in a recent study [55]. On the other hand, there are cases where mutualistic bacterial species can contribute to the spread and severity of diseases. This phenomenon often occurs between two or more bacterial species. For example, in the case of olive knot disease and in contrast to results obtained in the study by Broniarek-Niemiec et al. [55], the severity of the disease can be increased by the presence of non-pathogenic endophytic bacteria such as P. agglomerans and Erwinia toletana [61,62]. Therefore, the possibility of cooperation or coinfection between pathogenic and non-pathogenic bacteria should be considered as a significant factor in disease management approaches. Although the mechanisms of action of various antagonistic microorganisms are well studied [36,56,63], ongoing research aims to further explore the optimization and utilization of these biopesticidal bacteria to enhance plant health and reduce the reliance on conventional chemical pesticides.
3. Antibacterial Activity of Some Plant Waste Products
3.1. Importance of Management of Agricultural Byproducts and Wastes
In agriculture, the generation of byproducts and wastes is inevitable, and their improper disposal represents a major source of pollution to soil and groundwater. Although this is a global problem, the implementation of sustainable and circular economy practices for managing agricultural waste is poor or lacking. These wastes mainly originate from four major sectors, including livestock manure, byproducts, fruit and vegetable residues and crop residues. Exploring the potential uses of agricultural by-products and wastes not only addresses the issue of pollution but also opens opportunities for potential applications in various disciplines. The potential use and environmental management perspectives of these agricultural wastes have recently been comprehensively discussed [64,65]. From an environmental management standpoint, these waste materials can be harnessed for their value in areas such as bioenergy production, organic fertilizer production, soil amendment and bioremediation. Furthermore, agricultural wastes are generally rich in phenolic content and therefore present a new source of substances that can be used to develop biopesticides [30,64,65,66,67,68,69,70,71,72,73,74,75,76,77,78,79].
3.2. Antibacterial Properties of Olive Mill Wastewater
Olive mill wastewater (OMWW), also known as olive mill effluent (OME), is recognized as one of the by-products that are characterized by its bioactivity and high content of phenolic compounds such as hydroxytyrosol glucoside, hydroxytyrosol, tyrosol, caffeic acid, verbascoside, oleuropein, luteolin, quercetin, apigenin, rutin and EDA-like components [67,68]. However, the management of OMWW presents a challenge in olive-growing regions due to its large volume, resulting in water and soil pollution caused by the concentrated phenolics [68]. Therefore, it is necessary to control waste disposal and utilize OMWW in sustainable ways. The amount of phenolic compounds can be influenced by storage conditions and duration. Proper storage and handling of OMWW are essential to preserve the phenolic content and maximize its future applications. For example, Feki et al. [69] observed that storing OMWW at 25 °C in the dark for five months resulted in a decrease in the amount of most phenolic compounds, while the amount of hydroxytyrosol increased. Further studies would need to explore the optimal storage conditions and duration to maintain the desired phenolic composition and bioactivity of OMWW. Despite the potential risks associated with its disposal, OMWW has shown promising results as an antimicrobial agent in some studies. Direct use of OMWW in its raw form is not recommended as it is toxic to humans, animals and the environment [70]. However, studies have also shown that even low concentrations of bioactive compounds can inhibit the growth of bacteria in laboratory experiments with its effect surpassing copper treatment [71]. For instance, OMWW has exhibited bactericidal effects on species such as P. savastanoi pv. savastanoi and C. michiganensis, using the original concentrations of polyphenols present in the wastewater [72]. Another study demonstrated the bactericidal activity of OMWW obtained from the Leccino olive cultivar against the seed-borne bacterium X. campestris, without interfering with seed germination. The MIC of total polyphenols was determined to be 2500 µg/mL for X. campestris. It was proposed that the bactericidal activity was a result of the interaction between polyphenols and bacterial proteins, leading to protein denaturation [73]. It is worth noting that the phenol oleuropein only inhibited the growth of the Gram-positive bacterium B. subtilis, suggesting that the concentrations of certain phenolic compounds may affect beneficial organisms present in/on plant hosts. Therefore, the extraction of individual compounds should be considered as an optimal solution to preserve non-pathogenic organisms and the biodiversity of microorganisms associated with various plant species. Furthermore, different olive cultivars were found to significantly differ in their total phenolic content, indicating that the antimicrobial activity among extracts may vary. Extracts obtained from cvs. Frantoio, Mission, Taggiasca and Picual were effective against some foodborne bacteria, including Staphylococcus aureus, Escherichia coli, P. aeruginosa and Listeria monocytogenes [74,75]. In a study by Krid et al. [71], the application of phenolic extracts from OMWW, which contained high concentrations of hydroxytyrosol, 3,4-dihydroxy phenylethanol (tyrosol), catechol, para-coumaric acid and caffeic acid, against P. savastanoi pv. savastanoi inhibited the formation of knots in olive plants (cv. Chemlali) significantly stronger than treatment with a copper-based preparation. Copper sulphate (0.5%) inhibited the weight of knots by around 50%. The effectiveness of extracts was attributed to the high concentrations of hydroxytyrosol. In contrast to these results, the study by Medina et al. [76] determined the antibacterial properties of numerous polymeric fractions against bacterial strains belonging to the genera Erwinia, Pseudomonas and Clavibacter. Briefly, the results showed that the antibacterial effect is likely the result of synergy among ingredients. For example, the ethyl acetate extract of wastewater showed significant inhibition of P. savastanoi and E. amylovora growth at concentrations ranging from 5 to 20%, respectively, while the wastewater itself exhibited MIC at concentrations from 10 to 20%, depending on the targeted bacterial species.
Antibacterial Properties of Phenolic Compound Hydroxytyrosol
Hydroxytyrosol (HTyr) is a promising low molecular weight phenol found in OMWW, as well as in olive oil and olive leaves. It has antimicrobial activity against bacterial species such as R. radiobacter and P. savastanoi pv. savastanoi, which are known to induce tumors in plants [77]. Strong bactericidal activity has been observed with the application of hydroxytyrosol alone, as well as in its enriched form within OMWW extract. However, when hydroxytyrosol was used alone, even at its highest concentration (32.83 mg/g) it exhibited the weakest inhibition of both bacteria. Similar results were obtained by Medina et al. [76], where hydroxytyrosol alone did not show any antibacterial properties when used against P. savastanoi, whilst the antibacterial potential was determined against E. amylovora only at a concentration of 6.6 mmol/L. A greater inhibition of both bacteria was achieved with the hydroxytyrosol-enriched extract at its highest concentration (30.4 mg/mL). The effectiveness of both treatments was found to be concentration dependent, with lower concentrations showing decreased antibacterial activity. The strong bactericidal potential of OMWW extract is presumed to be the result of the synergy among different phenolic compounds present alongside HTyr. However, the cost of hydroxytyrosol remains a major disadvantage, and it is not clear if the efficiency of its use justifies the expense. Nonetheless, due to the estimated non-toxic properties of HTyr, it is considered an interesting molecule in the development of environmentally friendly pesticides for the future.
3.3. Other Agricultural and Agro-Industrial By-Products and Wastes Showing Antibacterial Properties
In addition to olive wastes, the studies listed in Table 1 investigated the antibacterial potential of various agricultural residues, pest plants, solid wastes and food industry by-products against economically important phytopathogenic bacterial species. Generally, the antibacterial properties against Gram-negative bacteria were relatively weaker, requiring higher concentrations (>1000 µg/mL) of the antimicrobials compared with the Gram-positive bacteria. In the study by Ditsawanon et al. [78], the antibacterial efficacy of hydrolates derived from agricultural waste products was comprehensively studied. The research focused on the antibacterial potential of hydrolates obtained from selected agricultural waste materials, including bagasse and rice straw, and agro-industrial residues such as coconut residue from coconut milk production and peanut seed coat from peanut-based snack production. The results indicated significant inhibition of growth among specific phytopathogenic bacterial species (Table 1). Remarkably, the utilization of purified protein hydrolate extracted from bagasse—an ATP-binding cassette domain-containing protein—exhibited a distinct mechanism of action compared with other studied peptides. For instance, an unidentified peptide and expansin were observed to impact the integrity of the cell wall membrane. In contrast, the ATP-binding cassette domain-containing protein demonstrated a different mechanism, targeting intercellular biological processes. The findings of this study underscore the potential significance of peptides for agricultural applications, particularly in managing phytopathogenic bacteria. Importantly, ATP-binding cassette domain-containing protein did not inhibit the growth of beneficial bacterial species B. subtilis and P. fluorescens in vitro. However, it did inhibit the growth of Pectobacterium carotovorum, implying its potential for targeted control.
The antimicrobial potential of various plant by-products was tested in a recent study [79]. Tested by-products (Table 1), obtained from different parts and processes of cultivated and weed species, exhibited selective antibacterial properties. In particular, the efficacy of selected treatments was strong against the Gram-positive species C. michiganensis subsp. nebraskensis. The antibacterial potential was determined for the solid waste derived from the distillation of aerial parts of three species within the Salvia genus. The key bioactive compound that was determined as the molecule of interest was rosmarinic acid. In contrast, treatments poorly affected the growth of Gram-negative species P. syringae pv. syringae. These findings highlight the challenging identification of effective antibacterial agents against Gram-negative bacterial strains.
Interestingly, fruit peels, commonly generated as household waste, have been studied for their antibacterial effects [80]. Some examples of potential antibacterial agents from fruit peels mentioned in the literature include peanut shells [81], pomegranate peel [82] and mandarin peel [83]. Other potential antimicrobials, such as jabuticaba (Myrciaria cauliflora) peel, grape (Vitis sp.) pomace, peels and seeds, as well as prickly pear (Opuntia ficus-indica) wastes, have been highlighted in studies focusing on their activity against human pathogenic bacteria [30]. These studies examined the antibacterial properties of extract and found it to be effective against various pathogenic bacteria such as Klebsiella pneumoniae. While studies did not specifically focus on plant pathogens, they suggest the investigated extracts could act as antibacterial agents for plant diseases as well. Jabuticaba, known for its anthocyanin-rich fruit peel, possesses compounds that exhibit antimicrobial activity. Similarly, grape pomace, peels and seeds contain bioactive compounds, including phenolics, flavonoids and tannins, which exhibit antimicrobial properties. Additionally, prickly pear extracts have shown antimicrobial activity against human pathogens. These extracts contain bioactive compounds, including phenolic compounds, which also may have potential applications in plant disease management, although more research is needed to explore their specific effects on plant-associated microorganisms.
4. Antibacterial Activity of Essential Oils and Plants Extracts
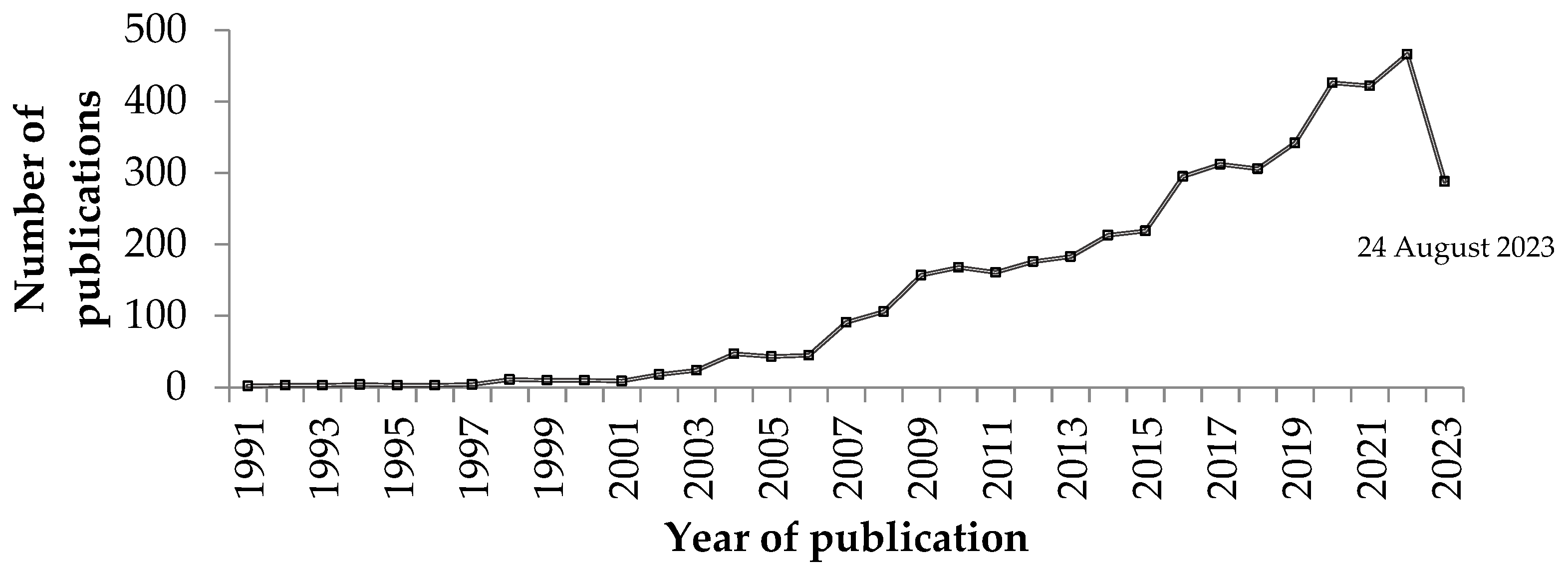
The urge to implement a new approach to controlling bacterial diseases is demonstrated by a total of 4570 scientific papers indexed in the Web of Science Core Collection, searched using the keywords “essential oil” and “plant bacteria” (Figure 1).
These papers discuss the antibacterial efficacy of numerous essential oils (EOs) against various plant, human and foodborne pathogenic bacteria. The research on this topic can be traced back to 1991, with an initial publication rate ranging from 2 to 18 papers per year until the early 2000s. However, since 2013, there has been a significant intensification of research efforts, resulting in a considerable increase in the number of published papers. In the past decade (2014 to present), there has been a consistent growth in research activity, with over 200 papers published annually. Over the last five years, the number of publications per year has ranged from 312 to 466, indicating a substantial interest and engagement in investigating the antibacterial properties of EOs. Notably, more than 50% of the listed papers were published between 2017 and 2023, indicating a recent surge in research focusing on this area. Plant extracts, particularly EOs, are known to encompass a diverse array of bioactive compounds, including phenolic compounds. These compounds offer potential alternatives for the control of phytopathogenic bacteria. Although the specific phenolic compositions may differ among plant extracts, they collectively contribute to the antimicrobial activity exhibited by these extracts. The increasing number of research papers in this field reflects the growing interest in seeking natural and environmentally friendly solutions for controlling bacterial pathogens.
4.1. Antibacterial Potential of Essential Oils and Essential Oil Compounds
4.1.1. Plant Species from the Lamiaceae Family
EOs have been investigated for their antimicrobial activity against a range of microorganisms, including fungi and human pathogenic bacteria, and as preservatives for fresh fruits [84,85,86]. EOs derived from plants are often readily accepted from a human perspective due to the traditional use of medicinal aromatic plants in many countries [87]. Chemical analyses of various plant species have revealed diverse plant metabolites, particularly high phenolic content, which have the potential for antimicrobial activity [88,89,90]. In addition to EOs, the most concentrated compounds that characterize the chemotypes of these oils are monoterpenoids and terpenoids with longer chains, such as menthol, geraniol, thymol, eugenol, carvacrol and others. These compounds contribute to the chemotypes of various plant species, including Citrus limon (lemon), Mentha x piperita (peppermint), Geranium sp. (geranium), Lavandula sp. (lavandin) and Cymbopogon citratus (lemongrass), among others [91]. The presence of specific compounds in EOs is dependent on the plant species. Commonly detected compounds in EOs also include α-phellandrene, limonene, d-limonene, carvacrol, thymol, α,β-thujone, camphor, carvone, menthol and menthone [92]. Some of these compounds have been shown to exhibit antimicrobial efficacy. Further research and exploration of the antimicrobial properties of these plant metabolites, both individually and in combination, can provide valuable insights into their potential applications in agriculture and plant disease management (Table 2). Notably, among the plant pathogenic bacteria, EOs derived from M. piperita (peppermint) have been frequently tested and shown antimicrobial potential. The use of EOs derived from Mentha plant species is expected to be more prevalent due to the presence of a high number of different species and cultivars within this genus. Studies suggest that various Mentha species are characterized by a high content of organic compounds with determined antibacterial activity, particularly phenols (terpenoids) such as eucalyptol, limonene, linalool, menthone, menthol, pulegone, linalyl acetate, carvone D, and more [90]. Nevertheless, the antibacterial potential of EOs is largely attributed to the synergy of various volatile compounds, rather than one specific compound [93,94]. According to the literature review by Wińska et al. [29], the EOs derived from Mentha species exhibit low antibacterial efficacy, except if the menthol is highly concentrated in oils. However, it is proposed that the Mentha EO can be used in combination with other EOs to induce higher sensitivity of bacteria to some antibiotics [95]. In the same study, carvacrol–thymol chemotypes of EOs of Thymus and Origanum species are highlighted. Moreover, combinations or synergistic effects of EOs, such as those derived from Thymus and Origanum species, have been found to exhibit antibacterial properties when applied undiluted, with higher susceptibility determined for Xanthomonas strains compared with Pseudomonas or Erwinia taxa [96]. Particularly low MIC values listed in Table 2, indicate high efficacy of Thymus and Origanum EOs as antimicrobials [96,97,98,99,100,101,102]. In the study by Bozkurt et al. [98], as well as previous studies, the EOs derived from Thymus and Origanum species, which are primarily composed of thymol and carvacrol, demonstrated the strongest inhibition of bacterial growth in vitro. Interestingly, the bioactivity against P. savastanoi pv. savastanoi was significantly greater compared with P. savastanoi pv. nerii, which are closely related pathovars known to induce tumors in host plants. The carvacrol chemotype Origanum EO was the most efficient antibacterial in the study by Todorović et al. [103], where the lowest MIC values (0.02–0.32 µg/mL) were determined against Pseudomonas, Xanthomonas and Clavibacter strains. Other EOs obtained from plants belonging to Lavandula, Citrus and Salvia taxa were effective at concentrations > 0.32 µg/mL. However, there are contrary observations in the literature, where the EO obtained from Satureja hortensis (summer savory), characterized by high concentrations of carvacrol (79.17%), showed lower antibacterial potential compared with the EO of Calamintha nepeta (calamint) with the highest concentrations of cis-Piperitone epoxide (48.66%) [104]. Generally, it was determined in in vitro studies that the application of EOs exhibits higher MIC values against species belonging to Pseudomonas genera than those belonging to Erwinia, Ralstonia, Xanthomonas, Clavibacter, etc. [96,102,103,104]. These findings suggest further investigation of specific biological mechanisms that may differentiate these species. Also, the observed variability in antibacterial activity highlights the complexity of interactions between EOs and bacterial strains within the Pseudomonas syringae complex. A lower efficiency against P. savastanoi compared with R. radiobacter was observed in the study by Caparrotta et al. [93], suggesting that susceptibility to the same concentrations of plant-based compounds, e.g., EOs, can significantly vary among targeted strains. Further, the EOs derived from basil, sage, rosemary and marjoram have shown relatively lower effectiveness against Gram-negative Pseudomonas and Xylella species, for example [97,105]. Specifically, the MIC of Salvia sclarea (clary sage) against X. fastidiosa was 500 µg/mL, because of which it is categorized as a weak antimicrobial [97]. Nonetheless, EOs from sage (Salvia) species are enumerated as promising antibacterial compounds as alternative antimicrobials due to determined antibacterial potential depending on specific plant species, suggesting that further evaluation and development of biopesticides derived from Salvia species may be worth exploring. Importantly, the plant’s chemical profile highly depends on genetics, climate and cultivation practice, thus suggesting the optimization of cultivation and identifying the most promising cultivar for further evaluation of less effective EOs, which could potentially act in synergy with some conventional pesticides or antibiotics. The oils from plant species belonging to the Lavandula genus can reduce antibiotic resistance of numerous human pathogenic bacterial species [106].
Of interest, O. vulgare (oregano) EOs were non-toxic to human neuroblastoma cell lines (IC50 = 50.5 µg/mL) [99], and therefore could be considered as safe compound for further evaluation for control of bacterial diseases in agriculture. However, it is important to mention that EOs and their compounds can exhibit phytotoxic effects on plant hosts [107], which highly depends on used concentrations [99,107]. Hence, future studies need to evaluate the phytotoxic effect of potential antibacterials to avoid damage to plants in the field. Also, studies warrant the investigation of combinations of some EOs to expand the spectrum of potential plant protection preparations, due to documented interaction between plant species—bacterial species—compound.
4.1.2. Plant Species Other Than Lamiaceae
Outside of the well-studied species of Lamiaceae, EOs derived from olive, almond and neem have demonstrated high efficacy against some human pathogens and could be potential pesticide candidates against plant pathogenic bacteria [108]. In the study by Sánchez-Hernández et al. [109], it was determined that Ginkgo biloba (ginkgo) EO exhibited strong antibacterial activity. At a concentration of 2 times MIC (1000 and 1500 μL/mL), the EO completely inhibited infection of C. michiganensis subsp. michiganensis in tomato plants and P. syringae pv. pisi in pea plants. Remarkably, no phytotoxicity was observed in any of the treatment groups, indicating the safety of G. biloba leaf extract as a natural pesticide. In the study by Tarakanov and Dzhalilov [100], two EOs obtained from Cinnamomum cassia (Chinese cinnamon) and O. vulgare (oregano), inhibited the growth of P. savastanoi pv. glycinea and Curtobacterium flaccumfaciens pv. flaccumfaciens in vitro and in vivo at MIC of 200 and 1600 µg/mL, respectively. In foliar treatments or direct application on infected seeds at their MBC values, the inhibition of disease severity was more than 50%, without phytotoxicity. The antibacterial properties of cinnamon and oregano EOs were attributed to the presence of cinnamaldehyde and carvacrol compounds. Specifically, these EO treatments were found to be more effective than the standard fungicide–bactericide Kocide. It is worth noting that the phytotoxicity of the applied treatments in vivo followed a dose-dependent pattern, with a safe threshold of 0.5% solution of EOs, below which no adverse effects on plant health were observed. Another study [2] determined the antibacterial potential of the EO of Tagetes minuta (khaki weed). This EO showed remarkable inhibitory activity against Xanthomonas and Pseudomonas bacteria at higher concentrations of treatment. Interestingly, the MIC value was lower for the Pseudomonas strain compared with Xanthomonas, indicating higher sensitivity of Pseudomonas to this EO. Furthermore, the EO of T. minuta exhibited stronger activity compared with the standard broad-spectrum bactericide Enrich BM (immunomodulator2-bromo-2-nitropropane1,3 diol). Minimal bactericidal concentrations of 950 µg/mL were determined for X. axonopodis pv. phaseoli (causal agent of bean blight) and P. savastanoi pv. phaseolicola (causal agent of halo blight of beans), while a bactericidal concentration of 1900 µg/mL was determined for X. axonopodis pv. manihotis (causal agent of cassava bacterial blight). Overall, all tested Gram-negative bacterial strains exhibited high sensitivity to the application of T. minuta crude oil. The antibacterial effectiveness of T. minuta EO is attributed to its high concentrations of monoterpenes and sesquiterpenes. Moreover, prolonged incubation of bacteria (48 h) further enhanced the inhibitory effect against Pseudomonas bacteria. These findings suggest that T. minuta EO possesses potent antibacterial properties, particularly against troublesome Pseudomonas strains, which is interesting since in the literature, the strongest antibacterial effect of plant compounds is often reported against Gram-positive strains. Moreover, the application of Laurus nobilis L. (bay laurel) EO showed interesting results [110]. Specifically, the undiluted EO was more selective towards P. syringae pv. pahseolicola, with less antibacterial effect against the Gram-positive B. subtilis.
In addition to Xanthomonas species, the EO of Citrus aurantium (sour orange) and Cymbopogon winterianus (lemongrass) were effective against X. citri subsp. citri with MIC values of 0.238 and 0.121 µL/mL, compared with Foeniculum vulgare (fennel), Pinus elliottii (pine) and Ocimum gratissimum (basil) with MIC of 1.81, 7.81 and 0.43 µL/mL [94]. Also, Janaćković et al. [111] determined that the EO extracted from flowering Ambrosia artemisiifolia (common ragweed) exhibited pronounced activity against several strains of Gram-negative phytopathogenic bacteria listed in Table 2. This finding suggests the potential use of this allergenic weed species and its EO in the control of plant diseases. The study highlights the possibility of utilizing natural resources, such as weed species, as a source of bioactive compounds. Similarly, in a study by Bouchekouk et al. [112], the EO obtained from Pteridium aquilinum (bracken fern) demonstrated high potency against three economically important phytopathogenic bacteria. This EO was characterized by a high concentration of the compound linalool. These results support the previous literature reports on the potential of linalool as a molecule of interest for the control of bacterial diseases. Regarding individual molecules, the terpenes (p-cymene, y-terpinene and limonene) exhibit lower or no antimicrobial efficiency [113]. However, p-cymene can enhance the antimicrobial potential of certain terpenoids, such as carvacrol, by affecting bacterial membranes and flagellar movement [105]. Another group of compounds, phenylpropenes, including eugenol, vanillin and cinnamaldehyde, have shown notable antibacterial activity against Gram-negative bacteria, with isoeugenol and eugenol exhibiting greater efficacy [114].
Promising EO against Rhizobium radiobacter and E. carotovora var. carotovora was derived from Thuja occidentalis (white cedar) exhibiting lower MIC values (400 and 350 mg/L), compared with other 17 EOs tested [115]. Nevertheless, most EOs in the same study showed similar MIC values against both bacterial strains, while the EO of Myrtus communis (common myrtle) was defined as weak antimicrobial (MIC > 1000 µg/mL). Notably, it is demonstrated that the EOs with diverse compounds in their chemical profile, showed stronger inhibition of bacterial growth. Specifically, in a study by Feizi et al. [116], the EO of Ziziphora clinopodioides showed greater antibacterial properties with a MIC value of 6 µL/mL against P. syringae pv. syringae compared with thymol chemotype Carum capticum EO with a MIC of 18 µL/mL. The effectiveness of these EOs is attributed to the synergy of compounds in their chemical profile. Conversely, [94] a stronger antibacterial effect was determined for EOs that have a simple composition. Overall, these contradicting observations warrant further investigation of individual compounds alone or in mixtures to confirm which compounds could act in synergy or interact as antagonists.
Further, Amyris balsamifera (Indian sandalwood) and Pogostemon patchouli (patchouli) EOs showed activity against X. fastidiosa, with inhibition of biofilm formation by 50% using concentrations of 250 and 125 µg/mL, suggesting their moderate antibacterial potential [97]. The most concentrated compounds of these EOs were α-Gurjuene and γ-eudesmol. Even though the bacterial biofilm is less sensitive to antimicrobials, its inhibition by 50% suggests further research targeting other bacterial species. For this, further research on the mode of action is necessary. An additional suggestion is testing compounds in mixtures/combinations to broaden the spectrum and the mode of action as suggested by Patel et al. [102]. For example, a compound that disrupts cell membranes could allow better penetration of compounds that interfere with intercellular processes in bacterial cells.
4.2. Antibacterial Properties of Plant Extracts
Generally, EOs show higher inhibition of bacterial growth compared with plant extracts. In the case of C. michiganensis subsp. michiganensis, the antibacterial activity of pure M. suaveolens (apple mint) and C. ladaniferus (gum rockrose) EOs was slightly higher than treatment with antibiotics [87]. However, EOs inhibited the germination of tomato seeds, while the plant extracts were less effective but did not significantly reduce germination. Inhibition of seed germination by application of EOs contradicts the previously mentioned use of OMWWs as antimicrobials against C. michiganensis species [71]. It is proposed that the inhibition of germination might be attributed to allelopathy [87]. Overall, it is concluded that methanolic plant extracts of C. ladaniferus can be used as an alternative control measure against C. michiganensis on tomato seeds without interfering with germination, despite their lower total phenolic content. The antibacterial activity of the plant extracts of C. ladaniferus is attributed to the presence of sesquiterpenoid viridiflorol, which has been previously suggested as a safe natural pesticide in agriculture [117].
In the study by Fontana et al. [118], the efficacy of leaf extracts from Moringa oleifera (moringa) was evaluated against E. amylovora, the causal agent of fire blight in apple and pear trees. Application of leaf extracts significantly reduced the symptoms of infection, particularly when the treatments were applied prior to the onset of infection on apple shoots. This finding suggests the potential use of M. oleifera as a natural alternative for the management of fire blight disease. By reducing the symptoms of infection, these extracts have the potential to contribute to the protection and health of widely cultivated apple trees. In another study, a crude extract obtained from Annona atemoya (atemoya), which was characterized by a high concentration of the phytochemical rutin, demonstrated strong activity against the gall-forming bacterium R. radiobacter [119]. In another study [100], a strong antibacterial effect was determined using ethanolic extract of Bergenia crassifolia (leather bergenia) against two economically important pathogens of soybean—P. savastanoi pv. glycinea and C. flaccumfaciens pv. flaccumfaciens. The antibacterial effect was attributed to the presence of 5-Methyl-3-methylenedihydro-2(3H)-furanone. A MIC of 1000 to 2500 µg/mL was determined against both bacteria, with lower values determined against P. savastanoi pv. glycinea. Although the plant compounds are often phytotoxic, when these treatments were applied in vivo using 0.5% solution of EOs and 13% of plant extracts, no damage was observed on plants. In addition, these results confirm the strong activity of EOs at low concentrations, thus suggesting their use as plant protection agents that could be implemented in formulations.
Moreover, the hydroalcoholic extract of Larrea tridentata (creosote bush) [120], showed a strong antibacterial effect against multidrug-resistant strains of C. michiganensis subsp. michiganensis, P. syringae and X. campestris in vitro. The MIC of extract alone, as well as the aqueous (LTAq-F) and organic fraction (LTEtOAc-F) of extract, ranged from 0.39 to 6.25 mg/mL. The lowest MIC and minimal bactericidal concentration (MBC) against targeted bacterial strains was lowest for L. tridentata organic fraction. Interestingly, the plant extract and organic fraction were bactericidal in nature, conversely to the application of the aqueous fraction. Also, the authors highlighted that the stronger efficiency of the organic fraction was due to a higher yield of bioactive compounds. The observed antibacterial activity was proposed to be influenced by the presence of lignans, which exhibit various mechanisms of action that can interfere with cell membranes and protein transport systems, leading to cell death. Also, it could be assumed that the antibacterial effect was influenced by the presence of the phenols thymol and carvacrol, as these compounds are constituents of numerous EOs and are often determined as the most efficient agents against pathogenic bacteria [120]. These findings highlight the potential use of an organic fraction of L. tridentata extract in the control of drug-resistant bacterial strains in agriculture. In the overall context of plant extract use, it is worth considering the extraction process type and solvent used, as alcoholic extracts often result in a stronger antibacterial effect than aqueous extracts.
4.3. Mechanism of Action of Plant-Based Compounds against Gram-Positive and Gram-Negative Bacterial Species
The antibacterial activity of EOs is believed to be attributed to the synergy of various mechanisms that target bacterial cells, giving rise to what is known as the “essential oils versatility”. EOs can exert toxicity on bacterial cells by causing cell degradation, cytoplasmic coagulation, leakage of metabolites and ions from the cytoplasm, alterations in proton motive force and membrane fatty acids, increased membrane permeability, disruptions in intra- and extracellular ATP and ATPases, and distortions in membrane proteins. They can also interfere with anti-quorum sensing mechanisms, leading to a decrease in proteolytic activity, biofilm formation and inhibition of factors involved in bacterial virulence [105,121]. Hydrophobic chemical components found in EOs often exhibit lower antibacterial activity against Gram-negative bacteria compared with Gram-positive bacteria. This is due to the presence of a complex lipopolysaccharide outer membrane in Gram-negative bacteria, which can block or delay the penetration of EOs through the membrane [114]. However, it is important to note that the outer membrane of Gram-negative bacteria is not completely impermeable, allowing some penetration of hydrophobic molecules. In contrast, Gram-positive bacteria have a more permeable membrane to hydrophobic compounds, making them more sensitive to EO and plant extract application [105]. Additionally, EOs from plant species belonging to the Lamiaceae family have demonstrated strong antibacterial activity against plant pathogenic bacteria even at extremely low concentrations. The efficacy of treatments at lower concentrations is attributed to their impact on cellular enzymes, while higher concentrations can lead to protein denaturation [105]. It is important to note that the antimicrobial activity of EOs and plant extracts can vary depending on the specific bacterial strain, concentration, and composition of the oil, due to the variabilities in aromatic and phenolic profile which depends on climate, genetics, environment and agricultural practice [118,122]. The mode of action of EO components is highly dependent on the specific position of functional groups within their molecular structures, such as hydroxyl and alkyl groups. Additionally, the effectiveness of EOs can vary among different bacterial strains, highlighting the importance of investigating the virulence and pathogenicity of individual strains for effective management. This variability is exemplified in a study conducted by Hsouna et al. [123], where the antibacterial properties of the menthol chemotype of M. piperita EO exhibited significant variations in efficiency against different strains of R. radiobacter (with MIC ranging from 10 to 12,500 µg/mL). The study also confirmed that the mechanism of action of Mentha EO involved the disruption of the bacterial membrane, even at concentrations as low as half of the MIC. Furthermore, in vivo application of Mentha EO resulted in complete prevention of disease occurrence in tomato plants inoculated with R. radiobacter.

{kind=link}
Table 2.
List of plant species tested for their antibacterial potential expressed as minimal inhibitory concentration (MIC) of their essential oils (EOs) and extracts against various plant pathogenic bacterial species studied.
Table 2.
List of plant species tested for their antibacterial potential expressed as minimal inhibitory concentration (MIC) of their essential oils (EOs) and extracts against various plant pathogenic bacterial species studied.
| Reference | Source | Bacterial Species | Plant Species | MIC | Determination Method | Mode of Action |
|---|---|---|---|---|---|---|
| [100] | Tarakanov and Dzhalilov | Pseudomonas savastanoi pv. glycinea Curtobacterium flaccumfaciens pv. flaccumfaciens | EOs Cinnamomum cassia (L.) J. Presl Thymus vulgaris L. Origanum vulgare L. Mentha longifolia (L.) Huds. Mentha piperita L. Syzygium aromaticum (L.) Merr. & L.M.Perry Lavandula angustifolia Chaix ex Vill. Mill. Achillea millefolium L. Allium sativum L. Citrus aurantiifolia (Christm.) Swingle Elettarria cardamomum (L.) Maton Citrus reticulata Hort. Pimpinella anisum L. Foeniculum vulgare Mill. Salvia officinalis L. Extracts Bergenia crassifolia (L.) Fritsch-ethanol Mellisa officinalis L.-ethanol Capsicum annuum L.-ethanol Sambucus nigra L.-ethanol Phytolacca americana L.-ethanol C. annuum-water Galega officinalis L.-water Arthemisia absinhtium L.-ethanol Phellodendron amurense Rupr.-ethanol Rosa pendulina L.-ethanol Chelidonium majus L.-ethanol Morus nigra L.-ethanol Spiraea salicifolia L.-water | 0.200–>3200 µg/mL 1000–100,000 µg/mL | Disc diffusion Broth microdilution | n.t. |
| [98] | Bozkurt et al. | Rhizobium radiobacter P. savastanoi pv. savastanoi P. savastanoi pv. nerii | Thymus sipyleus Boiss. Rosmarinus officinalis (L.) Schleid. Mentha spicata L. Laurus nobilis L. Origanum majorana Boiss. L. Ocimum basilicum L. M. officinalis Salvia officinalis L. Thymbra spicata var. spicata L. Lavandula stoechas L. subsp. stoechas Origanum syriacum L. Thymus Serpyllum L. Foeniculum vulgare Mill. EOs | undiluted | Disc diffusion | n.t. |
| [123] | Hsouna et al. | R. radiobacter | Mentha piperita L. EO | 10–12,500 µg/mL | Disc diffusion Broth microdilution | Disruption of cell membrane |
| [119] | Al-Baharwee et al. | R. radiobacter | Annona atemoya Mabb. | 15.6–31.3 µg/mL | Agar well diffusion Broth microdilution | Damage to cell membrane |
| [2] | Muthee-Gakuubi et al. | Xanthomonas axonopodis pv. phaseolicola X. axonopodis pv. manihotis P. savastanoi pv. phaseolicola | Tagetes minuta L. EO | 12,000–48,000 µg/mL | Disc diffusion Broth dilution | n.t. |
| [87] | Benali et al. | Clavibacter michiganensis subsp. michiganensis P. savastanoi pv. savastanoi | Mentha suaveolens L., Cistus ladanifer L. EOs and methanolic and ethanolic extracts | 190–780 µg/mL | Disc diffusion Broth microdilution | n.t. |
| [111] | Janaćković et al. | X. campestris pv. campestris Erwinia amylovora P. syringae pv. syringae X. arboricola pv. juglandis | Ambrosia artemisiifolia Michx. L.EO | 4–1,500,000 µg/mL | Broth microdilution | n.t. |
| [120] | Morales-Ubaldo et al. | C. michiganensis subsp. michiganensis P. syringae X. campestris | Hydroalcoholic extract Larrea tridentata (DC.) Coville | 3120–8000 µg/mL | Disc diffusion Broth microdilution | Affect proteins of the ATP binding cassette transport system |
| [112] | Bouchekouk et al. | E. amylovora P. carotovorum subsp. carotovorum P. savastanoi pv. savastanoi | Pteridium aquilinum (L.) Kuhn EO | 0.625–5.00 µL/mL | Disc diffusion Agar diffusion | n.t. |
| [99] | Grul’ová et al. | C. michiganensis P. syringae pv. phaseolicola P. savastanoi X. campestris | Origanum vulgare L. EO | 100–10,000 µL/mL | Disc diffusion | n.t. |
| [93] | Caparrotta et al. | R. radiobacter P. savastanoi pv. savastanoi | Boswellia papyrifera (Delile ex Caill.) Hochst. B. frereana Birdw. B. dalzielli Hutch. B. rivae Engl. B. neglecta S.Moore Canarium madagascariensis Engl. C. schweinfurtii Engl. Bursera bipinnata (DC.) Engl. B. microphylla A.Gray Commiphora guidotti Chiov. C. wildii Merxm. Hymenaea verrucosa Gaertn. EOs | 0.2% (v/v) | Overnight incubation in liquid medium without dilution | n.t. |
| [97] | Brentini Santiago et al. | Xyllela fastidiosa | Pogostemon patchouli Pellet. Amyris balsamifera L. | 125 µg/mL | Broth microdilution Microtitration plate assay | n.t. |
| Cinnamomum zeylanicum Blume Cedrus atlântica (Endl.) G.Manetti ex Carriere Commiphora myrha (T.Nees) Engl. Cupressus sempervirens L. Citrus paradisi Macfad. Boswellia carterri Flueck. Citrus aurantium L. Salvia sclarea L. Thymus vulgaris | 500 µg/mL | |||||
| Citrus bergamia Risso & Poit. Eucalyptus globulus Labill. Zingiber officinale Roscoe Cinnamomum camphora (L.) J.Presl Abies sibirica Ledeb. Melaleuca alternifolia (Maiden & Betche) Cheel | 1000 µg/mL | . | ||||
| [96] | Vasinauskiene et al. | E. carotovora subsp. carotovora X. vesicatoria P. marginalis pv. marginalis P. syringae pv. syringae P. syringae pv. tomato Bacillus sp. | Origanum vulgare Acorus calamus L. Achillea millefolium L. Achillea filipendulina Lam. Achillea cartilaginea Petri Carum carvi L. Mentha piperita | Undiluted | Disc diffusion | n.t. |
| [104] | Gormez et al. | R. radiobacter Bacillus pumilus C. michiganensis subsp. michiganensis Enterobacter intermedius Erwinia carotovora subsp. carotovora E. chrysanthemi P. cichorii P. corrugate P. fluorescens P. syringae pv. syringae P. syringae pv. phaseolicola P. syringae pv. pisi P. syringae pv. tabaci P. syringae pv. tomato Ralstonia solanacearum X. campestris pv. campestris X. vesicatoria | Calamintha nepeta (L.) Kuntze Satureja hortensis L. | 7.81–31.25 µg/mL | Disc diffusion Broth dilution | n.t. |
| [116] | Feizi et al. | P. syringae pv. syringae | Carum capticum Ziziphora clinopodioides Lam. | Undiluted | Disc diffusion Well diffusion Vapor phase test | n.t. |
| [110] | Mamoucha et al. | P. syringae pv. phaseolicola | Laurus nobilis L. | Undiluted and 1.25% (v/v) | Disc diffusion Well diffusion | n.t. |
| [102] | Patel et al. | P. cichorii P. syringae X. perforans | Nepeta cataria L. Origanum vulgare | 2.5–5.0 % (v/v) | Agar well diffusion | n.t. |
| [103] | Todorović et al. | P. tolaasii C. michiganensis subsp. nichiganensis X. campestris pv. phaseoli | Illicum verum Hooker Juniperus oxycedrus L. Eucalyptus globulus Labilardie Lavandula angustifolia Mill. Citrus lemon L. Cymbopogon flexuosus Stapf. Mentha piperita Origanum vulgare Pinus nigra L. Pinus pinaster Aiton Pinus silvestris L. Rosmarinus officinalis L. Salvia officinalis L. Abies alba Mill. Gaultheria procumbens L. | 0.02–>32 µg/mL of air | Well diffusion (volatiles) | n.t. |
| [109] | Sánchez-Hernández et al. | C. michiganensis subsp. michiganensis P. cichorii P. syringae pv. pisi P. syringae pv. syringae P. syringae pv. tomato X. vesicatoria | Hydroalcoholic extract: Ginko biloba L. | 500 µL/mL 750 µL/mL 750 µL/mL 750 µL/mL 750 µL/mL 1000 µL/mL | Agar dilution | n.t. |
| [115] | Badawy and Abdelgaleil | R. radiobacter E. carotovora subsp. carotovora | Artemisia Judaica L. Artemisia monosperma Delile Callistemon viminalis (Sol. ex Gaertn.) G.Don Citrus aurantifolia (Christm.) Swingle Citrus lemon Citrus paradisi Citrus sinensis (L.) Osbeck Cupressus macrocarpa Hartw. ex Gordon Cupressus sempervirens L. Myrtus communis L. Origanum vulgare Pelargonium graveolens L.’Her. Rosmarinus officinalis (L.) Schleid. Syzygium cumini (L.) Skeels Schinus molle L. Schinus terebinthifolius Raddi Thuja occidentalis L. Vitex agnus-castus L. | 350–>1000 µg/mL | Broth dilution | n.t. |
| [94] | Sauer et al. | X. citri subsp. citri | Citrus aurantium L. Cymbopogon winterianus Jowitt Foeniculum vulgare Gaertn Pinus eillottii Engelm Ocimum gratissimum L. | 0.121–7.81 µL/mL | Broth microdilution | n.t. |
n.t.—not tested.
4.4. Future Aspects of Potential Biopesticides in Agriculture
Stability and preservation are crucial in the practical application of biopesticides. To overcome the evaporative nature and enhance the stability of EOs, phenolic compounds and other plant-derived compounds, encapsulation in chitosan nanoparticles has been proposed as a promising solution for their application as antibacterial preparations in agriculture. Encapsulation ensures controlled release of the compounds over time and improves their stability by reducing evaporation and degradation [12,124,125]. Encapsulation has been found to reduce the MICs compared with the free forms of oils, indicating improved effectiveness [85,126]. The cost of extracting plant metabolites and the complexity of chemical synthesis present limitations to the widespread use of terpenoids and other plant compounds with antibacterial activity. Although the plant compounds are promising agents in the control of phytopathogenic bacteria, standardization of extraction is required with more studies to obtain reliable results across various agricultural and horticultural settings [77]. The extraction methods required for commercial production of plant extracts can be costly and demanding in terms of skilled personnel needed to obtain high-yield products [117]. However, due to the documented effectiveness of low concentrations of numerous bioactive compounds, it is possible to ensure preliminary low-cost preparations to determine their profitability in the agricultural economy.
5. Conclusions
Plant diseases cause significant yield losses in global plant production, which is a major concern considering the increasing human population and the demand for high-quality and high-yield food production. Also, farmers face challenges due to restrictions on the use of many active chemical compounds in plant protection. Public concerns about the toxicity of these compounds to the environment and human health have led to their limited use. Considering these demands, beneficial microorganisms show considerable prospects. The most promising results might be achieved by applying antagonist bacterial strains of the Pseudomonas and Bacillus genera. The advantage of microbial biopesticide use is its biodegradable nature and targeted action. In contrast, it is limited by the high cost of production, limited supply, slow action and reduced efficacy in field conditions. As an alternative, bacteriophages are emerging as effective antibacterials due to documented suppression of disease in in planta experiments, comparable to copper-based preparations. However, rapid evaluation of the resistance of phytopathogenic bacteria to bacteriophages has remained an important constraint.
Also, certain plant-derived compounds, such as hydroxytyrosol, have been determined to be safe for plants, supporting their potential use in agriculture. Furthermore, plant metabolites, particularly phenolic compounds present in EOs and plant extracts, exhibit significant antibacterial effects. The mechanism of action of EOs involves damaging bacterial cell membranes and interfering with essential cellular processes. The versatility of EOs is attributed to diversity in mode of action, often more potent against Gram-positive bacteria than Gram-negative. In general, EOs obtained from species belonging to the family Lamiaceae show the strongest potential as antibacterials. Purified EOs generally showed higher inhibition than plant extracts against numerous tested bacterial strains. When the use of plant extracts is considered, alcoholic extracts generally have a stronger effect than aqueous extracts, which warrants further research to determine their effect on plants and their long-term effect on soil and the environment. All plant-based compounds need to be stabilized for storage when considering wider use. Innovative approaches such as microencapsulation of EOs in chitosan have shown promise in enhancing their inhibitory potential against bacteria, their stability and controlled release in agricultural applications, offering potential solutions for more targeted and efficient antimicrobial treatments. Also, it is important to consider that single concentrations of plant-based compounds will inhibit bacterial growth differently depending on species and even the pathovar.
Overall, this review underscores the importance of the development of novel techniques to enhance the efficacy of antimicrobial compounds in the control of bacterial diseases in plants. Although extraction and the synthesis of plant metabolites and compounds can be costly and complex, their potential as an alternative to managing bacterial diseases in agriculture is evident. With a significant portion of global plant production losses attributed to bacterial diseases, there is a pressing need for effective and environmentally friendly biopesticides. We believe that this approach can contribute to the development of more efficient and targeted antimicrobial treatments in the future. Further research and optimization of application methods are needed to help establish the practicality in the context of plant disease management strategies in field conditions.
Author Contributions
Conceptualization and methodology, L.K.; validation, E.Đ., J.L. and S.G.; resources, S.G.; writing—original draft preparation, L.K.; writing—review and editing, S.G., J.L., E.Đ. and I.P.; visualization, L.K., J.L., E.Đ., I.P. and S.G.; supervision, E.Đ. and S.G.; project administration, S.G.; funding acquisition, E.Đ., I.P., J.L. and S.G. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Croatian Science Foundation Installation Research Project UIP-2020-02-7413, “Natural Bioactive Compounds as a Source of Potential Antimicrobial Agents in the Control of Bacterial and Other Fungal Pathogens of Olives”, Anti-Mikrobi-OL (AMO) and Slovenian Research Agency (P4-0431 Next-generation Agriculture).
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Schloss, P.D.; Handelsman, J. Status of the microbial census. Microbiol. Mol. Biol. Rev. 2004, 68, 686–691. [Google Scholar] [CrossRef]
- Muthee Gakuubi, M.; Wagacha, J.M.; Dossaji, S.F.; Wanzala, W. Chemical Composition and Antibacterial Activity of Essential Oils of Tagetes minuta (Asteraceae) against Selected Plant Pathogenic Bacteria. Int. J. Microbiol. 2016, 2016, 7352509. [Google Scholar] [CrossRef]
- Strange, R.N.; Scott, P.R. Plant Disease: A Threat to Global Food Security. Annu. Rev. Phytopathol. 2005, 43, 83–116. [Google Scholar] [CrossRef] [PubMed]
- Martins, P.M.M.; Merfa, M.V.; Takita, M.A.; De Souza, A.A. Persistence in phytopathogenic bacteria: Do we know enough? Front. Microbiol. 2018, 9, 1099. [Google Scholar] [CrossRef] [PubMed]
- Leyns, F.; De Cleene, M.; Swings, J.-G.; Deley, J. The host range of the genus Xanthomonas. Bot. Rev. 1984, 50, 308–356. [Google Scholar] [CrossRef]
- Timilsina, S.; Potnis, N.; Newberry, E.A.; Liyanapathiranage, P.; Iruegas-Bocardo, F.; White, F.F.; Goss, E.M.; Jones, J.B. Xanthomonas diversity, virulence and plant-pathogen interactions. Nat. Rev. 2020, 18, 415–427. [Google Scholar] [CrossRef]
- Catara, V.; Cubero, J.; Pothier, J.F.; Bosis, E.; Bragard, C.; Đermić, E.; Holeva, M.C.; Jacques, M.-A.; Petter, F.; Pruvost, O.; et al. Trends in Molecular Diagnosis and Diversity Studies for Phytosanitary Regulated Xanthomonas. Microorganisms 2021, 9, 862. [Google Scholar] [CrossRef]
- Mansfield, J.; Genin, S.; Magori, S.; Citovsky, V.; Sriariyanum, M.; Ronald, P.; Dow, M.A.X.; Verdier, V.; Beer, S.V.; Machado, M.A.; et al. Top 10 plant pathogenic bacteria in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 614–629. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Coulthurst, S.J.; Pritchard, L.; Hedley, P.E.; Ravensdale, M.; Humphris, S.; Burr, T.; Takle, G.; Brurberg, M.B.; Birch, P.R.; et al. Quorum sensing coordinates brute force and stealth modes of infection in the plant pathogen Pectobacterium atrosepticum. PLoS Pathog. 2008, 4, e1000093. [Google Scholar] [CrossRef]
- Quesada, J.M.; Penyalver, R.; Pérez-Panadés, J.; Salcedo, C.I.; Carbonell, E.A.; López, M.M. Dissemination of Pseudomonas savastanoi pv. savastanoi populations and subsequent appearance of olive knot disease. Plant Pathol. 2010, 59, 262–269. [Google Scholar] [CrossRef]
- Arrebola, E.; Cazorla, F.M.; Perez-García, A.; de Vicente, A. Chemical and metabolic aspects of antimetabolite toxins produced by Pseudomonas syringae pathovars. Toxins 2011, 3, 1089–1110. [Google Scholar] [CrossRef] [PubMed]
- Chouhan, S.; Sharma, K.; Guleria, S. Antimicrobial Activity of Some Essential Oils—Present Status and Future Perspectives. Medicines 2017, 4, 58. [Google Scholar] [CrossRef] [PubMed]
- Mole, B.M.; Baltrus, D.A.; Dangl, J.L.; Grant, S.R. Global virulence regulation networks in phytopathogenic. Trends Microbiol. 2007, 15, 363–371. [Google Scholar] [CrossRef] [PubMed]
- Kraepiel, Y.; Barny, M.A. Gram-negative phytopathogenic bacteria, all hemibiotrophs after all? Mol. Plant Pathol. 2016, 17, 313–316. [Google Scholar] [CrossRef]
- Cerboneschi, M.; Decorosi, F.; Biancalani, C.; Ortenzi, M.V.; Macconi, S.; Giovannetti, L.; Viti, C.; Campanella, B.; Onor, M.; Bramanti, E.; et al. Indole-3-acetic acid in plant-pathogen interactions: A key molecule for in planta bacterial virulence and fitness. Res. Microbiol. 2016, 167, 774–778. [Google Scholar] [CrossRef]
- Lovelace, A.H.; Dorhmi, S.; Hulin, M.T.; Li, Y.; Mansfield, J.W.; Ma, W. Effectors Identification in Plant Pathogens. Phytopathology 2023, 113, 637–650. [Google Scholar] [CrossRef]
- Zheng, X.; Weaver Spivey, N.; Zeng, W.; Liu, P.-P.; Qing Fu, Z.; Klessig, D.F.; Yang He, S.; Dong, X. Coronatine Promotes Pseudomonas syringae Virulence in Plants by Activating a Signaling Cascade that Inhibits Salicylic Acid Accumulation. Cell Host Microbe 2012, 11, 587–596. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Bhardwaj, M.; Phan Tran, L.-S. Jasmonic Acid at the Crossroads of Plant Immunity and Pseudomonas syringae Virulence. Int. J. Mol. Sci. 2020, 21, 7482. [Google Scholar] [CrossRef] [PubMed]
- Buonaurio, R.; Moretti, C.; da Silva, D.P.; Cortese, C.; Ramos, C.; Venturi, V. The olive knot disease as a model to study the role of interspecies bacterial communities in plant disease. Front. Plant Sci. 2015, 6, 434. [Google Scholar] [CrossRef] [PubMed]
- Mina, D.; Pereira, J.A.; Lino-Neto, T.; Baptista, P. Screening the Olive Tree Phyllosphere: Search and Find Potential Antagonists Against Pseudomonas savastanoi pv. savastanoi. Front. Microbiol. 2020, 11, 2051. [Google Scholar] [CrossRef]
- Parsek, M.R.; Greenberg, E.P. Sociomicrobiology: The connections between quorum sensing and biofilms. Trends Microbiol. 2005, 13, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Varympopi, A.; Dimopoulou, A.; Theologidis, I.; Karamanidou, T.; Kaldeli Kerou, A.; Vlachou, A.; Karfaridis, D.; Papafotis, D.; Hatzinikolaou, D.G.; Tsouknidas, A.; et al. Bactericides Based on Copper Nanoparticles Restrain Growth of Important Plant Pathogens. Pathogens 2020, 9, 1024. [Google Scholar] [CrossRef] [PubMed]
- Verhaegen, M.; Bergot, T.; Liebana, E.; Stancanelli, G.; Streissl, F.; Mingeot-Leclercq, M.-P.; Mahillon, J.; Bragard, C. On the use ofantibiotics to control plant pathogenic bacteria:a genetic and genomic perspective. Front. Microbiol. 2023, 14, 1221478. [Google Scholar] [CrossRef] [PubMed]
- Fenibo, E.O.; Ijoma, G.N.; Matambo, T. Bipesticides in Sustainable Agriculture: A Critical Sustainable Development Driver Governed by Green Chemistry Principles. Front. Susutain Food Syst. 2021, 5, 619058. [Google Scholar] [CrossRef]
- Warrior, P. Living system as natural crop-protection agents. Pest. Manag. Sci. 2000, 56, 681–687. [Google Scholar] [CrossRef]
- Kumar, J.; Ramlal, A.; Mallick, D.; Mishra, V. An Overview of Some Biopesticides and Their Importance in Plant Protection for Commercial Acceptance. Plants 2021, 10, 1185. [Google Scholar] [CrossRef] [PubMed]
- Hahlbrock, K.; Scheel, D. Physiology and molecular biology of phenylpropanoid metabolism. Ann. Rev. Plant Physiol. Plant Mol. Biology 1989, 40, 347–369. [Google Scholar] [CrossRef]
- Fleming, H.P.; Walter, W.M., Jr.; Etchells, J.L. Antimicrobial properties of oleuropein and products of its hydrolysis from green olives. Appl. Microbiol. 1973, 26, 777–782. [Google Scholar] [CrossRef] [PubMed]
- Wińska, K.; Mączka, W.; Łyczko, J.; Grabarczyk, M.; Czubaszek, A.; Szumny, A. Essential Oils as Antimicrobial Agents—Myth or Real Alternative? Molecules 2019, 24, 2130. [Google Scholar] [CrossRef]
- Gonçalves, L.A.; Lorenzo, J.M.; Trindade, M.A. Fruit and Agro-Industrial Waste Extracts as Potential Antimicrobials in Meat Products: A Brief Review. Foods 2021, 10, 1469. [Google Scholar] [CrossRef] [PubMed]
- Poveda, J.; Baptista, P. Filamentous fungi as biocontrol agents in olive (Olea europaea L.) diseases: Mycorrhizial and endophytic fungi. Crop Prot. 2021, 146, 105672. [Google Scholar] [CrossRef]
- Ma, J.T.; Dong, X.Y.; Li, Z.H.; Yan, H.; He, J.; Liu, J.K.; Feng, T. Antibacterial Metabolites from Kiwi Endophytic Fungus Fusarium tricinctum, a Potential Biocontrol Strain for Kiwi Canker Disease. J. Agric. Food Chem. 2023, 71, 7679–7688. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Liu, Z.; Duan, F.; Chen, Y.; Qiu, K.; Xiong, Q.; Lin, H.; Zhang, J.; Tan, H. Isolation, identification, and antibacterial evaluation of endophytic fungi from Gannan navel orange. Front. Microb. 2023, 14, 1172629. [Google Scholar] [CrossRef]
- Guzmán-Guzmán, P.; Kumar, A.; de los Santos-Villalobos, S.; Parra-Cota, F.I.; Orozco-Mosqueda, M.d.C.; Fadiji, A.E.; Hyder, S.; Babalola, O.O.; Santoyo, G. Trichoderma Species: Our Best Fungal Allies in the Biocontrol of Plant Diseases—A Review. Plants 2023, 12, 432. [Google Scholar] [CrossRef] [PubMed]
- Song, X.-Y.; Shen, Q.-T.; Xie, S.-T.; Chen, X.-L.; Sun, C.-Y.; Zhong, Y.-Z. Broad-spectrum antimicrobial activity and high stability of Trichokonins from Trichoderma koningii SMF2 against plant pathogens. FEMS Microbiol. Lett. 2006, 260, 119–125. [Google Scholar] [CrossRef]
- Sivasakthi, S.; Usharani, G.; Saranraj, P. Biocontrol potentiality of plant growth promoting bacteria (PGPR)—Pseudomonas fluorescens and Bacillus subtilis: A review. Afr. J. Agric. Res. 2014, 9, 1265–1277. [Google Scholar]
- Chet, I.; Ordentlich, A.; Shapira, R.; Oppenheim, A. Mechanisms of biocontrol of soil-borne plant pathogens by rhizobacteria. Plant Soil. 1990, 129, 85–92. [Google Scholar] [CrossRef]
- van Loon, L.C.; Bakker, P.A.H.M.; Pieterse, C.M.J. Systemic resistance induced by rhizosphere bacteria. Annu. Rev. Phytopathol. 1998, 36, 453–483. [Google Scholar] [CrossRef] [PubMed]
- Barea, J.M.; Pozo, M.J.; Azcón-Aguilar, C. Microbial co-operation in the rhizosphere. J. Exp. Bot. 2005, 56, 1761–1778. [Google Scholar] [CrossRef] [PubMed]
- Compant, S.; Duffy, B.; Nowak, J.; Clément, C.; Barka, E.A. Use of plant growth-promoting bacteria for biocontrol of plant diseases: Principles, mechanisms of action, and future prospects. Appl. Environ. Microbiol. 2005, 71, 4951–5959. [Google Scholar] [CrossRef]
- Chang, W.T.; Chen, Y.C.; Jao, C.L. Antifungal activity and enhancement of plant growth by Bacillus cereus grown on shellfish chitin wastes. Bioresour. Technol. 2007, 98, 1224–1230. [Google Scholar] [CrossRef] [PubMed]
- Kaymak, H.C. Potential of PGPR in Agricultural Inovations. In Plant Growth and Health Promoting Bacteria; Springer: Berlin/Heidelberg, Germany, 2010; pp. 45–79. [Google Scholar]
- Shen, X.; Hu, H.; Peng, H.; Wang, W.; Zhang, X. Comparative genomic analysis of four representative plant growth-promoting rhizobacteria in Pseudomonas. BMC Genom. 2013, 14, 271. [Google Scholar] [CrossRef] [PubMed]
- Bukhat, S.; Imran, A.; Javaid, S.; Shadid, M.; Majeed, A.; Naqqash, T. Communication of plants with microbial world: Exploring the regulatory networks for PGPR mediated defense signaling. Microbiol. Res. 2020, 238, 126486. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Liu, R.; You, M.P.; Barbetti, M.J.; Chen, Y. Pathogen Biocontrol Using Plant Growth-Promoting Bacteria (PGPR): Role of Bacterial Diversity. Microorganisms 2021, 9, 1988. [Google Scholar] [CrossRef] [PubMed]
- Aćimović, S.G.; Zeng, Q.; McGhee, G.; Sundin, G.W.; Wise, J.C. Control of fire blight (Erwinia amylovora) on apple trees with trunk-injected plant resistance inducers and antibiotics and assessment of induction of pathogenesis-related protein genes. Front. Plant Sci. 2015, 6, 16. [Google Scholar] [CrossRef]
- Ali, A.O.; Rashid, T.S. Biocontrol Activities of Olive Endophytic Bacteria Isolates against Pseudomonas savastanoi. Polytech. J. 2022, 12, 114–120. [Google Scholar] [CrossRef]
- Zhang, C.; Lu, S.; Wei, C.; Zhao, J. Bacterium strain 1-1-4 Pseudomonas fluorescens for biocontrol of peach crown gall disease. Acta Agric. Shanghai 1991, 7, 65–69. [Google Scholar]
- Mondal, K.K.; Singh, R.P.; Dureja, P.; Verma, J.P. Secondary metabolites of cotton rhizobacteria in the suppression of bacterial blight of cotton. Indian. Phytopathol. 2000, 53, 22–27. [Google Scholar]
- Stockwell, V.O.; Johnson, K.B.; Sugar, D.; Loper, J. Control of fire blight by Pseudomonas fluorescens A506 and Pantoea vagans C9-1 applied as single strains and mixed inocula. Phytopathology 2010, 100, 1330–1339. [Google Scholar] [CrossRef]
- Elsayed, T.R.; Grosch, R.; Smalla, K. Potato plant spheres and to a lesser extent the soil type influence the proportion and diversity of bacterial isolates with in vitro antagonistic activity towards Ralstonia solanacearum. FEMS Microbioly Ecol. 2021, 97, fiab038. [Google Scholar] [CrossRef] [PubMed]
- Bouaichi, A.; Benkirane, R.; El-kinany, S.; Habbadi, K.; Lougraimzi, H.; Sadik, S.; Benbouazza, A.; Achbani, E.H. Potential effect of antagonistic bacteria in the management of olive knot disease caused by Pseudomonas savastanoi pv. savastanoi. J. Microbiol. Biotech. Food Sci. 2019, 4, 1035–1040. [Google Scholar] [CrossRef]
- Krid, S.; Triki, M.A.; Gargouri, A.; Rhouma, A. Biocontrol of olive knot disease by Bacillus subtilis isolated from olive leaves. Ann. Microbiol. 2012, 62, 149–154. [Google Scholar] [CrossRef]
- Doksöz, S.; Bozkurt, I.A. Biological control of Pseudomonas savastanoi pv. savastanoi causing the olive knot disease with epiphytic and endophytic bacteria. J. Plant Pathol. 2022, 104, 65–78. [Google Scholar] [CrossRef]
- Broniarek-Niemiec, A.; Børve, J.; Puławska, J. Control of Bacterial Canker in Stone Fruit Trees by Chemical and Biological Products. Agronomy 2023, 13, 1166. [Google Scholar] [CrossRef]
- Vero, S.; Garmendia, G.; Allori, E.; Sanz, J.M.; Gonda, M.; Alconada, T.; Cavello, I.; Dib, J.R.; Diaz, M.A.; Nally, C.; et al. Microbial Biopesticides: Diversity, Scope, and Mechanisms Involved in Plant Disease Control. Diversity 2023, 15, 457. [Google Scholar] [CrossRef]
- Buttimer, C.; McAuliffe, O.; Ross, R.P.; Hill, C.; O’Mahony, J.; Coffey, A. Bacteriophages and Bacterial Plant Diseases. Front. Microbiol. 2017, 8, 34. [Google Scholar] [CrossRef] [PubMed]
- Bragard, C.; Dehnen-Schmutz, K.; Di Seiro, F.; Gonthier, P.; Jacques, M.-A.; Jacques Miret, J.A.; Fejer Justesen, A.; MacLeod, A.; Magnusson, C.S.; Guzzo, M.; et al. Effectivness of in planta control measures of Xylella fastidiosa. EFSA J. 2019, 17, 5666. [Google Scholar] [CrossRef]
- Carlsson, G.; Palmborg, C.; Huss-Danell, K. Discrimination against 15N in three N2-fixing Trifolium species as influenced by Rhizobium strain and plant age. Acta Agric. Scanda 2006, 56, 31–38. [Google Scholar] [CrossRef]
- Ahmed, A.; Hasnain, S. Auxin producing Bacillus sp.: Auxin quantification and effect on the growth Solanum tuberosum. Pure Appl. Chem. 2010, 82, 313–319. [Google Scholar] [CrossRef]
- Caballo-Ponce, E.; Meng, X.; Uzelac, G.; Halliday, N.; Cámara, M.; Licastro, D.; da Silva, D.P.; Ramos, C.; Venturi, V. Quorum Sensing in Pseudomonas savastanoi pv. savastanoi and Erwinia toletana: Role in Virulence and Interspecies Interactions in the Olive Knot. Appl. Environ. Microbiol. 2018, 84, e00950-18. [Google Scholar] [CrossRef] [PubMed]
- Vuletin Selak, G.; Raboteg Božiković, M.; Abrouk, D.; Bolčić, M.; Žanić, K.; Perica, S.; Normand, P.; Pujić, P. Pseudomonas ST1 and Pantoea Paga Strains Cohabit in Olive Knots. Microorganisms 2022, 10, 1528. [Google Scholar] [CrossRef] [PubMed]
- Pandit, M.A.; Kumar, J.; Gulati, S.; Bhandari, N.; Mehta, P.; Katyal, R.; Rawat, C.D.; Mishra, V.; Kaur, J. Major Biological Control Strategies for Plant Pathogens. Pathogens 2022, 11, 273. [Google Scholar] [CrossRef] [PubMed]
- Koul, B.; Yakoob, M.; Shah, M.P. Agricultural waste management strategies for environmental sustainability. Environ. Res. 2022, 206, 112285. [Google Scholar] [CrossRef] [PubMed]
- Nath, P.C.; Ojha, A.; Debnath, S.; Sharma, M.; Sridhar, K.; Nayak, P.K.; Inbaraj, B.S. Biogeneration of Valuable Nanomaterials from Agro-Wastes: A Comprehensive Review. Agronomy 2023, 13, 561. [Google Scholar] [CrossRef]
- Bouaziz, M.; Grayer, R.J.; Simmonds, M.S.J.; Damak, M.; Sayadi, S. Identification and antioxidant potential of flavonoids and low molecular weight phenols in olive cultivar Chemlali growing in Tunisia. J. Agric. Food Chem. 2005, 53, 236–241. [Google Scholar] [CrossRef]
- Obied, H.K.; Allen, M.S.; Bedgood, D.R.; Prenzler, P.D.; Robards, K. Bioactivity and analysis of biophenols recovered from olive mill waste. J. Agricult Food Chem. 2005, 53, 823–837. [Google Scholar] [CrossRef]
- Yakhlef, W.; Arhab, R.; Romero, C.; Brenes, M.; de Castro, A.; Medina, E. Phenolic composition and antimicrobial activity of Algerian olive products and by-products. LWT Food Sci. Technol. 2018, 93, 323–328. [Google Scholar] [CrossRef]
- Feki, M.; Allouche, N.; Bouaziz, M.; Gargoubi, A.; Sayadi, S. Effect of storage of olive mill wastewaters on hydroxytyrosol concentration. Eur. J. Lipid Sci. Tech. 2006, 108, 1021–1027. [Google Scholar] [CrossRef]
- Obied, H.K.; Bedgood, D.R., Jr.; Prenzler, P.D.; Robards, K. Bioscreening of Australian olive mill waste extracts: Biophenol content, antioxidant, antimicrobial and molluscicidal activities. Food Chem. Toxicol. 2007, 45, 1238–1248. [Google Scholar] [CrossRef]
- Krid, S.; Bouaziz, M.; Triki, M.A.; Gargouri, A.; Rhouma, A. Inhibition of olive knot disease by polyphenols extracted from olive mill waste waters. J. Plant Pathol. 2011, 93, 561–568. [Google Scholar]
- Capasso, R.; Evidente, A.; Schivo, L.; Orru’, G.; Marcialis, M.A.; Cristinzio, G. Antibacterial polyphenols from olive oil mill waste waters. J. Appl. Bacteriol. 1995, 79, 393–398. [Google Scholar] [CrossRef] [PubMed]
- Ciafardini, G.; Zullo, B.A. Antimicrobial activity od oil-mill waste water polyphenols on the phytopathogen Xanthomonas campestris spp. Ann. Microbiol. 2003, 53, 283–290. [Google Scholar]
- Caballero-Guerrero, B.; Garrido-Fernández, A.; Fermoso, F.G.; Rodríguez-Gutierrez, G.; Fernández-Prior, M.A.; Reinhard, C.; Nyström, L.; Benítez-Cabello, A.; Nóe Arroyo-López, F. Antimicrobial effects of treated olive mill waste on foodborne pathogens. LWT Food Sci. Tech. 2022, 164, 113628. [Google Scholar] [CrossRef]
- Russo, E.; Spallarossa, A.; Comite, A.; Pagliero, M.; Guida, P.; Belotti, V.; Caviglia, D.; Schito, A.M. Valorization and Potential Antimicrobial Use of Olive Mill Wastewater (OMW) from Italian Olive oil Production. Antioxidants 2022, 11, 903. [Google Scholar] [CrossRef] [PubMed]
- Medina, E.; Romero, C.; de los Santos, B.; de Castro, A.; García, A.; Romero, F.; Brenes, M. Antimicrobial Activity of Olive Solutions from Stored Alpeorujo against Plant Pathogenic Microorganisms. J. Agric. Food Chem. 2011, 59, 6927–6932. [Google Scholar] [CrossRef] [PubMed]
- Pannucci, E.; Caracciolo, R.; Romani, A.; Cacciola, F.; Dugo, P.; Bernini, R.; Varvaro, L.; Santi, L. An hydroxytyrosol enriched extract from olive mill wastewaters exerts antioxidant activity and antimicrobial activity on Pseudomonas savastanoi pv. savastanoi and Agrobacterium tumefaciens. Nat. Product. Res. 2019, 35, 2677–2684. [Google Scholar] [CrossRef]
- Ditsawanon, T.; Roytrakul, S.; Phaonakrop, N.; Charoenlappanit, S.; Thaisakun, S.; Parinthawong, N. Novel Small Antimicrobial Peptides Extracted from Agricultural Wastes Act against Phytopathogens but Not Rhizobacteria. Agronomy 2022, 12, 1841. [Google Scholar] [CrossRef]
- Chiocchio, I.; Mandrone, M.; Tacchini, M.; Guerrini, A.; Poli, F. Phytochemical Profile and In Vitro Bioactivities of Plant-Based By-Products in View of a Potential Reuse and Valorization. Plants 2023, 12, 795. [Google Scholar] [CrossRef] [PubMed]
- Saleem, M.; Saeed, M.T. Potential application of waste fruit peels (orange, yellow lemon and banana) as wide range natural antimicrobial agent. J. King Saud. Univ. Sci. 2020, 32, 805–810. [Google Scholar] [CrossRef]
- Terea, H.; Selloum, D.; Rebiai, A.; Bouafia, A.; Mya, O.B. Preparation and characterization of cellulose/ZnO nanoparticles extracted from peanut shells: Effects on antibacterial and antifungal activities. Biomass Convers. Biorefin. 2023, 1–12. [Google Scholar] [CrossRef]
- Ul-Islam, M.; Alhajaim, W.; Fatima, A.; Yasir, S.; Kamal, T.; Abbas, Y.; Khan, S.; Khan, A.H.; Manan, S.; Ullah, M.W.; et al. Development of low-cost bacterial cellulose-pomegranate peel extract-based antibacterial composite for potential biomedical applications. Int. Jo Biol. Macromol. 2023, 231, 123269. [Google Scholar] [CrossRef] [PubMed]
- Hyun Lee, K.; Chun, Y.; Hyun Lee, J.; Uk Lee, J.; Lee, T.; Young Yoo, H. Sustainable Utilization Strategy of Organic Waste via Fabrication of Bioelastomer with Antibacterial and Antioxidant Activities Using Mandarin Peel Extracts. Agriculture 2023, 13, 161. [Google Scholar] [CrossRef]
- D’agostino, M.; Tesse, N.; Frippiat, J.P.; Machouart, M.; Debourgogne, A. Essential Oils and Their Natural Active Compounds Presenting Antifungal Properties. Molecules 2019, 24, 3713. [Google Scholar] [CrossRef] [PubMed]
- de Almeida, J.M.; Crippa, B.L.; Martins Alencar de Souza, V.V.; Perez Alonso, V.P.; da Motta Santos Júnior, E.; Picone, C.F.S.; Prata, A.S.; Silva, N.C.C. Antimicrobial action of Oregano, Thyme, Clove, Cinnamon and Black pepper essential oils free and encapsulated against foodborne pathogens. Food Control 2023, 144, 109356. [Google Scholar] [CrossRef]
- Najmi, Z.; Calogero Scalia, A.; De Giglio, E.; Cometa, S.; Cochis, A.; Colasanto, A.; Locatelli, M.; Coisson, J.D.; Iriti, M.; Vallone, L.; et al. Screening of Different Essential Oils Based on Their Physicochemical and Microbiological Properties to Preserve Red Fruits and Improve Their Shelf Life. Foods 2023, 12, 332. [Google Scholar] [CrossRef]
- Benali, T.; Bouyahya, A.; Habbadi, K.; Zengin, G.; Khabbach, A.; Achbani, E.H.; Hammani, K. Chemical composition and antibacterial activity of the essential oil and extracts of Cistus ladaniferus subsp. ladanifer and Mentha suaveolens against phytopathogenic bacteria and their ecofriendly management of phytopathogenic bacteria. Biocat Agric. Biotech. 2020, 28, 101696. [Google Scholar] [CrossRef]
- Rossi, P.-G.; Berti, L.; Panighi, J.; Luciani, A.; Maury, J.; Muselli, A.; Serra, D.R.; Gonny, M.; Bolla, J.M. Antibacterial action of essential oils from Corsica. J. Essent. Oil Res. 2007, 19, 176–182. [Google Scholar] [CrossRef]
- Amensour, M.; Sendra, E.; Perez-Alvarez, J.A.; Skali-Senhaji, N.; Abrini, J.; Fernández-López, J. Antioxidant activity and chemical content of methanol and ethanol extracts from leaves of rockrose (Cistus ladaniferus). Plant Foods Hum. Nutr. 2010, 65, 170–178. [Google Scholar] [CrossRef]
- de Sousa Barros, A.; de Morais, S.M.; Ferreira, P.A.T.; Vieira, I.G.P.; Craveiro, A.A.; Fontenelle, R.O.S.; de Menezes, J.E.S.A.; da Silva, F.W.F.; de Sousa, H.A. Chemical composition and functional properties of essential oils from Mentha species. Ind. Crop Prod. 2015, 76, 557–564. [Google Scholar] [CrossRef]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological effects of essential oils—A review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef] [PubMed]
- Shaaban, H.A.H.; El-Ghorab, A.H.; Takayuki, S. Bioactivity of essential oils and their volatile aroma components: Review. J. Essent. Oil Res. 2012, 24, 203–2012. [Google Scholar] [CrossRef]
- Caparrotta, S.; Comparini, D.; Marone, E.; Kimmenfield, R.; Luzzietti, L.; Taiti, C.; Mancuso, S. Correlation between VOC fingerprinting and antimicrobial activity of several essential oils extracted by plant resins against A. tumefaciens and P. savastanoi. Flavour. Fragr. J. 2019, 5, 377–387. [Google Scholar] [CrossRef]
- Sauer, A.V.; Santos, E.M.; Gonçalves-Zuliani, A.M.O.; Nocchi, P.T.R.; Nunes, W.M.C.; Bonato, C.M. Bacteriostatic and Bactericidal Activity In Vitro of Different Essential Oils as Alternative Treatments to Control Xanthomonas citri subsp. citri. Acta Hortic. 2015, 1065, 931–936. [Google Scholar] [CrossRef]
- Rathod, T.; Padalia, H.; Chand, S. Chemical constituents of Mentha piperita and Pongamia pinnata essential oils and their synergistic anticandidal activity with some antibiotics against multidrug resistant clinical isolates of Candida. J. Pharm. Phytochem. 2017, 6, 579–589. [Google Scholar]
- Vasinauskiene, M.; Radušiene, J.; Zitikaite, I.; Surviliene, E. Antibacterial activities of essential oils from aromatic and medicinal plants against growth of phytopathogenic bacteria. Agron. Res. 2006, 4, 437–440. [Google Scholar]
- Brentini Santiago, M.; da Silva Moraes, T.; Massuco, J.E.; Silva, L.O.; Lucarini, R.; da Silva, D.F.; Manzini Vieira, T.; Miller Crotti, A.E.; Gomes Martins, C.H. In vitro evaluation of essential oils for potential antibacterial effects against Xyllela fastidiosa. J. Phytopathol. 2018, 166, 790–798. [Google Scholar] [CrossRef]
- Bozkurt, I.A.; Soylu, S.; Kara, M.; Soylu, E.M. Chemical Composition and Antibacterial Activity of Essential Oils Isolated from Medicinal Plants against Gall Forming Plant Pathogenic Bacterial Disease Agents. KSU J. Agric. Nat. 2020, 23, 1474–1482. [Google Scholar] [CrossRef]
- Grul’ová, D.; Caputo, L.; Elshafie, H.S.; Baranová, B.; De Martino, L.; Sedlák, V.; Gogal’ová, Z.; Poráčová, B.; Camele, I.; De Feo, V. Thymol Chemotype Origanum vulgare L. Essential Oil as a Potential Selective Bio-Based Herbicide on Monocot Plant Species. Molecules 2020, 25, 595. [Google Scholar] [CrossRef] [PubMed]
- Tarakanov, R.I.; Dzhalilov, F.S.U. Using of Essential Oils and Plant Extracts against Pseudomonas savastanoi pv. glycinea and Curtobacterium flaccumfaciens pv. flaccumfaciens on Soybean. Plants 2022, 11, 2989. [Google Scholar] [CrossRef] [PubMed]
- György, É.; Laslo, É.; Salamon, B. Antimicrobial impacts of selected Lamiaceae plants on bacteria isolated from vegetables and their application in edible films. Food Biosci. 2023, 51, 102280. [Google Scholar] [CrossRef]
- Patel, H.K.; Gomes, E.N.; Wu, Q.; Patel, N.; Kobayashi, D.Y.; Wang, C.; Simon, J.E. Volatile metabolites from new cultivars of catnip and oregano as potential antibacterial and insect repellent agents. Front. Plant Sci. 2023, 14, 1124305. [Google Scholar] [CrossRef] [PubMed]
- Todorović, B.; Potočnik, I.; Rekanović, E.; Stepanović, M.; Kostić, M.; Ristić, M.; Milijašević-Marčić, S. Toxicity of twenty-two plant essential oils against pathogenic bacteria of vegetables and mashrooms. J. Environ. Sci. Health B 2016, 51, 832–839. [Google Scholar] [CrossRef]
- Gormez, A.; Bozari, S.; Yanmis, D.; Gulluce, M.; Sahin, F.; Agar, G. Chemical Composition and Antibacterial Activity of Essential Oils of Two Species of Lamiaceae against Phytopathogenic bacteria. Pol. J. Microbiol. 2015, 64, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Nazzaro, F.; Fratianni, F.; De Martino, L.; Coppola, R.; De Feo, V. Effect of Essential Oils on Pathogenic Bacteria. Pharmaceuticals 2013, 6, 1451–1474. [Google Scholar] [CrossRef]
- Tardugno, R.; Serio, A.; Pellati, F.; D’Amato, S.; Chaves López, C.; Bellardi, M.G.; Di Vito, M.; Savini, V.; Paparella, A.; Benvenuti, S. Lavandula × intermedia and Lavandula angustifolia essential oils: Phytochemical composition and antimicrobial activity against foodborne pathogens. Nat. Prod. Res. 2019, 33, 3330–3335. [Google Scholar] [CrossRef]
- Verdeguer, M.; Sánchez-Moreiras, A.M.; Araniti, F. Phytotoxic Effects and Mechanism of Action of Essential Oils and Terpenoids. Plants 2020, 9, 1571. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, R.K.; Dwivedi, P.; Ahmad, S. Screening of Antibacterial Activity of Six Plant Essential Oils against Pathogenic Bacterial Strains. Asian J. MedSci. 2010, 2, 152–158. [Google Scholar]
- Sánchez-Hernández, E.; González-García, V.; Palacio-Bielsa, A.; Lorenzo-Vidal, B.; Buzón-Durán, L.; Martín-Gil, J.; Martín-Ramos, P. Antibacterial Activity of Ginko biloba Extracts against Clavibacter michiganensis subsp. michiganensis, Pseudomonas spp., and Xanthomonas vesicatoria. Horticulturae 2023, 9, 461. [Google Scholar] [CrossRef]
- Mamoucha, S.; Prombona, A.; Galeou, A. Evaluation of the antibacterial activity of essential oil of Laurus nobilis against Pseudomonas syringae pv. phaseolicola and potential biocidal action. Hell. Plant Prot. J. 2023, 16, 29–39. [Google Scholar] [CrossRef]
- Janaćković, P.; Rajčević, N.; Gavrilović, M.; Novaković, J.; Radulović, M.; Miletić, M.; Janakiev, T.; Dimkić, I.; Marin, P.D. Essential Oil Composition of Ambrosia artemisiifolia and Its Antibacterial Activity against Phytopathogens. Biol. Life Sci. Forum 2022, 15, 22. [Google Scholar] [CrossRef]
- Bouchekouk, C.; Zohra Kara, F.; Tail, G.; Saidi, F.; Benabdelkader, T. Essential oil composition and antibacterial activity of Pteridium aquilinum (L.) Kuhn. Biol. Futur. 2019, 70, 56–61. [Google Scholar] [CrossRef]
- Dorman, H.J.D.; Deans, S.G. Antimicrobial agents from plants: Antibacterial activity of plant volatile oils. J. Appl. Microbiol. 2000, 88, 308–316. [Google Scholar] [CrossRef] [PubMed]
- Hyldgaard, M.; Mygind, M.; Meyer, R.L. Essential oils in food preservation: Mode of action, synergies and interactions with food matrix components. Front. Microbiol. 2012, 3, 12. [Google Scholar] [CrossRef]
- Badawy, M.E.I.; Abdelgaleil, S.A.M. Composition and antimicrobial activity of essential oils isolated from Egyptian plants against plant pathogenic bacteria and fungi. Ind. Crops Prod. 2014, 52, 776–782. [Google Scholar] [CrossRef]
- Feizi, H.; Tahan, V.; Kariman, K. In vitro antibacterial activity of essential oils from Carum copticum and Ziziphora clinopodioides plants against the phytopathogen Pseudomonas syringae pv. syringae. Plant Biosyst. 2023, 157, 487–492. [Google Scholar] [CrossRef]
- Shukal, S.; Chen, X.; Zhang, C. Systematic engineering for high-yield production of viridiflorol and amorphadiene in auxotrophic Escherichia coli. Metab. Eng. 2019, 55, 170–178. [Google Scholar] [CrossRef]
- Fontana, R.; Macchi, G.; Caproni, A.; Sicurella, M.; Buratto, M.; Salvatori, F.; Pappadà, M.; Manfredini, S.; Baldisserotto, A.; Marconi, P. Control of Erwinia amylovora Growth by Moringa oleifera Leaf Extracts: In Vitro and in Planta Effects. Plants 2022, 11, 957. [Google Scholar] [CrossRef]
- Al-Baharwee, N.I.K.; Qasim, W.S.; Abdlla, Y.A. Phytochemical analysis of Annona atemoya seed extract by HPLC and their ability to inhibit the growth of Agrobacterium tumefaciens. Biodiversitas 2023, 24, 603–608. [Google Scholar] [CrossRef]
- Morales-Ubaldo, A.L.; Rivero-Perez, N.; Avila-Ramos, F.; Aquino-Torres, E.; Prieto-Méndez, J.; Hetta, H.F.; El-Saber Batiha, G.; Zaragoza-Bastida, A. Bactericidal Activity of Larrea tridentata Hydroalcoholic Extract against Phytopathogenic bacteria. Agronomy 2021, 11, 957. [Google Scholar] [CrossRef]
- Diao, W.R.; Hua, Q.P.; Zhang, H.; Xu, J.G. Chemical composition, antibacterial activity and mechanism of action of essential oil from seeds of fennel (Foeniculum vulgare Mill.). Food Control 2014, 35, 109–116. [Google Scholar] [CrossRef]
- Barut, M.; Tansi, L.S.; Akyuz, A.M.; Karaman, S. Quality and yield of different basil (Ocimum basilicum L.) cultivars with various planting and cutting times under hot Mediterranean climate. Appl. Ecol. Environ. Res. 2021, 19, 3115–3136. [Google Scholar] [CrossRef]
- Hsouna, A.B.; Touj, N.; Hammami, I.; Dridi, K.; Al-Ayed, A.S.; Hamdi, N. Chemical Composition and in vivo Efficacy of the Essential Oil of Mentha piperita L. in the Suppression of Crown Gall Disease on Tomato Plants. J. Oleo Sci. 2019, 68, 419–426. [Google Scholar] [CrossRef]
- Vinceković, M.; Maslov Bandić, L.; Jurić, S.; Jalšenjak, N.; Čaić, A.; Živičnjak, E.; Đermić, E.; Karoglan, M.; Osrečak, M.; Topolovec-Pintarić, S. The enhancement of bioactive potential in Vitis vinifera leaves by application of microspheres loaded with biological and chemical agents. J. Plant Nutr. 2019, 42, 543–558. [Google Scholar] [CrossRef]
- Jurić, S.; Sopko Stracenski, K.; Król-Kilińska, Ż.; Žutić, I.; Uher, S.F.; Đermić, E.; Vinceković, M. The enhancement of plant secondary metabolites content in Lactuca sativa L. by encapsulated bioactive agents. Sci. Rep. 2020, 10, 3737. [Google Scholar] [CrossRef] [PubMed]
- Melchionna Albuquerque, P.; Gomes Azevedo, S.; Pereira de Andrade, C.; Corrêa de Souza D’Ambros, N.; Martins Pérez, M.T.; Manzato, L. Biotechnological Applications of Nanoencapsulated Essential Oils: A Review. Polymers 2022, 14, 5495. [Google Scholar] [CrossRef]
Figure 1.
Graphical presentation of increasing trend of scientific research focusing on the use of essential oils and their ingredients against various bacterial species based on data from Web of Science Core Collection using the keywords “essential oil” and “plant bacteria”. Available online: https://www.webofscience.com/wos/woscc/summary/e9e2769f-ace1-4e30-9da4-9daffa8c0efa-9f82a284/relevance/1 (accessed on 24 August 2023).
Figure 1.
Graphical presentation of increasing trend of scientific research focusing on the use of essential oils and their ingredients against various bacterial species based on data from Web of Science Core Collection using the keywords “essential oil” and “plant bacteria”. Available online: https://www.webofscience.com/wos/woscc/summary/e9e2769f-ace1-4e30-9da4-9daffa8c0efa-9f82a284/relevance/1 (accessed on 24 August 2023).

Table 1.
List of plant wastes tested for the antibacterial potential expressed as minimal inhibitory concentration (MIC) values against various plant pathogenic bacterial species used in different studies.
Table 1.
List of plant wastes tested for the antibacterial potential expressed as minimal inhibitory concentration (MIC) values against various plant pathogenic bacterial species used in different studies.
| Reference | Source | Bacterial Species | Plant Waste/By-Products | MIC | Determination Method | Mode of Action |
|---|---|---|---|---|---|---|
| [73] | Ciafardini and Zullo | Xanthomonas campestris | OMWW phenols–cv. Leccino | 2500 µg/mL | Agar diffusion | protein cross-linking and protein-denaturation |
| [72] | Capasso et al. | Pseudomonas. syringae pv. savastanoi Clavibacter michiganensis | OMWW polyphenols and related compounds | 5 × 10−4 mol/L | Broth dilution | damage to cell membrane |
| [76] | Medina et al. | Erwinia uredovora E. toletana E. amylovora P. savastanoi P. syringae C. michiganensis | Olive wastewater of stored semisolid olive residue | 10–20% (v/v) | Broth dilution | n.t. |
| [77] | Pannucci et al. | P. savastanoi pv. savastanoi Rhizobium radiobacter | Hydroxytyrosol–HTyr Hydroxytyrosol enriched extract of OMWW-HTE | 100–300 300–500 µg/mL | Disc diffusion Subculture method | n.t. |
| [79] | Chicchio et al. | P. syringae pv. syringae C. michiganensis subsp. nebraskense | Methanolic extracts of wastes/by-products: Abutilon theophrasti Medik. Achillea millefolium L. Allium cepa L. Artemisia absinthium L. Beta vulgaris L. Camelina sativa (L.) Crantz Castnea sativa Mill. Cicer arietinum L. Cichorium intybus L. Cucurbita pepo L. Cupressus sempervirens L. Echinochloa crus-galli (L.) P. Beauv. Erigeron canadensis L. Helianthis annus L. Laurus nobilis L. Lavandula angustifolia Mill. Melissa officinalis L. Origanum vulgare L. Phaseolus vulgaris L. Prunus amygdalus Batsch Rosa damascena Solanum lycopersicum L. Solanum tuberosum L. Sorghum bicolor (L.) Moench Thymus vulgaris L. Triticum aestivum L. Vitis vinifera L. | >1000 µg/mL | Broth microdilution | n.t. |
| Helichrysum italicum (Roth) G. Don | 125–>1000 µg/mL | n.t. | ||||
| Salvia officinalis L. Salvia rosmarinus Schleid. | 500–>1000 µg/mL | n.t | ||||
| Salvia sclarea L. | 1000–>1000 µg/mL | n.t. | ||||
| [78] | Ditsawanon et al. | X. oryzae pv. oryzae X. citri Pectobacterium carotovorum R. rhizogenes | Agricultural waste (rice, corn, sugarcane, bagasse) Agro-industrial waste (soybean, peanut, coconut, coffee, fish) | § | Broth dilution | Disruption of cell wall membrane and intercellular biological processes |
§ protein hydrolites and small peptides; n.t.—not tested.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Košćak, L.; Lamovšek, J.; Đermić, E.; Prgomet, I.; Godena, S. Microbial and Plant-Based Compounds as Alternatives for the Control of Phytopathogenic Bacteria. Horticulturae 2023, 9, 1124. https://doi.org/10.3390/horticulturae9101124
AMA Style
Košćak L, Lamovšek J, Đermić E, Prgomet I, Godena S. Microbial and Plant-Based Compounds as Alternatives for the Control of Phytopathogenic Bacteria. Horticulturae. 2023; 9(10):1124. https://doi.org/10.3390/horticulturae9101124
Chicago/Turabian StyleKošćak, Laura, Janja Lamovšek, Edyta Đermić, Iva Prgomet, and Sara Godena. 2023. "Microbial and Plant-Based Compounds as Alternatives for the Control of Phytopathogenic Bacteria" Horticulturae 9, no. 10: 1124. https://doi.org/10.3390/horticulturae9101124
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.