
CsSE59 Encoding Invertase/Pectin Methyl Esterase Inhibitor Is a Candidate Gene Conferring the Virescent True Leaf Phenotype in Cucumber
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials, Growth Conditions, and Phenotype Identification
2.2. Genetic Analysis
2.3. Photosynthetic Parameter Determination
2.4. Measurement of Chlorophyll Fluorescence Kinetic Parameters
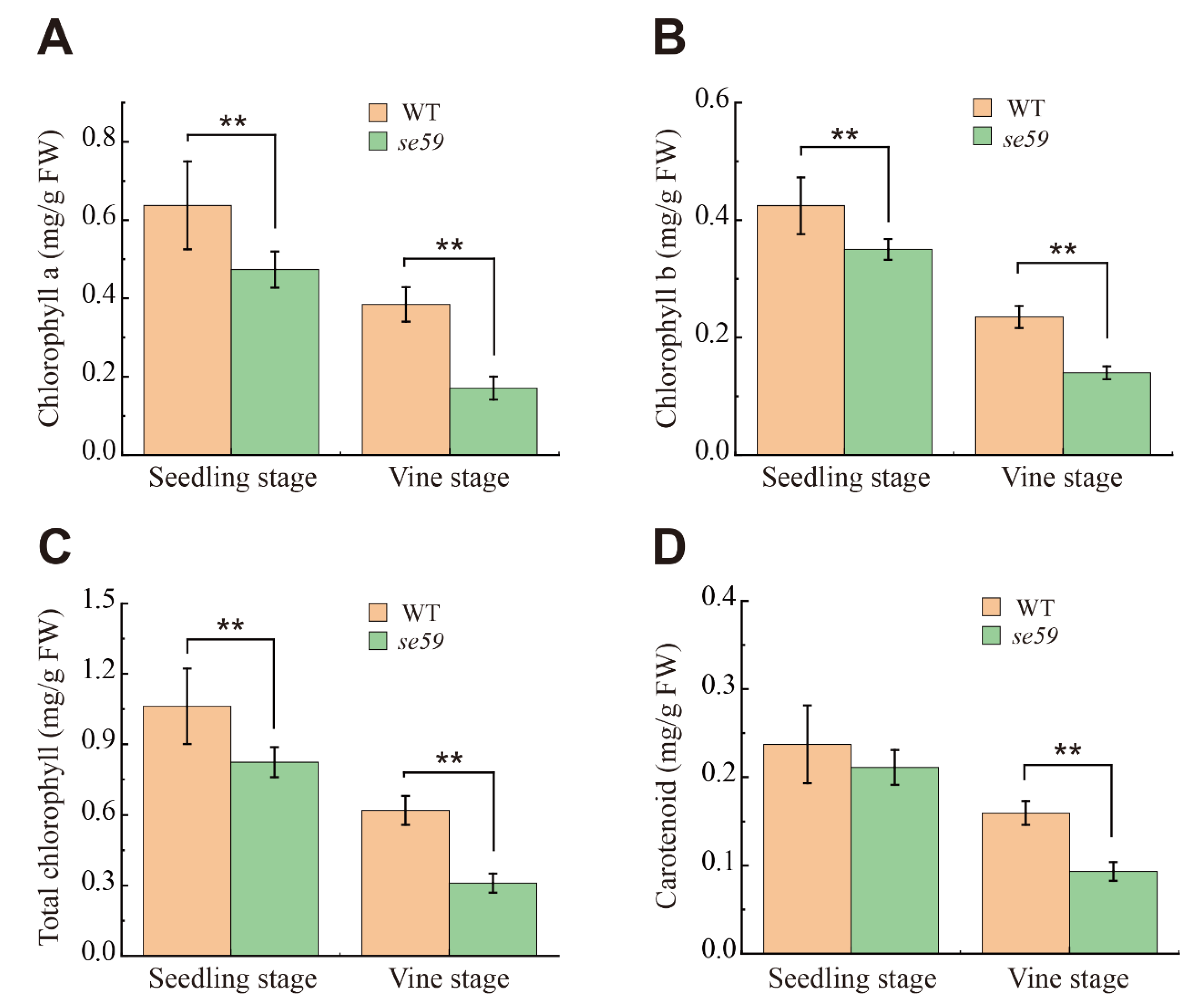
2.5. Measurement of Pigment Contents
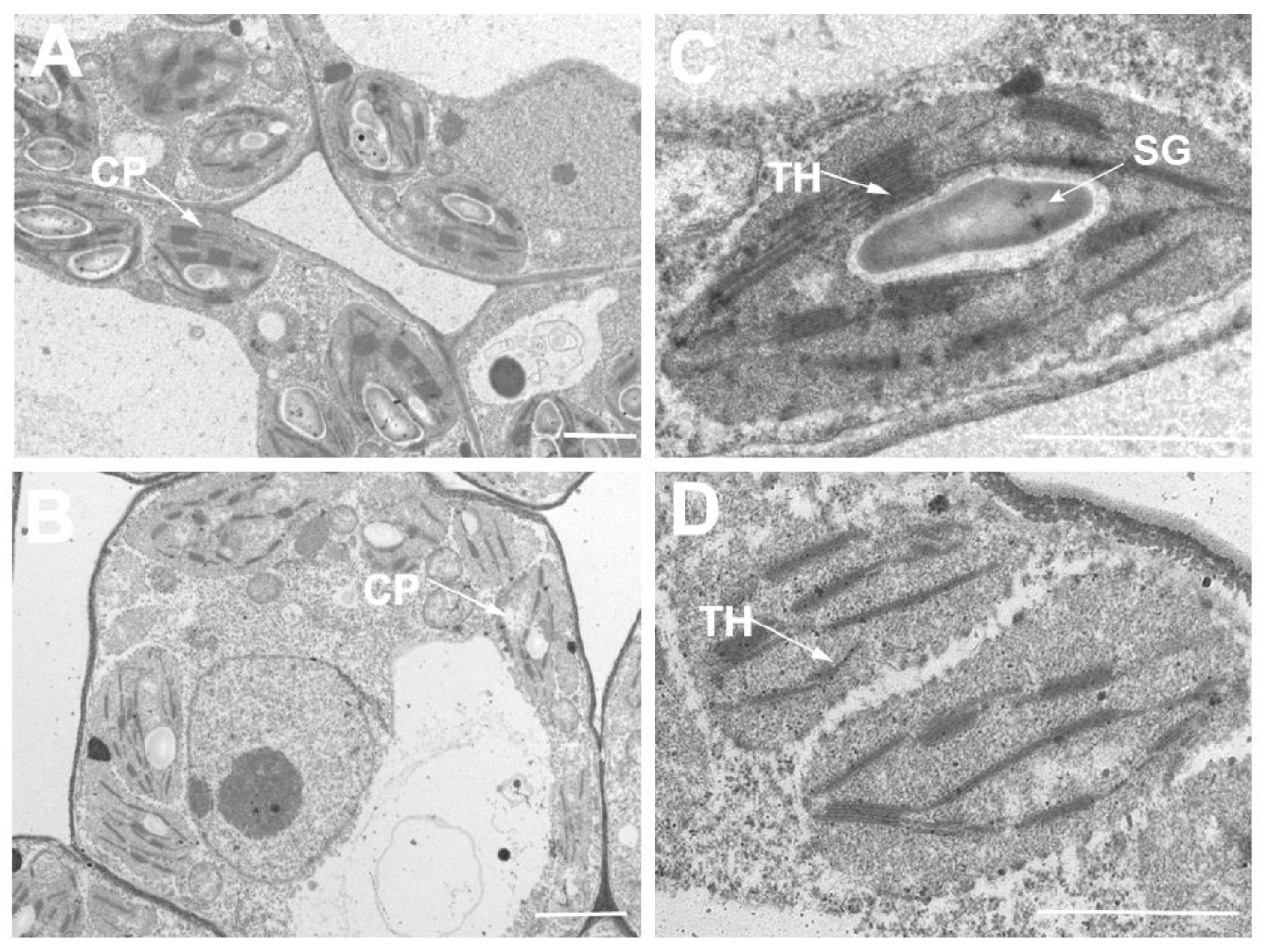
2.6. Transmission Electron Microscopy Observation
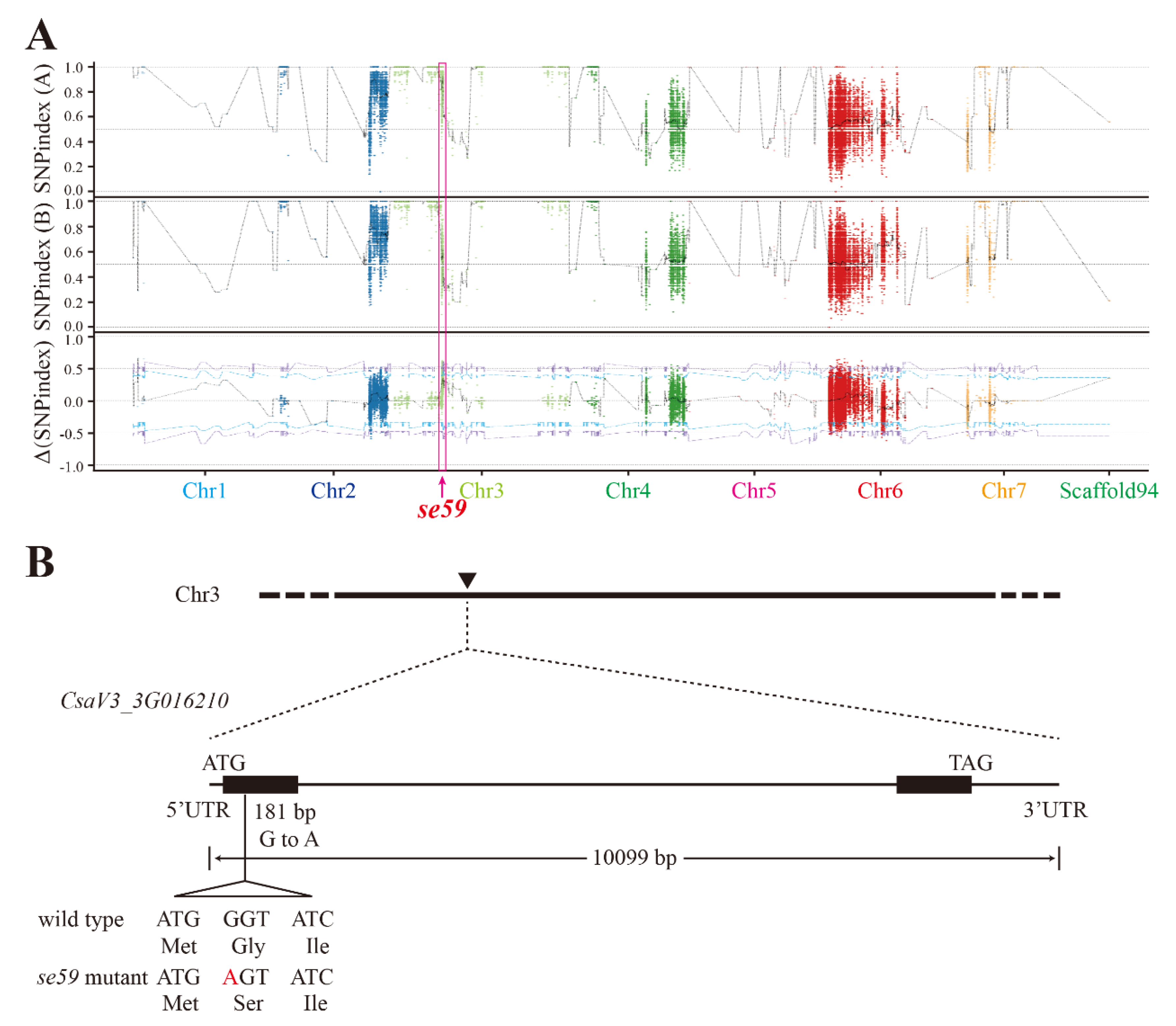
2.7. BSA-Seq Analysis of se59 Locus
2.8. RNA-Seq Analysis
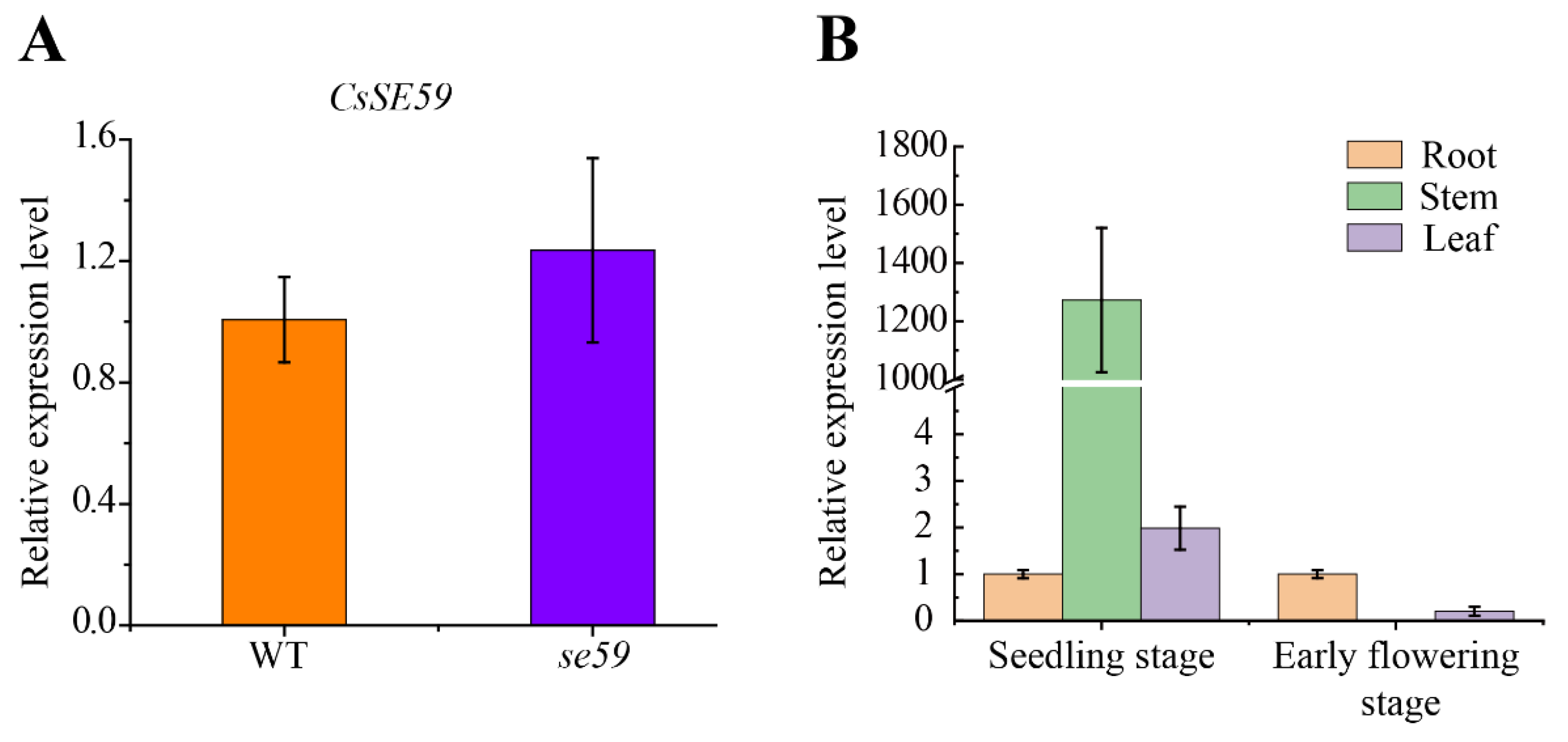
2.9. RNA Extraction and qRT-PCR
2.10. Statistical Analysis
3. Results
3.1. The se59 Produced a Virescent Leaf Phenotype
3.2. Genetic Analysis of se59 Locus
3.3. The se59 Has Defective Chloroplast Development, Chlorophyll Synthesis and Photosynthetic Capacity at Early Leaf Stage
3.4. Identification of Candidate Genes for se59 Mutant
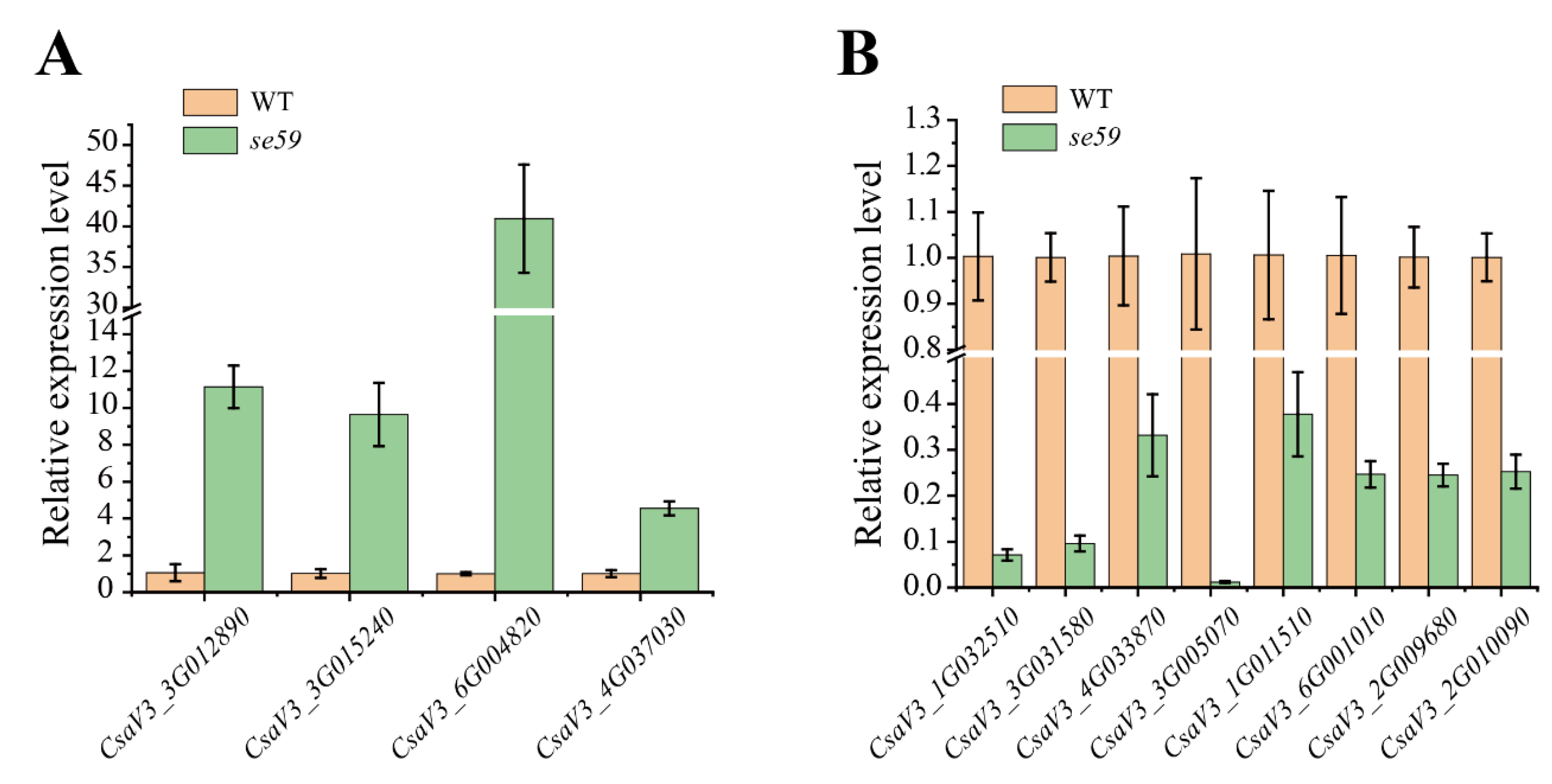
3.5. Sequence and Expression Analysis of CsSE59 Gene
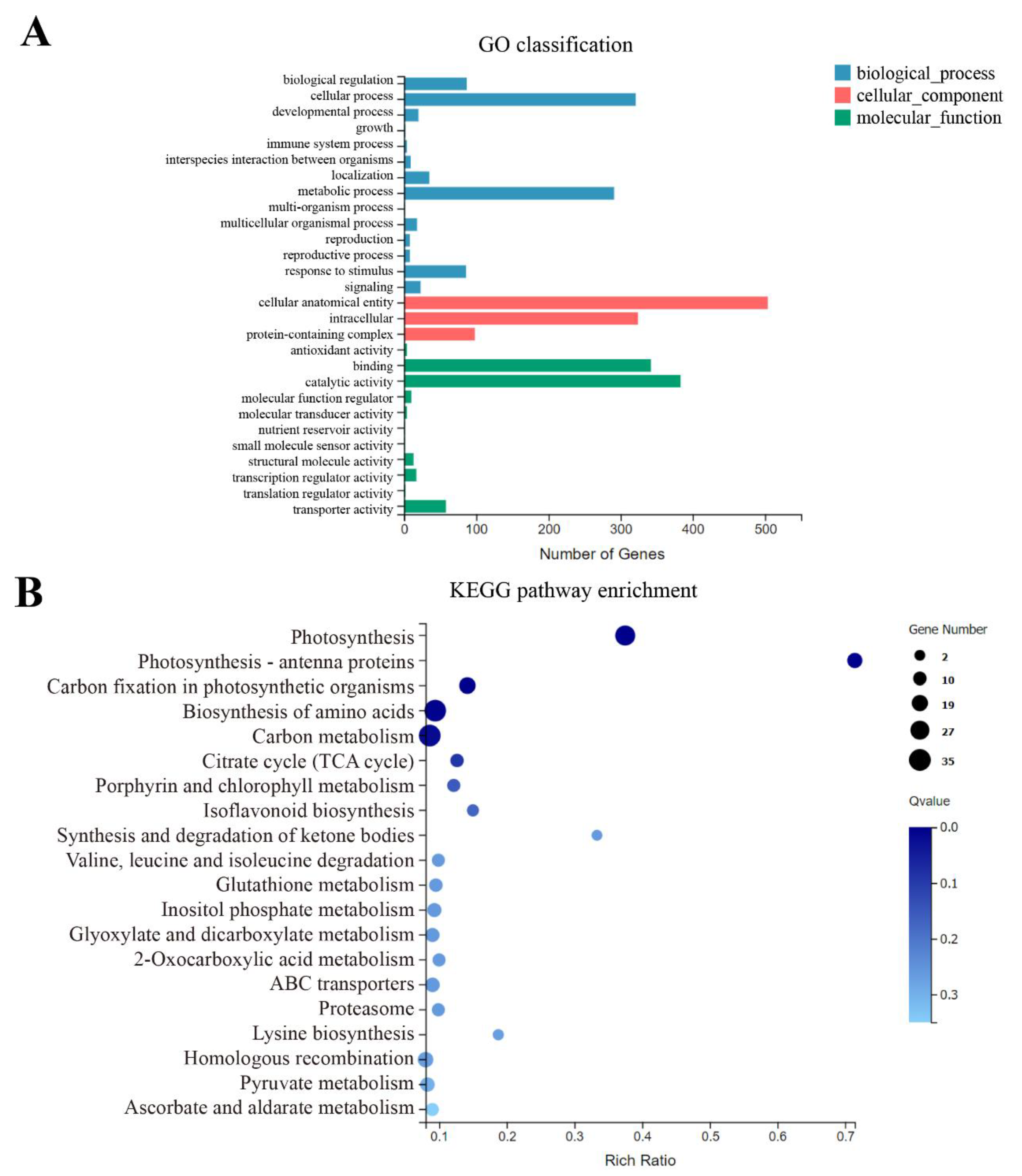
3.6. Comparative Transcriptome Analysis between se59 Mutant and Wild Type
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Mao, G.; Wei, H.; Hu, W.; Ma, Q.; Zhang, M.; Wang, H.; Yu, S. Fine mapping and molecular characterization of the virescent gene vsp in Upland cotton (Gossypium hirsutum). Theor. Appl. Genet. 2019, 132, 2069–2086. [Google Scholar] [CrossRef]
- Chen, N.; Wang, P.; Li, C.; Wang, Q.; Pan, J.; Xiao, F.; Wang, Y.; Zhang, K.; Li, C.; Yang, B.; et al. A Single Nucleotide Mutation of the IspE Gene Participating in the MEP Pathway for Isoprenoid Biosynthesis Causes a Green-Revertible Yellow Leaf Phenotype in Rice. Plant Cell Physiol. 2018, 59, 1905–1917. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.J.; Zhang, H.Q.; Wang, Y.; He, F.; Liu, J.L.; Xiao, X.; Shu, Z.F.; Li, W.; Wang, G.H. Mapped clone and functional analysis of leaf-color gene Ygl7 in a rice hybrid (Oryza sativa L. ssp. indica). PLoS ONE 2014, 9, e99564. [Google Scholar] [CrossRef] [PubMed]
- Waters, B.M.; Chu, H.-H.; DiDonato, R.J.; Roberts, L.A.; Eisley, R.B.; Lahner, B.; Salt, D.E.; Walker, E.L. Mutations in Arabidopsis yellow stripe-like1 and yellow stripe-like3 reveal their roles in metal ion homeostasis and loading of metal ions in seeds. Plant Physiol. 2006, 141, 1446–1458. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Liu, C.; An, X.; Wu, H.; Feng, Y.; Wang, H.; Sun, D. Identification and genetic mapping of a novel incompletely dominant yellow leaf color gene, Y1718, on chromosome 2BS in wheat. Euphytica 2017, 213, 141. [Google Scholar] [CrossRef]
- Marco, G.D.; Giardi, M.T.; Massacci, A.; Tricoli, D. Photosynthetic properties of leaves of a yellow green mutant of wheat compared to its wild type. Photosynth Res. 1989, 21, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Barry, C.S.; Mcquinn, R.P.; Chung, M.Y.; Besuden, A.; Giovannoni, J.J. Amino acid substitutions in homologs of the STAY-GREEN protein are responsible for the green-flesh and chlorophyll retainer mutations of tomato and pepper. Plant Physiol. 2008, 147, 179–187. [Google Scholar] [CrossRef]
- Nothnagel, P.S. Inheritance and mapping of a yellow leaf mutant of carrot (Daucus carota). Plant Breed. 2003, 122, 339–342. [Google Scholar] [CrossRef]
- Zou, J.J.; Singh, R.J.; Hymowitz, T. Association of the yellow leaf (y10) mutant to soybean chromosome 3. J. Hered. 2003, 94, 352–355. [Google Scholar] [CrossRef]
- Pasini, L.; Bruschini, S.; Bertoli, A.; Mazza, R.; Fracheboud, Y.; Marocco, A. Photosynthetic performance of cold-sensitive mutants of maize at low temperature. Physiol. Plant. 2005, 124, 362–370. [Google Scholar] [CrossRef]
- Park, S.Y.; Yu, J.W.; Park, J.S.; Li, J.; Yoo, S.C.; Lee, N.Y.; Lee, S.K.; Jeong, S.W.; Seo, H.S.; Koh, H.J.; et al. The senescence-induced staygreen protein regulates chlorophyll degradation. Plant Cell 2007, 19, 1649–1664. [Google Scholar] [CrossRef]
- Ma, X.; Sun, X.; Li, C.; Huan, R.; Sun, C.; Wang, Y.; Xiao, F.; Wang, Q.; Chen, P.; Ma, F.; et al. Map-based cloning and characterization of the novel yellow-green leaf gene ys83 in rice (Oryza sativa). Plant Physiol. Biochem. 2017, 111, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Gong, X.; Jiang, Q.; Xu, J.; Zhang, J.; Teng, S.; Lin, D.; Dong, Y. Disruption of the rice plastid ribosomal protein s20 leads to chloroplast developmental defects and seedling lethality. G3 (Bethesda) 2013, 3, 1769–1777. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Zhang, X.; He, B. A chlorophyll-deficient rice mutant with impaired chlorophyllide esterification in chlorophyll biosynthesis. Plant Physiol. 2007, 145, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Xu, J.; Chen, Y.; Mao, C.; Zhang, S.; Bai, Y.; Jiang, D.; Wu, P. Molecular cloning and characterization of OsCHR4, a rice chromatin-remodeling factor required for early chloroplast development in adaxial mesophyll. Planta 2012, 236, 1165–1176. [Google Scholar] [CrossRef]
- Cui, X.; Wang, Y.; Wu, J.; Han, X.; Gu, X.; Lu, T.; Zhang, Z. The RNA editing factor DUA1 is crucial to chloroplast development at low temperature in rice. New Phytol. 2019, 221, 834–849. [Google Scholar] [CrossRef]
- Zhang, K.; Li, Y.; Zhu, W.; Wei, Y.; Njogu, M.K.; Lou, Q.; Li, J.; Chen, J. Fine Mapping and Transcriptome Analysis of Virescent Leaf Gene v-2 in Cucumber (Cucumis sativus L.). Front. Plant Sci. 2020, 11, 570817. [Google Scholar] [CrossRef]
- Sugimoto, H.; Kusumi, K.; Tozawa, Y.; Yazaki, J.; Kishimoto, N.; Kikuchi, S.; Iba, K. The virescent-2 mutation inhibits translation of plastid transcripts for the plastid genetic system at an early stage of chloroplast differentiation. Plant Cell Physiol. 2004, 45, 985–996. [Google Scholar] [CrossRef]
- Sugimoto, H.; Kusumi, K.; Noguchi, K.; Yano, M.; Yoshimura, A.; Iba, K. The rice nuclear gene, VIRESCENT 2, is essential for chloroplast development and encodes a novel type of guanylate kinase targeted to plastids and mitochondria. Plant J. 2007, 52, 512–527. [Google Scholar] [CrossRef]
- Yoo, S.C.; Cho, S.H.; Sugimoto, H. Rice virescent3 and stripe1 encoding the large and small subunits of ribonucleotide reductase are required for chloroplast biogenesis during early leaf development. Plant Physiol. 2009, 150, 388–401. [Google Scholar] [CrossRef]
- Kusumi, K.; Sakata, C.; Nakamura, T. A plastid protein NUS1 is essential for build-up of the genetic system for early chloroplast development under cold stress conditions. Plant J. 2011, 68, 1039–1050. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Xue, D.; Li, X.; Long, Y.; Zeng, X.; Liu, Y. Characterization and molecular mapping of a new virescent mutant in rice. J. Genet. Genom. 2014, 41, 353–356. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Liu, X.; Liang, S.; Wang, R.; Li, Y.; Zhao, J.; Shao, J.; An, L.; Yu, F. A Putative Chloroplast Thylakoid Metalloprotease VIRESCENT3 Regulates Chloroplast Development in Arabidopsis thaliana. J. Biol. Chem. 2016, 291, 3319–3332. [Google Scholar] [CrossRef] [PubMed]
- Song, M.; Wei, Q.; Wang, J. Fine Mapping of CsVYL, Conferring Virescent Leaf through the Regulation of Chloroplast Development in Cucumber. Front. Plant Sci. 2018, 9, 432. [Google Scholar] [CrossRef]
- Ding, Y.; Yang, W.; Su, C.; Ma, H.; Pan, Y.; Zhang, X.; Li, J. Tandem 13-Lipoxygenase Genes in a Cluster Confers Yellow-Green Leaf in Cucumber. Int. J. Mol. Sci. 2019, 20, 3102. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Zhang, H.; Xie, C.; Wang, J.; Zhang, J.; Wang, H.; Weng, Y.; Chen, P.; Li, Y. A mutation in CsHD encoding a histidine and aspartic acid domain-containing protein leads to yellow young leaf-1 (yyl-1) in cucumber (Cucumis sativus L.). Plant Sci. 2020, 293, 110407. [Google Scholar] [CrossRef]
- Zhang, T.; Dong, X.; Yuan, X. Identification and characterization of CsSRP43, a major gene controlling leaf yellowing in cucumber. Hortic. Res. 2022, 9, 212. [Google Scholar] [CrossRef]
- Ke, X.; Shen, J.; Niu, Y. Cucumber NUCLEAR FACTOR-YC2/-YC9 target translocon component CsTIC21 in chloroplast photomorphogenesis. Plant Physiol. 2023, 192, 2822–2837. [Google Scholar] [CrossRef]
- Gao, M.; Hu, L.; Li, Y.; Weng, Y. The chlorophyll-deficient golden leaf mutation in cucumber is due to a single nucleotide substitution in CsChlI for magnesium chelatase I subunit. Theor. Appl. Genet. 2016, 129, 1961–1973. [Google Scholar] [CrossRef]
- Xiong, L.; Du, H.; Zhang, K. A Mutation in CsYL2.1 Encoding a Plastid Isoform of Triose Phosphate Isomerase Leads to Yellow Leaf 2.1 (yl2.1) in Cucumber (Cucumis sativus L.). Int. J. Mol. Sci. 2020, 22, 322. [Google Scholar] [CrossRef]
- Miao, H.; Zhang, S.; Wang, M.; Wang, Y.; Weng, Y.; Gu, X. Fine Mapping of Virescent Leaf Gene v-1 in Cucumber (Cucumis sativus L.). Int. J. Mol. Sci. 2016, 17, 1602. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Wang, J.; Xing, G.; Li, M.; Li, S. Integrating physiology, genetics, and transcriptome to decipher a new thermo-sensitive and light-sensitive virescent leaf gene mutant in cucumber. Front. Plant Sci. 2022, 13, 972620. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Wang, R. Change Law of Hyperspectral Data in Related with Chlorophyll and Carotenoid in Rice at Different Developmental Stages. Rice Science 2004, 11, 274–282. [Google Scholar]
- Abe, A.; Kosugi, S.; Yoshida, K.; Natsume, S.; Takagi, H.; Kanzaki, H.; Matsumura, H.; Yoshida, K.; Mitsuoka, C.; Tamiru, M.; et al. Genome sequencing reveals agronomically important loci in rice using MutMap. Nat. Biotechnol. 2012, 30, 174–178. [Google Scholar] [CrossRef]
- Takagi, H.; Abe, A.; Yoshida, K.; Kosugi, S.; Natsume, S.; Mitsuoka, C.; Uemura, A.; Utsushi, H.; Tamiru, M.; Takuno, S.; et al. QTL-seq: Rapid mapping of quantitative trait loci in rice by whole genome resequencing of DNA from two bulked populations. Plant J. 2013, 74, 174–183. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Jenkins, C.L. Effects of the Phosphoenolpyruvate Carboxylase Inhibitor 3,3-Dichloro-2-(Dihydroxyphosphinoylmethyl) propenoate on Photosynthesis: C(4) Selectivity and Studies on C(4) Photosynthesis. Plant Physiol. 1989, 89, 1231–1237. [Google Scholar] [CrossRef]
- Fischer, R.A.; Rees, D.; Sayre, K.D.; Lu, Z.M.; Condon, A.G.; Saavedra, A.L. Wheat yield progress associated with higher stomatal conductance and photosynthetic rate, and cooler canopies. Crop Sci. 1998, 38, 1467–1475. [Google Scholar] [CrossRef]
- Zhou, X.S.; Xing, W.D.; Quan, S.S. High photosynthetic efficiency of a rice (Oryza sativa L.) xantha mutant. Photosynthetica 2006, 44, 316–319. [Google Scholar] [CrossRef]
- Gough, J.; Karplus, K.; Hughey, R.; Chothia, C. Assignment of homology to genome sequences using a library of hidden Markov models that represent all proteins of known structure. J. Mol. Biol. 2001, 313, 903–919. [Google Scholar] [CrossRef]
- Ruan, Y.L.; Jin, Y.; Yang, Y.J.; Li, G.J.; Boyer, J.S. Sugar input, metabolism, and signaling mediated by invertase: Roles in development, yield potential, and response to drought and heat. Mol. Plant 2010, 3, 942–955. [Google Scholar] [CrossRef] [PubMed]
- Tauzin, A.S.; Giardina, T. Sucrose and invertases, a part of the plant defense response to the biotic stresses. Front. Plant Sci. 2014, 5, 293. [Google Scholar] [CrossRef] [PubMed]
- Liao, S.; Wang, L.; Li, J.; Ruan, Y.L. Cell wall invertase is essential for ovule development through sugar signaling rather Than provision of carbon nutrients. Plant Physiol. 2020, 183, 1126–1144. [Google Scholar] [CrossRef] [PubMed]
- Bate, N.J.; Niu, X.; Wang, Y.; Reimann, K.S.; Helentjaris, T.G. An invertase inhibitor from maize localizes to the embryo surrounding region during early kernel development. Plant Physiol. 2014, 134, 246–254. [Google Scholar] [CrossRef] [PubMed]
- Raiola, A.; Camardella, L.; Giovane, A. Two Arabidopsis thaliana genes encode functional pectin methylesterase inhibitors11The industrial utilization of Arabidopsis and kiwi PMEIs is patent pending it, no. RM2003A000346. FEBS Lett. 2004, 557, 199–203. [Google Scholar] [CrossRef]
- Lionetti, V. PECTOPLATE: The simultaneous phenotyping of pectin methylesterases, pectinases, and oligogalacturonides in plants during biotic stresses. Front. Plant Sci. 2015, 6, 331. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Material | Pn (μm/m2·s) | Tr (μm/m2·s) |
|---|---|---|
| HB | 30.07 ± 2.3 | 3.3 ± 3.4 |
| se59 | 19.8 ± 4.49 * | 5.93 ± 0.46 |
| Material | F0 | Fv/Fm | Y (II) | qP | NPQ | ETR |
|---|---|---|---|---|---|---|
| HB | 416 ± 34.6 | 0.71 ± 0.06 | 0.60 ± 0.02 | 0.83 ± 0.3 | 0.06 ± 0.08 | 11.1 ± 0.4 |
| se59 | 403 ± 6.81 | 0.67 ± 0.01 | 0.53 ± 0.47 | 0.79 ± 0.5 | 0.18 ± 0.01 * | 9.8 ± 0. 8 |
| Chromo-Some | Position | Gene | Reference | Alteration | SNP_index | Type | Amino Acid Change |
|---|---|---|---|---|---|---|---|
| Chr.6 | 12112564 | CsaV3_3G 016210 | G | A | 1 | exon | G to S |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, Y.; Liao, L.; Liu, L.; Xiao, L.; Zhou, Z.; Zhou, Y.; Hu, Z.; Liu, S. CsSE59 Encoding Invertase/Pectin Methyl Esterase Inhibitor Is a Candidate Gene Conferring the Virescent True Leaf Phenotype in Cucumber. Horticulturae 2023, 9, 951. https://doi.org/10.3390/horticulturae9090951
Zhou Y, Liao L, Liu L, Xiao L, Zhou Z, Zhou Y, Hu Z, Liu S. CsSE59 Encoding Invertase/Pectin Methyl Esterase Inhibitor Is a Candidate Gene Conferring the Virescent True Leaf Phenotype in Cucumber. Horticulturae. 2023; 9(9):951. https://doi.org/10.3390/horticulturae9090951
Chicago/Turabian StyleZhou, Yuelong, Liting Liao, Liu Liu, Lingdi Xiao, Zixian Zhou, Yong Zhou, Zhaoyang Hu, and Shiqiang Liu. 2023. "CsSE59 Encoding Invertase/Pectin Methyl Esterase Inhibitor Is a Candidate Gene Conferring the Virescent True Leaf Phenotype in Cucumber" Horticulturae 9, no. 9: 951. https://doi.org/10.3390/horticulturae9090951