
Breeding Approaches for Controlled Conditions of Artificial Light Culture for Small Radish and Radish (Raphanus sativus L.)
 , , , and
, , , and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Using the Methodology of Accelerated Transgressive Breeding in the Creating of New Forms of Small Radish and Radish for Controlled Conditions of Light Culture
- -
- taking into account the ecological and genetic organization of quantitative traits;
- -
- taking into account the modular structure of the targeted traits, represented by the complex of the resulting trait and its components;
- -
- taking into account the independence of inheritance of modular components of selected traits;
- -
- the use of growing regimes, which ensure a decrease in the modification and an increase in the clarity of the manifestation of genotypic variability, as well as regimes that accelerate the development of plants in CEAL;
- -
- assessment of the degree of manifestation of the components of the targeted traits in the modeling in CEAL of the main limiting environmental factors operating in the growing region;
- -
- selection of parental pairs based on the principle of favorable complementarity of different components of selectable traits determined by genes or blocks of genes that provide transgressions due to complementary interactions;
- -
- carrying out and studying hybrids F1 and F2, comparing them with parental forms and among themselves under controlled conditions, in order to obtain and identify transgressive genotypes with a predictable complex of economically valuable traits;
- -
- obtaining of stable lines according to the studied traits from selected transgressive genotypes by inbreeding and stabilizing the selection using growing regimes that accelerate generative development [22].
2.2. Growing Plants under Controlled Conditions
2.3. Statistical Analysis
3. Results
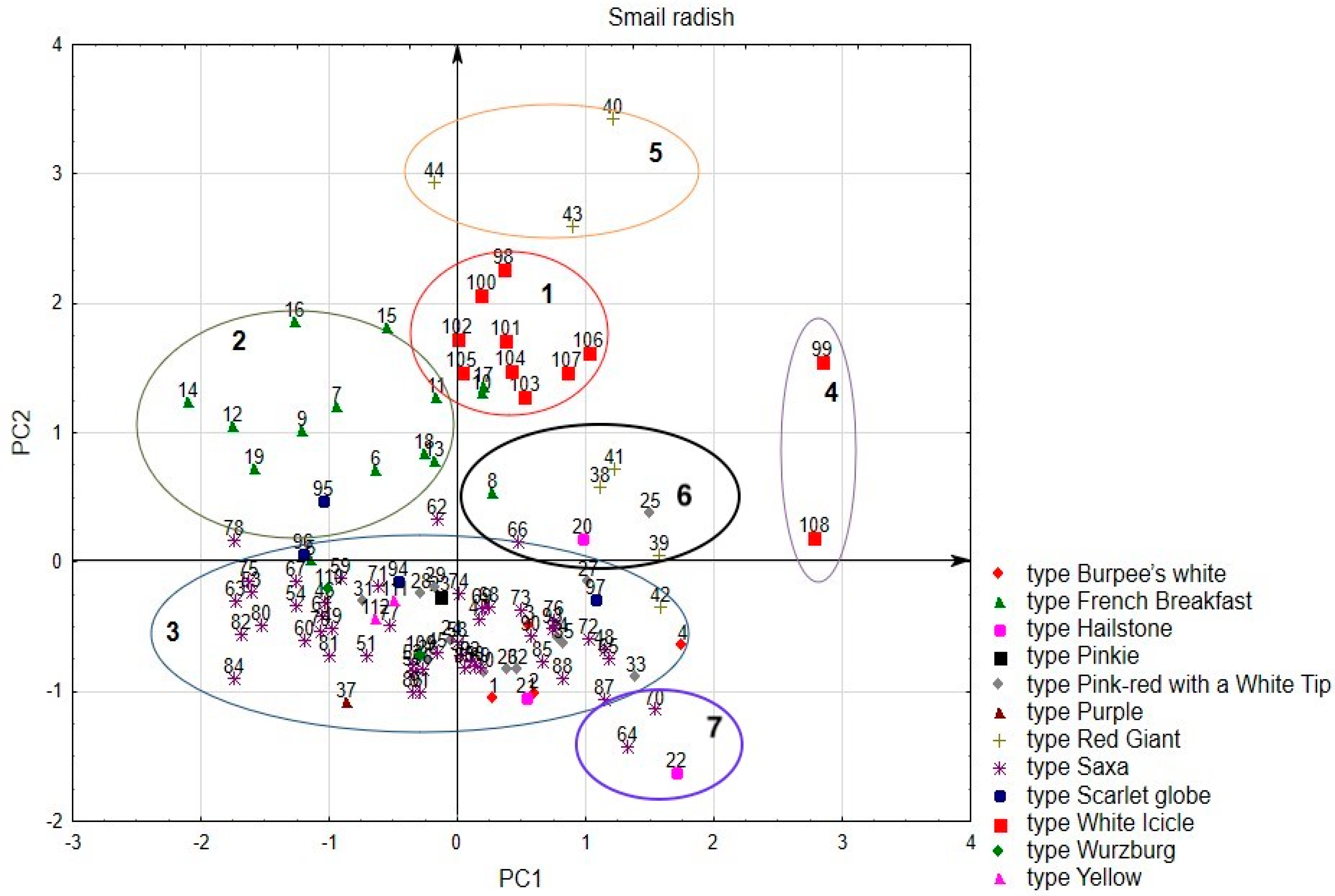
3.1. Principal Component Analysis (PCA)
3.2. Breeding under Artificial Light Culture Conditions and Obtaining Transgressions by the Mass of the Root of Raphanus sativus L.
3.3. Resistance to Premature Bolting
3.4. The Compactness of a Leaf Rosette and Other Economically Valuable Traits
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Hansen, J.; Hellin, J.; Rosenstock, T.; Fisher, E.; Cairns, J.; Stirling, C.; Campbell, B. Climate risk management and rural poverty reduction. Agric. Syst. 2019, 172, 28–46. [Google Scholar] [CrossRef]
- Vermeulen, S.J.; Dinesh, D.; Howden, S.M.; Cramer, L.; Thornton, P.K. Transformation in practice: A review of empirical cases of transformational adaptation in agriculture under climate change. Front. Sustain. Food Syst. 2018, 2, 65. [Google Scholar] [CrossRef] [Green Version]
- Ksenofontov, M.Y.; Polzikov, D.A. On the issue of the impact of climate change on the development of Russian agriculture in the long term. Stud. Russ. Econ. Dev. 2020, 31, 304–311. [Google Scholar] [CrossRef]
- Vyas, S.; Khatri-Chhetri, A.; Aggarwal, P.; Thornton, P.; Campbell, B.M. Perspective: The gap between intent and climate action in agriculture. Glob. Food Sec. 2022, 32, 100612. [Google Scholar] [CrossRef]
- Rouphael, Y.; Kyriacou, M.C.; Petropoulos, S.A.; De Pascale, S.; Colla, G. Improving vegetable quality in controlled environments. Sci. Hortic. 2018, 234, 275–289. [Google Scholar] [CrossRef]
- Martin, M.; Weidner, T.; Gullström, C. Estimating the potential of building integration and regional synergies to improve the environmental performance of urban vertical farming. Front. Sustain. Food Syst. 2022, 6, 109. [Google Scholar] [CrossRef]
- Kozai, T.; Niu, G. Role of the plant factory with artificial lighting (PFAL) in urban areas. In Plant Factory, 2nd ed.; Kozai, T., Niu, G., Takagaki, M., Eds.; Academic Press: London, UK, 2020; pp. 7–34. ISBN 978-0-12-816691-8. [Google Scholar]
- Shamshiri, R.; Kalantari, F.; Ting, K.C.; Thorp, K.R.; Hameed, I.A.; Weltzien, C.; Shad, Z.M. Advances in greenhouse automation and controlled environment agriculture: A transition to plant factories and urban agriculture. Int. J. Agric. Biol. Eng. 2018, 11, 1–22. [Google Scholar] [CrossRef]
- Saito, K.; Ishigami, Y.; Goto, E. Evaluation of the light environment of a plant factory with artificial light by using an optical simulation. Agronomy 2020, 10, 1663. [Google Scholar] [CrossRef]
- Gómez, C.; Currey, C.J.; Dickson, R.W.; Kim, H.J.; Hernández, R.; Sabeh, N.C.; Burnett, S.E. Controlled environment food production for urban agriculture. HortScience 2019, 54, 1448–1458. [Google Scholar] [CrossRef]
- Ahmed, H.A.; Tong, Y.; Li, L.; Sahari, S.Q.; Almogahed, A.M.; Cheng, R. Integrative effects of CO2 concentration, illumination intensity and air speed on the growth, gas exchange and light use efficiency of lettuce plants grown under artificial lighting. Horticulturae 2022, 8, 270. [Google Scholar] [CrossRef]
- O'sullivan, C.A.; Bonnett, G.D.; McIntyre, C.L.; Hochman, Z.; Wasson, A.P. Strategies to improve the productivity, product diversity and profitability of urban agriculture. Agric. Syst. 2019, 174, 133–144. [Google Scholar] [CrossRef]
- Lubna, F.A.; Lewus, D.C.; Shelford, T.J.; Both, A.J. What you may not realize about vertical farming. Horticulturae 2022, 8, 322. [Google Scholar] [CrossRef]
- Van Delden, S.H.; SharathKumar, M.; Butturini, M.; Graamans, L.J.A.; Heuvelink, E.; Kacira, M.; Marcelis, L.F.M. Current status and future challenges in implementing and upscaling vertical farming systems. Nat. Food 2021, 2, 944–956. [Google Scholar] [CrossRef] [PubMed]
- SharathKumar, M.; Heuvelink, E.; Marcelis, L.F. Vertical farming: Moving from genetic to environmental modification. Trends Plant Sci. 2020, 25, 724–727. [Google Scholar] [CrossRef]
- Folta, K.M. Breeding new varieties for controlled environments. Plant Biol. 2019, 21, 6–12. [Google Scholar] [CrossRef]
- Awika, H.O.; Mishra, A.K.; Gill, H.; DiPiazza, J.; Avila, C.A.; Joshi, V. Selection of nitrogen responsive root architectural traits in spinach using machine learning and genetic correlations. Sci. Rep. 2021, 11, 9536. [Google Scholar] [CrossRef]
- Artemyeva, A.M.; Sinyavina, N.G.; Panova, G.G.; Chesnokov, Y.V. Biological features of Brassica rapa L. vegetable leafy crops when growing in an intensive light culture. Agric. Biol. 2021, 56, 103–120. [Google Scholar] [CrossRef]
- Egorova, K.V.; Sinyavina, N.G.; Artemyeva, A.M.; Kocherina, N.V.; Chesnokov, Y.V. QTL Analysis of the Content of Some Bioactive Compounds in Brassica rapa L. Grown under Light Culture Conditions. Horticulturae 2021, 7, 583. [Google Scholar] [CrossRef]
- Kurina, A.B.; Kornyukhin, D.L.; Solovyeva, A.E.; Artemyeva, A.M. Genetic diversity of phenotypic and biochemical traits in VIR radish (Raphanus sativus L.) germplasm collection. Plants 2021, 10, 1799. [Google Scholar] [CrossRef]
- Samantara, K.; Bohra, A.; Mohapatra, S.R.; Prihatini, R.; Asibe, F.; Singh, L.; Varshney, R.K. Breeding More Crops in Less Time: A Perspective on Speed Breeding. Biology 2022, 11, 275. [Google Scholar] [CrossRef]
- Kochetov, A.A.; Mirskaya, G.V.; Sinyavina, N.G.; Egorova, K.V. Transgressive Breeding: A methodology for accelerated creation of new forms of plants with a predictable complex of economically valuable traits. Russ. Agric. Sci. 2021, 47, 40–50. [Google Scholar] [CrossRef]
- Wanga, M.A.; Shimelis, H.; Mashilo, J.; Laing, M.D. Opportunities and challenges of speed breeding: A review. Plant Breed. 2021, 140, 185–194. [Google Scholar] [CrossRef]
- Ahmar, S.; Gill, R.A.; Jung, K.H.; Faheem, A.; Qasim, M.U.; Mubeen, M.; Zhou, W. Conventional and molecular techniques from simple breeding to speed breeding in crop plants: Recent advances and future outlook. Int. J. Mol. Sci. 2020, 21, 2590. [Google Scholar] [CrossRef] [Green Version]
- Watson, A.; Ghosh, S.; Williams, M.J.; Cuddy, W.S.; Simmonds, J.; Rey, M.D.; Hickey, L.T. Speed breeding is a powerful tool to accelerate crop research and breeding. Nat. Plants 2018, 4, 23–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chesnokov, Y.V.; Kanash, E.V.; Mirskaya, G.V.; Kocherina, N.V.; Rusakov, D.V.; Lohwasser, U.; Börner, A. QTL mapping of diffuse reflectance indices of leaves in hexaploid bread wheat (Triticum aestivum L.). Russ. J. Plant Physiol. 2019, 66, 77–86. [Google Scholar] [CrossRef]
- Chesnokov, Y.V.; Mirskaya, G.V.; Kanash, E.V.; Kocherina, N.V.; Rusakov, D.V.; Lohwasser, U.; Börner, A. QTL identification and mapping in soft spring wheat (Triticum aestivum L.) under controlled agroecological and biological testing area conditions with and without nitrogen fertilizer. Russ. J. Plant Physiol. 2018, 65, 123–135. [Google Scholar] [CrossRef]
- Yanaeva, D.A. Cultivars and hybrids of the garden radish for the growing according to cassette technology. Potato Veg. 2015, 2, 19–21. (In Russian) [Google Scholar]
- Singh, B.K. Radish (Raphanus sativus L.): Breeding for higher yield, better quality and wider adaptability. In Advances in Plant Breeding Strategies: Vegetable Crops; Al-Khayri, J.M., Jain, S.M., Johnson, D.V., Eds.; Springer: Cham, Switzerland, 2021; Volume 8, pp. 275–304. ISBN 978-3-030-66964-5. [Google Scholar]
- Gamba, M.; Asllanaj, E.; Raguindin, P.F.; Glisic, M.; Franco, O.H.; Minder, B.; Muka, T. Nutritional and phytochemical characterization of radish (Raphanus sativus): A systematic review. Trends Food Sci. Technol. 2021, 113, 205–218. [Google Scholar] [CrossRef]
- Gutiérrez, R.M.P.; Perez, R.L. Raphanus sativus (Radish): Their chemistry and biology. Sci. World J. 2004, 4, 811–837. [Google Scholar] [CrossRef]
- Keyata, E.O.; Tola, Y.B.; Bultosa, G.; Forsido, S.F. Proximate, mineral, and anti-nutrient compositions of underutilized plants of Ethiopia: Figl (Raphanus sativus L.), Girgir (Eruca sativa L) and Karkade (Hibiscus sabdariffa): Implications for in-vitro mineral bioavailability. Food Res. Int. 2020, 137, 109724. [Google Scholar] [CrossRef]
- Manivannan, A.; Kim, J.H.; Kim, D.S.; Lee, E.S.; Lee, H.E. Deciphering the nutraceutical potential of Raphanus sativus—A comprehensive overview. Nutrients 2019, 11, 402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goyeneche, R.; Roura, S.; Ponce, A.; Vega-Gálvez, A.; Quispe-Fuentes, I.; Uribe, E.; Di Scala, K. Chemical characterization and antioxidant capacity of red radish (Raphanus sativus L.) leaves and roots. J. Funct. Foods 2015, 16, 256–264. [Google Scholar] [CrossRef]
- Sarkar, T.; Salauddin, M.; Roy, S.; Chakraborty, R.; Rebezov, M.; Shariati, M.A.; Rengasamy, K.R.R. Underutilized green leafy vegetables: Frontier in fortified food development and nutrition. Crit. Rev. Food Sci. Nutr. 2022, 11, 1–55. [Google Scholar] [CrossRef] [PubMed]
- Noman, O.M.; Nasr, F.A.; Alqahtani, A.S.; Al-zharani, M.; Cordero, M.A.W.; Alotaibi, A.A.; Daoud, A. Comparative study of antioxidant and anticancer activities and HPTLC quantification of rutin in white radish (Raphanus sativus L.) leaves and root extracts grown in Saudi Arabia. Open Chem. 2021, 19, 408–416. [Google Scholar] [CrossRef]
- Koley, T.K.; Khan, Z.; Oulkar, D.; Singh, B.K.; Maurya, A.; Singh, B.; Banerjee, K. High resolution LC-MS characterization of phenolic compounds and the evaluation of antioxidant properties of a tropical purple radish genotype. Arab. J. Chem. 2020, 13, 1355–1366. [Google Scholar] [CrossRef]
- Jahangir, M.; Abdel-Farid, I.B.; de Vos, C.H.R.; Jonker, H.H.; Choi, Y.H.; Verpoorte, R. Metabolomic variation of Brassica rapa var. rapa (var. raapstelen) and Raphanus sativus L. at different developmental stages. Pak. J. Bot. 2014, 46, 1445–1452. [Google Scholar]
- Li, X.; Wang, J.; Qiu, Y.; Wang, H.; Wang, P.; Zhang, X.; Li, X. SSR-sequencing reveals the inter-and intraspecific genetic variation and phylogenetic relationships among an extensive collection of Radish (Raphanus) germplasm resources. Biology 2021, 10, 1250. [Google Scholar] [CrossRef]
- Panova, G.G.; Chernousov, I.N.; Udalova, O.R.; Alexandrov, A.V.; Karmanov, I.V.; Anikina, L.M.; Yakushev, V.P. Scientific basis for large year-round yields of high-quality crop products under artificial lighting. Russ. Agric. Sci. 2015, 41, 335–339. [Google Scholar] [CrossRef]
- Ermakov, E.I.; Zheltov, Y.I.; Milto, N.E.; Kucherov, V.I. Soil for Growing Plants “Agrophyte”. Russian Federation Patent 2081555, 13 July 1997. (In Russian). [Google Scholar]
- Kochetov, A.A.; Sinyavina, N.G. Small Radish Raphanus sativus var. sativus Peterburgskiy Fioletovyy. Russian Federation Patent 11,518 (8,058,521), 25 March 2021. (In Russian). [Google Scholar]
- Kochetov, A.A.; Artem’eva, A.M. Daikon Peterburgskiy. Russian Federation Patent 6392 (8,953,530), 22 March 2012. (In Russian). [Google Scholar]
- Sazonova, L.V. Guidelines for the Study and Maintenance of the World Collection of Root Vegetables; VIR: Leningrad, Russia, 1989; p. 88. (In Russian) [Google Scholar]
- Cattell, R.B. The Scree test for the number of factors. Multivar. Behav. Res. 1966, 1, 245–276. [Google Scholar] [CrossRef]
- Dospekhov, B.A. Field Experience Methodology (with the Basics of Statistical Processing of Research Results), 5th ed., add and revised; Agropromizdat: Moscow, Russia, 1985; 351p. [Google Scholar]
- Benildo, G.D.L.R. Genomic and epigenomic bases of transgressive segregation–New breeding paradigm for novel plant phenotypes. Plant Sci. 2019, 288, 110213. [Google Scholar]
- Manzoor, A.; Bashir, M.A.; Naveed, M.S.; Cheema, K.L.; Cardarelli, M. Role of different abiotic factors in inducing pre-harvest physiological disorders in radish (Raphanus sativus). Plants 2021, 10, 2003. [Google Scholar] [CrossRef] [PubMed]
- Iwata, H.; Niikura, S.; Matsuura, S.; Takano, Y.; Ukai, Y. Interaction between genetic effects and soil type in diallel analysis of root shape and size of Japanese radish (Raphanus sativus L.). Breed. Sci. 2004, 54, 313–318. [Google Scholar] [CrossRef] [Green Version]
- Ting, F.S.-T.; Wren, M.J. Storage organ development in radish (Raphanus sativus L.). 1. A comparison of development in seedlings and rooted cuttings of two contrasting varieties. Ann. Bot. 1980, 46, 267–276. [Google Scholar] [CrossRef]
- Dolan, L.; Janmaat, K.; Willemsen, V.; Linstead, P.; Poethig, S.; Roberts, K.; Scheres, B. Cellular organisation of the Arabidopsis thaliana root. Development 1993, 119, 71–84. [Google Scholar] [CrossRef]
- Zaki, H.E.; Yokoi, S.; Takahata, Y. Identification of genes related to root shape in radish (Raphanus sativus) using suppression subtractive hybridization. Breed. Sci. 2010, 60, 130–138. [Google Scholar] [CrossRef] [Green Version]
- Makarova, G.A.; Ivanova, T.I. Inheritance of root and leaf characteristics in radish plants. Genetiks 1983, 19, 304–311. (In Russian) [Google Scholar]
- Wei, Q.; Wang, J.; Wang, W.; Hu, H.; Yan, Y.; Bao, C.; Hu, T. Identification of QTLs controlling radish root shape using multiple populations. Horticulturae 2022, 8, 931. [Google Scholar] [CrossRef]
- Yu, X.; Choi, S.R.; Dhandapani, V.; Rameneni, J.J.; Li, X.; Pang, W.; Lim, Y.P. Quantitative trait loci for morphological traits and their association with functional genes in Raphanus sativus. Front. Plant Sci. 2016, 7, 255. [Google Scholar] [CrossRef] [Green Version]
- Tsuro, M.; Suwabe, K.; Kubo, N.; Matsumoto, S.; Hirai, M. Mapping of QTLs controlling root shape and red pigmentation in radish, Raphanus sativus L. Breed. Sci. 2008, 58, 55–61. [Google Scholar] [CrossRef] [Green Version]
- Sinyavina, N.G.; Kochetov, A.A.; Egorova, K.V.; Kocherina, N.V.; Chesnokov, Y.V. Genetic-Biochemical Studies and Morphobiological Assessment of Small Radish (Raphanus sativus L.) under Artificial Light Culture Conditions. Russ. J. Genet. 2022, 58, 662–670. [Google Scholar] [CrossRef]
- Kochetov, A.A.; Sinyavina, N.G. Creation of new forms of Raphanus sativus L. with the predicted complex of economically valuable traits using the methodology of accelerated plant breeding. Potato Veg. 2019, 10, 29–34, (In Russian with English Abstract). [Google Scholar]
- Nie, S.; Li, C.; Xu, L.; Wang, Y.; Huang, D.; Muleke, E.M.; Liu, L. De novo transcriptome analysis in radish (Raphanus sativus L.) and identification of critical genes involved in bolting and flowering. BMC Genom. 2016, 17, 389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, T.; Wei, Q.; Wang, W.; Hu, H.; Mao, W.; Zhu, Q.; Bao, C. Genome-wide identification and characterization of CONSTANS-like gene family in radish (Raphanus sativus). PLoS ONE 2018, 13, e0204137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Qiu, X.; Tan, Q.; Xiao, Q.; Mei, S. A comparative metabolomics study of flavonoids in radish with different skin and flesh colors (Raphanus sativus L.). J. Agric. Food Chem. 2020, 68, 14463–14470. [Google Scholar] [CrossRef]
- Ugarova, S.V.; Zelenin, A.V. The use the genetically difficultly inherited trait of purple root color in breeding program for the complicated trait in radish. Veg. Crop Russ. 2017, 4, 66–69, (In Russian with English Abstract). [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Wei, Q.; Wang, W.; Hu, H.; Yan, Y.; Wang, Y.; Bao, C. Understanding the nutraceutical diversity through a comparative analysis of the taproot metabolomes of different edible radish types via UHPLC–Q–TOF–MS. Food Chem. 2023, 403, 134469. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Factor Loadings (Varimax Raw) Extraction: Principal Component | |||||
|---|---|---|---|---|---|---|
| PC1 | PC2 | PC3 | PC4 | PC5 | PC6 | |
| Rosette Height | 0.89 | 0.25 | 0.05 | 0.08 | 0.15 | 0.06 |
| Rosette Diameter | 0.74 | 0.15 | 0.17 | 0.20 | 0.09 | 0.25 |
| Leaf Length | 0.89 | 0.09 | 0.07 | 0.13 | 0.26 | 0.10 |
| Leaf Width | 0.91 | 0.12 | 0.05 | 0.17 | 0.05 | 0.00 |
| Root Length | 0.23 | 0.86 | 0.08 | 0.37 | 0.15 | −0.04 |
| Root Diameter | −0.02 | −0.88 | −0.07 | 0.39 | 0.01 | 0.13 |
| Root Index | 0.21 | 0.94 | 0.07 | 0.14 | 0.14 | −0.05 |
| Plant Weight | 0.72 | 0.14 | 0.15 | 0.50 | 0.25 | 0.13 |
| Root Weight | 0.29 | 0.06 | 0.10 | 0.91 | 0.14 | 0.10 |
| Duration of Vegetative Phase | 0.29 | 0.18 | 0.17 | 0.16 | 0.91 | −0.04 |
| Number of bolted plants | 0.15 | −0.14 | −0.15 | 0.09 | −0.03 | 0.95 |
| Degree of pubescence | −0.13 | −0.12 | −0.95 | −0.09 | −0.15 | 0.15 |
| Expl.Var | 3.75 | 2.60 | 1.05 | 1.53 | 1.07 | 1.05 |
| Prp.Totl | 0.31 | 0.22 | 0.09 | 0.13 | 0.09 | 0.09 |
| % total variance | 45.9 | 19.1 | 9.8 | 7.7 | 4.6 | 4.8 |
| % cumulative proportion of variance | 45.9 | 65.1 | 74.8 | 82.5 | 87.4 | 92.1 |
| Accession | Plant Weight. g | Root Weight. g | Leaves Weight. g | Ei | Ai | Rosette Height. cm | Rosette Diameter. cm | Number of Leaves | Leaf Length.cm | Leaf Width.cm | Root Length.cm | Root Diameter. cm | Ri |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ‘Viola’ Cv | 25.02 ± 8.41 a 33.59 | 14.44 ± 5.91 a 40.92 | 10.58 ± 3.39 a 32.02 | 0.57 ± 0.08 a 13.96 | 1.42 ± 0.54 37.89 | 17.26 ± 3.03 a 17.57 | 19.53 ± 2.52 a 12.93 | 5.58 ± 0.69 a 12.41 | 21.42 ± 2.76 a 12.86 | 6.32 ± 0.84 a 13.25 | 3.78 ± 0.62 a 16.41 | 3.29 ± 0.67 a 20.47 | 1.19 ± 0.28 a 23.82 |
| ‘Pernot’ Cv | 29.91 ± 13.20 b 44.15 | 18.86 ± 8.60 b 45.59 | 11.04 ± 5.64 b 51.10 | 0.63 ± 0.07 11.10 | 1.80 ± 0.49 27.36 | 18.07 ± 2.94 b 16.29 | 19.61 ± 3.25 b 16.56 | 5.75 ± 0.75 b 13.07 | 21.53 ± 3.84 b 17.79 | 6.05 ± 1.47 b 24.26 | 8.29 ± 1.67 ab 20.11 | 1.94 ± 0.30 ab 15.35 | 4.36 ± 1.06 ab 24.23 |
| F1 ‘Viola’ × ‘Pernot’ Cv | 77.81 ± 25.69 abc 33.01 | 48.11 ± 16.94 abc 35.21 | 29.69 ± 11.26 abc 37.94 | 0.62 ± 0.07 11.88 | 1.71 ± 0.51 29.76 | 24.10 ± 1.48 abc 6.15 | 23.00 ± 1.89 abc 8.23 | 6.70 ± 0.73 ab 10.93 | 28.50 ± 3.84 abc 13.47 | 9.80 ± 1.65 abc 16.83 | 6.95 ± 1.46 abc 21.06 | 3.66 ± 0.58 bc 15.94 | 1.90 ± 0.31 ab 16.25 |
| ‘PF’ * (cultivar) Cv | 48.00 ± 13.66 abcd 28.45 | 30.75 ± 10.51 abcd 34.17 | 17.25 ± 5.09 abc 29.52 | 0.64 ± 0.06 a 9.72 | 1.84 ± 0.56 30.37 | 18.18 ± 2.26 cd 12.42 | 19.92 ± 2.50 cd 12.42 | 6.79 ± 0.61 ab 9.05 | 22.51 ± 2.63 cd 11.70 | 7.04 ± 1.05 bc 14.88 | 5.83 ± 1.00 abcd 17.18 | 3.32 ± 0.58 bd 17.44 | 1.79 ± 0.36 ab 19.97 |
| ‘PK’ ** (line) Cv | 41.37 ± 12.97 abc 31.35 | 26.53 ± 8.59 abc 32.38 | 14.84 ± 4.78 c 32.21 | 0.64 ± 0.09 a 14.06 | 1.79 ± 0.56 31.28 | 17.33 ± 1.67 c 9.64 | 18.84 ± 2.13 c 11.31 | 7.03 ± 0.61 ab 8.66 | 20.25 ± 3.13 c 17.15 | 6.69 ± 0.92 c 13.75 | 5.55 ± 0.85 abc 15.13 | 3.24 ± 0.57 b 17.59 | 1.71 ± 0.33 ab 19.30 |
| Accession | Plant Weight. g | Root Weight. g | Leaves Weight. g | Ei | Ai | Rosette Height. cm | Rosette Diameter. cm | Number of Leaves | Leaf Length. cm | Leaf Width. cm | Root Length. cm | Root Diameter. cm | Ri |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ‘Octava’ * Cv | 27.01 ± 10.85 a 40.19 | 14.47 ± 7.47 a 51.63 | 12.54 ± 3.89 a 31.03 | 0.51 ± 0.09 a 18.53 | 1.11 ± 0.38 a 34.35 | 20.56 ± 4.33 a 21.06 | 23.11 ± 2.35 a 10.16 | 6.28 ± 0.83 a 13.16 | 26.72 ± 5.33 a 19.96 | 8.97 ± 1.79 a 20.00 | 4.27 ± 1.22 a 28.63 | 3.17 ± 0.73 a 22.98 | 1.38 ± 0.41 a 29.90 |
| ‘Viola’ * Cv | 25.02 ± 8.41 b 33.59 | 14.44 ± 5.91 b 40.92 | 10.58 ± 3.39 b 32.02 | 0.57 ± 0.08 b 13.96 | 1.42 ± 0.54 b 37.89 | 17.26 ± 3.03 ab 17.57 | 19.53 ± 2.52 b 12.93 | 5.50 ± 0.69 b 12.41 | 21.42 ± 2.76 ab 12.86 | 6.32 ± 0.84 ab 13.25 | 3.78 ± 0.62 b 16.41 | 3.29 ± 0.67 b 20.47 | 1.19 ± 0.28 b 23.82 |
| ‘Peterburgskiy’ ** Cv | 90.84 ± 57.07 abc 62.83 | 47.18 ± 38.01 abc 80.56 | 43.65 ± 20.99 abc 48.08 | 0.47 ± 0.13 bc 27.12 | 0.99 ± 0.49 c 49.87 | 22.31 ± 2.18 bc 9.77 | 33.86 ± 6.96 abc 20.56 | 10.63 ± 2.86 abc 26.91 | 27.13 ± 4.53 bc 16.69 | 8.19 ± 1.67 bc 20.37 | 11.15 ± 2.57 abc 23.04 | 2.78 ± 0.89 c 31.99 | 4.27 ± 1.17 abc 27.50 |
| F1 ‘Octava’ × ‘Peterburgskiy’ Cv | 108.80 ± 33.10 abd 30.42 | 79.82 ± 30.03 abcd 37.62 | 28.98 ± 13.10 abc 45.20 | 0.72 ± 0.11 abc 14.64 | 3.12 ± 1.48 abcd 47.37 | 23.50 ± 2.93 bd 12.46 | 25.86 ± 3.24 bcd 12.59 | 8.36 ± 1.39 abc 16.66 | 26.57 ± 1.45 b 5.47 | 8.86 ± 1.36 b 15.41 | 9.95 ± 1.68 abd 16.89 | 4.19 ± 0.74 abcd 17.61 | 2.42 ± 0.50 abcd 20.50 |
| F1 ‘Viola’ × ‘Peterburgskiy’ Cv | 148.09 ± 37.15 abce 25.09 | 105.45 ± 31.14 abce 29.53 | 42.64 ± 10.05 abd 23.57 | 0.71 ± 0.05 abc 7.60 | 2.52 ± 0.71 abc 28.34 | 23.92 ± 1.32 abe 5.52 | 30.92 ± 2.29 abde 7.41 | 8.85 ± 1.41 abc 15.88 | 27.00 ± 2.63 be 11.70 | 9.62 ± 1.40 bc 14.58 | 8.90 ± 1.29 abce 14.45 | 5.18 ± 0.73 abcde 13.99 | 1.75 ± 0.36 c 20.86 |
| ‘Octava’ × ‘Peterburgskiy’ (line) Cv | 68.65 ± 30.37 abde 29.67 | 44.94 ± 16.34 abde 36.55 | 23.71 ± 5.94 bcd 25.05 | 0.64 ± 0.08 ac 12.61 | 1.93 ± 0.61 acd 31.85 | 18.52 ± 2.61 cde 14.09 | 20.9 6± 3.54 cde 16.87 | 8.91 ± 1.24 abc 13.91 | 23.09 ± 2.61 ace 11.30 | 9.00 ± 1.35 b 14.89 | 4.84 ± 0.82 cdef 16.90 | 4.16 ± 0.85 abce 20.53 | 1.19 ± 0.22 cd 18.33 |
| ‘Viola’ × ‘Peterburgskiy’ (line) Cv | 91.54 ± 27.20 abe 29.71 | 63.03 ± 21.78 abe 34.55 | 28.51 ± 10.62 abcd 37.24 | 0.69 ± 0.08 abc 12.14 | 2.34 ± 0.70 abcd 30.01 | 18.91 ± 2.80 cde 14.80 | 23.73 ± 4.10 bce 17.29 | 8.91 ± 1.53 abc 17.15 | 24.67 ± 3.07 b 12.44 | 8.30 ± 1.47 b 17.68 | 7.11 ± 1.51 abcdef 21.31 | 4.32 ± 0.88 abce 20.42 | 1.67 ± 0.29 cd 17.25 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sinyavina, N.G.; Kochetov, A.A.; Kocherina, N.V.; Egorova, K.V.; Kurina, A.B.; Panova, G.G.; Chesnokov, Y.V. Breeding Approaches for Controlled Conditions of Artificial Light Culture for Small Radish and Radish (Raphanus sativus L.). Horticulturae 2023, 9, 678. https://doi.org/10.3390/horticulturae9060678
Sinyavina NG, Kochetov AA, Kocherina NV, Egorova KV, Kurina AB, Panova GG, Chesnokov YV. Breeding Approaches for Controlled Conditions of Artificial Light Culture for Small Radish and Radish (Raphanus sativus L.). Horticulturae. 2023; 9(6):678. https://doi.org/10.3390/horticulturae9060678
Chicago/Turabian StyleSinyavina, Nadezhda G., Alexei A. Kochetov, Natalia V. Kocherina, Ksenia V. Egorova, Anastasia B. Kurina, Gayane G. Panova, and Yuriy V. Chesnokov. 2023. "Breeding Approaches for Controlled Conditions of Artificial Light Culture for Small Radish and Radish (Raphanus sativus L.)" Horticulturae 9, no. 6: 678. https://doi.org/10.3390/horticulturae9060678