
Evaluation of Growth and Photosynthetic Rate of Cucumber Seedlings Affected by Far-Red Light Using a Semi-Open Chamber and Imaging System

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Target Crops and Environmental Condition
2.2. Treatment of Far-Red Light
2.3. Plant Growth Analysis
2.4. Measurement of Whole Canopy Gas Exchange Rate
2.5. Imaging System Configuration
2.6. Data Analysis
3. Results
3.1. Growth of Cucumber Seedlings in Different Light Quality Treatments
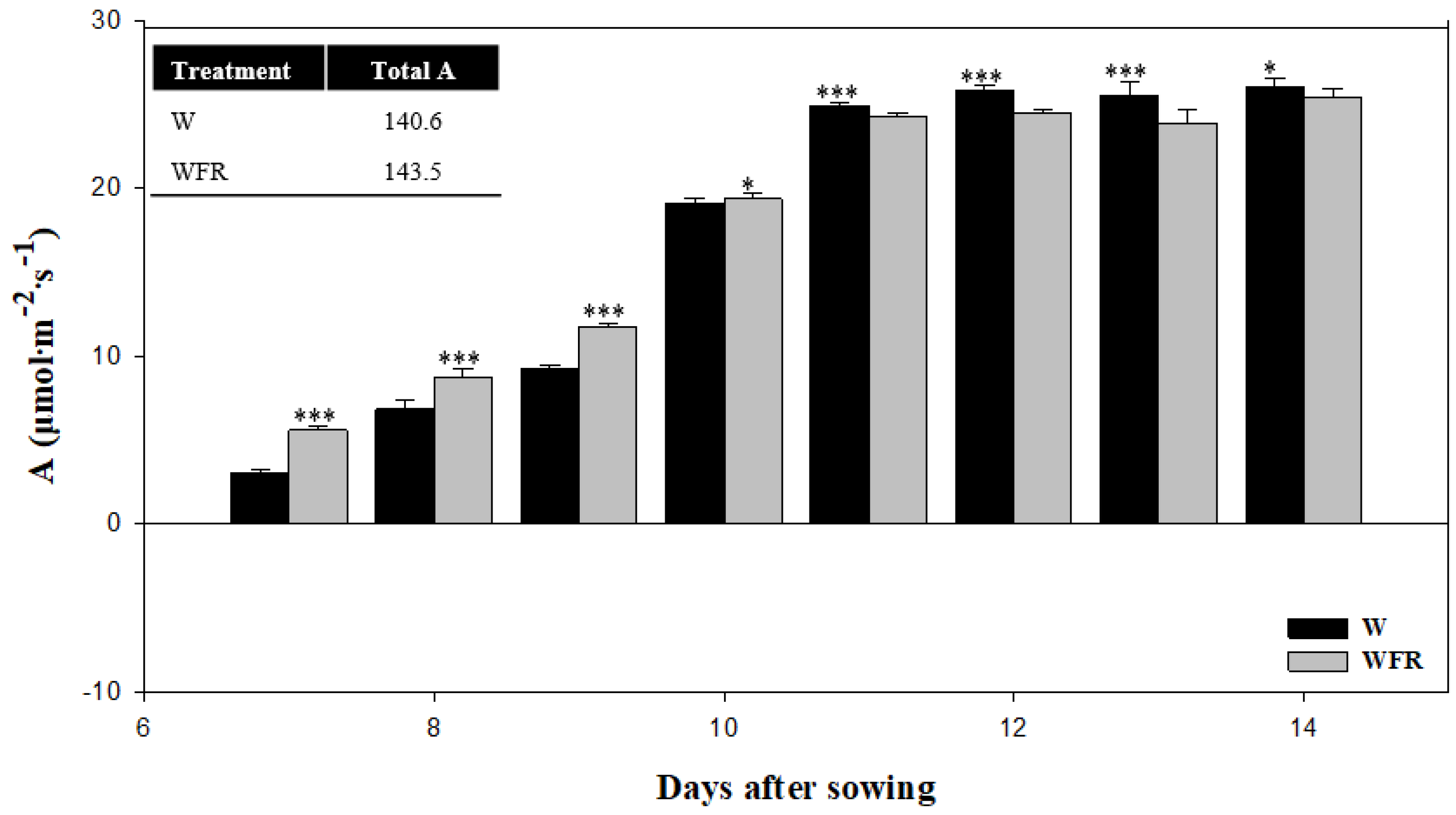
3.2. Photosynthetic Rate of Cucumber Whole Canopy in Different Light Quality Treatments
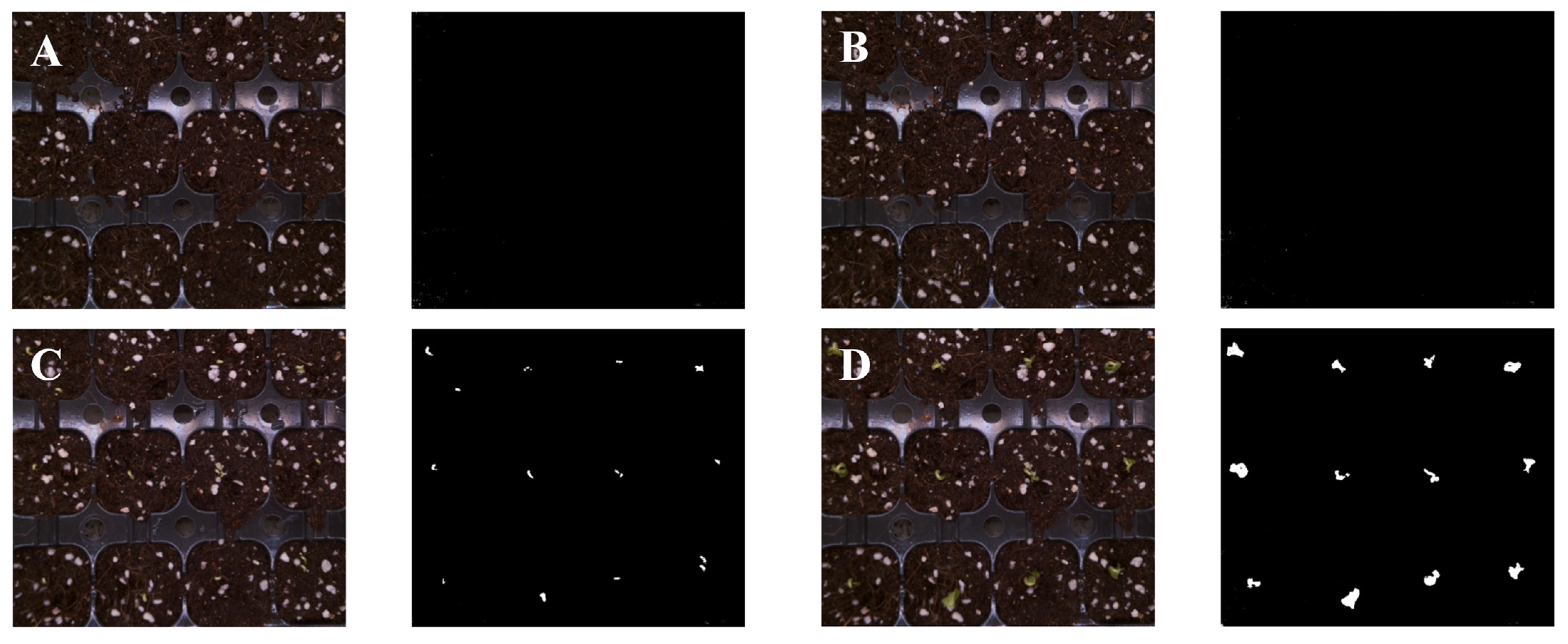
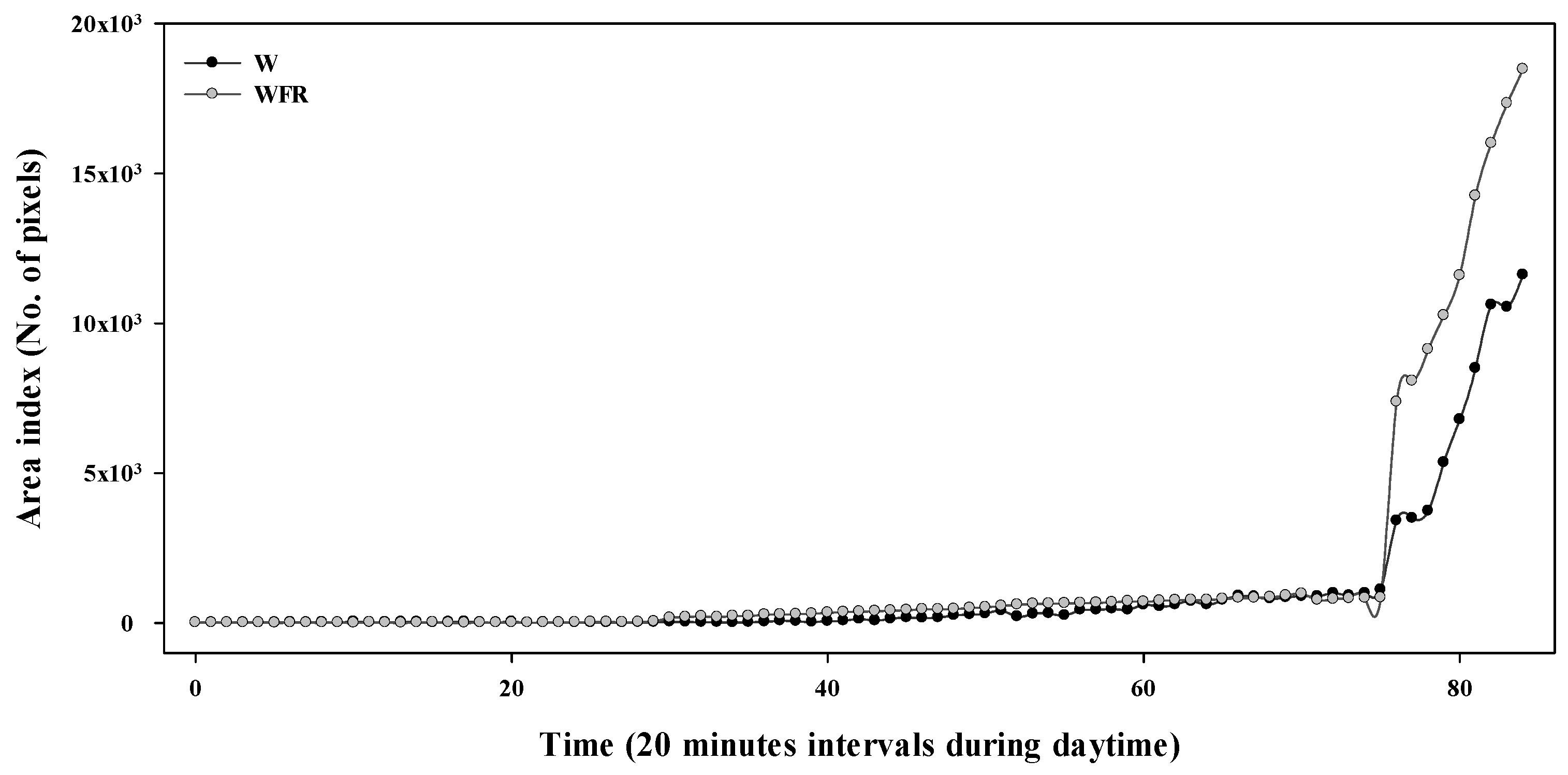
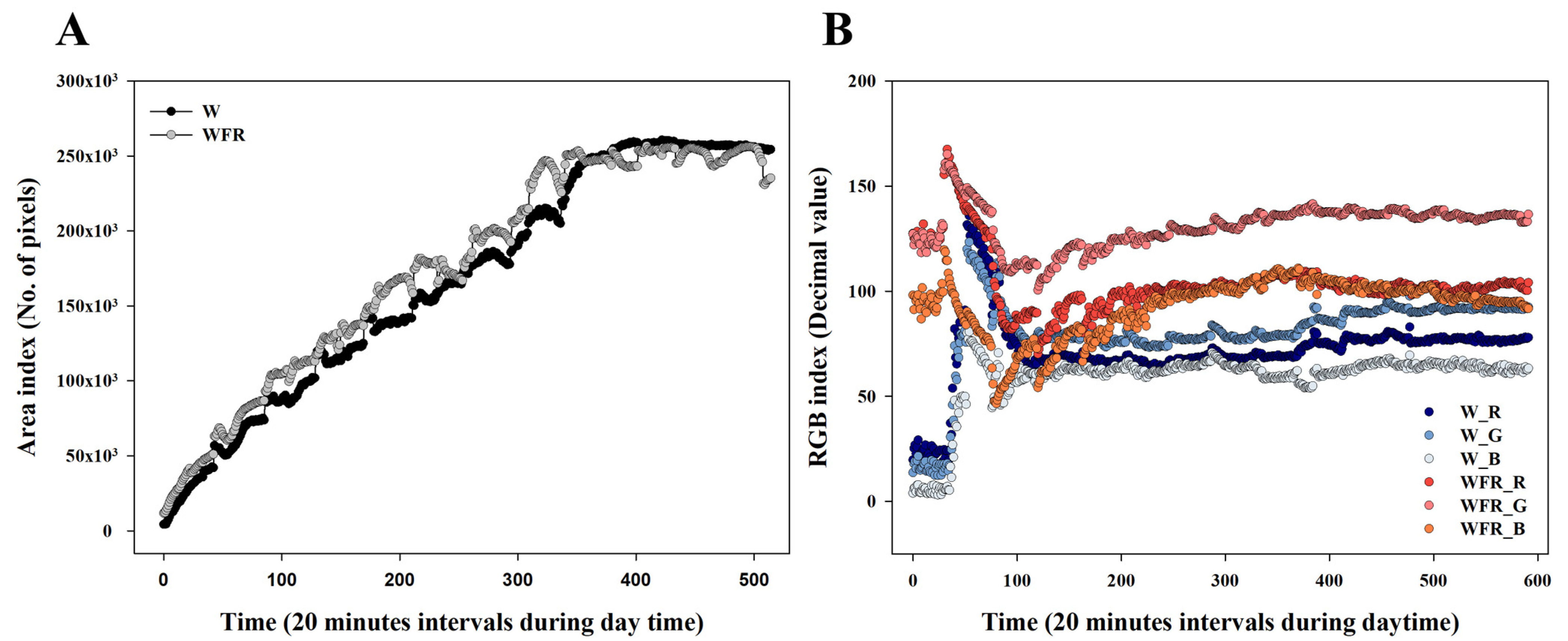
3.3. Growth of Cucumber Seedlings in Image
3.4. Pearson Correlation between Light Quality and Plant Growth
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kalaitzoglou, P.; van Ieperen, W.; Harbinson, J.; van der Meer, M.; Martinakos, S.; Weerheim, K.; Nicole, C.C.S.; Marcelis, L.F.M. Effects of Continuous or End-of-Day Far-Red Light on Tomato Plant Growth, Morphology, Light Absorption, and Fruit Production. Front. Plant Sci. 2019, 10, 322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhen, S.; Bugbee, B. Far-Red Photons Have Equivalent Efficiency to Traditional Photosynthetic Photons: Implications for Redefining Photosynthetically Active Radiation. Plant Cell Environ. 2020, 43, 1259–1272. [Google Scholar] [CrossRef] [PubMed]
- Jin, W.; Urbina, J.L.; Heuvelink, E.; Marcelis, L.F.M. Adding Far-Red to Red-Blue Light-Emitting Diode Light Promotes Yield of Lettuce at Different Planting Densities. Front. Plant Sci. 2021, 11, 2219. [Google Scholar] [CrossRef] [PubMed]
- Kasperbauer, M.J.; Peaslee, D.E. Morphology and Photosynthetic Efficiency of Tobacco Leaves That Received End-of-Day Red and Far Red Light during Development. Plant Physiol. 1973, 52, 440–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.T.; Zhang, Y.Q.; Yang, Q.C.; Li, T. Overhead Supplemental Far-Red Light Stimulates Tomato Growth under Intra-Canopy Lighting with LEDs. J. Integr. Agric. 2019, 18, 62–69. [Google Scholar] [CrossRef] [Green Version]
- Shibuya, T.; Endo, R.; Kitaya, Y.; Hayashi, S. Growth Analysis and Photosynthesis Measurements of Cucumber Seedlings Grown under Light with Different Red to Far-Red Ratios. HortScience 2016, 51, 843–846. [Google Scholar] [CrossRef] [Green Version]
- Bae, J.H.; Park, S.Y.; Oh, M.M. Supplemental Irradiation with Far-Red Light-Emitting Diodes Improves Growth and Phenolic Contents in Crepidiastrum denticulatum in a Plant Factory with Artificial Lighting. Hortic. Environ. Biotechnol. 2017, 58, 357–366. [Google Scholar] [CrossRef]
- Yang, F.; Liu, Q.; Cheng, Y.; Feng, L.; Wu, X.; Fan, Y.; Raza, M.A.; Wang, X.; Yong, T.; Liu, W.; et al. Low Red/Far-Red Ratio as a Signal Promotes Carbon Assimilation of Soybean Seedlings by Increasing the Photosynthetic Capacity. BMC Plant Biol. 2020, 20, 148. [Google Scholar] [CrossRef] [Green Version]
- Girardin, P.; Tollenaar, M. Effects of Intraspecific Interference on Maize Leaf Azimuth. Crop Sci. 1994, 34, 151–155. [Google Scholar] [CrossRef]
- Maddonni, G.A.; Otegui, M.E.; Andrieu, B.; Chelle, M.; Casal, J.J. Maize Leaves Turn Away from Neighbors. Plant Physiol. 2002, 130, 1181–1189. [Google Scholar] [CrossRef]
- Momokawa, N.; Kadono, Y.; Kudoh, H. Effects of Light Quality on Leaf Morphogenesis of a Heterophyllous Amphibious Plant, Rotala hippuris. Ann. Bot. 2011, 108, 1299–1306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.J.; Hahn, E.J.; Heo, J.W.; Paek, K.Y. Effects of LEDs on Net Photosynthetic Rate, Growth and Leaf Stomata of Chrysanthemum Plantlets In Vitro. Sci. Hortic. 2004, 101, 143–151. [Google Scholar] [CrossRef]
- Li, Q.; Kubota, C. Effects of Supplemental Light Quality on Growth and Phytochemicals of Baby Leaf Lettuce. Environ. Exp. Bot. 2009, 67, 59–64. [Google Scholar] [CrossRef]
- Lee, M.J.; Park, S.Y.; Oh, M.M. Growth and Cell Division of Lettuce Plants under Various Ratios of Red to Far-Red Light-Emitting Diodes. Hortic. Environ. Biotechnol. 2015, 56, 186–194. [Google Scholar] [CrossRef]
- Zhen, S.; van Iersel, M.W. Far-Red Light Is Needed for Efficient Photochemistry and Photosynthesis. J. Plant Physiol. 2017, 209, 115–122. [Google Scholar] [CrossRef] [Green Version]
- Fan, D.Y.; Hope, A.B.; Smith, P.J.; Jia, H.; Pace, R.J.; Anderson, J.M.; Chow, W.S. The Stoichiometry of the Two Photosystems in Higher Plants Revisited. Biochim. Biophys. Acta BBA Bioenerg. 2007, 1767, 1064–1072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glick, R.E.; McCauley, S.W.; Melis, A. Effect of Light Quality on Chloroplast-Membrane Organization and Function in Pea. Planta 1985, 164, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Wientjes, E.; Philippi, J.; Borst, J.W.; van Amerongen, H. Imaging the Photosystem I/Photosystem II Chlorophyll Ratio inside the Leaf. Biochim. Biophys. Acta BBA Bioenerg. 2017, 1858, 259–265. [Google Scholar] [CrossRef]
- Laisk, A.; Talts, E.; Oja, V.; Eichelmann, H.; Peterson, R.B. Fast Cyclic Electron Transport around Photosystem I in Leaves under Far-Red Light: A Proton-Uncoupled Pathway? Photosynth. Res. 2010, 103, 79–95. [Google Scholar] [CrossRef]
- Chow, W.S.; Hope, A.B. Electron Fluxes through Photosystem I in Cucumber Leaf Discs Probed by Far-Red Light. Photosynth. Res. 2004, 81, 77–89. [Google Scholar] [CrossRef]
- Zahedi, S.M.; Sarikhani, H. The Effect of End of Day Far-Red Light on Regulating Flowering of Short-Day Strawberry (Fragaria × ananassa Duch. cv. Paros) in a Long-Day Situation. Russ. J. Plant Physiol. 2017, 64, 83–90. [Google Scholar] [CrossRef]
- Jorissen, H.J.M.M.; Quest, B.; Lindner, I.; Marsac, N.T.D.; Gärtner, W. Phytochromes with Noncovalently Bound Chromophores: The Ability of Apophytochromes to Direct Tetrapyrrole Photoisomerization. Photochem. Photobiol. 2002, 75, 554–559. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Deng, X.W. Dissecting the Phytochrome A-Dependent Signaling Network in Higher Plants. Trends Plant Sci. 2003, 8, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Wu, S.; Zhai, H.; Zhou, P.; Song, M.; Su, L.; Xi, Y.; Li, Z.; Cai, Y.; Meng, F.; et al. Arabidopsis Phytochrome B Promotes SPA1 Nuclear Accumulation to Repress Photomorphogenesis under Far-Red Light. Plant Cell 2013, 25, 115–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheerin, D.J.; Menon, C.; Oven-Krockhaus, S.Z.; Enderle, B.; Zhu, L.; Johnen, P.; Schleifenbaum, F.; Stierhof, Y.D.; Huq, E.; Hiltbrunner, A. Light-Activated Phytochrome A and B Interact with Members of the SPA Family to Promote Photomorphogenesis in Arabidopsis by Reorganizing the COP1/SPA Complex. Plant Cell 2015, 27, 189–201. [Google Scholar] [CrossRef] [Green Version]
- Seo, H.S.; Watanabe, E.; Tokutomi, S.; Nagatani, A.; Chua, N.H. Photoreceptor Ubiquitination by COP1 E3 Ligase Desensitizes Phytochrome A Signaling. Genes Dev. 2004, 18, 617–622. [Google Scholar] [CrossRef] [Green Version]
- Xie, Y.; Liu, Y.; Ma, M.; Zhou, Q.; Zhao, Y.; Zhao, B.; Wang, B.; Wei, H.; Wang, H. Arabidopsis FHY3 and FAR1 Integrate Light and Strigolactone Signaling to Regulate Branching. Nat. Commun. 2020, 11, 1995. [Google Scholar] [CrossRef] [Green Version]
- Keuskamp, D.H.; Pollmann, S.; Voesenek, L.A.C.J.; Peeters, A.J.M.; Pierik, R. Auxin Transport through PIN-FORMED 3 (PIN3) Controls Shade Avoidance and Fitness during Competition. Proc. Natl. Acad. Sci. USA 2010, 107, 22740–22744. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Li, G. Auxin-Dependent Cell Elongation during the Shade Avoidance Response. Front. Plant Sci. 2019, 10, 914. [Google Scholar] [CrossRef] [Green Version]
- Sheerin, D.J.; Hiltbrunner, A. Molecular Mechanisms and Ecological Function of Far-Red Light Signalling. Plant Cell Environ. 2017, 40, 2509–2529. [Google Scholar] [CrossRef]
- Samiei, S.; Rasti, P.; Ly Vu, J.; Buitink, J.; Rousseau, D. Deep Learning-Based Detection of Seedling Development. Plant Methods 2020, 16, 103. [Google Scholar] [CrossRef] [PubMed]
- Hoecker, U.; Xu, Y.; Quail, P.H. SPA1: A New Genetic Locus Involved in Phytochrome A–Specific Signal Transduction. Plant Cell 1998, 10, 19–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saijo, Y.; Sullivan, J.A.; Wang, H.; Yang, J.; Shen, Y.; Rubio, V.; Ma, L.; Hoecker, U.; Deng, X.W. The COP1–SPA1 Interaction Defines a Critical Step in Phytochrome A-Mediated Regulation of HY5 Activity. Genes Dev. 2003, 17, 2642–2647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ai, K.; Su, H.; Zhou, H.; Cao, K.; Zou, Z. Effects of Different R:Fr Ratio on Chlorophyll Biosynthesis in Tomato Leaves under Salt Stress. North Hortic. 2019, 424, 14–22. [Google Scholar]
- Ji, Y.; Nuñez Ocaña, D.; Choe, D.; Larsen, D.H.; Marcelis, L.F.M.; Heuvelink, E. Far-Red Radiation Stimulates Dry Mass Partitioning to Fruits by Increasing Fruit Sink Strength in Tomato. New Phytol. 2020, 228, 1914–1925. [Google Scholar] [CrossRef]
- Shibuya, T.; Endo, R.; Yuba, T.; Kitaya, Y. The Photosynthetic Parameters of Cucumber as Affected by Irradiances with Different Red:Far-Red Ratios. Biol. Plant. 2014, 59, 198–200. [Google Scholar] [CrossRef]
- Woolley, D.W.; Shaw, E.; Med, B.J.; Pharmacol Exptl Therap, J.; Robert Emerson, B.; Chalmers, R.; Cederstrand, C. Some factors influencing the long-wave limit of photosynthesis. Proc. Natl. Acad. Sci. USA 1957, 43, 133–143. [Google Scholar] [CrossRef] [Green Version]
- Tan, T.; Li, S.; Fan, Y.; Wang, Z.; Ali Raza, M.; Shafiq, I.; Wang, B.; Wu, X.; Yong, T.; Wang, X.; et al. Far-Red Light: A Regulator of Plant Morphology and Photosynthetic Capacity. Crop J. 2022, 10, 300–309. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Light Sources | Blue (400–500 nm) | Green (500–600 nm) | Red (600–700 nm) | Far-Red (700–780 nm) | PPFD (400–700 nm) | PFD (380–780 nm) | Red/Far-Red | DLI (mol) |
|---|---|---|---|---|---|---|---|---|
| W | 57.3b | 199.4b | 172.0a | 58.2b | 428.7b | 487.3a | 3.0a | 28.0a |
| (%) z | 11.8 | 40.9 | 35.3 | 11.9 | 88.0 | 100 | ||
| WFR | 47.1c | 162.9c | 145.5b | 126.9a | 355.5c | 482.8a | 1.15a | 27.8a |
| (%) | 9.8 | 33.7% | 30.1 | 26.3 | 73.6 | 100 | ||
| Sunlight | 160.9a | 216.5a | 179.8a | 111a | 557.2a | 680.0b | 1.6b | 25.9b |
| (%) | 23.7 | 31.8 | 26.4 | 16.3 | 81.9 | 100 |
| Treatment | Plant Height (cm) | Stem Diameter (mm) | Leaf Length (cm) | Leaf Width (cm) | Leaf Area (cm2) |
|---|---|---|---|---|---|
| W | 3.28 ± 0.14 | 4.11 ± 0.28 | 5.93 ± 0.15 | 8.08 ± 0.28 | 72.15 ± 5.33 |
| WFR | 7.06 ± 0.39 | 4.17 ± 0.26 | 6.40 ± 0.14 | 9.25 ± 0.33 | 83.95 ± 6.95 |
| Significance | *** | NS | * | *** | ** |
| Treatment | No. of Leaves | Shoot Fresh Weight (g) | Shoot Dry Weight (g) | Root Fresh Weight (g) | Root Dry Weight (g) | S/R Ratio |
|---|---|---|---|---|---|---|
| W | 2.00 ± 0.00 | 3.05 ± 0.27 | 0.55 ± 0.08 | 1.83 ± 0.59 | 0.06 ± 0.02 | 1.8 ± 0.7 |
| WFR | 2.00 ± 0.00 | 3.55 ± 0.35 | 0.56 ± 0.10 | 1.87 ± 0.30 | 0.08 ± 0.00 | 2.0 ± 0.4 |
| Significance | NS | ** | NS | NS | NS | NS |
| Treatment | Compactness | LAI | LAR | LUE |
|---|---|---|---|---|
| W | 0.17 ± 0.04 | 0.18 ± 0.01 | 132.2 ± 15.6 | 20.4 ± 3.0 |
| WFR | 0.08 ± 0.01 | 0.21 ± 0.02 | 152.7 ± 22.9 | 20.9 ± 3.7 |
| Significance | *** | ** | NS | NS |
| Pearson Correlation with Far-Red Light | |||||||
|---|---|---|---|---|---|---|---|
| Plant Height | Stem Diameter | Leaf Length | Leaf Width | Leaf Area | Shoot Fresh Weight | Shoot Dry Weight | |
| coefficient | 0.92 *** | 0.12 * | 0.53 ** | 0.90 *** | 0.71 ** | 0.64 ** | 0.05 NS |
| Root fresh weight | Root dry weight | S/R ratio | Compactness | LAI | LAR | LUE | |
| coefficient | 0.04 NS | 0.45 ** | 0.10 * | −0.21 * | 0.71 *** | 0.49 ** | 0.07 NS |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moon, Y.H.; Yang, M.; Woo, U.J.; Sim, H.S.; Lee, T.Y.; Shin, H.R.; Jo, J.S.; Kim, S.K. Evaluation of Growth and Photosynthetic Rate of Cucumber Seedlings Affected by Far-Red Light Using a Semi-Open Chamber and Imaging System. Horticulturae 2023, 9, 98. https://doi.org/10.3390/horticulturae9010098
Moon YH, Yang M, Woo UJ, Sim HS, Lee TY, Shin HR, Jo JS, Kim SK. Evaluation of Growth and Photosynthetic Rate of Cucumber Seedlings Affected by Far-Red Light Using a Semi-Open Chamber and Imaging System. Horticulturae. 2023; 9(1):98. https://doi.org/10.3390/horticulturae9010098
Chicago/Turabian StyleMoon, Yu Hyun, Myongkyoon Yang, Ui Jeong Woo, Ha Seon Sim, Tae Yeon Lee, Ha Rang Shin, Jung Su Jo, and Sung Kyeom Kim. 2023. "Evaluation of Growth and Photosynthetic Rate of Cucumber Seedlings Affected by Far-Red Light Using a Semi-Open Chamber and Imaging System" Horticulturae 9, no. 1: 98. https://doi.org/10.3390/horticulturae9010098