
Determination of Grafting Success and Carbohydrate Distributions of Foxy Grape (Vitis labrusca L.) Varieties Grafted on Different American Grape Rootstocks


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Characteristics of Cultivars and Rootstocks
2.1.1. Cultivars
‘Rizessi’
‘Çeliksu’
‘Ülkemiz’
‘Rizellim’
‘Rizpem’
2.1.2. Rootstocks
140Ru
SO4
110R
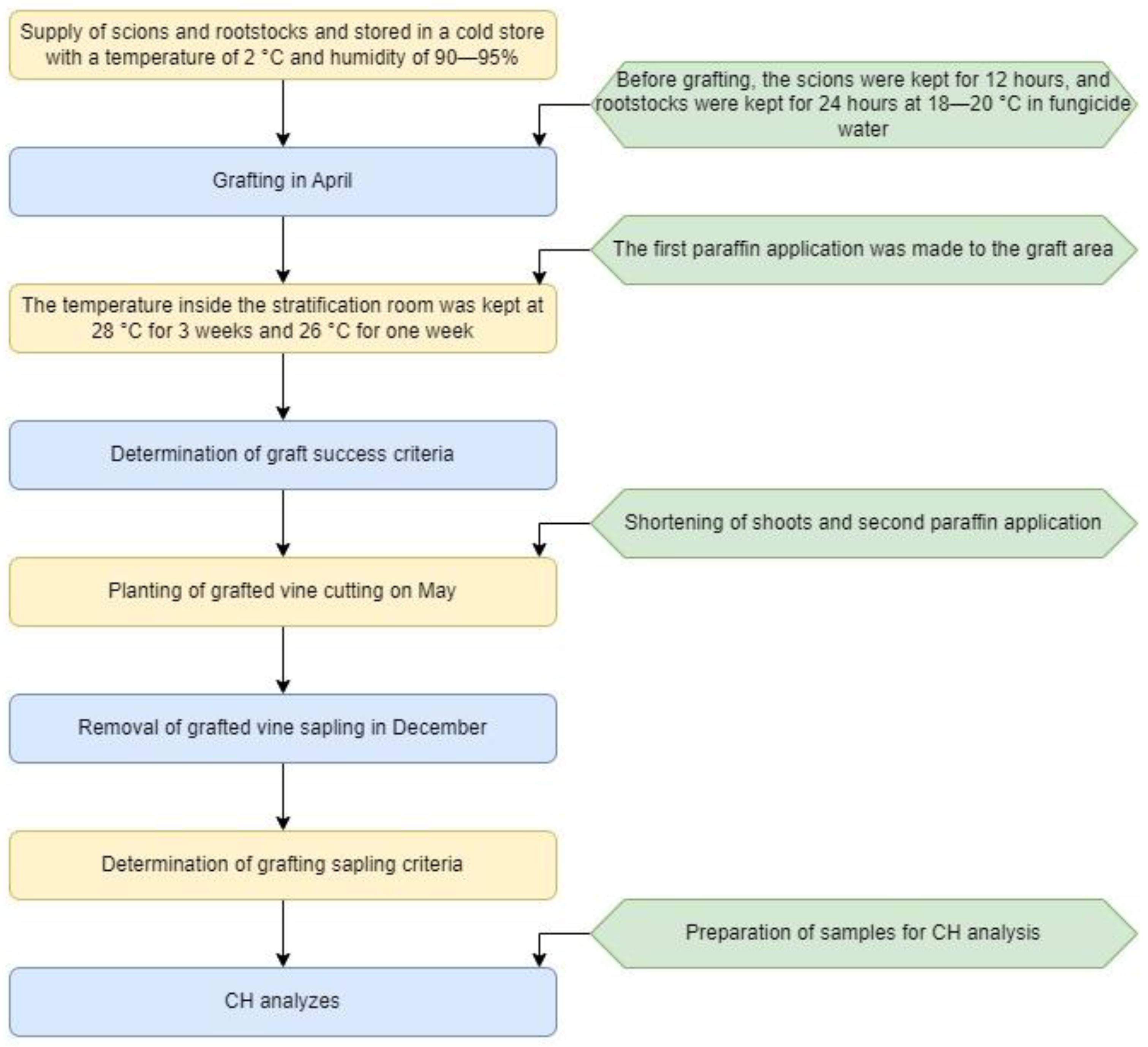
2.2. Production of Grafted Vine Saplings
2.3. Graft Success Criteria
2.4. Grafted Vine Sapling Criteria
2.5. Carbohydrate Analyses
2.6. Statistical Analysis
3. Results
3.1. Graft Success
3.2. Grafted Vine Sapling
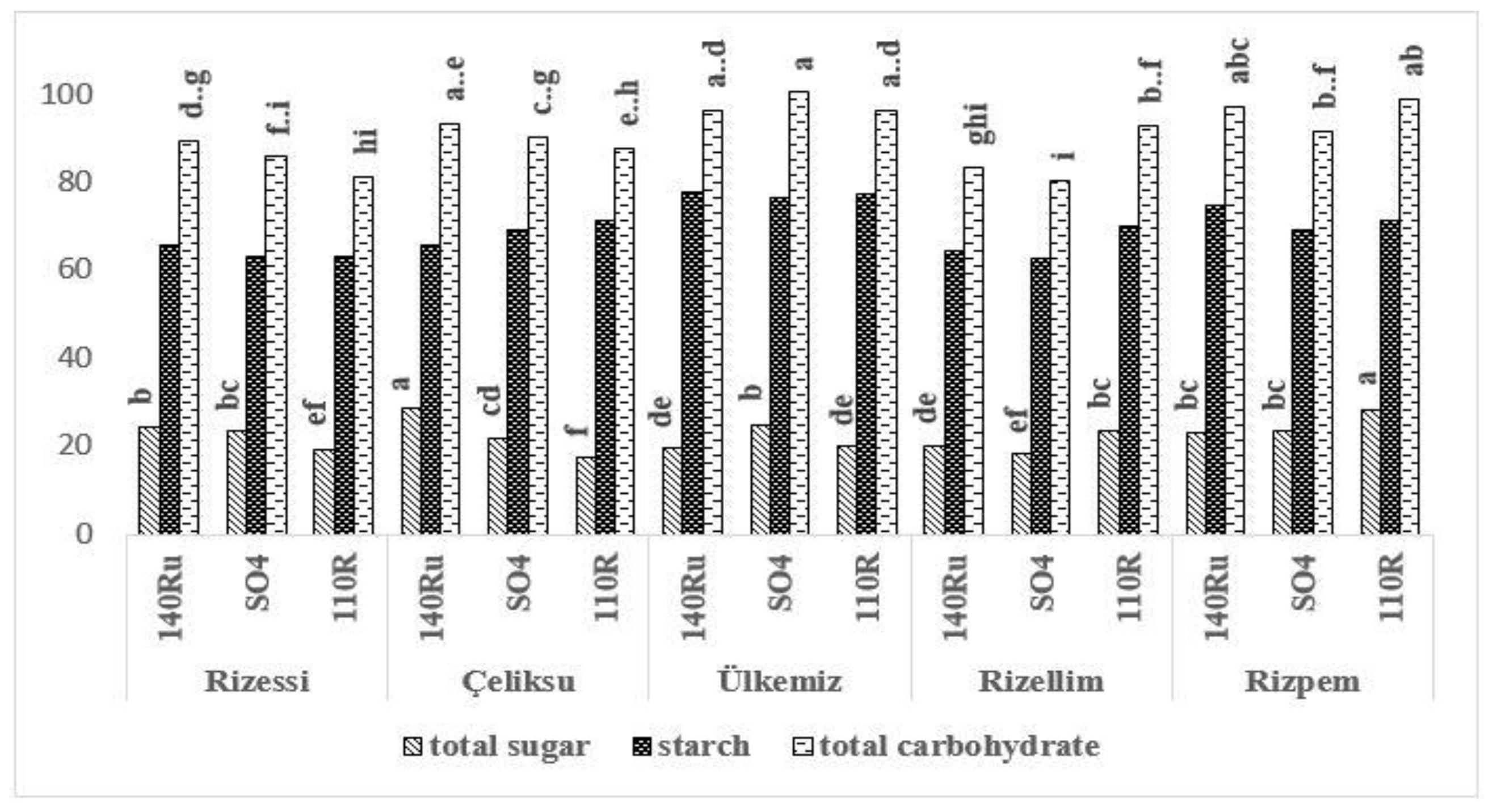
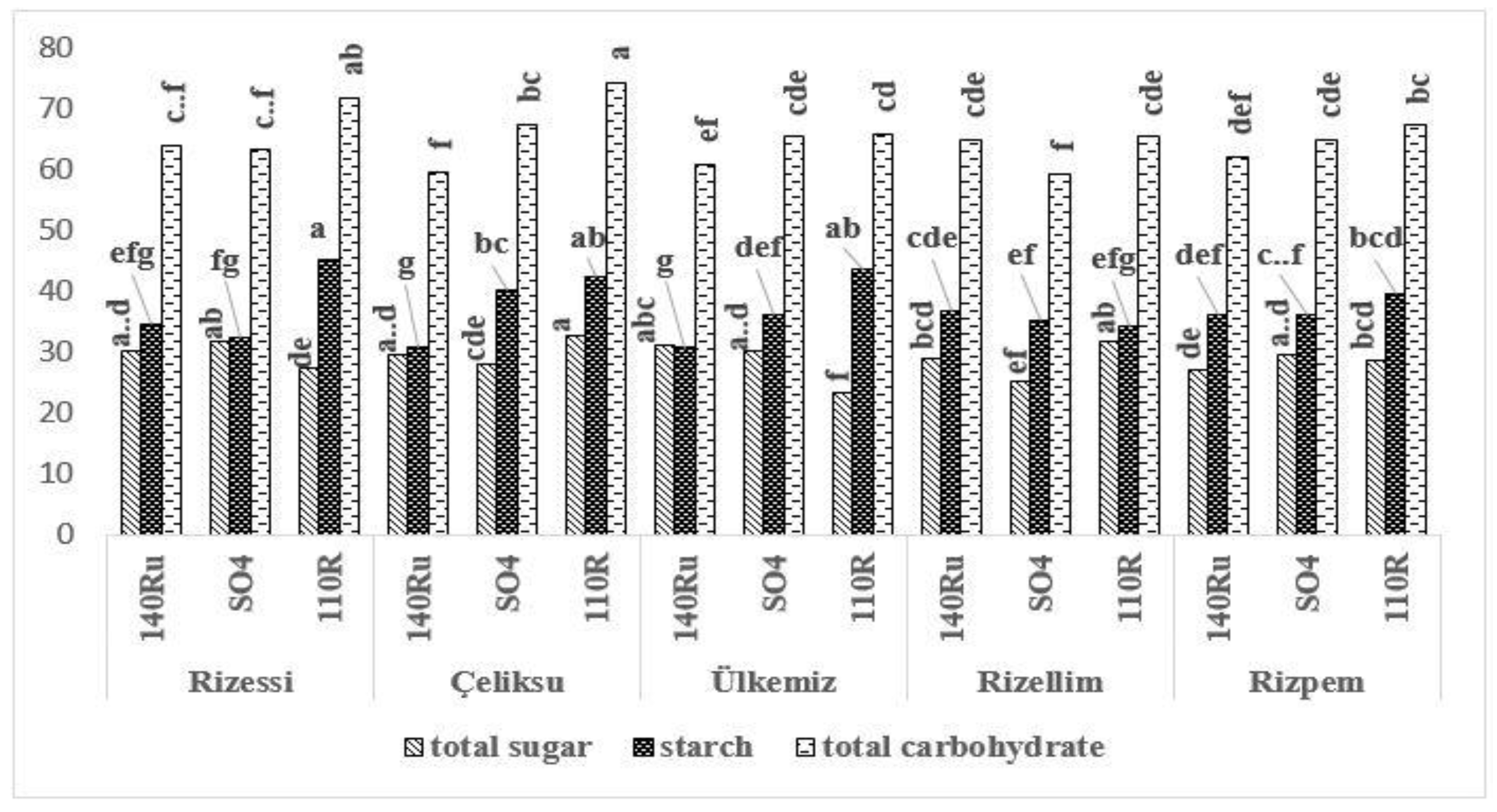
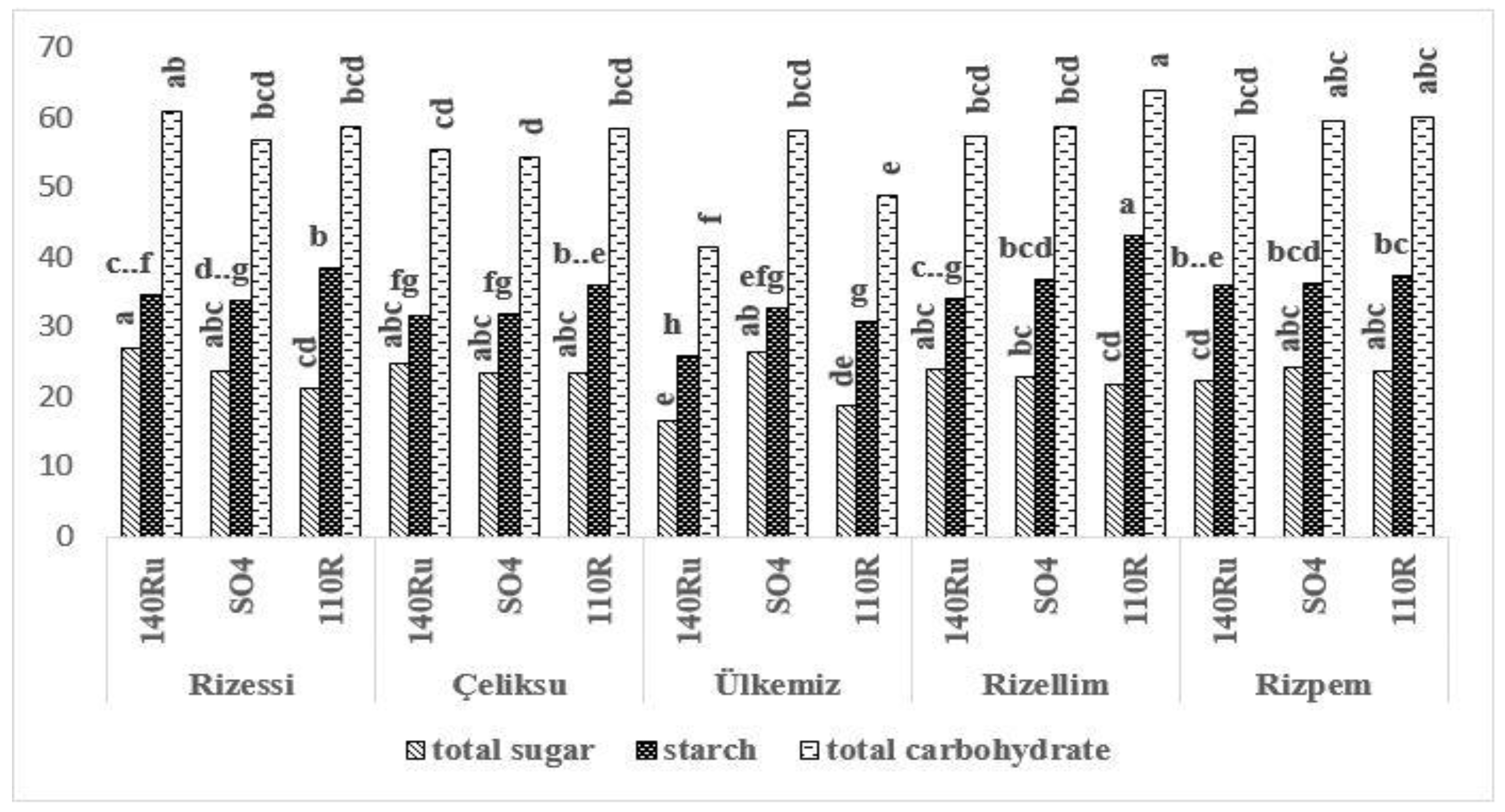
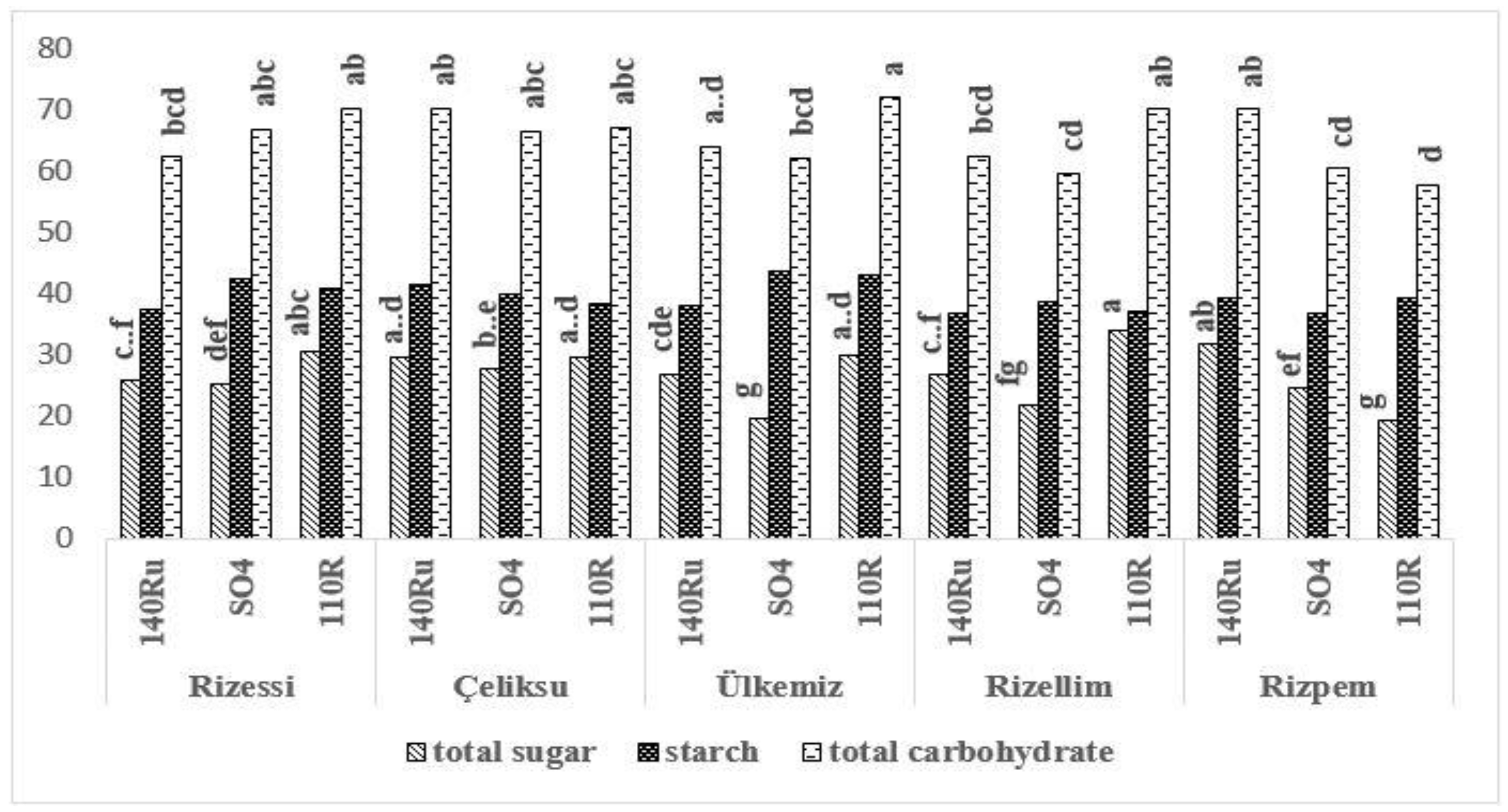
3.3. Carbohydrate Accumulations
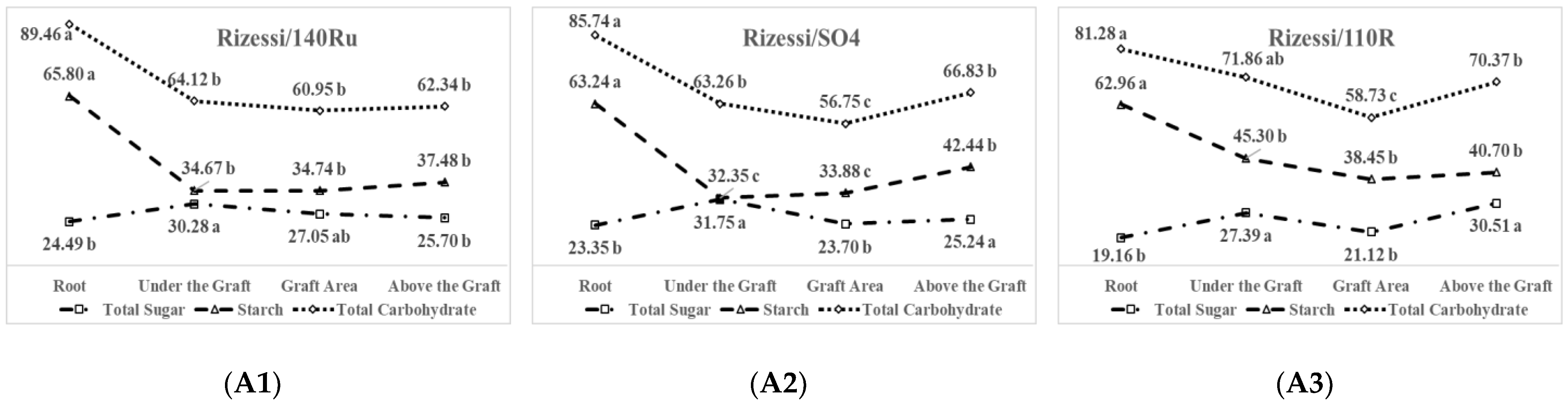
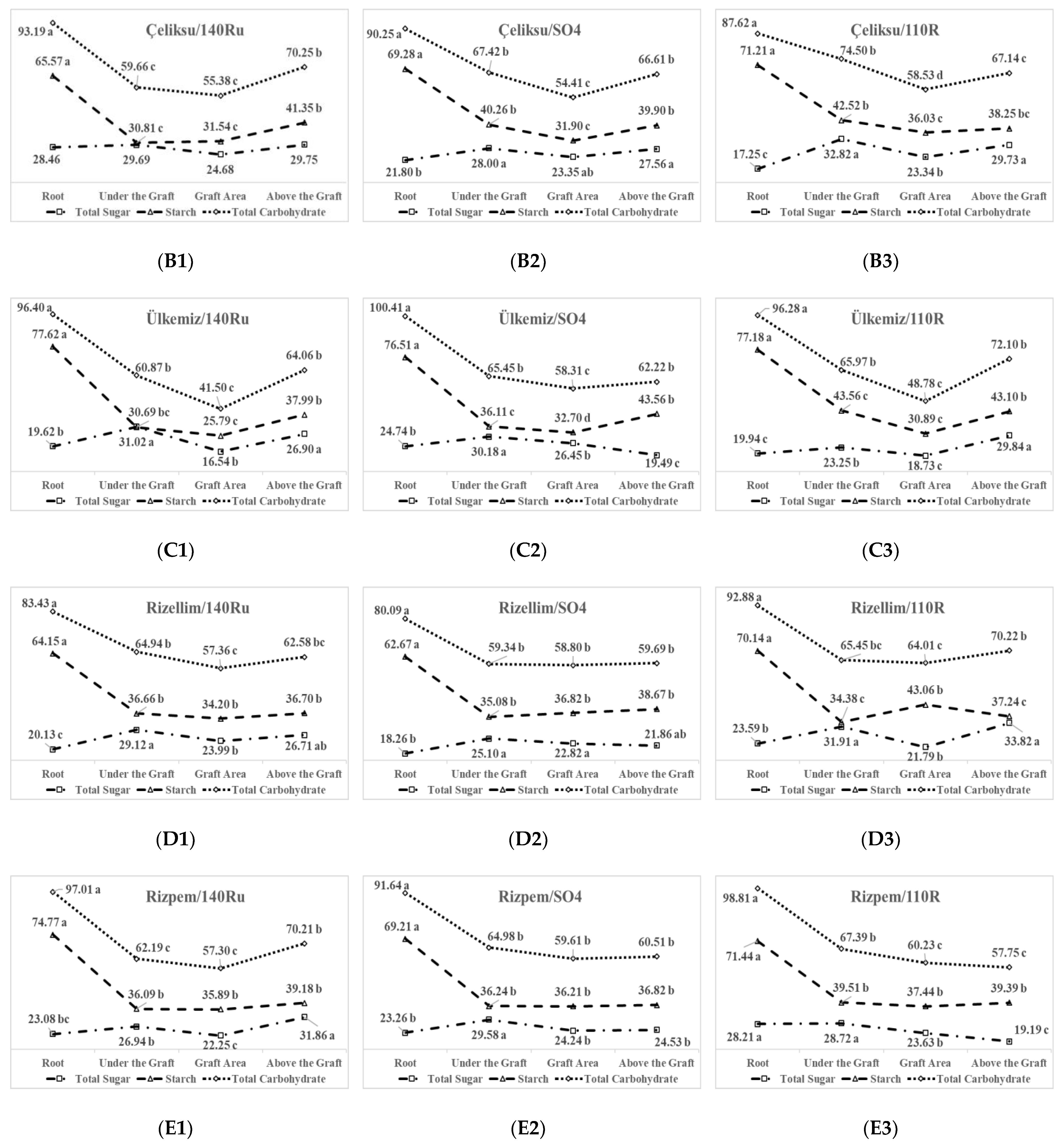
3.4. Carbohydrate Distributions
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kızılaslan, N.; Somak, E. Tokat ili Erbaa İlçesinde bağcılık işletmelerinde tarımsal ilaç kullanımında üreticilerin bilinç düzeyi. Gaziosmanpaşa Bilimsel Araştırma Derg 2013, 4, 79–93. [Google Scholar]
- Tekdal, D.; Sarlar, S. Yerel asma genetik kaynakları ve önemi. Bağbahçe Bilim Dergisi 2016, 3, 20–26. [Google Scholar]
- Köse, B.; Çelik, H. Phenological changes of shoot carbohydrates and plant growth characteristics in Vitis Labrusca L. grape. Sci. Pap.-Ser. B Hortic. 2017, LXI, 257–268. [Google Scholar]
- Cangi, R.; Celik, H.; Odabas, F.; Islam, A. Determination of ampelographic characters of some natural foxy grape (Vitis labrusca L.) types grown in Northern Turkey (in Trabzon province). Asian J. Plant Sci. 2006, 5, 373–377. [Google Scholar]
- Çelik, H.; Köse, B.; Cangi, R. Determination of Fox grape genotypes (Vitis labrusca L.) grown in Northeastern Anatolia. Hort. Sci 2008, 35, 162–170. [Google Scholar]
- Çelik, H.; Köse, B.; Ateş, S. Karadeniz bölgesinden selekte edilerek tescillenen yeni kokulu üzüm (Vitis labrusca L.) çeşitleri. Bahçe 2018, 47, 299–309. [Google Scholar]
- Gargin, S.; Altindisli, A. A Research on the affinity coefficients of Red Globe grape variety with 140 R, 41 B rootstocks. In Proceedings of the BIO Web of Conferences, Online, 4 November 2014; p. 01004. [Google Scholar]
- Köse, B.; Karabulut, B.; Ceylan, K. Effect of rootstock on grafted grapevine quality. Eur. J. Hortic. Sci. 2014, 79, 197–202. [Google Scholar]
- Rizk-Alla, M.; Sabry, G.; Abd-El-Wahab, M. Influence of some rootstocks on the performance of red globe grape cultivar. J. Am. Sci. 2011, 7, 71–81. [Google Scholar]
- Verma, S.; Singh, S.; Patel, V.; Singh, K. Grafting and stionic effect studies in grape. J. Rural. Agriculural Res. 2010, 10, 1–3. [Google Scholar]
- Troncoso de Arce, A.; Matte, C.; Cantos, M.; Lavee, S. Evaluation of salt tolerance of in vitro-grown grapevine rootstock varieties. Vitis 1999, 38, 55–60. [Google Scholar]
- Loreti, F.; Massai, R. State of the art on peach rootstocks and orchard systems. In Proceedings of the VI International Peach Symposium 713, Santiago, Chile, 30 July 2006; pp. 253–268. [Google Scholar]
- Reynolds, A.G.; Wardle, D.A. Rootstocks impact vine performance and fruit composition of grapes in British Columbia. Hort Technol. 2001, 11, 419–427. [Google Scholar] [CrossRef] [Green Version]
- Çelik, H.; Odabaş, F. Bağcılıkta aşıların kontrollü şartlarda kaynaştırılması ve tüplü asma fidanı üretimi. OMÜ Ziraat Fakültesi Derg. 1995, 10, 169–178. [Google Scholar]
- Hartmann, H.T.; Kester, D.E. Plant Propagation: Principles and Practices; Prentice-Hall: Hoboken, NJ, USA, 1975. [Google Scholar]
- Çelik, H. Üzüm Yetiştiriciliği; Pazar Ziraat Odası Egitim Yay, Pazar Ofset: Rize, Turkey, 2004; p. 121. [Google Scholar]
- Çelik, S. Bağcılık (Ampeloloji); Trakya Üniversitesi, Tekirdağ Ziraat Fakültesi Bahçe Bitkileri Bölümü: Tekirdağ, Türkiye, 1998; Volume 1, 425p. [Google Scholar]
- Wang, Y.; Kollmann, R. Vascular differentiation in the graft union of in-vitro grafts with different compatibility. Structural and functional aspects. J. Plant Physiol. 1996, 147, 521–533. [Google Scholar] [CrossRef]
- Pina, A.; Errea, P. A review of new advances in mechanism of graft compatibility–incompatibility. Sci. Hortic. 2005, 106, 1–11. [Google Scholar] [CrossRef]
- Cangi, R.; Balta, F.; Doğan, A. Aşılı asma fidanı üretiminde kullanılan katlama ortamlarının fidan randıman ve kalitesi üzerine etkilerinin anatomik ve histolojik olarak incelenmesi. Turk. J. Agric. For 2000, 24, 393–398. [Google Scholar]
- Assunção, M.; Santos, C.; Brazão, J.; Eiras-Dias, J.; Fevereiro, P. Understanding the molecular mechanisms underlying graft success in grapevine. BMC Plant Biol. 2019, 19, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Tedesco, S.; Pina, A.; Fevereiro, P.; Kragler, F. A phenotypic search on graft compatibility in grapevine. Agronomy 2020, 10, 706. [Google Scholar] [CrossRef]
- Estrada-Luna, A.; Lopez-Peralta, C.; Cardenas-Soriano, E. In vitro micrografting and the histology of graft union formation of selected species of prickly pear cactus (Opuntia spp.). Sci. Hortic. 2002, 92, 317–327. [Google Scholar] [CrossRef]
- Pina, A.; Errea, P.; Schulz, A.; Martens, H.J. Cell-to-cell transport through plasmodesmata in tree callus cultures. Tree Physiol. 2009, 29, 809–818. [Google Scholar] [CrossRef] [Green Version]
- Bahar, E. Hidroponik Yöntemlerle Aşılı Köklü Asma Fidanı Üretimi. Ph.D. Thesis, Trakya Üniversitesi (Tekirdağ) Fen Bilimleri Enstitüsü, Terkidağ, Türkiye, 1996. [Google Scholar]
- Vršič, S.; Pulko, B.; Kocsis, L. Factors influencing grafting success and compatibility of grape rootstocks. Sci. Hortic. 2015, 181, 168–173. [Google Scholar] [CrossRef]
- Gökbayrak, Z.; Söylemezoğlu, G.; Akkurt, M.; Celik, H. Determination of grafting compatibility of grapevine with electrophoretic methods. Sci. Hortic. 2007, 113, 343–352. [Google Scholar] [CrossRef]
- Evert, R.F.; Eichhorn, S.E. Raven Biology of Plants, 8th ed.; Peter Marshall: New York, NY, USA, 2013. [Google Scholar]
- Gibon, Y.; Eva-Theresa, P.Y.L.; Sulpice, R.; Lunn, J.E.; Hoehne, M.; Günther, M.; Stitt, M. Adjustment of growth, starch turnover, protein content and central metabolism to a decrease of the carbon supply when Arabidopsis is grown in very short photoperiods. Plant Cell Environ. 2009, 32, 859–874. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.M.; Stitt, M. Coordination of carbon supply and plant growth. Plant Cell Environ. 2007, 30, 1126–1149. [Google Scholar] [CrossRef] [PubMed]
- Sulpice, R.; Pyl, E.-T.; Ishihara, H.; Trenkamp, S.; Steinfath, M.; Witucka-Wall, H.; Gibon, Y.; Usadel, B.; Poree, F.; Piques, M.C. Starch as a major integrator in the regulation of plant growth. Proc. Natl. Acad. Sci. USA 2009, 106, 10348–10353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Körner, C. Carbon limitation in trees. J. Ecol. 2003, 91, 4–17. [Google Scholar] [CrossRef] [Green Version]
- Loescher, W.H.; McCamant, T.; Keller, J.D. Carbohydrate reserves, translocation, and storage in woody plant roots. HortScience 1990, 25, 274–281. [Google Scholar] [CrossRef] [Green Version]
- Keller, M. The Science of Grapevines; Academic Press: Cambridge, MA, USA, 2015. [Google Scholar]
- Spicer, R. Symplasmic networks in secondary vascular tissues: Parenchyma distribution and activity supporting long-distance transport. J. Exp. Bot. 2014, 65, 1829–1848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salleo, S.; Lo Gullo, M.; Trifilo, P.; Nardini, A. New evidence for a role of vessel-associated cells and phloem in the rapid xylem refilling of cavitated stems of Laurus nobilis L. Plant Cell Environ. 2004, 27, 1065–1076. [Google Scholar] [CrossRef]
- O’Brien, M.J.; Leuzinger, S.; Philipson, C.D.; Tay, J.; Hector, A. Drought survival of tropical tree seedlings enhanced by non-structural carbohydrate levels. Nat. Clim. Chang. 2014, 4, 710–714. [Google Scholar] [CrossRef]
- Hoch, G. Carbon reserves as indicators for carbon limitation in trees. In Progress in Botany; Springer: Bielefeld, Germany, 2015; pp. 321–346. [Google Scholar]
- Plavcová, L.; Jansen, S. The role of xylem parenchyma in the storage and utilization of nonstructural carbohydrates. In Functional and Ecological Xylem Anatomy; Springer: Cham, Switzerland, 2015; pp. 209–234. [Google Scholar]
- Hardy, P. Selective diffusion of basic and acidic products of CO2 fixation into the transpiration stream in grapevine. J. Exp. Bot. 1969, 20, 856–862. [Google Scholar] [CrossRef]
- Hardy, P.; Possingham, J. Studies on translocation of metabolites in the xylem of grapevine shoots. J. Exp. Bot. 1969, 20, 325–335. [Google Scholar] [CrossRef]
- Andersen, P.C.; Brodbeck, B.V. Diurnal and temporal changes in the chemical profile of xylem exudate from Vitis rotundifolia. Physiol. Plant. 1989, 75, 63–70. [Google Scholar] [CrossRef]
- Ağaoğlu, Y. Bilimsel ve uygulamalı bağcılık (Asma Biyolojisi); Kavaklıdere Eğitim Yayınları: Ankara, Türkiye, 1999; Volume 1, p. 205. [Google Scholar]
- Whiting, J.R. Selection of grapevine rootstocks and clones for greater Victoria; Department of Primary Industries: Tatura, Australia, 2003. [Google Scholar]
- Rules, L. Selecting a Rootstock for a Winegrape Vineyard. Available online: https://www.lodigrowers.com/selecting-a-rootstock-for-a-winegrape-vineyard/ (accessed on 20 February 2021).
- Shaffer, R.G.; Sampalo, T.; Pinkerton, J.; Vasconcelos, M. Grapevine rootstocks for Oregon vineyards. Corvallis Or. Ext. Serv. Or. State Univ. 2004, 1–10. [Google Scholar]
- Goldammer, T. Grape Grower’s Handbook. A Guide to Viticulture for Wine Production; Apex Publisher: Centreville, VG, USA, 2015; 728p. [Google Scholar]
- Cangi, R. Aşılı Asma Fidanı Üretimi ve aşı Kaynaşmasının Anatomik, Histolojik ve Biyokimyasal Olarak İncelenmesi. Ph.D. Thesis, Yüzüncü Yıl Üniversitesi, Van, Türkiye, 1996. [Google Scholar]
- Çelik, H. The effects of different grafting methods applied by manual grafting units on grafting success in grapevines. Turkish J. Agric. For. 2000, 24, 499–504. [Google Scholar]
- Çelik, H. Samsun ili Fidanlık Şartlarında Aşılama Yoluyla Aşırı asma Fidanı Üretiminde aşı Tipi ve Aşılama Zamanlarının Etkileri. Ph.D. Thesis, Ondokuz Mayıs Üniversitesi Fen Bilimleri Enstitüsü, Samsun, Türkiye, 1995. [Google Scholar]
- Köse, B.; Ateş, S.; Çelik, H. Farklı anaçlar üzerine aşılı kokulu kara üzüm (Vitis labrusca L.) ve Şiraz (Vitis vinifera L.) üzüm çeşitlerinin fidan randımanı ve gelişimi üzerine ağır bünyeli toprakların etkileri. Harran Tarım Ve Gıda Bilim. Derg. 2016, 20, 135–145. [Google Scholar] [CrossRef] [Green Version]
- Köse, B.; Çelik, H.; Karabulut, B. Determination of callusing performance and vine sapling characteristics on different rootstocks of ‘Merzifon Karası’ grape variety (Vitis vinifera L.). Anadolu Tarım Bilim. Derg. 2015, 30, 87–94. [Google Scholar] [CrossRef] [Green Version]
- Miller, D.; Howell, G.S.; Flore, J.A. Effect of shoot number on potted grapevines: I. Canopy development and morphology. Am. J. Enol. Vitic. 1996, 47, 244–250. [Google Scholar]
- Scott, T.A., Jr.; Melvin, E.H. Determination of dextran with anthrone. Anal. Chem. 1953, 25, 1656–1661. [Google Scholar] [CrossRef]
- Candolfi-Vasconcelos, M.; Koblet, W. Yield, fruit quality, bud fertility and starch reserves of the wood as a function of leaf removal in Vitis vinifera-evidence of compensation and stress recovering. Vitis 1990, 29, 199–221. [Google Scholar]
- Dardeniz, A.; Şahin, A.O. Aşılı asma fidanı üretiminde farklı çeşit ve anaç kombinasyonlarının vejetatİf gelişme ve fidan randımanı üzerine etkileri. Bahçe 2005, 34, 1–10. [Google Scholar]
- Tangolar, S.; Gök, S.; Ergenoglu, F.; Kamiloglu, Ö. Rooting characteristics of several grape cultivars and rootstocks. V Temp. Zone Fruit Trop. Subtrop. 1996, 441, 403–406. [Google Scholar] [CrossRef]
- Tunçel, R.; Dardeniz, A. Aşılı asma çeliklerinin fidanlıktaki vejetatif gelişimi ve randımanları üzerine katlamanın etkileri. Int. J. Agric. Nat. Sci. 2013, 6, 118–122. [Google Scholar]
- Ghule, V.; Zagade, P.; Bhor, V.; Somkuwar, R. Rootstock affects graft success, growth and physiological parameters of grape varieties (Vitis vinifera L.). Int. J. Curr. Microbiol. App. Sci 2019, 8, 799–805. [Google Scholar] [CrossRef]
- Hamdan, A.J.; Basheer-Salimia, R. Preliminary compatibility between some table-grapevine scion and phylloxera-resistant rootstock cultivars. Jordan J. Agric. Sci. 2010, 6, 1–10. [Google Scholar]
- Çelik, H.; Ağaoğlu, Y.; Fidan, Y.; Marasalı, B.; Söylemezoğlu, G. Genel Bağcılık; Sunfidan AŞ Mesleki Kitaplar Serisi 1; Fersa Matbaacılık: Ankara, Türkiye, 1998; 256p, ISBN 975-96656-0-3. [Google Scholar]
- Çakır, A.; Yücel, B. Narince ve Kalecik Karası üzüm çeşitlerinin 1103 Paulsen Amerikan asma anacı ile aşı performanslarının belirlenmesi. Turk. J. Agric. Nat. Sci. 2016, 3, 311–317. [Google Scholar]
- Bekişli, M.; Gürsöz, S.; Bilgiç, C. Aşılı Asma fidanı üretiminde bazı anaç-çeşit kombinasyonlarının katlama odası performanslarının incelenmesi. Harran Tarım Ve Gıda Bilim. Derg. 2015, 19, 24–37. [Google Scholar]
- Blasi, L.; Pommer, C. Evaluation of the initial development of different types of grapevine cv. Italia plants. Rev. Do Set. De Cienc. Agrar. 2000, 17, 131–142. [Google Scholar]
- Baydar, N.; Ece, M. Isparta koşullarında aşılı asma fidanı üretiminde farklı çeşit/anaç kombinasyonlarının karşılaştırılması. Süleyman Demirel Üniversitesi Fen Bilim. Enstitüsü Derg. 2005, 9, 1–5. [Google Scholar]
- Sivritepe, N.; Türkben, C. Müşküle üzüm çeşidinde farklı anaçların aşıda başarı ve fidan randımanı üzerine etkileri. Uludağ Üniversitesi Ziraat Fakültesi Derg. 2001, 15, 47–58. [Google Scholar]
- Vršič, S.; Valdhuber, J.; Pulko, B. Compatibility of the rootstock Börner with various scion varieties. Vitis 2004, 43, 155–156. [Google Scholar]
- Jogaiah, S.; Oulkar, D.; Banerjee, K.; Sharma, J.; Patil, A.; Maske, S.; Somkuwar, R. Biochemically induced variations during some phenological stages in Thompson Seedless grapevines grafted on different rootstocks. S. Afr. J. Enol. Vitic. 2013, 34, 36–45. [Google Scholar] [CrossRef] [Green Version]
- Akçaman, S.; Dardeniz, A. Red Globe üzüm çeşidinde farklı anaç kombinasyonlarının açık köklü aşılı fidan randımanları üzerine etkileri. Çomü Ziraat Fakültesi Derg. 2021, 9, 211–217. [Google Scholar] [CrossRef]
- Andrews, P.K.; Marquez, C.S. Graft incompatibility. Hortic. Rev. 1993, 15, 183–232. [Google Scholar]
- Çakır, A.; Yücel, B. Aşılı tüplü (kaplı) asma fidanı üretiminde farklı köklendirme ortamlarının kök ve sürgün gelişimi üzerine etkileri. Türk Doğa Ve Fen Derg. 2016, 5, 18–25. [Google Scholar]
- Holzapfel, B.P.; Smith, J.P. Developmental stage and climatic factors impact more on carbohydrate reserve dynamics of Shiraz than cultural practice. Am. J. Enol. Vitic. 2012, 63, 333–342. [Google Scholar] [CrossRef]
- Weyand, K.M.; Schultz, H.R. Long-term dynamics of nitrogen and carbohydrate reserves in woody parts of minimally and severely pruned Riesling vines in a cool climate. Am. J. Enol. Vitic. 2006, 57, 172–182. [Google Scholar]
- Zufferey, V.; Murisier, F.; Vivin, P.; Belcher, S.; Lorenzini, F.; Spring, J.-L.; Viret, O. Carbohydrate reserves in grapevine (Vitis vinifera L. ‘Chasselas’): The influence of the leaf to fruit ratio. Vitis 2012, 51, 103–110. [Google Scholar]
- Phillips, N.; Reynolds, A.; Di Profio, F. Nonstructural carbohydrate concentrations in dormant grapevine scionwood and rootstock impact propagation success and vine growth. HortTechnology 2015, 25, 536–550. [Google Scholar] [CrossRef] [Green Version]
- Prats-Llinàs, M.T.; García-Tejera, O.; Marsal, J.; Girona, J. Water stress during the post-harvest period affects new root formation but not starch concentration and content in Chardonnay grapevine (Vitis vinifera L.) perennial organs. Sci. Hortic. 2019, 249, 461–470. [Google Scholar] [CrossRef]
- Bates, T.R.; Dunst, R.M.; Joy, P. Seasonal dry matter, starch, and nutrient distribution in ‘Concord‘ grapevine roots. HortScience 2002, 37, 313–316. [Google Scholar] [CrossRef] [Green Version]
- Carlquist, S. Comparative wood anatomy. Systematic ecological and evolutionary aspects of dicotyledon wood. Int. For. Rev. 2001, 3, 330. [Google Scholar]
- Falchi, R.; Petrussa, E.; Zancani, M.; Casolo, V.; Beraldo, P.; Nardini, A.; Sivilotti, P.; Calderan, A.; Herrera, J.C.; Peterlunger, E. Summer drought stress: Differential effects on cane anatomy and non-structural carbohydrate content in overwintering Cabernet Sauvignon and Syrah vines. In Proceedings of the BIO Web of Conferences, Online, 1 April 2019; p. 03007. [Google Scholar]
- Pfautsch, S.; Renard, J.; Tjoelker, M.G.; Salih, A. Phloem as capacitor: Radial transfer of water into xylem of tree stems occurs via symplastic transport in ray parenchyma. Plant Physiol. 2015, 167, 963–971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Earles, J.M.; Knipfer, T.; Tixier, A.; Orozco, J.; Reyes, C.; Zwieniecki, M.A.; Brodersen, C.R.; McElrone, A.J. In vivo quantification of plant starch reserves at micrometer resolution using X-ray micro CT imaging and machine learning. New Phytol. 2018, 218, 1260–1269. [Google Scholar] [CrossRef] [Green Version]
- Hunter, J.; Volschenk, C.; Le Roux, D.; Fouché, G.; Adams, L. Plant Material Quality: A Compilation of Research; ARC Infruitec-Nietvoorbij Private Bag X5026, 7599; ARC Infruitec-Nietvoorbij: Stellenbosch, South Africa, 2004; p. 18. [Google Scholar]
- Bahar, E.; Korkutal, İ.; Kök, D. Hidroponik kültür ve fidanlık koşullarında yetiştirilen aşılı asma fidanlarının karbonhidrat ve azot İçerikleri İle bağdaki tutma performansları üzerine araştırmalar. Akdeniz Üniversitesi Ziraat Fakültesi Derg. 2008, 21, 15–26. [Google Scholar]
- Vršič, S.; Pulko, B.; Valdhuber, J. Influence of defoliation on carbohydrate reserves of young grape-vines in the nursery. Eur. J. Hortic. Sci 2009, 74, 218–222. [Google Scholar]
- Cutler, D.F.; Botha, C.E.J.; Stevenson, D.W. Plant Anatomy: An Applied Approach; Blackwell: Oxford, UK, 2008. [Google Scholar]
- Cookson, S.J.; Prodhomme, D.; Chambaud, C.; Hevin, C.; Valls Fonayet, J.; Hilbert, G.; Trossat-Magnin, C.; Richard, T.; Bortolami, G.; Gambetta, G.A. Understanding scion-rootstock interactions at the graft interface of grapevine. Acta Hortic. 2019, 1248, 369–374. [Google Scholar] [CrossRef]
- Martínez-Ballesta, M.C.; Alcaraz-López, C.; Muries, B.; Mota-Cadenas, C.; Carvajal, M. Physiological aspects of rootstock–scion interactions. Sci. Hortic. 2010, 127, 112–118. [Google Scholar] [CrossRef]
- Çakırlar, H.; Doğan, C.; Özmen, E. Açıklamalı Genel Botanik ve Bitki Anatomisi Atlası, 4th ed.; Palme Yayıncılık: Ankara, Türkiye, 2017; 181p. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | Rootstock | Callus Formation Rate (%) | Bud Burst (%) | Callus Development Level (0–4) | Root Formation Rate (%) |
|---|---|---|---|---|---|
| Rizessi | 140Ru | 82.00 | 75.33 | 2.18 gh | 41.33 ef |
| SO4 | 100.00 | 93.33 | 3.55 cde | 82.67 abcd | |
| 110R | 99.33 | 95.33 | 3.91 a | 50.00 e | |
| Çeliksu | 140Ru | 90.67 | 90.67 | 2.31 fg | 51.33 e |
| SO4 | 99.33 | 90.67 | 3.71 abcd | 80.00 bcd | |
| 110R | 98.67 | 96.67 | 3.68 abcd | 73.33 cd | |
| Ülkemiz | 140Ru | 85.33 | 78.00 | 1.95 h | 71.33 d |
| SO4 | 99.33 | 80.67 | 3.83 ab | 84.67 abcd | |
| 110R | 99.33 | 89.33 | 3.65 bcd | 86.00 abc | |
| Rizellim | 140Ru | 92.00 | 90.67 | 2.52 f | 33.33 f |
| SO4 | 100.00 | 98.00 | 3.53 de | 84.67 abcd | |
| 110R | 100.00 | 96.00 | 3.34 e | 80.67 bcd | |
| Rizpem | 140Ru | 95.00 | 75.00 | 2.47 f | 45.00 ef |
| SO4 | 100.00 | 85.00 | 3.80 abc | 88.33 ab | |
| 110R | 100.00 | 88.33 | 3.67 abcd | 90.00 a | |
| LSD | N.S | N.S | 1.14 p < 0.01 | 9.72 p < 0.01 | |
| Cultivar | Rootstock | Sapling Rate (%) | First-Class Sapling Rate (%) | Shoot Development Level (0–4) | Primarily Shoot Diameter (mm) | Primarily Shoot Length (cm) |
|---|---|---|---|---|---|---|
| Rizessi | 140Ru | 46.24 d | 35.72 c | 3.55 | 6.50 ab | 137.15 ab |
| SO4 | 72.79 bc | 65.31 ab | 3.04 | 6.11 abcd | 93.13 cde | |
| 110R | 48.98 d | 38.24 c | 3.26 | 6.30 abc | 104.45 cd | |
| Çeliksu | 140Ru | 48.57 d | 38.10 c | 3.26 | 6.30 abc | 141.55 ab |
| SO4 | 85.62 a | 67.90 ab | 2.92 | 5.04 ef | 75.95 def | |
| 110R | 70.39 c | 60.83 b | 3.48 | 5.64 cde | 110.24 bc | |
| Ülkemiz | 140Ru | 22.87 e | 16.31 d | 3.05 | 5.40 def | 95.86 cde |
| SO4 | 45.67 d | 37.61 c | 3.10 | 5.40 def | 84.39 cdef | |
| 110R | 47.22 d | 34.72 c | 3.30 | 5.88 bcde | 93.24 cde | |
| Rizellim | 140Ru | 45.89 d | 36.08 c | 3.59 | 6.95 a | 151.79 a |
| SO4 | 86.00 a | 75.33 a | 3.35 | 5.53 cdef | 87.05 cdef | |
| 110R | 81.94 ab | 70.83 ab | 3.44 | 5.65 bcde | 101.68 cd | |
| Rizpem | 140Ru | 27.14 e | 11.27 d | 2.62 | 4.71 f | 67.28 ef |
| SO4 | 53.33 d | 36.67 c | 2.74 | 5.61 cde | 76.28 def | |
| 110R | 28.33 e | 15.00 d | 2.81 | 5.98 bcd | 59.84 f | |
| LSD | 8.33 p < 0.01 | 7.71 p < 0.05 | N.S | 0.85 p < 0.05 | 32.07 p < 0.05 | |
| Cultivar | Rootstock | Graft Area Diameter (mm) | Rootstock Diameter (mm) | Root Development Level (0–4) | Root Length (cm) | Number of Roots (pcs) |
|---|---|---|---|---|---|---|
| Rizessi | 140Ru | 19.92 abc | 10.20 bcd | 3.35 | 56.46 a | 13.32 |
| SO4 | 19.66 abc | 10.56 bcd | 3.26 | 52.07 abc | 13.59 | |
| 110R | 20.49 a | 10.21 bcd | 3.00 | 43.97 def | 11.82 | |
| Çeliksu | 140Ru | 19.83 abc | 9.75 def | 2.87 | 50.62 abcd | 9.84 |
| SO4 | 18.46 cde | 10.13 cde | 3.23 | 55.65 ab | 15.37 | |
| 110R | 20.31 a | 11.62 a | 3.16 | 37.89 fg | 9.46 | |
| Ülkemiz | 140Ru | 14.14 f | 9.31 ef | 2.61 | 48.35 cd | 9.68 |
| SO4 | 17.71 de | 10.56 bcd | 3.40 | 51.60 abc | 13.22 | |
| 110R | 18.59 bcd | 9.96 de | 3.10 | 46.11 cde | 9.28 | |
| Rizellim | 140Ru | 20.30 a | 10.12 cde | 2.76 | 47.05 cde | 10.25 |
| SO4 | 19.68 abc | 10.98 ab | 3.39 | 48.99 bcd | 16.97 | |
| 110R | 20.16 ab | 10.79 abc | 2.95 | 40.09 efg | 9.06 | |
| Rizpem | 140Ru | 14.25 f | 9.09 f | 2.77 | 35.07 g | 12.04 |
| SO4 | 16.95 e | 9.02 f | 3.06 | 50.99 abcd | 16.76 | |
| 110R | 17.12 de | 9.32 ef | 3.11 | 49.19 bcd | 11.69 | |
| LSD | 0.92 p < 0.01 | 0.83 p < 0.05 | N.S | 7.18 p < 0.01 | N.S | |
| Root Total Carbohydrates | UTG Total Carbohydrates | GA Total Carbohydrates | ATG Total Carbohydrates | |
|---|---|---|---|---|
| Sapling rate | −0.361 * | 0.133 | 0.387 ** | 0.083 |
| First-class sapling rate | −0.400 * | 0.137 | 0.358 * | 0.072 |
| Shoot development level | −0.335 * | 0.063 | 0.164 | 0.060 |
| Primarily shoot diameter | −0.279 | 0.074 | 0.155 | −0.037 |
| Primarily shoot length | −0.281 | −0.085 | −0.020 | 0.076 |
| Graft area diameter | −0.434 * | 0.182 | 0.423 * | 0.082 |
| Rootstock diameter | −0.352 * | 0.261 | 0.261 | 0.168 |
| Root development level | −0.121 | 0.143 | 0.339 * | −0.351 * |
| Number of roots | −0.157 | −0.078 | 0.288 | −0.443 * |
| Root length | 0.071 | −0.291 | −0.046 | −0.310 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karabulut, B.; Çelik, H. Determination of Grafting Success and Carbohydrate Distributions of Foxy Grape (Vitis labrusca L.) Varieties Grafted on Different American Grape Rootstocks. Horticulturae 2022, 8, 949. https://doi.org/10.3390/horticulturae8100949
Karabulut B, Çelik H. Determination of Grafting Success and Carbohydrate Distributions of Foxy Grape (Vitis labrusca L.) Varieties Grafted on Different American Grape Rootstocks. Horticulturae. 2022; 8(10):949. https://doi.org/10.3390/horticulturae8100949
Chicago/Turabian StyleKarabulut, Besim, and Hüseyin Çelik. 2022. "Determination of Grafting Success and Carbohydrate Distributions of Foxy Grape (Vitis labrusca L.) Varieties Grafted on Different American Grape Rootstocks" Horticulturae 8, no. 10: 949. https://doi.org/10.3390/horticulturae8100949