
Effects of Arbuscular Mycorrhizal Inoculation by Indigenous Fungal Complexes on the Morpho-Physiological Behavior of Argania spinosa Subjected to Water Deficit Stress


 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Soil/Root Sampling
2.2. Preparation of the Inoculum
2.3. Plant Material
2.4. Experimental Setup
2.5. Analysis of the Effects of Controlled Mycorrhization on the Morphophysiological Responses of the Argan Tree Tolerance to Water Stress
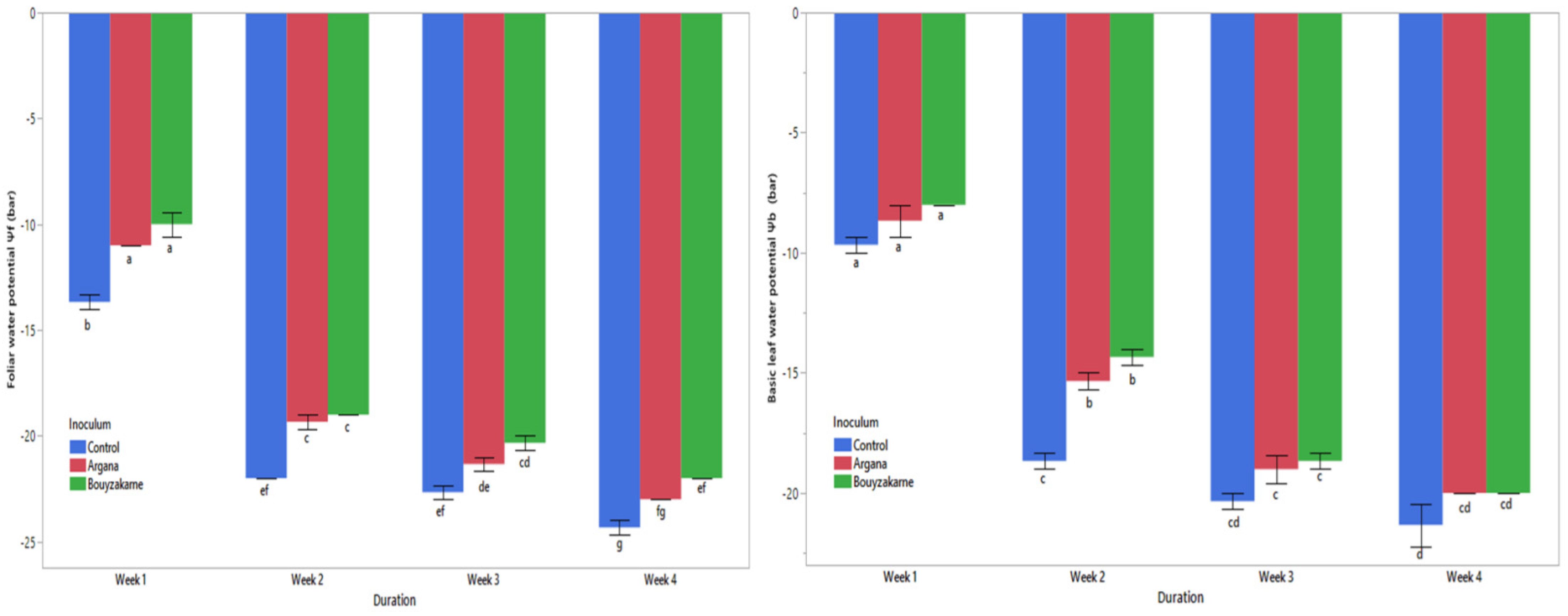
2.5.1. Leaf Water Potential
Basic Leaf Water Potential (Ψb)
Leaf Water Potential at Noon (Ψf)
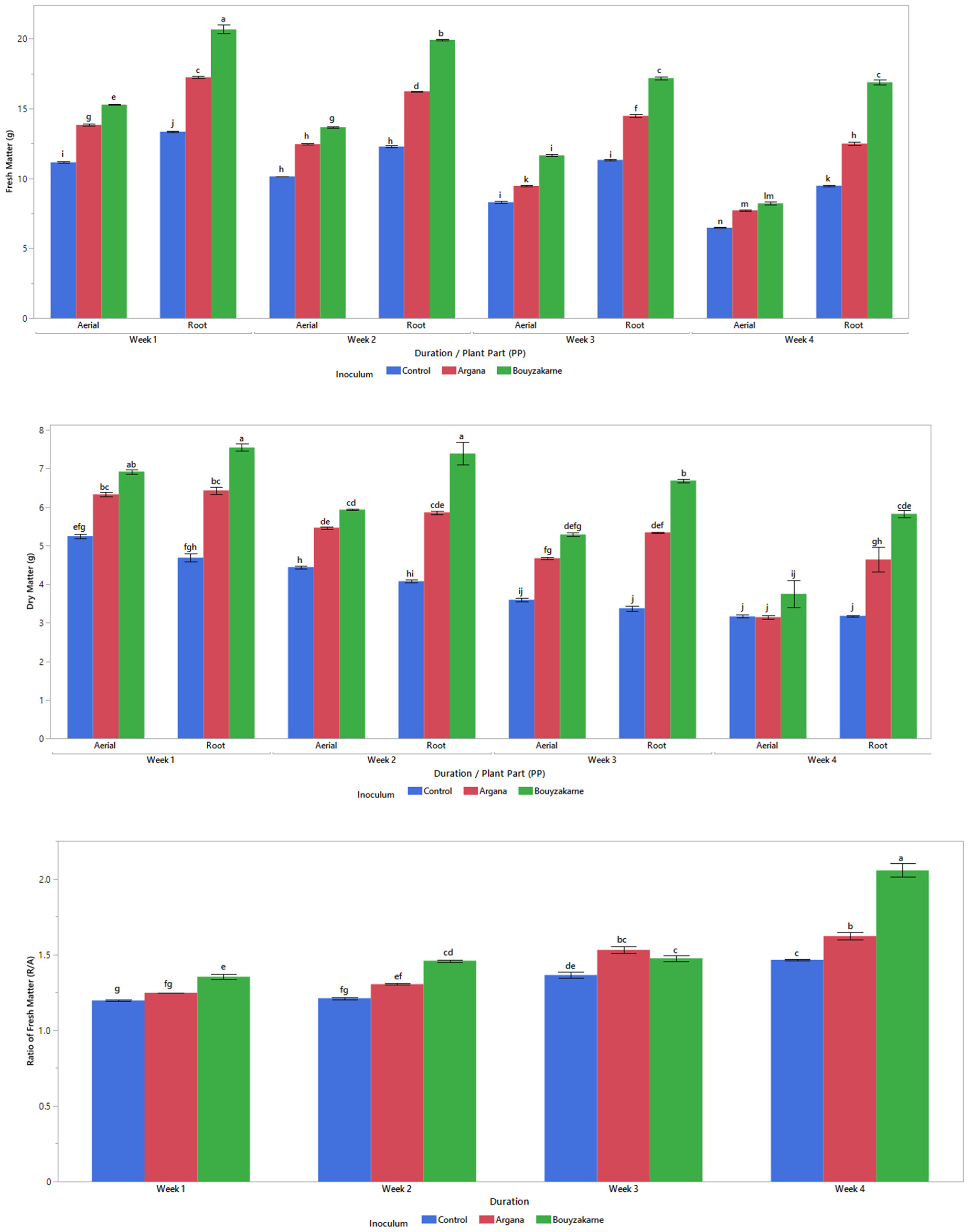
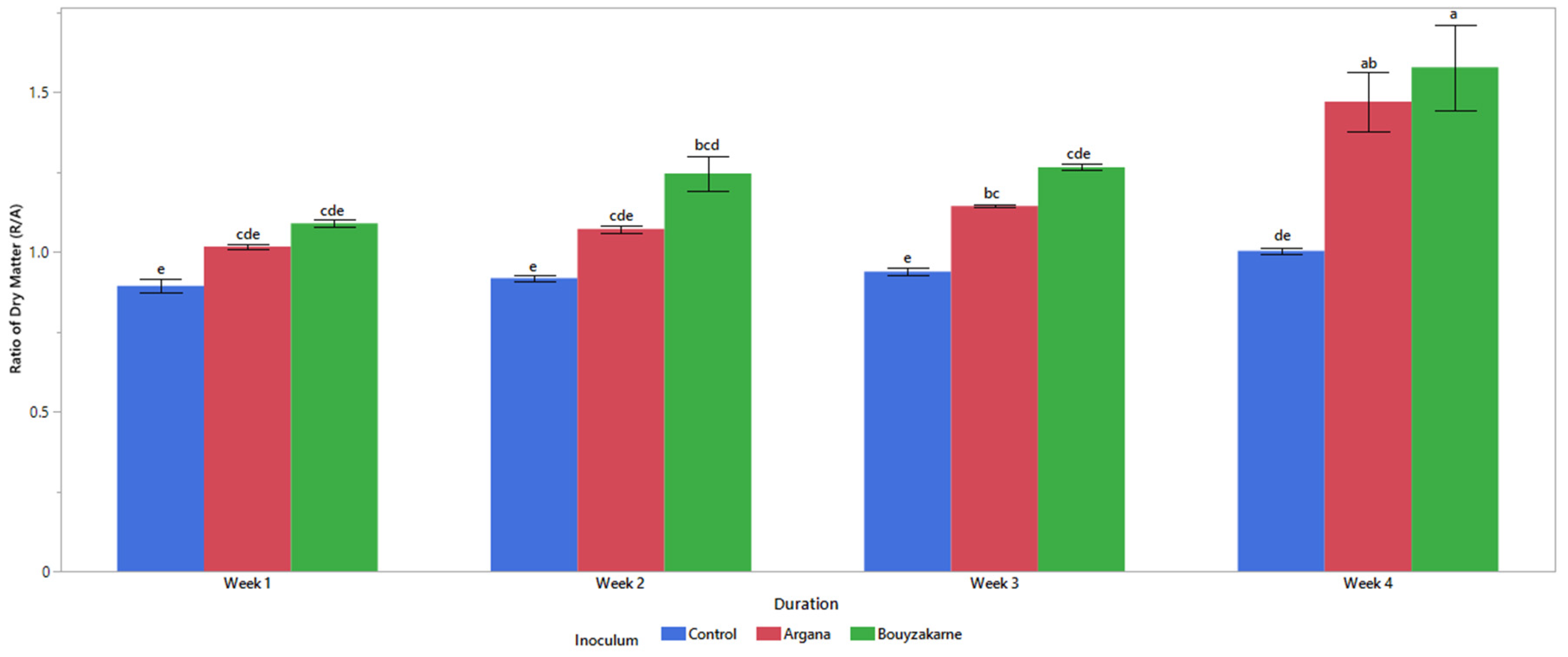
2.5.2. Biomass
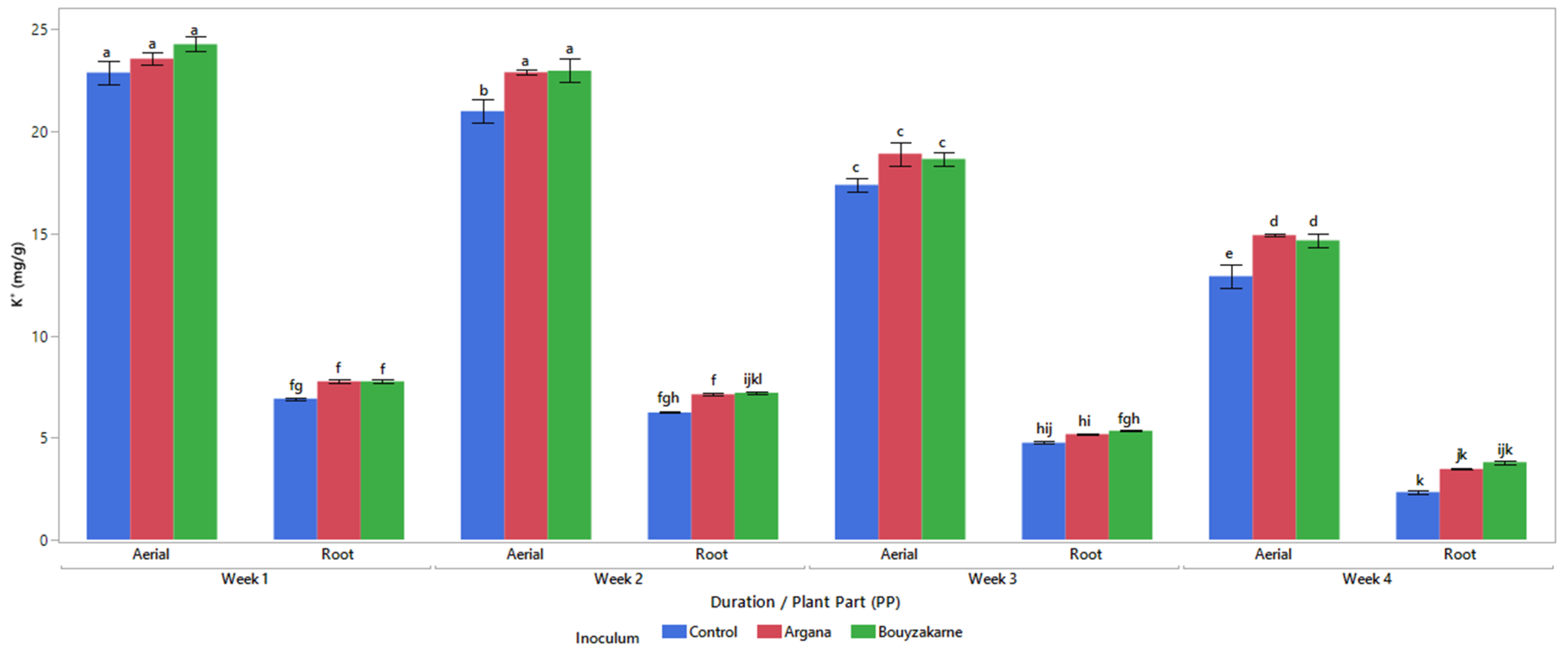
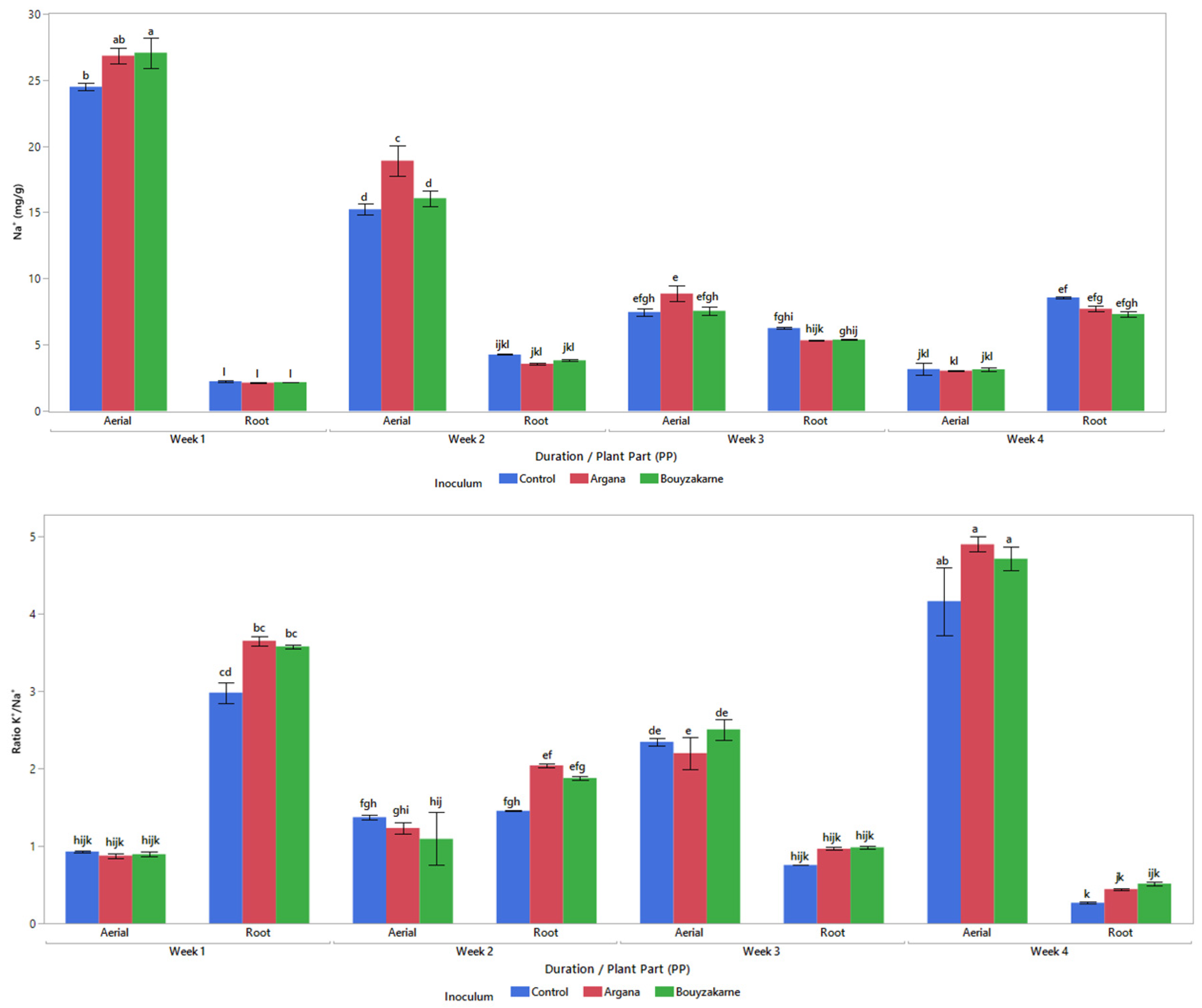
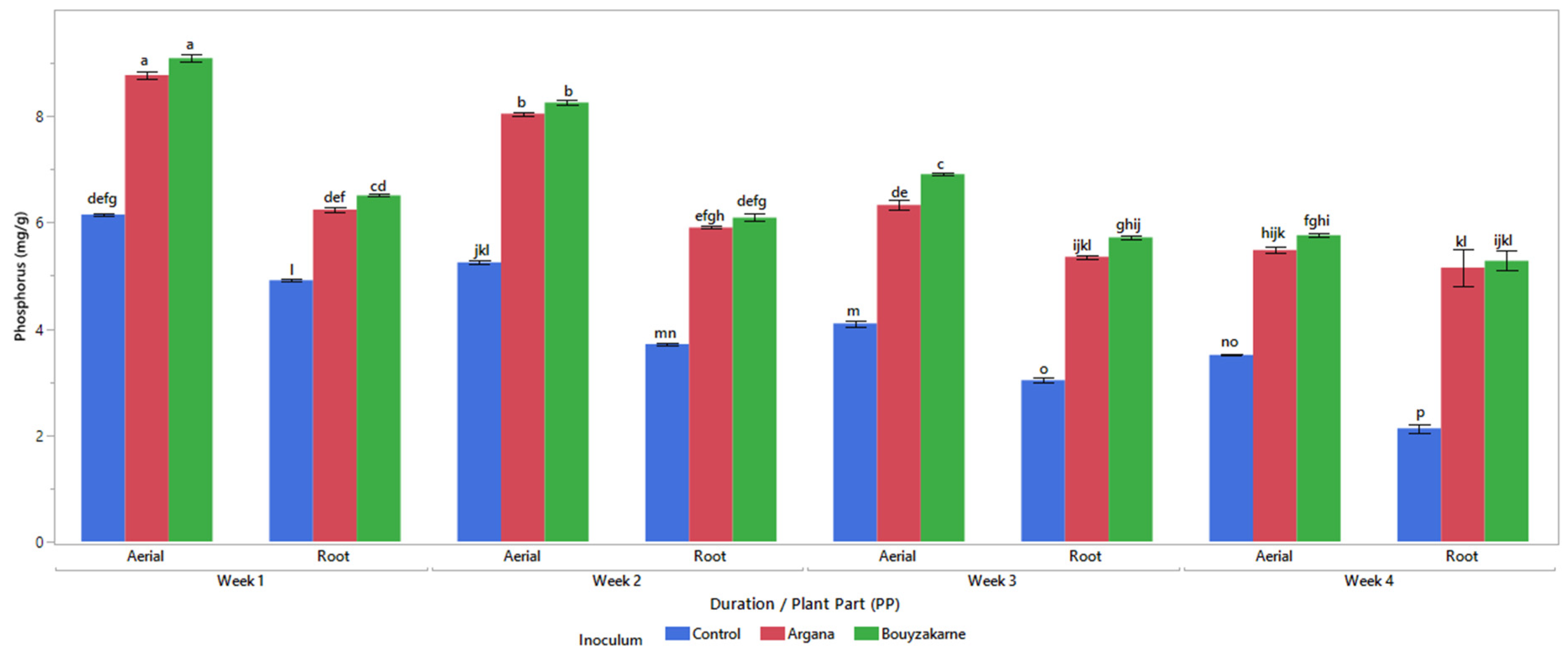
2.5.3. Mineral Content
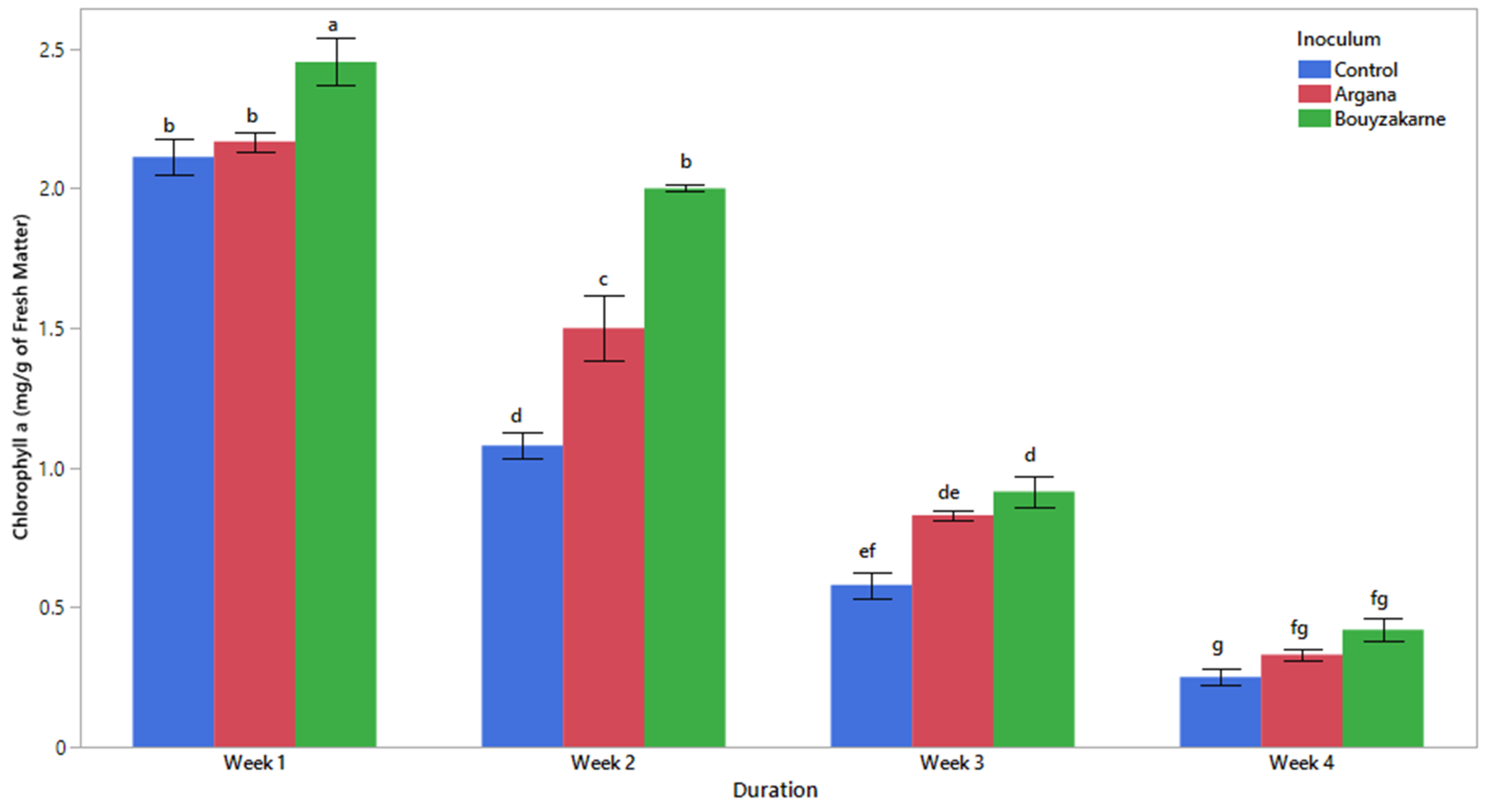
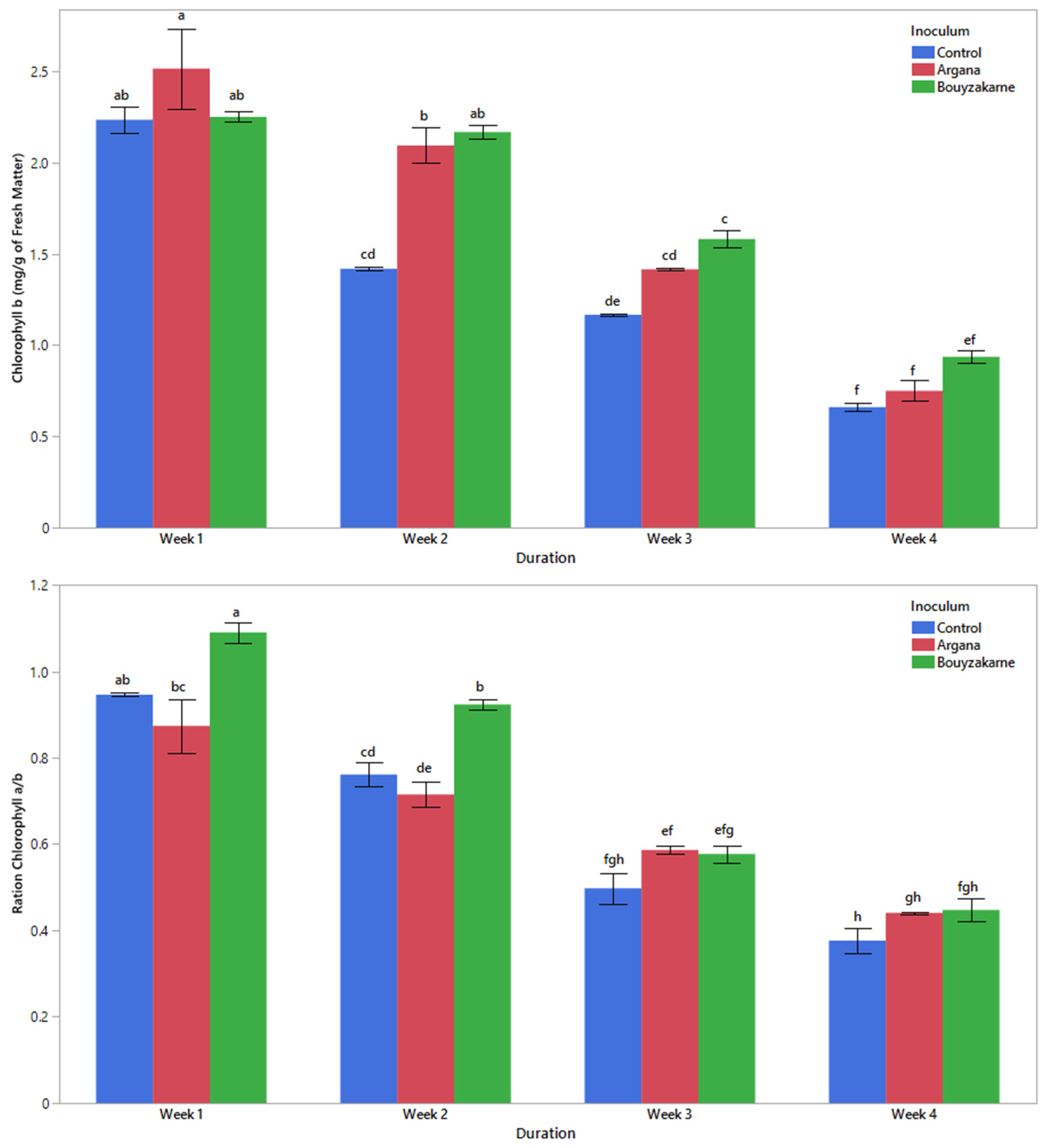
2.5.4. Chlorophyll Pigment Content
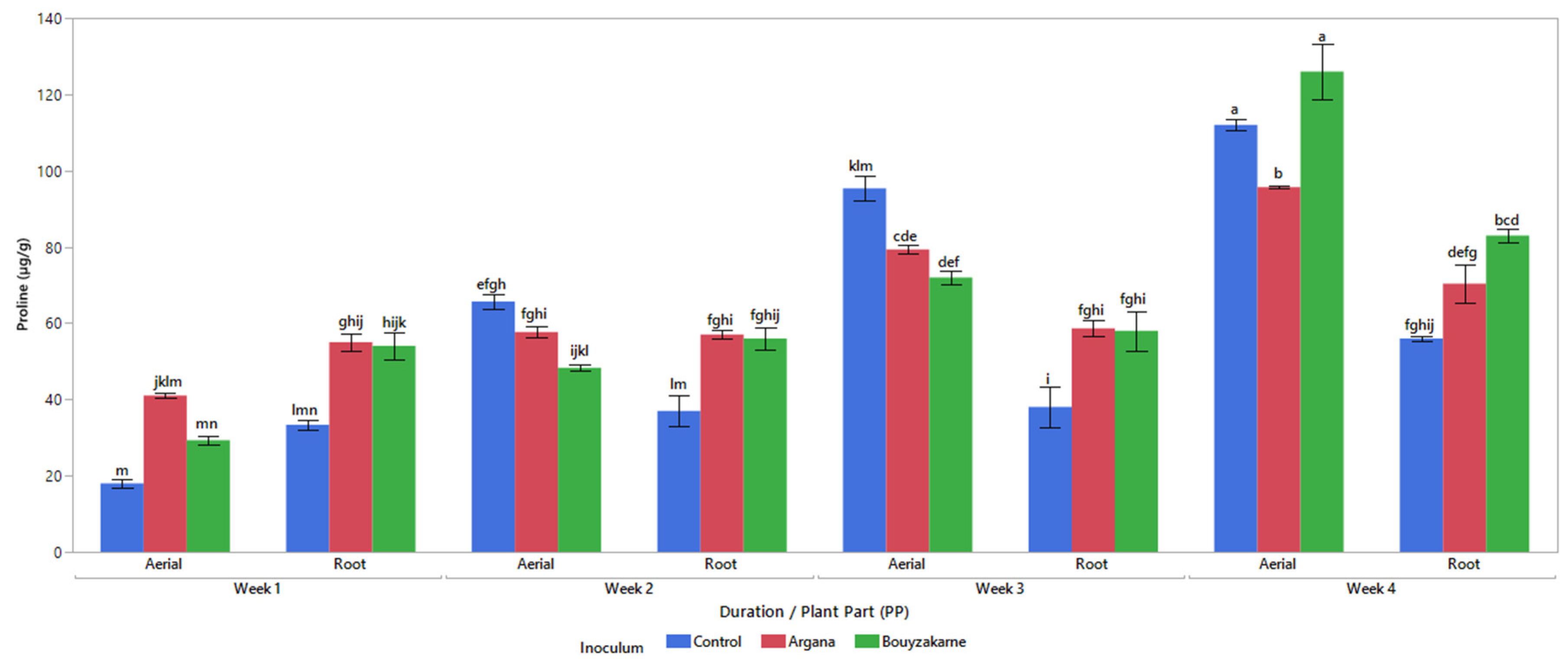
2.5.5. Free Proline Content
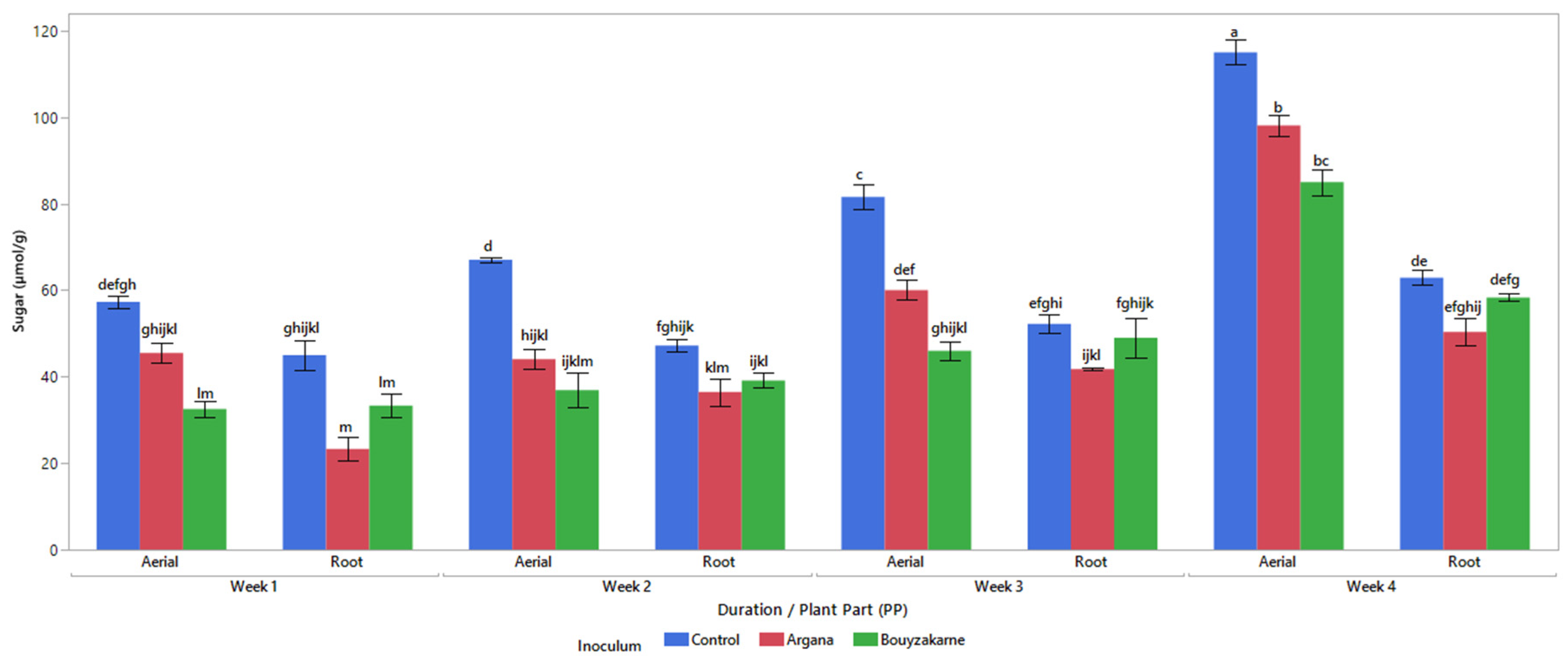
2.5.6. Total Soluble Sugar Content
2.6. Statistical Analyses
3. Results
3.1. Analysis of Variance of the Parameters Measured for the Aerial and Root Parts
3.2. Morphophysiological Characters
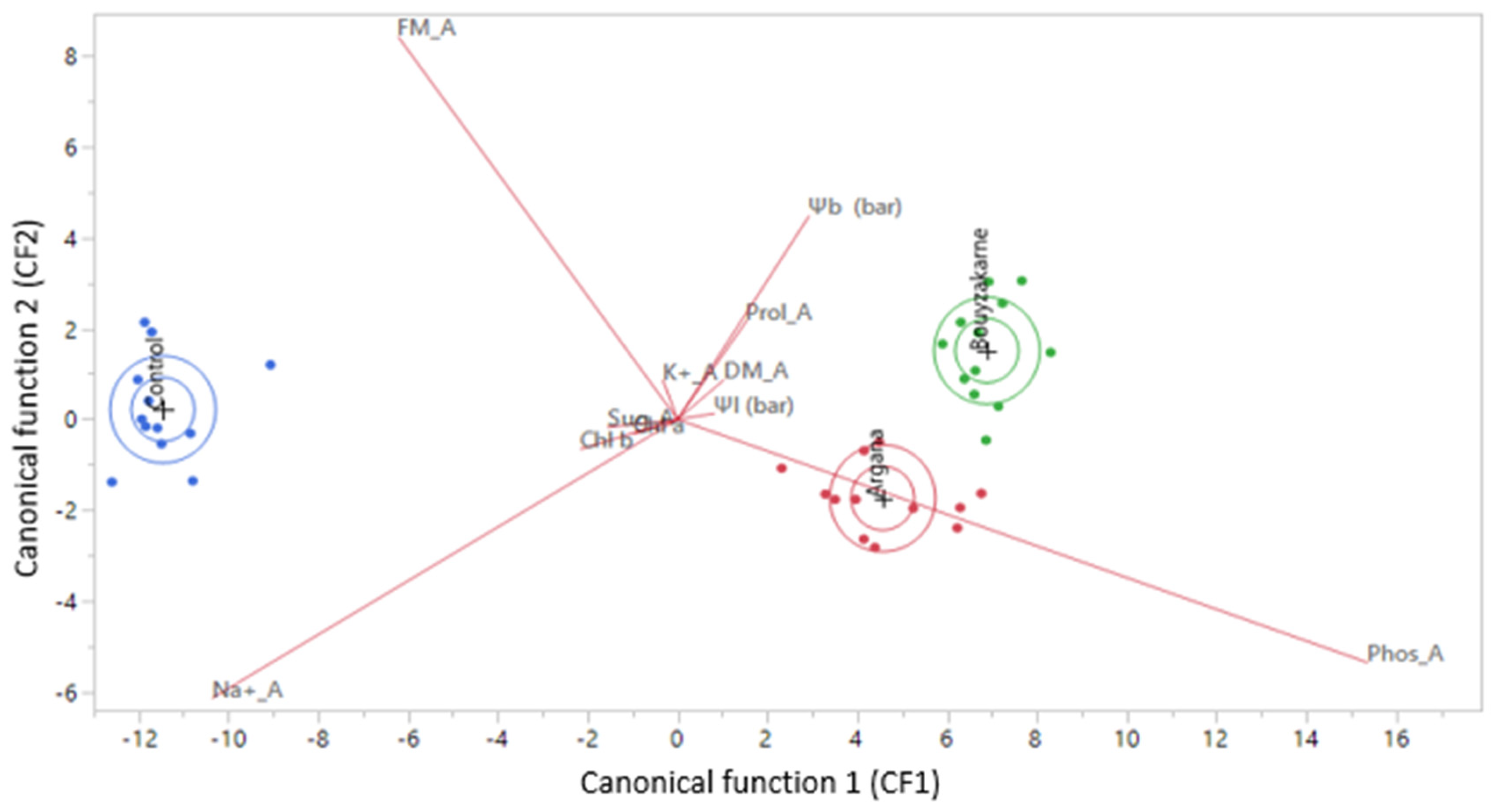
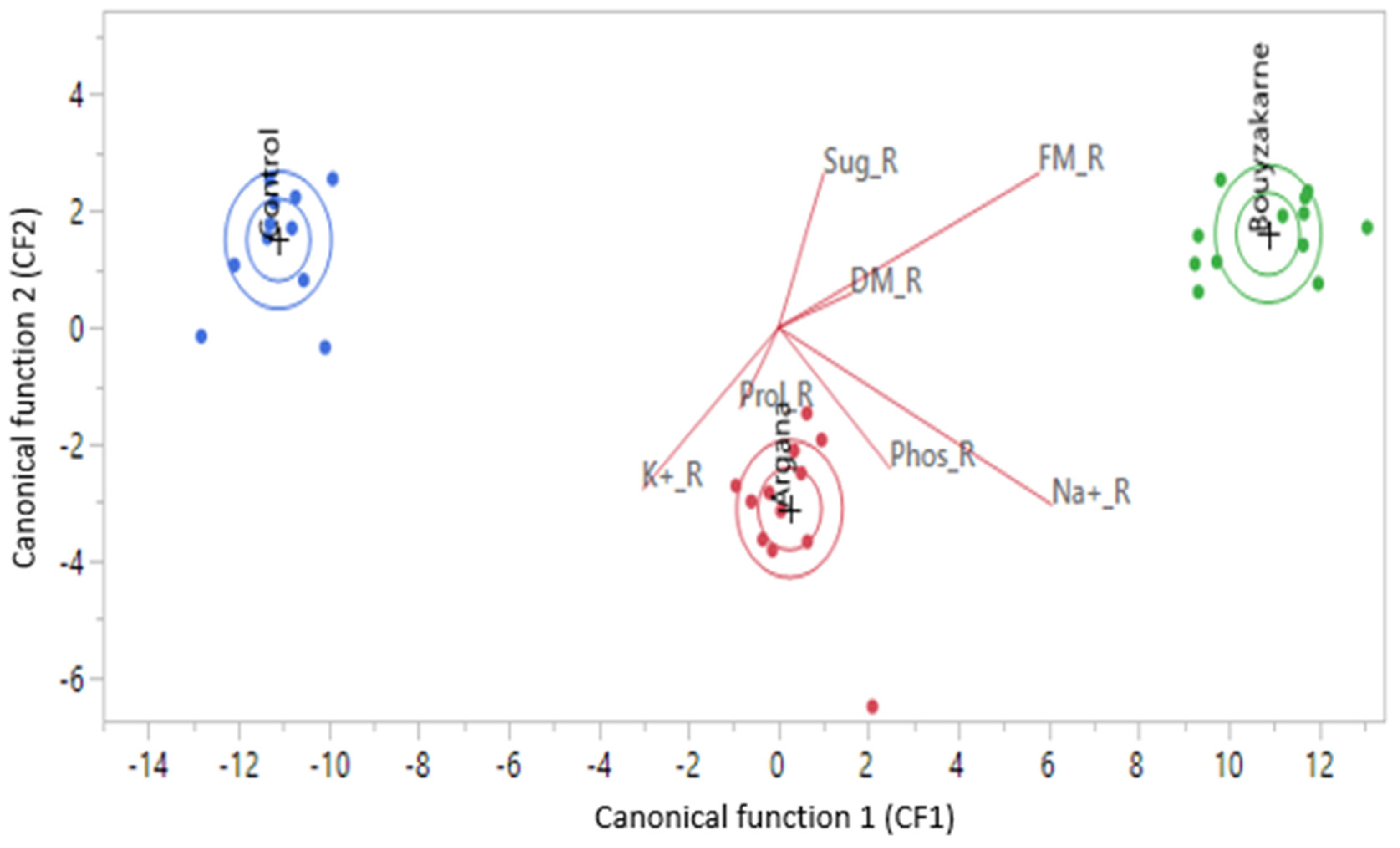
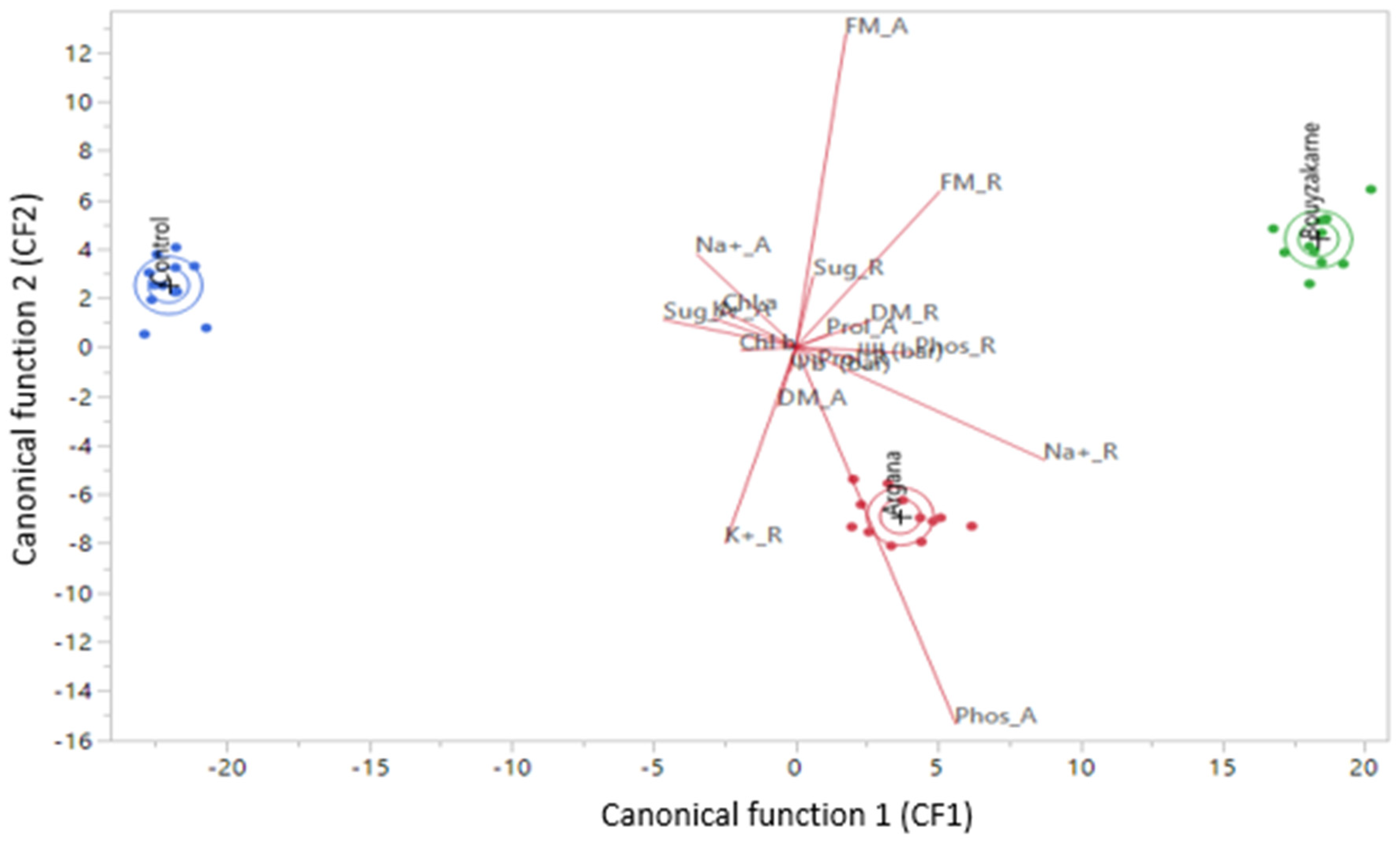
3.3. Canonical Discriminant Analysis of Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nouaim, R.; Chaussod, R.; El Aboudi, A.; Schnabel, C.; Peltier, J.P. L’arganier: Essai de Synthèse des Connaissances sur cet Arbre. Dans: Physiologie des Arbres et Arbustes en Zones Aride et Semi-Aride; Groupe d’étude de l’arbre: Paris, France, 1991; pp. 373–388. [Google Scholar]
- Bethlenfalvay, G.J.; Brown, M.S.; Ames, R.N.; Thomas, R.S. Effects of Drought on Host and Endophyte Development in Mycorrhizal Soybeans in Relation to Water Use and Phosphate Uptake. Physiol. Plant. 1988, 72, 565–571. [Google Scholar] [CrossRef]
- Oliet, J.; Puértolas, J.; Planelles, R.; Jacobs, D. Nutrient Loading of Forest Tree Seedlings to Promote Stress Resistance and Field Performance: A Mediterranean Perspective. New For. 2013, 44, 649–669. [Google Scholar] [CrossRef]
- Calvo-Polanco, M.; Sánchez-Romera, B.; Aroca, R.; Asins, M.J.; Declerck, S.; Dodd, I.C.; Martínez-Andújar, C.; Albacete, A.; Ruiz-Lozano, J.M. Exploring the Use of Recombinant Inbred Lines in Combination with Beneficial Microbial Inoculants (AM Fungus and PGPR) to Improve Drought Stress Tolerance in Tomato. Environ. Exp. Bot. 2016, 131, 47–57. [Google Scholar] [CrossRef] [Green Version]
- Fischlin, A.; Midgley, G.F.; Price, J.T.; Leemans, R.; Gopal, B.; Turley, C. Ecosystems, Their Properties, Goods and Services. Climate Change 2007: Impacts, Adaptation and Vulnerability. Contribution of Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Parry, M.L., Canziani, O.F., Palutikof, J.P., van der Linden, P.J., Hanson, C.E., Eds.; Cambridge University Press: Cambridge, UK, 2007; pp. 211–272. [Google Scholar]
- Salehi-Lisar, S.Y.; Bakhshayeshan-Agdam, H. Drought Stress in Plants: Causes, Consequences, and Tolerance. In Drought Stress Tolerance in Plants, Vol 1: Physiology and Biochemistry; Hossain, M.A., Wani, S.H., Bhattacharjee, S., Burritt, D.J., Tran, L.-S.P., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 1–16. ISBN 978-3-319-28899-4. [Google Scholar] [CrossRef]
- Kaushal, M.; Wani, S.P. Rhizobacterial-Plant Interactions: Strategies Ensuring Plant Growth Promotion under Drought and Salinity Stress. Agric. Ecosyst. Environ. 2016, 231, 68–78. [Google Scholar] [CrossRef]
- Xiong, L.; Wang, R.; Mao, G.; Koczan, J.M. Identification of drought tolerance determinants by genetic analysis of root response to drought stress and abscisic acid. Plant Physiol. 2006, 142, 1065–1074. [Google Scholar] [CrossRef] [Green Version]
- Chun, S.C.; Paramasivan, M.; Chandrasekaran, M. Proline Accumulation Influenced by Osmotic Stress in Arbuscular Mycorrhizal Symbiotic Plants. Front. Microbiol. 2018, 9, 2525. [Google Scholar] [CrossRef] [Green Version]
- Asmelash, F.; Bekele, T.; Birhane, E. The potential role of arbuscular mycorrhizal fungi in the restoration of degraded lands. Front. Microbiol. 2016, 26, 1095. [Google Scholar] [CrossRef] [Green Version]
- Duponnois, R.; Hafidi, M.; NDoya, I.; Ramanankierana, H.; Bà, M.A. Des Champignons Symbiotiques Contre La Désertification: Écosystèmes Méditerranéens, Tropicaux et Insulaires; IRD ORSTOM: Marseille, France, 2013; ISBN 978-2-7099-1827-5. [Google Scholar]
- Smith, S.E.; Smith, F.A. Fresh Perspectives on the Roles of Arbuscular Mycorrhizal Fungi in Plant Nutrition and Growth. Mycologia 2012, 104, 1–13. [Google Scholar] [CrossRef]
- Ruiz-Lozano, J.M.; Azcón, R. Hyphal Contribution to Water Uptake in Mycorrhizal Plants as Affected by the Fungal Species and Water Status. Physiol. Plant. 1995, 95, 472–478. [Google Scholar] [CrossRef]
- Navarro García, A.; Del Pilar Bañón Árias, S.; Morte, A.; Sánchez-Blanco, M.J. Effects of Nursery Preconditioning through Mycorrhizal Inoculation and Drought in Arbutus Unedo L. Plants. Mycorrhiza 2011, 21, 53–64. [Google Scholar] [CrossRef]
- Mycorrhizal Symbiosis, 3rd ed. Available online: https://www.elsevier.com/books/mycorrhizal-symbiosis/smith/978-0-12-370526-6 (accessed on 10 February 2022).
- Effects of Arbuscular Mycorrhizal Fungus and Humic Acid on the Seedling Development and Nutrient Content of Pepper Grown under Saline Soil Conditions. Available online: https://scialert.net/abstract/?doi=jbs.2005.568.574 (accessed on 10 February 2022).
- Sheng, M.; Tang, M.; Chen, H.; Yang, B.; Zhang, F.; Huang, Y. Influence of Arbuscular Mycorrhizae on Photosynthesis and Water Status of Maize Plants under Salt Stress. Mycorrhiza 2008, 18, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Song, F.; Xu, H. Influence of Arbuscular Mycorrhiza on Lipid Peroxidation and Antioxidant Enzyme Activity of Maize Plants under Temperature Stress. Mycorrhiza 2010, 20, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Filho, J.A.C.; Sobrinho, R.R.; Pascholati, S.F. Arbuscular Mycorrhizal Symbiosis and Its Role in Plant Nutrition in Sustainable Agriculture. In Agriculturally Important Microbes for Sustainable Agriculture; Meena, V.S., Mishra, P.K., Bisht, J.K., Pattanayak, A., Eds.; Springer: Singapore, 2017; pp. 129–164. ISBN 978-981-10-5342-9. [Google Scholar]
- Augé, R.M. Water Relations, Drought and Vesicular-Arbuscular Mycorrhizal Symbiosis. Mycorrhiza 2001, 11, 3–42. [Google Scholar] [CrossRef]
- Bousselmane, F.; Kenny, L.; Achouri, M. Effet des mycorhizes à vésicules et arbuscules sur la croissance et la nutrition de l’arganier (Argania spinosa L.). Rev. Maroc. Des. Sci. Agron. Vétérinaires 2002, 22, 193–198. [Google Scholar]
- Echairi, A.; Nouaim, R.; Chaussod, R. Intérêt de La Mycorhization Contrôlée Pour La Production de Plants d’arganier (Argania spinosa) En Conditions de Pépinière. Sci. Changements Planét./Sécher. 2008, 19, 277–281. [Google Scholar] [CrossRef]
- El Mrabet, E.; Lahcen, O.; A., E.; Msanda, F.; Abbas, Y. L’efficacité de l’inoculation Mycorhyzienne et de l’addition Du Bio-Compost Sur Le Développement d’Argania Spinosa Sur Le Champ. In Proceedings of the Actes du 2e Congrès International de l’arganier, Agadir, Morocco, 9–11 December 2013. [Google Scholar]
- Ouallal, I.; Abbas, Y.; Ech-cheddadi, S.; Ouajdi, M.; Ouhadach, M.; El Yacoubi, H.; Kerdouh, B.; El Goumi, Y.; Rochdi, A. Diversité des champignons endomycorhiziens de l’arganier et potentiel mycorhizogène des sols rhizosphériques des arganeraies du Sud-Ouest marocain. Bois For. Des. Tropiques. 2019, 338, 73–86. [Google Scholar] [CrossRef]
- Rochdi, A.; Ouallal, I.; ElYacoubi, H.; Abbas, Y. Pouvoir mycorhizogene des sols rhizospheriques des arganeraies du sud-ouest du maroc. In Proceedings of the Actes du 5ème Congrès International de l’Arganier, Maroc, Agadir, Morocco, 10–11 December 2019; pp. 431–435. [Google Scholar]
- Arnon, D.I. Copper Enzymes in Isolated Chloroplasts. Polyphenoloxidase in Beta Vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Saurin, N.; Tisseyre, B.; Lebon, E. Comment Mesurer La Contrainte Hydrique de La Vigne, de La Plante Au Vignoble. Innov. Agron. 2014, 38, 143–158. [Google Scholar]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid Determination of Free Proline for Water-Stress Studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, k.L.; Hamilton, J.K.; Reberg, P.A.; Smith, F. Colorimetric Method For Determination Of Sugars And Related Substanses. Analatycal. Chemistry 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Villagra, P.E.; Cavagnaro, J.B. Water stress effects on the seedling growth of Prosopis argentina and Prosopis alpataco. J. Arid. Environ. 2006, 64, 390–400. [Google Scholar] [CrossRef]
- Boudjabi, S.; Kribaa, M.; Chenchouni, H. Growth, physiology and yield of durum wheat (Triticum durum) treated with sewage sludge under water stress conditions. Excli J. 2015, 14, 320–334. [Google Scholar] [PubMed]
- Posta, K.; Duc, N.H. Benefits of Arbuscular Mycorrhizal Fungi Application to Crop Production under Water Scarcity. Drought Detect Solut 2020. Available online: https://pdfs.semanticscholar.org/93d8/475065c847e7b2574eb228c730d33fb2c961.pdf (accessed on 1 March 2022).
- Bernardo, L.; Carletti, P.; Badeck, F.; Rizza, F.; Morcia, C.; Ghizzoni, R.; Rouphael, Y.; Colla, G.; Terzi, V.; Lucini, L. Metabolomic responses triggered by arbuscular mycorrhiza enhance tolerance to water stress in wheat cultivars. Plant Physiol. Biochem. 2019, 137, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Dashti, N.H.; Smith, D.L.; Cherian, V.M. PGPR to Alleviate the Stress of Suboptimal Root Zone Temperature on Leguminous Plant Growth. In Use of Microbes for the Alleviation of Soil Stresses, Volume 1; Miransari, M., Ed.; Springer: New York, NY, USA, 2014; pp. 111–137. ISBN 978-1-4614-9466-9.34. [Google Scholar]
- Marulanda, A.; Barea, J.M.; Azcón, R. An Indigenous Drought-Tolerant Strain of Glomus intraradices Associated with a Native Bacterium Improves Water Transport and Root Development in Retama sphaerocarpa. Microbiol. Ecol. 2006, 52, 670. [Google Scholar] [CrossRef]
- Ruiz-Lozano, J.M.; Aroca, R. Modulation of Aquaporin Genes by the Arbuscular Mycorrhizal Symbiosis in Relation to Osmotic Stress Tolerance. In Symbioses and Stress; Seckbach, J., Grube, M., Eds.; Cellular Origin, Life in Extreme Habitats and Astrobiology; Springer: Dordrecht, The Netherlands, 2010; Volume 17, pp. 357–374. ISBN 978-90-481-9448-3. [Google Scholar]
- Ruth, B.; Khalvati, M.; Schmidhalter, U. Quantification of mycorrhizal water uptake via high-resolutio on-line water content sensors. Plant Soil 2011, 342, 459–468. [Google Scholar] [CrossRef]
- Rillig, M.C. Arbuscular mycorrhizae, glomalin, and soil aggregation. Candian J. Soil Sci. 2004, 84, 355–363. [Google Scholar] [CrossRef] [Green Version]
- Driver, J.D.; Holben, W.E.; Rillig, M.C. Characterization of glomalin as a hyphal wall component of arbuscular mycorrhizal fungi. Soil Biol. Biochem. 2005, 37, 101–106. [Google Scholar] [CrossRef]
- Zarik, L.; Meddich, A.; Hijri, M.; Hafidi, M.; Ouhammou, A.; Ouahmane, L.; Duponnois, R.; Boumezzough, A. Use of Arbuscular Mycorrhizal Fungi to Improve the Drought Tolerance of Cupressus Atlantica G. C. R. Biol. 2016, 339, 185–196. [Google Scholar] [CrossRef]
- Abbaspour, H.; Saeidi-Sar, S.; Afshari, H.; Abdel-Wahhab, M.A. Tolerance of Mycorrhiza Infected Pistachio (Pistacia Vera L.) Seedling to Drought Stress under Glasshouse Conditions. J. Plant Physiol. 2012, 169, 704–709. [Google Scholar] [CrossRef]
- Guissou, T.; Bâ, A.; Plenchette, C.; Guinko, S.; Duponnois, R. Effets Des Mycorhizes à Arbuscules Sur La Tolérance à Un Stress Hydrique de Quatre Arbres Fruitiers: Balanites Aegyptiaca (L.) Del., Parkiabiglobosa (Jacq.) Benth., Tamarindus Indica L et Zizyphus Mauritiana Lam. Sci. Changements Planét./Secher. 2001, 12, 121–127. [Google Scholar]
- Destinoble, A. Effet de la Symbiose Mycorhizienne Arbusculaire sur la Composition Minérale du Poireau Cultivé en Présence de Carbonate de Calcium. Master’s Thesis, Laval University, Quebec, QC, Canada, 2017; 90p. [Google Scholar]
- Subramanian, K.S.; Charest, C. Nutritional, Growth, and Reproductive Responses of Maize ( Zea Mays L.) to Arbuscular Mycorrhizal Inoculation during and after Drought Stress at Tasselling. Mycorrhiza 1997, 7, 25–32. [Google Scholar] [CrossRef]
- Jesus, E.d.C.; Schiavo, J.A.; de Faria, S.M. Dependência de micorrizas para a nodulação de leguminosas arbóreas tropicais. Rev. Árvore 2005, 29, 545–552. [Google Scholar] [CrossRef]
- Quiroga, G.; Erice, G.; Aroca, R.; Chaumont, F.; Ruiz-Lozano, J.M. Contribution of the Arbuscular Mycorrhizal Symbiosis to the Regulation of Radial Root Water Transport in Maize Plants under Water Deficit. Environ. Exp. Bot. 2019, 167, 103821. [Google Scholar] [CrossRef]
- Wu, Q.; Xia, R. Effects of Arbuscular Mycorrhizal Fungi on Leaf Solutes and Root Absorption Areas of Trifoliate Orange 590 Seedlings under Water Stress Conditions. Front. For. China 2006, 1, 312–317. [Google Scholar] [CrossRef]
- Farahani, H.A.; Lebaschi, M.H.; Hamidi, A. Effects of Arbuscular Mycorrhizal Fungi, Phosphorus and Water Stress on Quantity and Quality Characteristics of Coriander. Adv. Nat. Appl. Sci. 2008, 2, 55–60. [Google Scholar]
- Montaño, N.M.; Alarcón, A.; Camargo-Ricalde, S.L.; Hernández-Cuevas, L.V.; Álvarez-Sánchez, J.; González-Chávez, M.d.C.A.; Gavito, M.E.; Sánchez-Gallen, I.; Ramos-Zapata, J.; Guadarrama, P.; et al. Research on Arbuscular Mycorrhizae in Mexico: An Historical Synthesis and Future Prospects. Symbiosis 2012, 57, 111–126. [Google Scholar] [CrossRef]
- de Azevedo Neto, A.D.; Prisco, J.T.; Enéas-Filho, J.; de Abreu, C.E.B.; Gomes-Filho, E. Effect of Salt Stress on Antioxidative Enzymes and Lipid Peroxidation in Leaves and Roots of Salt-Tolerant and Salt-Sensitive Maize Genotypes. Environ. Exp. Bot. 2006, 56, 87–94. [Google Scholar] [CrossRef]
- Abdul Jaleel, C.; Gopi, R.; Sankar, B.; Manivannan, P.; Kishorekumar, A.; Sridharan, R.; Panneerselvam, R. Studies on Germination, Seedling Vigour, Lipid Peroxidation and Proline Metabolism in Catharanthus Roseus Seedlings under Salt Stress. S. Afr. J. Bot. 2007, 73, 190–195. [Google Scholar] [CrossRef] [Green Version]
- Marulanda, A.; Porcel, R.; Barea, J.M.; Azcón, R. Drought Tolerance and Antioxidant Activities in Lavender Plants Colonized by Native Drought-Tolerant or Drought-Sensitive Glomus Species. Microbiol. Ecol. 2007, 54, 543–552. [Google Scholar] [CrossRef]
- Koide, R.T.; Kabir, Z. Extraradical Hyphae of the Mycorrhizal Fungus Glomus Intraradices Can Hydrolyse Organic Phosphate. New Phytol. 2000, 148, 511–517. [Google Scholar] [CrossRef]
- Benito, B.; González-Guerrero, M. Unravelling Potassium Nutrition in Ectomycorrhizal Associations. New Phytol. 2014, 201, 707–709. [Google Scholar] [CrossRef] [PubMed]
- Scheloske, S.; Maetz, M.; Schneider, T.; Hildebrandt, U.; Bothe, H.; Povh, B. Element Distribution in Mycorrhizal and Nonmycorrhizal Roots of the Halophyte Aster Tripolium Determined by Proton Induced X-ray Emission. Protoplasma 2004, 223, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Hashem, A.; Alqarawi, A.A.; Radhakrishnan, R.; Al-Arjani, A.-B.F.; Aldehaish, H.A.; Egamberdieva, D.; Allah, E.A. Arbuscular mycorrhizal fungi regulate the oxidative system, hormones and ionic equilibrium to trigger salt stress tolerance in Cucumis sativus L. Saudi J. Biol. Sci. 2018, 25, 1102–1114. [Google Scholar] [CrossRef] [PubMed]
- Perner, H.; Schwarz, D.; Bruns, C.; Mäder, P.; George, E. Effect of Arbuscular Mycorrhizal Colonization and Two Levels of Compost Supply on Nutrient Uptake and Flowering of Pelargonium Plants. Mycorrhiza 2007, 17, 469–474. [Google Scholar] [CrossRef]
- Baslam, M.; Esteban, R.; García-Plazaola, J.I.; Goicoechea, N. Effectiveness of Arbuscular Mycorrhizal Fungi (AMF) for Inducing the Accumulation of Major Carotenoids, Chlorophylls and Tocopherol in Green and Red Leaf Lettuces. Appl. Microbiol. Biotechnol. 2013, 97, 3119–3128. [Google Scholar] [CrossRef]
- Boutasknit, A.; Baslam, M.; Ait-El-Mokhtar, M.; Anli, M.; Ben-Laouane, R.; Douira, A.; El Modafar, C.; Mitsui, T.; Wahbi, S.; Meddich, A. Arbuscular Mycorrhizal Fungi Mediate Drought Tolerance and Recovery in Two Contrasting Carob (Ceratonia siliqua L.) Ecotypes by Regulating Stomatal, Water Relations, and (In)Organic Adjustments. Plants 2020, 9, 80. [Google Scholar] [CrossRef] [Green Version]
- Olsson, P.A.; Hammer, E.C.; Pallon, J.; van Aarle, I.M.; Wallander, H. Elemental Composition in Vesicles of an Arbuscular Mycorrhizal Fungus, as Revealed by PIXE Analysis. Fungal Biol. 2011, 115, 643–648. [Google Scholar] [CrossRef]
- Jung, N.C.; Tamai, Y. Polyphosphate (Phytate) Formation in Quercus Acutissima-Scleroderma Verrucosum Ectomycorrhizae Supplied with Phosphate. J. Plant Interact. 2013, 8, 291–303. [Google Scholar] [CrossRef]
- Garcia, K.; Zimmermann, S.D. The Role of Mycorrhizal Associations in Plant Potassium Nutrition. Front. Plant Sci. 2014, 5, 337. [Google Scholar] [CrossRef] [Green Version]
- Al-Karaki, G.N. The Role of Mycorrhiza in the Reclamation of Degraded Lands in Arid Environments. In Developments in Soil Classification, Land Use Planning and Policy Implications; Shahid, S., Taha, F., Abdelfattah, M., Eds.; Springer: Dordrecht, The Netherlands, 2013; pp. 823–836. [Google Scholar] [CrossRef]
- Laxa, M.; Liebthal, M.; Telman, W.; Chibani, K.; Dietz, K.-J. The Role of the Plant Antioxidant System in Drought Tolerance. Antioxidants 2019, 8, 94. [Google Scholar] [CrossRef] [Green Version]
- Slama, F. Effet Du NaCl Sur La Croissance et La Nutrition Minérale de Six Espèces de Plantes Cultivées. Agrochimica 1986, 30, 137–147. [Google Scholar]
- Zhu, X.C.; Song, F.B.; Liu, S.Q.; Liu, T.D.; Zhou, X. Arbuscular Mycorrhizae Improves Photosynthesis and Water Status of Zea mays L. under Drought Stress. Plant Soil Environ. 2012, 58, 186–191. [Google Scholar] [CrossRef] [Green Version]
- Zaouchi, Y.; Rezgui, S.; Bettaieb, T. Influence of Mycorrhization on Adaptation Capacity of Jacaranda Mimosifolia D. Don Grown in Urban Conditions. J. New Sci. 2015, 18, 679–688. [Google Scholar]
- Zhang, Y.; Zhong, C.L.; Chen, Y.; Chen, Z.; Jiang, Q.B.; Wu, C.; Pinyopusarerk, K. Improving Drought Tolerance of Casuarina Equisetifolia Seedlings by Arbuscular Mycorrhizas under Glasshouse Conditions. New For. 2010, 40, 261–271. [Google Scholar] [CrossRef]
- ElYacoubi, H.; Ayolié, K.; Rochdi, A. In Vitro Cellular Salt Tolerance of Troyer Citrange: Changes in Growth and Solutes Accumulation in Callus Tissue. Int. J. Agric. Biol. 2010, 12, 187–193. [Google Scholar]
- Wu, Q.-S.; Xia, R.-X. Arbuscular Mycorrhizal Fungi Influence Growth, Osmotic Adjustment and Photosynthesis of Citrus under Well-Watered and Water Stress Conditions. J. Plant Physiol. 2006, 163, 417–425. [Google Scholar] [CrossRef]
- Pavithra, D.; Yapa, N. Arbuscular Mycorrhizal Fungi Inoculation Enhances Drought Stress Tolerance of Plants. Groundw. Sustain. Dev. 2018, 7, 490–494. [Google Scholar] [CrossRef]
- Porcel, R.; Ruiz-Lozano, J.M. Arbuscular Mycorrhizal Influence on Leaf Water Potential, Solute Accumulation, and Oxidative Stress in Soybean Plants Subjected to Drought Stress. J. Exp. Bot. 2004, 55, 1743–1750. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.-S.; Xia, R.-X.; Zou, Y.-N.; Wang, G.-Y. Osmotic Solute Responses of Mycorrhizal Citrus (Poncirus Trifoliata) Seedlings to Drought Stress. Acta Physiol. Plant. 2007, 6, 543–549. [Google Scholar] [CrossRef]
- Subramanian, K.S.; Charest, C. Influence of Arbuscular Mycorrhizae on the Metabolism of Maize under Drought Stress. Mycorrhiza 1995, 5, 273–278. [Google Scholar] [CrossRef]
- Lu, J.Y.; Liu, M.; Mao, Y.M.; Shen, L.Y. Effects of Vesicular-Arbuscular Mycorrhizae on the Drought Resistance of Wild Jujube (Zizyphs Spinosus Hu) Seedlings. Front. Agric. China 2007, 1, 468–471. [Google Scholar] [CrossRef]
- Irigoyen, J.J.; Einerich, D.W.; Sánchez-Díaz, M. Water Stress Induced Changes in Concentrations of Proline and Total Soluble Sugars in Nodulated Alfalfa (Medicago Sativd) Plants. Physiol. Plant. 1992, 84, 55–60. [Google Scholar] [CrossRef]
- Yooyongwech, S.; Phaukinsang, N.; Cha-um, S.; Supaibulwatana, K. Arbuscular Mycorrhiza Improved Growth Performance in Macadamia Tetraphylla L. Grown under Water Deficit Stress Involves Soluble Sugar and Proline Accumulation. Plant Growth Regul. 2013, 69, 285–293. [Google Scholar] [CrossRef]
- Muthukumar, T.; Udaiyan, K. Arbuscular Mycorrhizal Fungal Composition in Semi-Arid Soils of Western Ghats, Southern 678 India. Curr. Sci. 2002, 82, 624–628. [Google Scholar]
- Talaat, N.B.; Shawky, B.T. Influence of Arbuscular Mycorrhizae on Yield, Nutrients, Organic Solutes, and Antioxidant Enzymes of Two Wheat Cultivars under Salt Stress. J. Plant Nutr. Soil Sci. 2011, 174, 283–291. [Google Scholar] [CrossRef]
- Ruiz-Lozano, J.M.; Porcel, R.; Azcón, C.; Aroca, R. Regulation by Arbuscular Mycorrhizae of the Integrated Physiological Response to Salinity in Plants: New Challenges in Physiological and Molecular Studies. J. Exp. Bot. 2012, 63, 4033–4044. [Google Scholar] [CrossRef] [Green Version]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Station | Coordination | Province | Altitude | Average Annual Rainfall (mm) | Annual Mean Temperature (°C) | Tree Density/Hectare | Use |
|---|---|---|---|---|---|---|---|
| Bouyzakarne | N: 29.1879 W: 009.7421 | Goulmim Es-smara | 690 | 120 | 21 | 5 | Pasture |
| Argana | N: 30.7544 W: 009.1545 | Taroudante | 735 | 500 | 20.9 | 30 | Pasture |
| Source of Variation | df | Proline (µg/g) | Sugar (µmol/g) | K+ (mg/g) | Na+ (mg/g) | Ratio K/Na | Phosphorus (mg/g) | Fresh Matter (g) | Dry Matter (g) | |
| Inoculum | 2 | 1094.11 *** | 4854.55 *** | 24.29 *** | 4.63 * | 0.97 ** | 97.52 *** | 315.09 *** | 58.29 *** | |
| Duration (D) | 3 | 26,295.61 *** | 15,808.79 *** | 514.83 *** | 814.34 *** | 11.54 *** | 60.00 *** | 269.06 *** | 49.83 *** | |
| Plant Part (PP) | 1 | 4232.00 *** | 6610.39 *** | 3493.50 *** | 1328.96 *** | 7.45 *** | 38.75 *** | 353.82 *** | 6.28 *** | |
| Inoculum*Duration | 6 | 2437.89 *** | 15.24 ns | 1.61 ns | 11.23 * | 0.41 ns | 0.77 ns | 4.60 *** | 1.81 *** | |
| Inoculum*PP | 2 | 2413.00 *** | 1825.16 *** | 1.81 * | 18.60 *** | 0.35 ns | 0.27 ns | 45.31 *** | 8.44 *** | |
| D*PP | 3 | 9422.11 *** | 3271.16 *** | 73.04 *** | 2123.73 *** | 110.55 *** | 6.01 *** | 8.34 *** | 2.95 *** | |
| Model | 17 | 45,894.72 *** | 32,385.28 *** | 4109.09 *** | 4301.49 *** | 131.26 *** | 203.32 *** | 996.22 *** | 127.60 *** | |
| Error | 54 | 2371.89 | 1225.20 | 14.97 | 35.10 | 3.62 | 4.06 | 7.26 | 2.77 | |
| C. Total | 71 | 48,266.61 | 33,610.48 | 4124.05 | 4336.59 | 134.87 | 207.37 | 1003.48 | 130.37 | |
| Source of Variation | df | Ratio FM (A/R) | Ratio DM (A/R) | Chlorophyll a (mg/L) | Chlorophyll b (mg/L) | Ratio Chlorophyll a/b | Ψb (bar) | Ψf (bar) | ||
| Inoculum | 2 | 0.1014 *** | 0.5024 *** | 13.72 ** | 16.06 *** | 0.0524 ns | 33.50 *** | 50.89 *** | ||
| Duration (D) | 3 | 0.1899 *** | 0.2587 *** | 302.83 *** | 195.33 *** | 1.4910 *** | 746.33 *** | 715.67 *** | ||
| Inoculum*D | 6 | 0.0208 *** | 0.0349 * | 12.54 ** | 14.83 *** | 0.0515 ns | 9.83 * | 3.33 ns | ||
| Model | 11 | 0.3122 *** | 0.7960 *** | 329.09 *** | 226.22 *** | 1.5949 *** | 789.67 *** | 769.89 *** | ||
| Error | 24 | 0.0073 | 0.0426 | 17.84 | 10.67 | 0.7603 | 13.33 | 6.00 | ||
| C. Total | 35 | 0.3195 | 0.8386 | 346.93 | 236.89 | 2.3552 | 803.00 | 775.89 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ouallal, I.; Abbas, Y.; ElYacoubi, H.; Imtara, H.; Al Zain, M.N.; Ouajdi, M.; El Goumi, Y.; Alzamel, N.M.; Mohammed Noman, O.; Rochdi, A. Effects of Arbuscular Mycorrhizal Inoculation by Indigenous Fungal Complexes on the Morpho-Physiological Behavior of Argania spinosa Subjected to Water Deficit Stress. Horticulturae 2022, 8, 280. https://doi.org/10.3390/horticulturae8040280
Ouallal I, Abbas Y, ElYacoubi H, Imtara H, Al Zain MN, Ouajdi M, El Goumi Y, Alzamel NM, Mohammed Noman O, Rochdi A. Effects of Arbuscular Mycorrhizal Inoculation by Indigenous Fungal Complexes on the Morpho-Physiological Behavior of Argania spinosa Subjected to Water Deficit Stress. Horticulturae. 2022; 8(4):280. https://doi.org/10.3390/horticulturae8040280
Chicago/Turabian StyleOuallal, Imane, Younes Abbas, Houda ElYacoubi, Hamada Imtara, Mashail N. Al Zain, Mohemed Ouajdi, Younes El Goumi, Nurah M. Alzamel, Omar Mohammed Noman, and Atmane Rochdi. 2022. "Effects of Arbuscular Mycorrhizal Inoculation by Indigenous Fungal Complexes on the Morpho-Physiological Behavior of Argania spinosa Subjected to Water Deficit Stress" Horticulturae 8, no. 4: 280. https://doi.org/10.3390/horticulturae8040280