
Vanadium Toxicity Induced Changes in Growth, Antioxidant Profiling, and Vanadium Uptake in Pepper (Capsicum annum L.) Seedlings

 ,
, 
Abstract
:1. Introduction
2. Material and Methods
2.1. Pepper Seed, Reagent, and Growth Conditions
2.2. Seedling Growth and Treatment
2.3. Growth Variables
2.4. Root Morphology
2.5. Gas Exchange Parameters and SPAD Index
2.6. Photosynthetic Pigments
2.7. Malonaldehyde (MDA) and Hydrogen Peroxide (H2O2), Catalase (CAT), and Superoxide Dismutase (SOD) Measurement
2.8. Proline and Soluble Sugars Content
2.9. V Determination, Uptake and Translocation
2.10. Statistical Analysis
3. Results
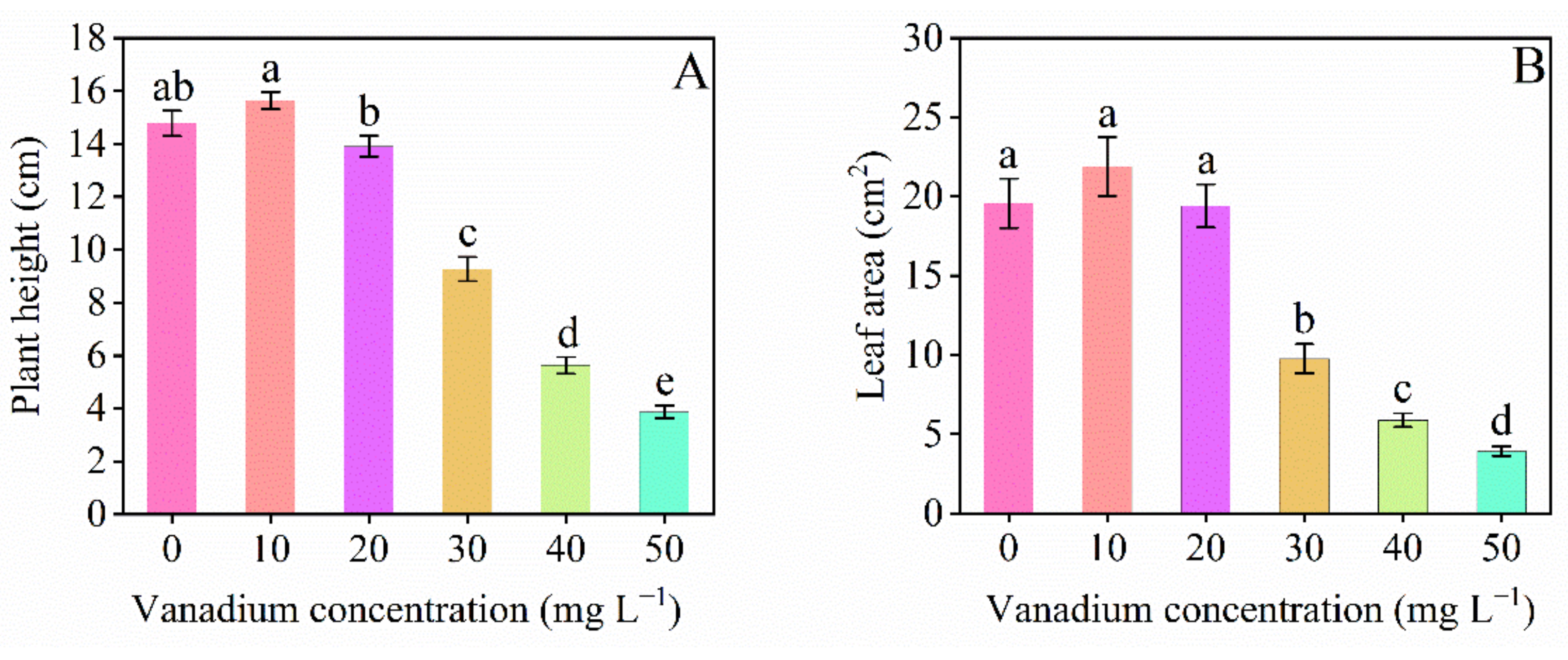
3.1. Plant Growth and Growth Tolerance Index
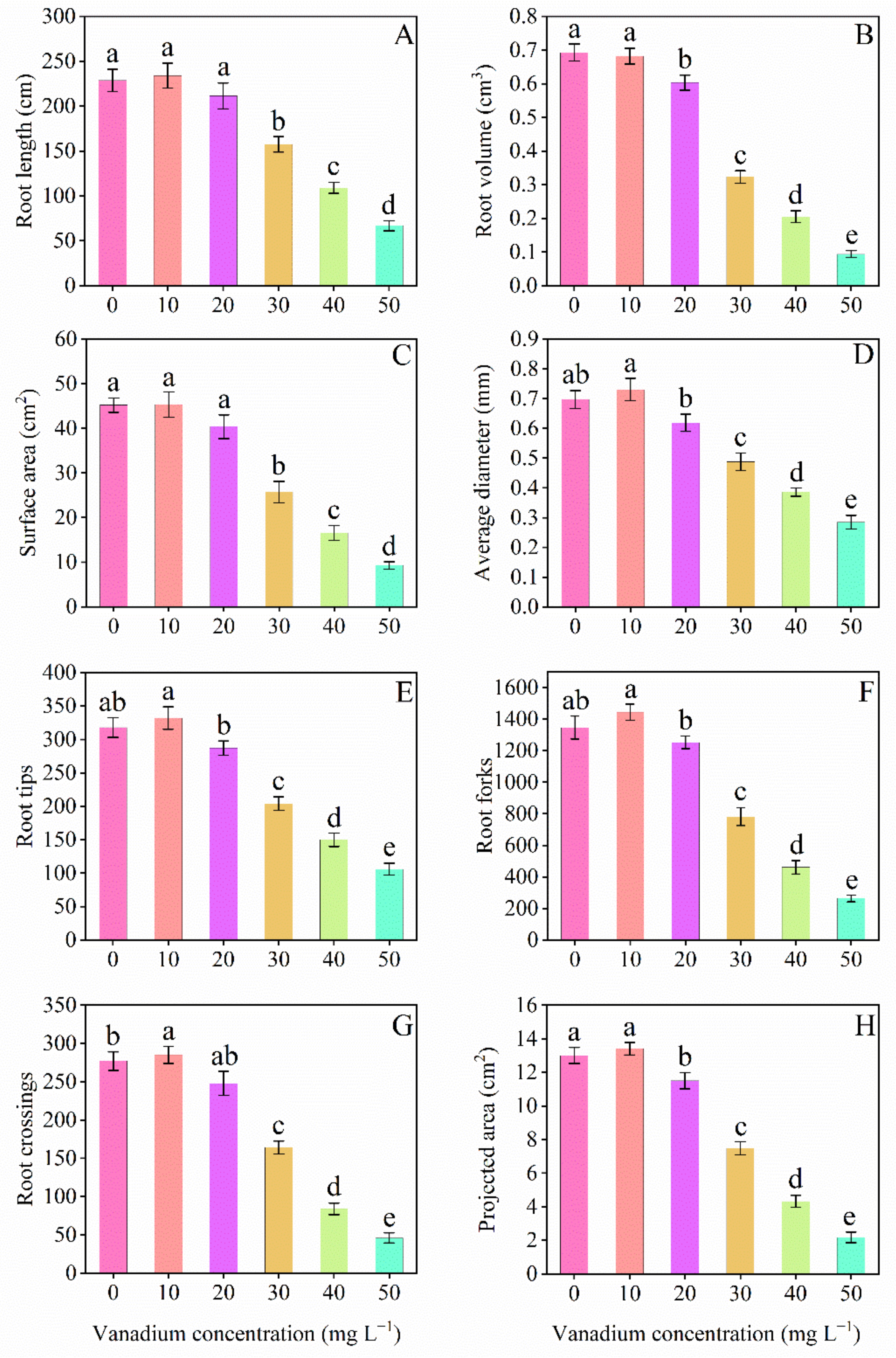
3.2. Root Morphology
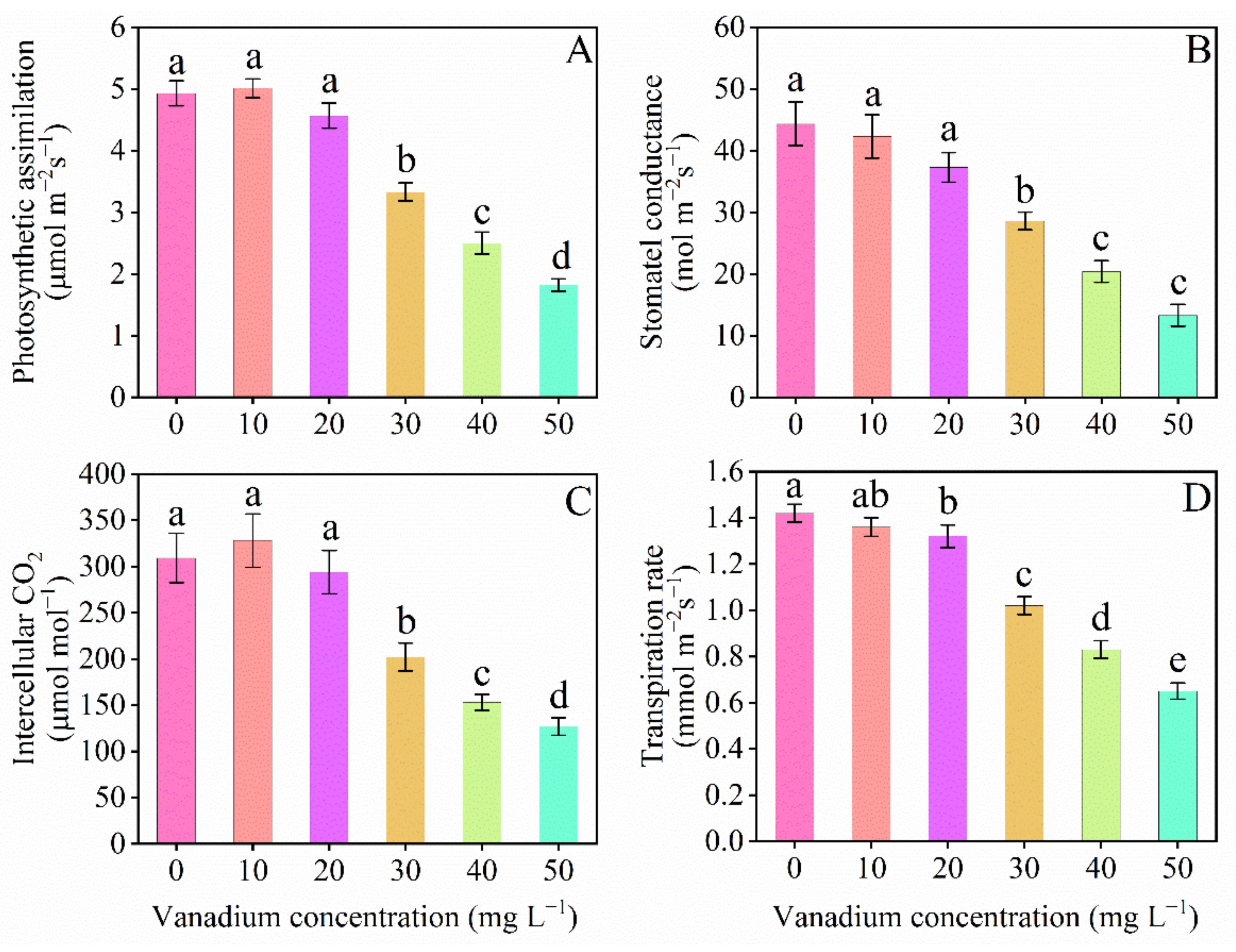
3.3. Leaf Gas Exchange Elements
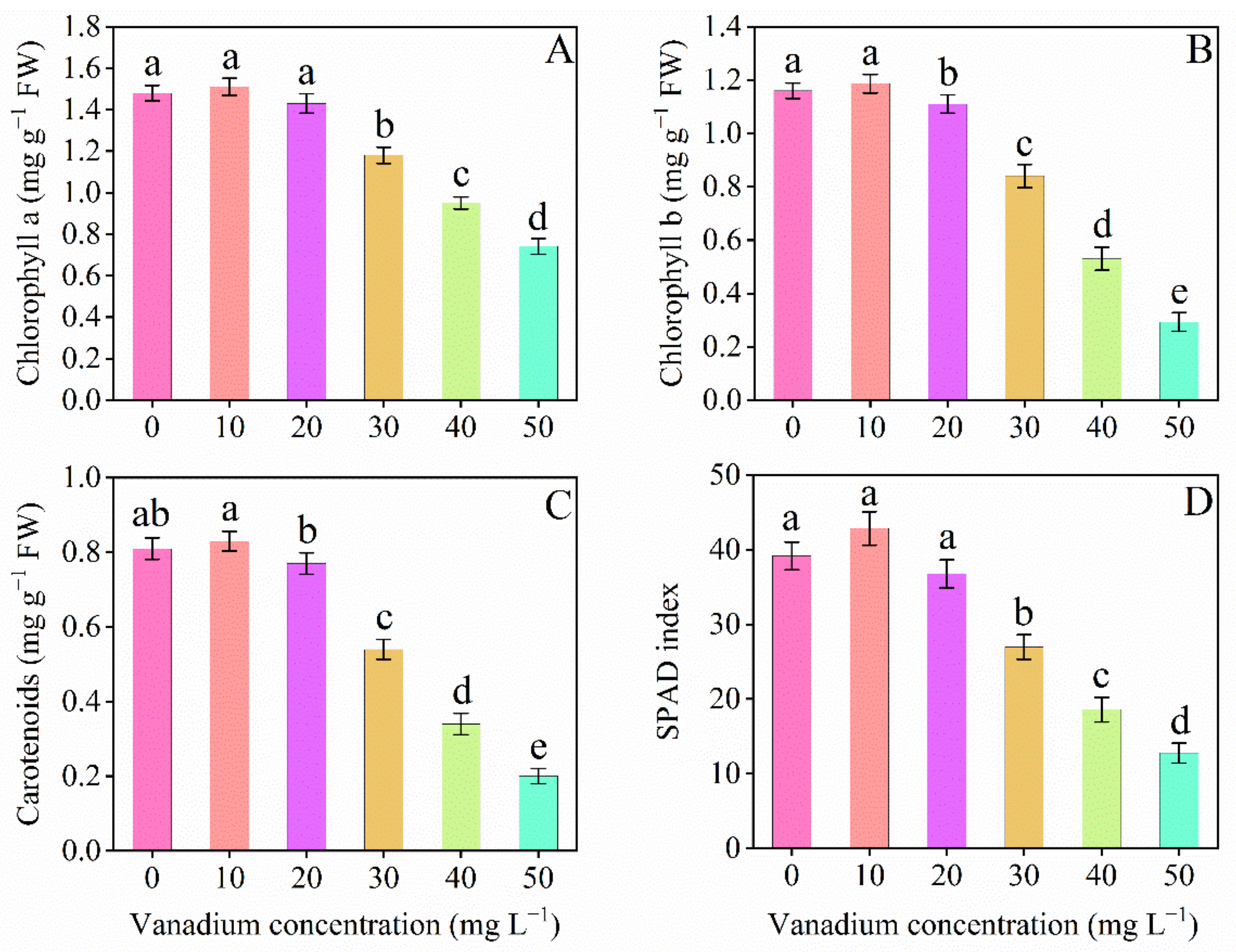
3.4. SPAD Index and Pigments Content
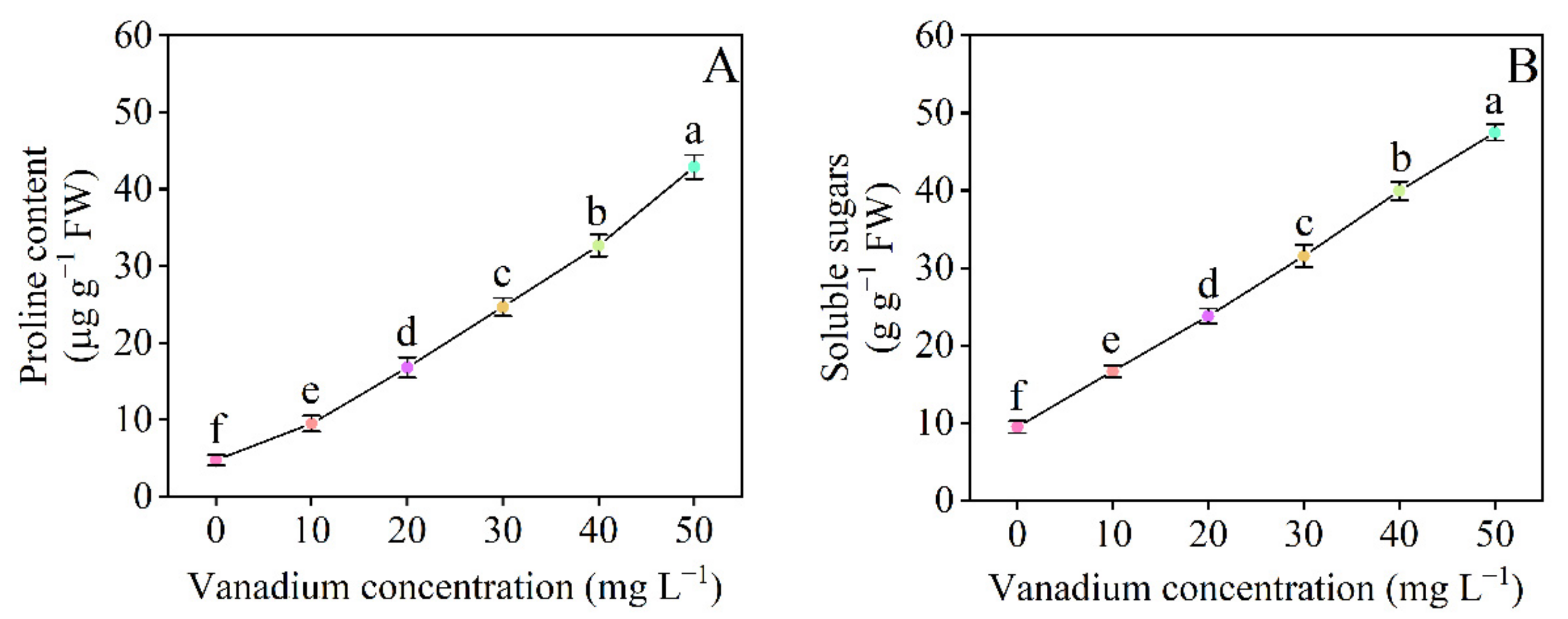
3.5. Proline and Soluble Sugar Content
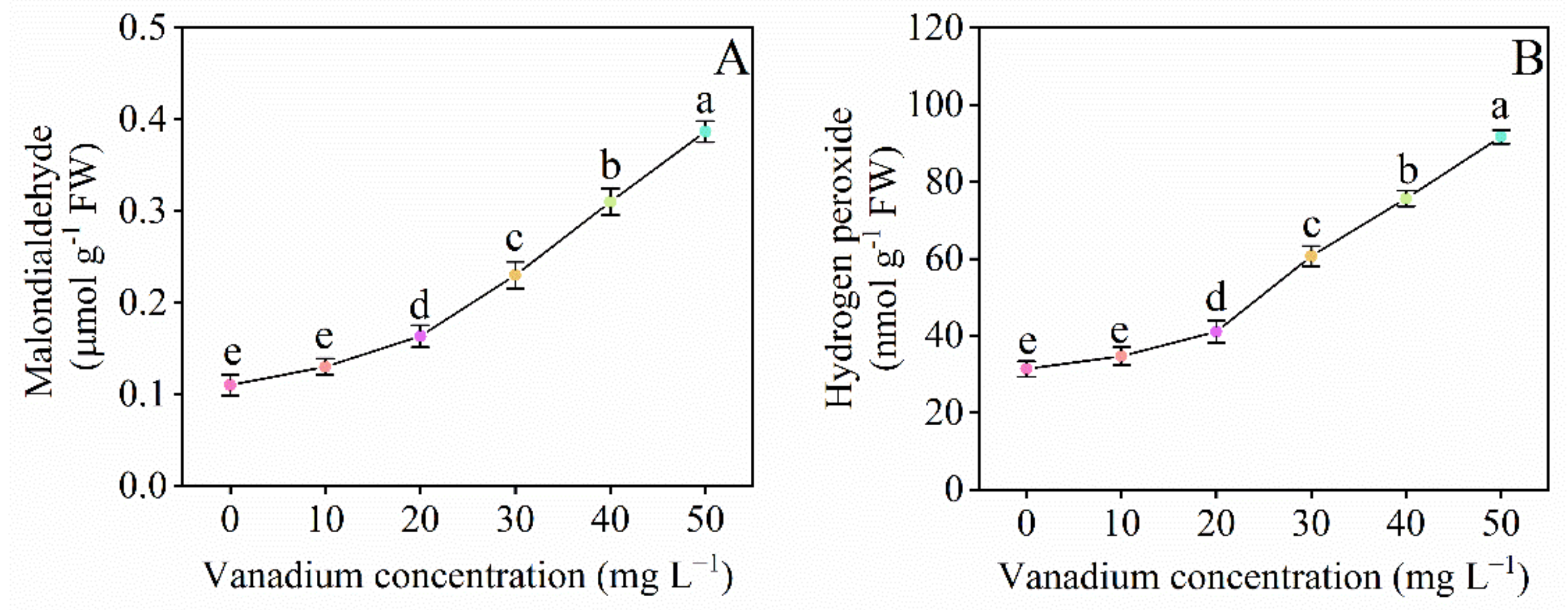
3.6. Malonaldehyde and Hydrogen Peroxide Content
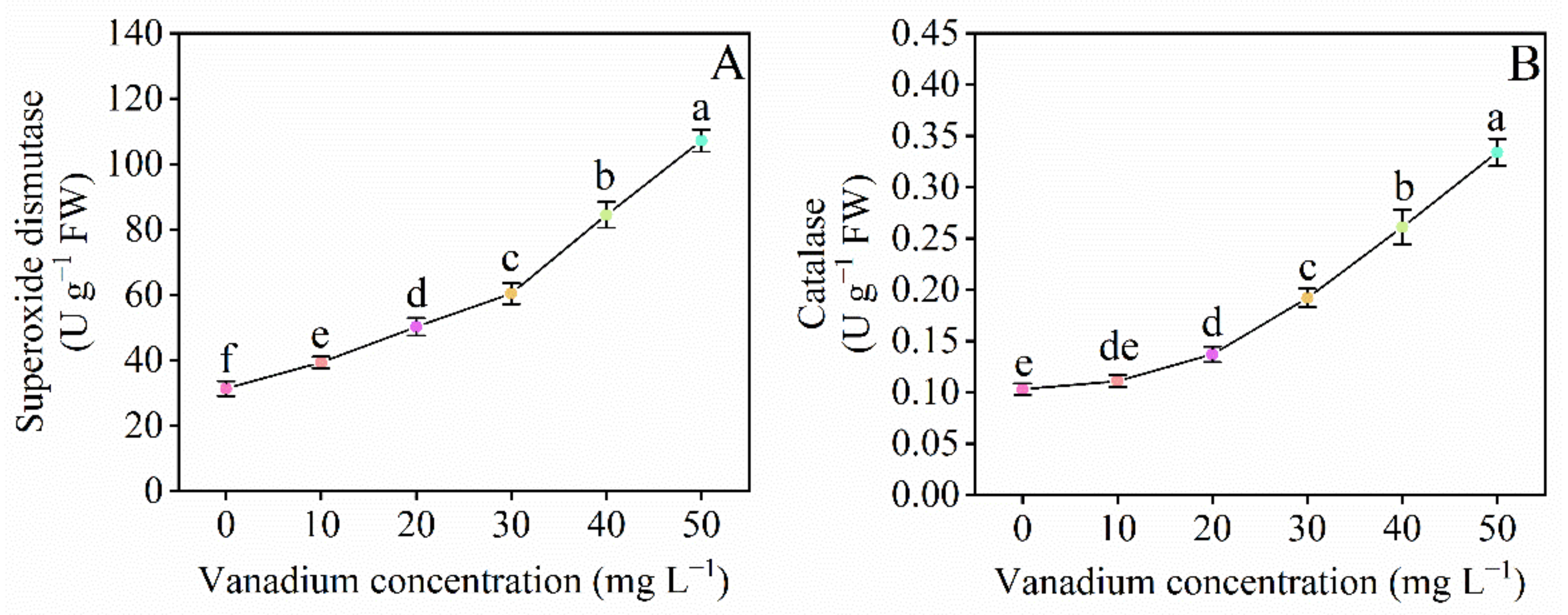
3.7. Catalase and Superoxide Dismutase
3.8. Plant Tissues Vanadium Concentration, Uptake, and Root to Shoot Translocation
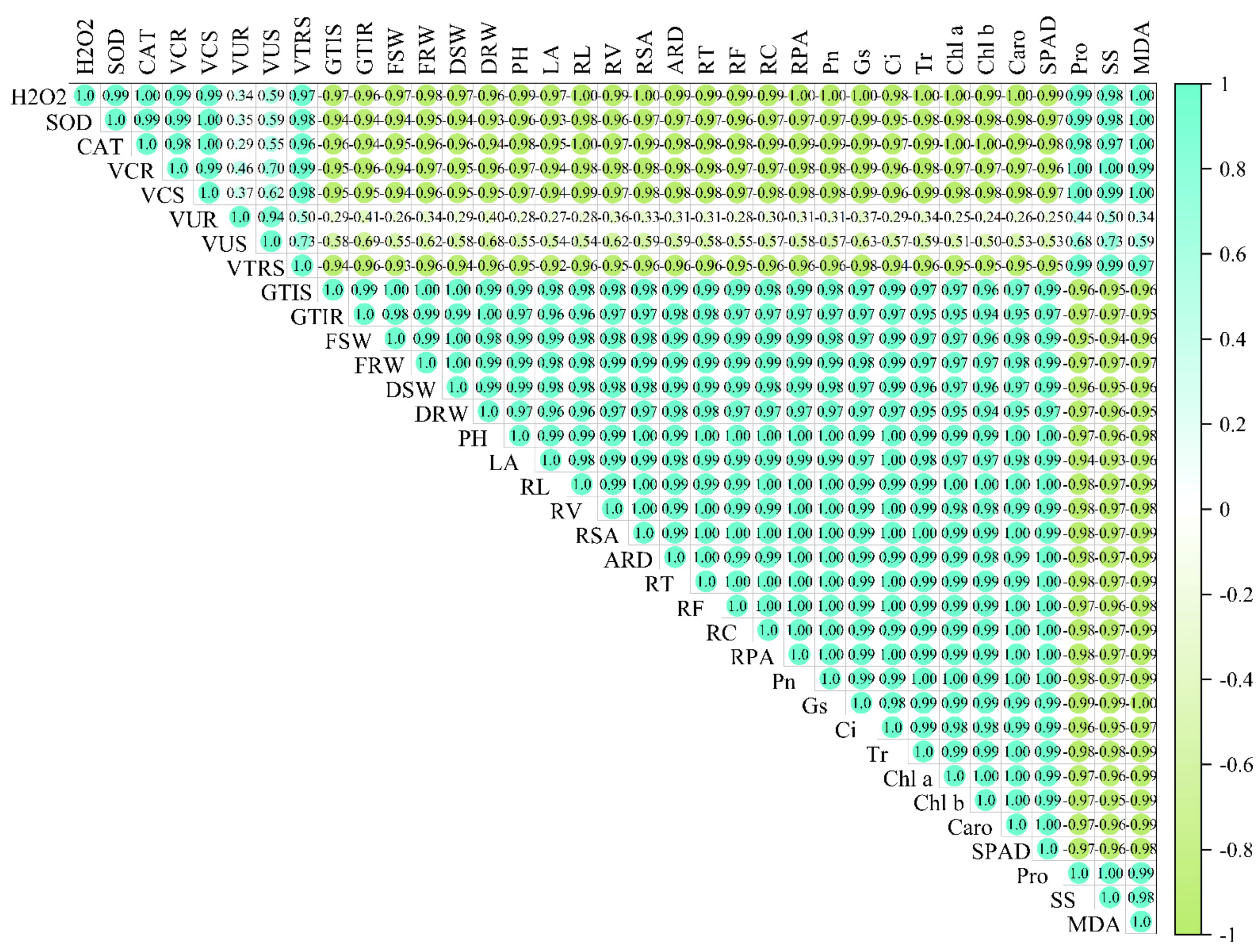
3.9. Pearson’s Correlation Coefficient
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Harmanescu, M.; Alda, L.M.; Bordean, D.M.; Gogoasa, I.; Gergen, I. Heavy metals health risk assessment for population via consumption of vegetables grown in old mining area; a case study: Banat County Romania. Chem. Cent. J. 2011, 5, 64. [Google Scholar] [CrossRef] [Green Version]
- Imtiaz, M.; Mushtaq, M.A.; Nawaz, M.A.; Ashraf, M.; Rizwan, M.S.; Mehmood, S.; Aziz, O.; Rizwan, M.; Virk, M.S.; Shakeel, Q.; et al. Physiological and anthocyanin biosynthesis genes response induced by vanadium stress in mustard genotypes with distinct photosynthetic activity. Environ. Toxicol. Pharm. 2018, 62, 20–29. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Tang, Y.; Yang, K.; Rouff, A.A.; Elzinga, E.J.; Huang, J.H. Leaching characteristics of vanadium in mine tailings and soils near a vanadium titanomagnetite mining site. J. Hazard. Mater. 2014, 264, 498–504. [Google Scholar] [CrossRef]
- Tian, L.Y.; Yang, J.Y.; Huang, J.H. Uptake and speciation of vanadium in the rhizosphere soils of rape (Brassica juncea L.). Environ. Sci. Pollut. Res. 2015, 22, 9215–9223. [Google Scholar] [CrossRef] [PubMed]
- Witz, G.; Osmond, F. Essai sur l’application des propriete’s de l’oxycellulose au dosage du vanadium. Bull. Soc. Chim. 1986, 2, 309–314. [Google Scholar] [CrossRef]
- Bertrand, G. The biogeochemistry of vanadium. Bull. Am. Mus. Nat. Hist. 1950, 94, 409–455. [Google Scholar]
- Larsson, M.A.; Baken, S.; Gustafsson, J.P.; Hadialhejazi, G.; Smolders, E. Vanadium bioavailability and toxicity to soil microorganisms and plants. Environ. Toxicol. Chem. 2013, 32, 2266–2273. [Google Scholar] [CrossRef] [Green Version]
- Imtiaz, M.; Rizwan, M.S.; Xiong, S.; Li, H.; Ashraf, M.; Shahzad, S.M.; Shahzad, M.; Rizwan, M.; Tu, S. Vanadium, recent advancements and research prospects: A review. Environ. Int. 2015, 80, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Olness, A.; Gesch, R.; Forcella, F.; Archer, D.; Rinke, J. Importance of vanadium and nutrient ionic ratios on the development of hydroponically grown cuphea. Ind. Crops Prod. 2005, 21, 165–171. [Google Scholar] [CrossRef]
- García-Jiménez, A.; Trejo-Téllez, L.I.; Guillén-Sánchez, D.; Gómez-Merino, F.C. Vanadium stimulates pepper plant growth and flowering, increases concentrations of amino acids, sugars and chlorophylls, and modifies nutrient concentrations. PLoS ONE 2018, 13, e0201908. [Google Scholar] [CrossRef] [Green Version]
- Vachirapatama, N.; Jirakiattiku, Y.; Dicinoski, G.W.; Townsend, A.T.; Haddad, P.R. Effect of vanadium on plant growth and its accumulation in plant tissues. Songklanakarin J. Sci. Technol. 2011, 33, 255–261. [Google Scholar]
- Imtiaz, M.; Tu, S.; Xie, Z.; Han, D.; Ashraf, M.; Rizwan, M.S. Growth, V uptake, and antioxidant enzymes responses of chickpea (Cicer arietinum L.) genotypes under vanadium stress. Plant Soil. 2015, 390, 17–27. [Google Scholar] [CrossRef]
- Jahan, M.S.; Guo, S.; Sun, J.; Shu, S.; Wang, Y.; Abou El-Yazied, A.; Alabdallah, N.M.; Hikal, M.; Mohamed, M.H.; Ibrahim, M.F.; et al. Melatonin-mediated photosynthetic performance of tomato seedlings under high-temperature stress. Plant Physiol. Biochem. 2021, 167, 309–320. [Google Scholar] [CrossRef] [PubMed]
- Altaf, M.M.; Diao, X.P.; Shakoor, A.; Imtiaz, M.; Altaf, M.A.; Khan, L.U. Delineating Vanadium (V) Ecological Distribution, Its Toxicant Potential, and Effective Remediation Strategies from Contaminated Soils. J. Soil Sci. Plant Nutr. 2021, 1–19. [Google Scholar] [CrossRef]
- Nawaz, M.A.; Jiao, Y.; Chen, C.; Shireen, F.; Zheng, Z.; Imtiaz, M.; Bie, Z.; Huang, Y. Melatonin pretreatment improves vanadium stress tolerance of watermelon seedlings by reducing vanadium concentration in the leaves and regulating melatonin biosynthesis and antioxidant-related gene expression. J. Plant Physiol. 2018, 220, 115–127. [Google Scholar] [CrossRef]
- Li, Q.; Wang, G.; Wang, Y.R.; Yang, D.; Guan, C.F. Foliar application of salicylic acid alleviates the cadmium toxicity by modulation the reactive oxygen species in potato. Ecotoxicol. Environ. Saf. 2019, 172, 317–325. [Google Scholar] [CrossRef]
- Jahan, M.S.; Guo, S.; Baloch, A.R.; Sun, J.; Shu, S.; Wang, Y.; Ahammed, G.J.; Kabir, K.; Roy, R. Melatonin alleviates nickel phytotoxicity by improving photosynthesis, secondary metabolism and oxidative stress tolerance in tomato seedlings. Ecotoxicol. Environ. Saf. 2020, 197, 110593. [Google Scholar] [CrossRef]
- Alyemeni, M.N.; Ahanger, M.A.; Wijaya, L.; Alam, P.; Bhardwaj, R.; Ahmad, P. Selenium mitigates cadmium-induced oxidative stress in tomato (Solanum lycopersicum L.) plants by modulating chlorophyll fluorescence, osmolyte accumulation, and antioxidant system. Protoplasma 2018, 255, 985–986. [Google Scholar] [CrossRef] [Green Version]
- Andresen, E.; Küpper, H. Cadmium toxicity in plants. In Cadmium: From Toxicity to Essentiality; Springer: Dordrecht, The Netherlands, 2013; pp. 395–413. [Google Scholar] [CrossRef]
- Altaf, M.A.; Shahid, R.; Ren, M.X.; Khan, L.U.; Altaf, M.M.; Jahan, M.S.; Nawaz, M.A.; Naz, S.; Shahid, S.; Lal, M.K.; et al. Protective Mechanisms of Melatonin Against Vanadium Phytotoxicity in Tomato Seedlings: Insights into Nutritional Status, Photosynthesis, Root Architecture System, and Antioxidant Machinery. J. Plant Growth Regul. 2021, 1–17. [Google Scholar] [CrossRef]
- Altaf, M.M.; Diao, X.P.; ur Rehman, A.; Imtiaz, M.; Shakoor, A.; Altaf, M.A.; Younis, H.; Fu, P.; Ghani, M.U. Effect of vanadium on growth, photosynthesis, reactive oxygen species, antioxidant enzymes, and cell death of rice. J. Soil Sci. Plant Nutr. 2020, 20, 2643–2656. [Google Scholar] [CrossRef]
- Howard, L.R.; Talcott, S.Y.; Brenes, C.H.; Villalon, B. Changes in phytochemical and antioxidant activity of selected pepper cultivars (Capsicum sp.) as influenced by maturity. J. Agric. Food Chem. 2000, 48, 1713–1720. [Google Scholar] [CrossRef]
- Altaf, M.A.; Shahid, R.; Altaf, M.A.; Ren, M.X.; Tan, K.; Xiang, W.Q.; Qadir, A.; Shakoor, A.; Altaf, M.M. Effect of NPK, organic manure and their combination on growth, yield and nutrient uptake of chilli (Capsicum annum L.). Hortic. Int. J. 2019, 3, 217–222. [Google Scholar] [CrossRef]
- Wiszniewska, A.; Hanus-Fajerska, E.; Muszyńska, E.; Smoleń, S. Comparative assessment of response to cadmium in heavy metal-tolerant shrubs cultured in vitro. Water Air Soil Pollut. 2017, 228, 304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altaf, M.A.; Shahid, R.; Ren, M.X.; Naz, S.; Altaf, M.M.; Qadir, A.; Anwar, M.; Shakoor, A.; Hayat, F. Exogenous melatonin enhances salt stress tolerance in tomato seedlings. Biol. Plant. 2020, 64, 604–615. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, Y.; Wen, W.; Shi, Z.; Gu, Q.; Ahammed, G.J.; Cao, K.; Jahan, M.S.; Shu, S.; Wang, J.; et al. Hydrogen peroxide mediates spermidine induced autophagy to alleviate salt stress in cucumber. Autophagy 2020, 10, 2876–2890. [Google Scholar] [CrossRef] [PubMed]
- Lichtenhaler, K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef] [Green Version]
- Altaf, M.A.; Shahid, R.; Ren, M.X.; Altaf, M.M.; Khan, L.U.; Shahid, S.; Jahan, M.S. Melatonin alleviates salt damage in tomato seedling: A root architecture system, photosynthetic capacity, ion homeostasis, and antioxidant enzymes analysis. Sci. Hortic. 2021, 285, 110145. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.T.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Amjad, M.; Raza, H.; Murtaza, B.; Abbas, G.; Imran, M.; Shahid, M.; Asif Naeem, M.; Zakir, A.; Mohsin Iqbal, M.M. Nickel toxicity induced changes in nutrient dynamics and antioxidant profiling in two Maize (Zea mays L.) Hybrids. Plants 2020, 9, 5. [Google Scholar] [CrossRef] [Green Version]
- Heidari, J.; Amooaghaie, R.; Kiani, S. Impact of chitosan on nickel bioavailability in soil, the accumulation and tolerance of nickel in Calendula tripterocarpa. Int. J. Phytoremediat. 2020, 22, 1175–1184. [Google Scholar] [CrossRef] [PubMed]
- Pilon-Smits, A.H.E.; Quinn, F.C.; Tapken, W.; Malagoli, M.; Schiavon, M. Physiological functions of beneficial elements. Curr. Opin Plant Biol. 2009, 12, 267–274. [Google Scholar] [CrossRef]
- Tang, Y.T.; Qiu, R.L.; Zeng, X.W.; Ying, R.R.; Yu, F.M.; Zhou, X.Y. Lead, zinc, cadmium hyperaccumulation and growth stimulation in Arabis paniculate Franch. Environ. Exp. Bot. 2009, 66, 126–134. [Google Scholar] [CrossRef]
- Shafeeq-ur-Rahman; Qi, X.; Xiao, Y.; Muhammad, I.A.; Muhammad, S.; Muhammad, Z. Silicon and its application methods improve physiological traits and antioxidants in Triticum aestivum (L.) under cadmium stress. J. Soil Sci. Plant Nutr. 2020, 20, 1110–1121. [Google Scholar] [CrossRef]
- Liang, Y.C.; Sun, W.C.; Zhu, Y.G.; Christie, P. Mechanisms of silicon-mediated alleviation of abiotic stresses in higher plants: A review. Environ. Pollut. 2007, 147, 422–428. [Google Scholar] [CrossRef] [Green Version]
- Wissuwa, M.; Kretzschmar, T.; Rose, T.J. From promise to application: Root traits for enhanced nutrient capture in rice breeding. J. Exp. Bot. 2016, 67, 3605–3615. [Google Scholar] [CrossRef] [Green Version]
- Altaf, M.A.; Shahid, R.; Ren, M.X.; Altaf, M.M.; Jahan, M.S.; Khan, L.U. Melatonin Mitigates Nickel Toxicity by Improving Nutrient Uptake Fluxes, Root Architecture System, Photosynthesis, and Antioxidant Potential in Tomato Seedling. J. Soil Sci. Plant Nutr. 2021, 21, 1842–1855. [Google Scholar] [CrossRef]
- Meisch, H.U.; Benzschawel, H.; Bielig, H.J. The role of vanadium in green plants. Arch. Microbiol. 1977, 114, 67–70. [Google Scholar] [CrossRef]
- Cai, S.Y.; Zhang, Y.; Xu, Y.P.; Qi, Z.Y.; Li, M.Q.; Ahammed, G.J.; Xia, X.J.; Shi, K.; Zhou, Y.H.; Reiter, R.J.; et al. HsfA1a upregulates melatonin biosynthesis to confer cadmium tolerance in tomato plants. J. Pineal Res. 2017, 62, e12387. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.Y.; Qi, C.D.; Li, S.; Wang, Z.; Wang, X.; Wang, J.; Ren, S.; Li, X.; Zhang, N.; Guo, Y.D. Melatonin alleviates copper toxicity via improving copper sequestration and ROS scavenging in cucumber. Plant Cell Physiol. 2019, 60, 562–574. [Google Scholar] [CrossRef] [PubMed]
- Yu-chen, G.; Liu, Y.Y.; Wang, R.Y.; Wang, S.; Lu, X.P.; Wang, B. Effect of mercury stress on Khudsar photosynthetic characteristics of two kinds of warm season turf grass. Int. J. Environ. Monit. Anal. 2015, 3, 293–297. [Google Scholar] [CrossRef]
- Hasan, M.; Ahammed, G.J.; Yin, L.; Shi, K.; Xia, X.; Zhou, Y.; Yu, J.; Zhou, J. Melatonin mitigates cadmium phytotoxicity through modulation of phytochelatins biosynthesis, vacuolar sequestration, and antioxidant potential in Solanum lycopersicum L. Front. Plant Sci. 2015, 6, 601. [Google Scholar] [CrossRef]
- Abedini, M.; Mohammadian, F. Vanadium effects on phenolic content and photosynthetic pigments of sunflower. South West J. Hortic. Biol. Environ. 2018, 9, 77–86. [Google Scholar]
- Sarafi, E.; Tsouvaltzis, P.; Chatzissavvidis, C.; Siomos, A.; Therios, I. Melatonin and resveratrol reverse the toxic effect of high boron (B) and modulate biochemical parameters in pepper plants (Capsicum annuum L.). Plant Physiol. Biochem. 2017, 112, 173–182. [Google Scholar] [CrossRef]
- Hasan, M.K.; Liu, C.X.; Pan, Y.T.; Ahammed, G.J.; Qi, Z.Y.; Zhou, J. Melatonin alleviates low-sulfur stress by promoting sulfur homeostasis in tomato plants. Sci. Rep. 2018, 8, 10182. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, M.H.; Alamri, S.; Alsubaie, Q.D.; Ali, H.M.; Ibrahim, A.A.; Alsadon, A. Potential roles of melatonin and sulfur in alleviation of lanthanum toxicity in tomato seedlings. Ecotoxicol. Environ. Saf. 2019, 180, 656–667. [Google Scholar] [CrossRef] [PubMed]
- Aihemaiti, A.; Jiang, J.; Blaney, L. The detoxification effect of liquid digestate on vanadium toxicity to seed germination and seedling growth of dog’s tail grass. J. Hazard. Mater. 2019, 369, 456–464. [Google Scholar] [CrossRef] [PubMed]
- Chary, N.S.; Kamala, C.; Raj, D.S.S. Assessing risk of heavy metals from consuming food grown on sewage irrigated soils and food chain transfer. Ecotoxicol. Environ. Saf. 2008, 69, 513–524. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, S.; Ansari, M.; Aslam, R. Sequential effects of cadmium on plant growth, biochemical and cyto-physiological aspects, antioxidant activity, and molecular screening of protein by SDS-PAGE in Trigonella. Toxicol. Environ. Chem. 2012, 94, 1557–1570. [Google Scholar] [CrossRef]
- Imtiaz, M.; Rizwan, M.S.; Mushtaq, M.A.; Yousaf, B.; Ashraf, M.; Ali, M.; Yousuf, A.; Rizwan, M.; Din, M.; Dai, Z. Interactive effects of vanadium and phosphorus on their uptake, growth and heat shock proteins in chickpea genotypes under hydroponic conditions. Environ. Exp. Bot. 2017, 134, 72–81. [Google Scholar] [CrossRef] [Green Version]
- Okunlola, G.O.; Akinwale, R.O.; Adelusi, A.A. Proline and soluble sugars accumulation in three pepper species (Capsicum spp) in response to water stress imposed at different stages of growth. Sci. Cold Arid. Reg. 2018, 8, 205–211. [Google Scholar] [CrossRef]
- Shah, K.; Kumar, R.G.; Verma, S.; Dubey, R. Effect of cadmium on lipid peroxidation, superoxide anion generation and activities of antioxidant enzymes in growing rice seedlings. Plant Sci. J. 2001, 161, 1135–1144. [Google Scholar] [CrossRef]
- Xiao, S.; Li, Y.Y.; Quan, F.Z.; Wei, B.W. Effect of Pb toxicity on leaf growth, antioxidant enzyme activities, and photosynthesis in cuttings and seedlings of Jatropha curcas L. Environ. Sci. Pollut. Res. 2012, 19, 893–902. [Google Scholar] [CrossRef]
- Zembala, M.; Filek, M.; Walas, S.; Mrowiec, H.; Kornaś, A.; Miszalski, Z.; Hartikainen, H. Effect of selenium on macro-and microelement distribution and physiological parameters of rape and wheat seedlings exposed to cadmium stress. Plant Soil 2010, 329, 457–468. [Google Scholar] [CrossRef]
- Ahammed, G.J.; Wu, M.; Wang, Y.; Yan, Y.; Mao, Q.; Ren, J.; Ma, R.; Liu, A.; Chen, S. Melatonin alleviates iron stress by improving iron homeostasis, antioxidant defense and secondary metabolism in cucumber. Sci. Hortic. 2020, 265, 109205. [Google Scholar] [CrossRef]
- Ulhassan, Z.; Huang, Q.; Gill, R.A.; Ali, S.; Mwamba, T.M.; Ali, B.; Hina, F.; Zhou, W. Protective mechanisms of melatonin against selenium toxicity in Brassica napus: Insights into physiological traits, thiol biosynthesis and antioxidant machinery. BMC Plant Biol. 2019, 19, 507. [Google Scholar] [CrossRef]
- Posmyk, M.M.; Kuran, H.; Marciniak, K.; Janas, K.M. Pre-sowing seed treatment with melatonin protects red cabbage seedlings against toxic copper ion concentrations. J. Pineal Res. 2008, 45, 24–31. [Google Scholar] [CrossRef]
- Rascio, N.; Della-Vecchia, F.; La-Rocca, N.; Barbato, R.; Pagliano, C.; Raviolo, M.; Gonnelli, C.; Gabbrielli, R. Metal accumulation and damage in rice (cv. Vialone nano) seedlings exposed to cadmium. Environ. Exp. Bot. 2008, 62, 267–278. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vanadium (mg L−1) | Biomass Yield Per Plant (g) | Growth Tolerance Index (%) | ||||
|---|---|---|---|---|---|---|
| Fresh | Dry | Shoot | Root | |||
| Shoot | Root | Shoot | Root | |||
| 0 | 5.310 ± 0.217 b | 0.728 ± 0.015 b | 1.483 ± 0.054 b | 0.106 ± 0.001 b | 100 | 100 |
| 10 | 6.381 ± 0.125 a | 0.791 ± 0.007 a | 1.722 ± 0.066 a | 0.114 ± 0.001 a | 116 | 107 |
| 20 | 4.793 ± 0.229 c | 0.586 ± 0.015 c | 1.266 ± 0.058 c | 0.077 ± 0.003 c | 85 | 72 |
| 30 | 2.843 ± 0.138 d | 0.389 ± 0.012 d | 0.845 ± 0.056 d | 0.051 ± 0.002 d | 56 | 48 |
| 40 | 1.473 ± 0.153 e | 0.193 ± 0.010 e | 0.510 ± 0.052 e | 0.036 ± 0.001 e | 34 | 34 |
| 50 | 0.929 ± 0.111 f | 0.135 ± 0.007 f | 0.351 ± 0.041 f | 0.024 ± 0.001 f | 23 | 22 |
| Vanadium (mg L−1) | V Concentration | V Uptake | V Translocation (Root to Shoot) | ||
|---|---|---|---|---|---|
| Root | Shoot | Root | Shoot | ||
| 0 | 4.851 ± 0.351 f | 1.530 ± 0.202 f | 0.513 ± 0.043 c | 2.261 ± 0.284 c | 0.316 ± 0.037 d |
| 10 | 12.543 ± 0.967 e | 4.573 ± 0.661 e | 1.428 ± 0.095 ab | 7.903 ± 1.251 b | 0.361 ± 0.025 cd |
| 20 | 19.697 ± 1.380 d | 8.376 ± 0.481 d | 1.515 ± 0.051 a | 10.665 ± 1.074 a | 0.426 ± 0.007 c |
| 30 | 31.011 ± 1.035 c | 13.963 ± 0.739 c | 1.555 ± 0.039 a | 11.743 ± 0.508 a | 0.449 ± 0.009 bc |
| 40 | 39.577 ± 1.523 b | 20.806 ± 1.122 b | 1.434 ± 0.003 ab | 10.537 ± 0.771 a | 0.529 ± 0.049 ab |
| 50 | 51.876 ± 1.735 a | 29.409 ± 1.213 a | 1.266 ± 0.118 b | 10.221 ± 1.006 ab | 0.569 ± 0.042 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Altaf, M.A.; Shu, H.; Hao, Y.; Zhou, Y.; Mumtaz, M.A.; Wang, Z. Vanadium Toxicity Induced Changes in Growth, Antioxidant Profiling, and Vanadium Uptake in Pepper (Capsicum annum L.) Seedlings. Horticulturae 2022, 8, 28. https://doi.org/10.3390/horticulturae8010028
Altaf MA, Shu H, Hao Y, Zhou Y, Mumtaz MA, Wang Z. Vanadium Toxicity Induced Changes in Growth, Antioxidant Profiling, and Vanadium Uptake in Pepper (Capsicum annum L.) Seedlings. Horticulturae. 2022; 8(1):28. https://doi.org/10.3390/horticulturae8010028
Chicago/Turabian StyleAltaf, Muhammad Ahsan, Huangying Shu, Yuanyuan Hao, Yan Zhou, Muhammad Ali Mumtaz, and Zhiwei Wang. 2022. "Vanadium Toxicity Induced Changes in Growth, Antioxidant Profiling, and Vanadium Uptake in Pepper (Capsicum annum L.) Seedlings" Horticulturae 8, no. 1: 28. https://doi.org/10.3390/horticulturae8010028