
Effects of Drought on Yield and Nutraceutical Properties of Beans (Phaseolus spp.) Traditionally Cultivated in Veneto, Italy

 , , , ,
, , , ,  ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
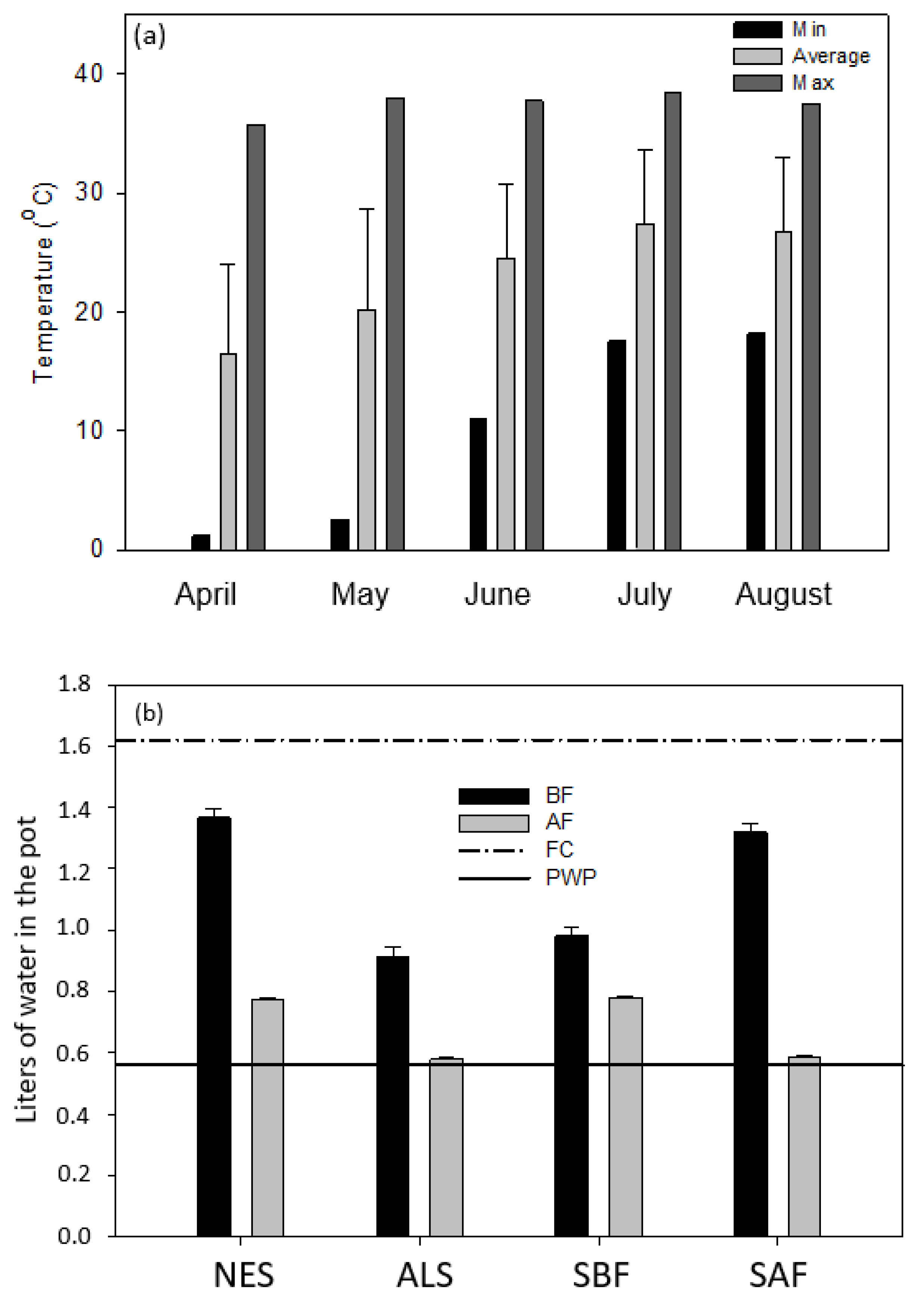
2.2. Experimental Design
2.3. Plant Development and Yield
- i.
- Vegetative phase: from germination until first flower opens;
- ii.
- Flowering: from the first flowers opening until the first pods are visible;
- iii.
- Fresh pods: from the first green pod being visible until the first pod starts to dry and ripen;
- iv.
- Dry pods: from when the pods and seeds ripen until the plant’s senescence.
2.4. Seeds’ Nutraceutical Characterization
2.5. Statistical Analysis
3. Results
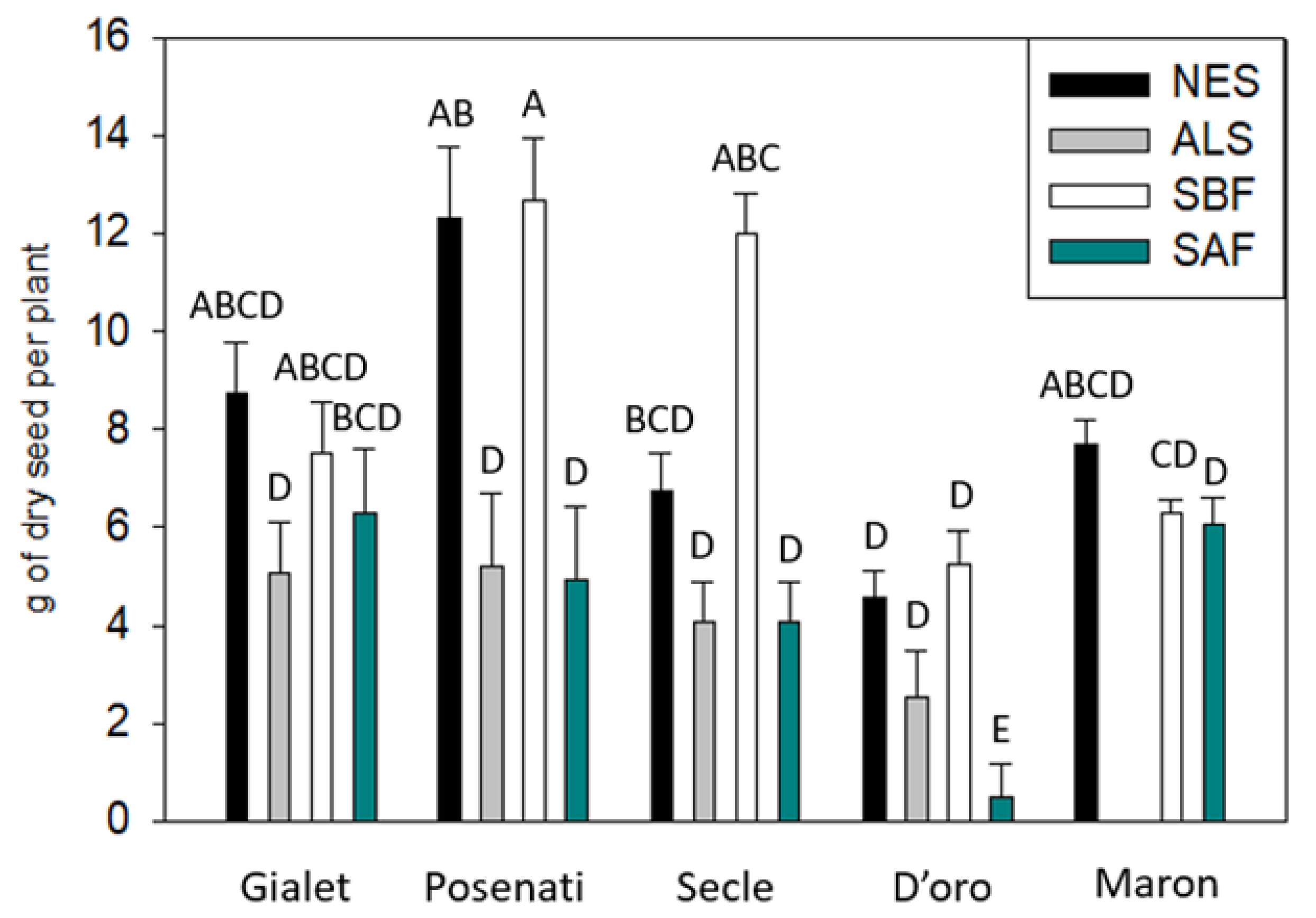
3.1. Yield
3.2. Nutraceutical Properties
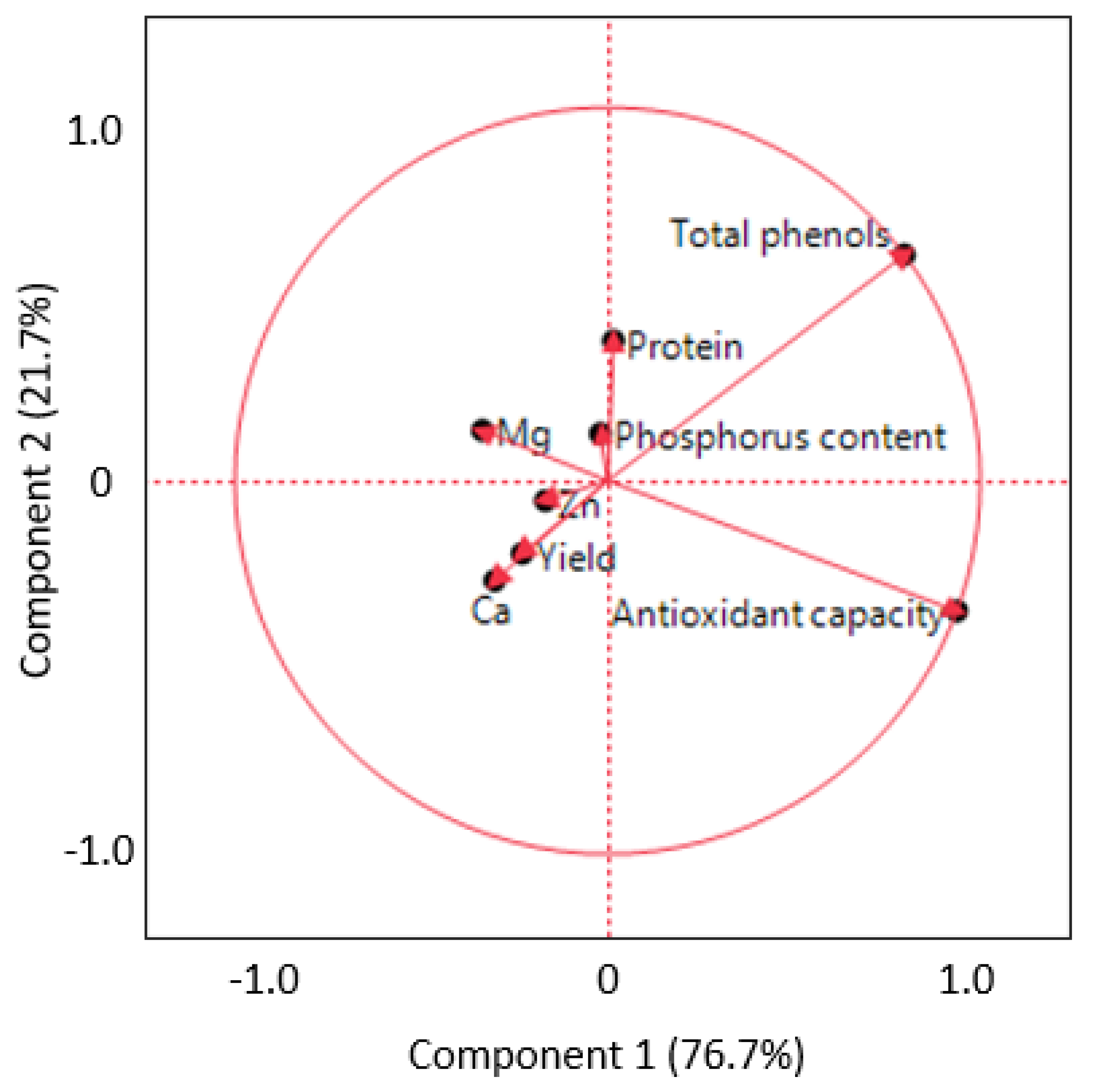
3.3. Principal Component Analysis and Correlations
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. Pulses Contribute to Food Security. Available online: http://www.fao.org/fileadmin/user_upload/pulses-2016/docs/factsheets/FoodSecurity_EN_PRINT.pdf (accessed on 30 August 2020).
- Jimenez-Lopez, J.C.; Singh, K.B.; Clemente, A.; Nelson, M.N.; Ochatt, S.; Smith, P.M.C. Editorial: Legumes for Global Food Security. Front. Plant. Sci. 2020, 11, 926. [Google Scholar] [CrossRef] [PubMed]
- Blair, M.W. Mineral biofortification strategies for food staples: The example of common bean. J. Agric. Food Chem. 2013, 61, 8287–8294. [Google Scholar] [CrossRef] [PubMed]
- McClean, P.E.; Raatz, B. Common Bean Genomes: Mining New Knowledge of a Major Societal Crop. In The Common Bean Genome; de la Vega, M.P., Marsolais, F., Santalla, M., Eds.; Springer International Printer: Gewebestrasse, Switzerland, 2017; p. 295. ISBN 978-3-319-63526-2. [Google Scholar]
- Jones, A. Phaseoulus Beans: Post-Harvest Operations. Available online: http://www.fao.org/3/a-av015e.pdf (accessed on 13 January 2020).
- Calles, T. Preface to special issue on leguminous pulses. Plant. Cell Tiss Organ. Cult. 2016, 127, 541–542. [Google Scholar] [CrossRef] [Green Version]
- Peix, A.; Ramírez-Bahena, M.H.; Velázquez, E.; Bedmar, E.J. Bacterial Associations with Legumes. CRC Crit. Rev. Plant. Sci. 2015, 34, 17–42. [Google Scholar] [CrossRef]
- Courty, P.E.; Smith, P.; Koegel, S.; Redecker, D.; Wipf, D. Inorganic Nitrogen Uptake and Transport in Beneficial Plant Root-Microbe Interactions. CRC Crit. Rev. Plant. Sci. 2015, 34, 4–16. [Google Scholar] [CrossRef]
- FAO. How to Feed the World in 2050; FAO: Rome, Italy, 2009. [Google Scholar]
- Arnoldi, A.; Zanoni, C.; Lammi, C.; Boschin, G. The Role of Grain Legumes in the Prevention of Hypercholesterolemia and Hypertension. CRC Crit. Rev. Plant. Sci. 2015, 34, 144–168. [Google Scholar] [CrossRef]
- Guzmán-Maldonado, S.H.; Acosta-Gallegos, J.; Paredes-López, O. Protein and mineral content of a novel collection of wild and weedy common bean (Phaseolus vulgaris L). J. Sci. Food Agric. 2000, 80, 1874–1881. [Google Scholar] [CrossRef]
- Mitchell, C.J.; Milan, A.M.; Mitchell, S.M.; Zeng, N.; Ramzan, F.; Sharma, P.; Knowles, S.O.; Roy, N.C.; Sjödin, A.; Wagner, K.-H.; et al. The effects of dietary protein intake on appendicular lean mass and muscle function in elderly men: A 10-wk randomized controlled trial. Am. J. Clin. Nutr. 2017, 106, 1375–1383. [Google Scholar] [CrossRef] [Green Version]
- Rand, W.M.; Pellett, P.L.; Young, V.R. Meta-analysis of nitrogen balance studies for estimating protein requirements in healthy adults. Am. J. Clin. Nutr. 2003, 77, 109–127. [Google Scholar] [CrossRef] [Green Version]
- Díaz-Batalla, L.; Widholm, J.M.; Fahey, G.C.; Castaño-Tostado, E.; Paredes-López, O. Chemical Components with Health Implications in Wild and Cultivated Mexican Common Bean Seeds (Phaseolus vulgaris L.). J. Agric. Food Chem. 2006, 54, 2045–2052. [Google Scholar] [CrossRef]
- Ombra, M.N.; D’acierno, A.; Nazzaro, F.; Riccardi, R.; Spigno, P.; Zaccardelli, M.; Pane, C.; Maione, M.; Fratianni, F. Phenolic Composition and Antioxidant and Antiproliferative Activities of the Extracts of Twelve Common Bean (Phaseolus vulgaris L.) Endemic Ecotypes of Southern Italy before and after Cooking. Oxid. Med. Cell. Longev. 2016, 2016, 1398298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oroian, M.; Escriche, I. Antioxidants: Characterization, natural sources, extraction and analysis. Food Res. Int. 2015, 74, 10–36. [Google Scholar] [CrossRef] [PubMed]
- Cardador-Martínez, A.; Albores, A.; Bah, M.; Calderón-Salinas, V.; Castaño-Tostado, E.; Guevara-González, R.; Shimada-Miyasaka, A.; Loarca-Piña, G. Relationship among antimutagenic, antioxidant and enzymatic activities of methanolic extract from common beans (Phaseolus vulgaris L). Plant. Foods Hum. Nutr. 2006, 61, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Kalogeropoulos, N.; Chiou, A.; Ioannou, M.; Karathanos, V.T.; Hassapidou, M.; Andrikopoulos, N.K. Nutritional evaluation and bioactive microconstituents (phytosterols, tocopherols, polyphenols, triterpenic acids) in cooked dry legumes usually consumed in the Mediterranean countries. Food Chem. 2010, 121, 682–690. [Google Scholar] [CrossRef]
- Ranilla, L.G.; Genovese, M.I.; Lajolo, F.M. Effect of different cooking conditions on phenolic compounds and antioxidant capacity of some selected Brazilian bean (Phaseolus vulgaris L.) cultivars. J. Agric. Food Chem. 2009, 57, 5734–5742. [Google Scholar] [CrossRef]
- NIH–U.S. (National Center for Complementary and Integrative Health) Antioxidants: In Depth|NCCIH. Available online: https://www.nccih.nih.gov/health/antioxidants-in-depth (accessed on 7 July 2020).
- USDA (U.S. Department of Agriculture) and HHS (U.S. Department of Health and Human Services). 2015–2020 Dietary Guidelines for Americans, 8th Edition. Available online: https://health.gov/our-work/food-nutrition/previous-dietary-guidelines/2015 (accessed on 30 August 2020).
- Alloway, B.J. Zinc In Soils and Crop Nutrition, 2nd ed.; IZA and IFA: Paris, France, 2008; ISBN 978-90-8133-310-8. [Google Scholar]
- Wessells, K.R.; Brown, K.H. Estimating the Global Prevalence of Zinc Deficiency: Results Based on Zinc Availability in National Food Supplies and the Prevalence of Stunting. PLoS ONE 2012, 7, e50568. [Google Scholar] [CrossRef] [Green Version]
- Cakmak, I.; McLaughlin, M.J.; White, P. Zinc for better crop production and human health. Plant. Soil 2017, 411, 1–4. [Google Scholar] [CrossRef]
- Larochelle, C.; Katungi, E.; Beebe, S. Disaggregated Analysis of Bean Consumption Demand and Contribution to Household Food Security in Uganda; CGIAR: Cali, Colombia, 2015. [Google Scholar]
- Bitocchi, E.; Rau, D.; Bellucci, E.; Rodriguez, M.; Murgia, M.L.; Gioia, T.; Santo, D.; Nanni, L.; Attene, G.; Papa, R. Beans (Phaseolus ssp.) as a model for understanding crop evolution. Front. Plant. Sci. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Zeven, A.C. The introduction of the common bean (Phaseolus vulgaris L.) into Western Europe and the phenotypic variation of dry beans collected in the Netherlands in 1946. Euphytica 1997, 94, 319–328. [Google Scholar] [CrossRef]
- De Ron, A.M.; González, A.M.; Rodiño, A.P.; Santalla, M.; Godoy, L.; Papa, R. History of the common bean crop: Its evolution beyond its areas of origin and domestication. Arbor 2016, 192, a317. [Google Scholar] [CrossRef] [Green Version]
- Albala, K. Phaseolus vulgaris: Mexico and the World. In Beans: A History; Bloomsbury Academic: London, UK, 2007; p. 240. ISBN 978-1-8452-0430-3. [Google Scholar]
- Venora, G.; Grillo, O.; Ravalli, C.; Cremonini, R. Identification of Italian landraces of bean (Phaseolus vulgaris L.) using an image analysis system. Sci. Hortic. 2009, 121, 410–418. [Google Scholar] [CrossRef]
- Piergiovanni, A.R.; Lioi, L. Italian Common Bean Landraces: History, Genetic Diversity and Seed Quality. Diversity 2010, 2, 837–862. [Google Scholar] [CrossRef] [Green Version]
- Spagnoletti Zeuli, P.L.; Baser, N.; Riluca, M.; Laghetti, G.; Logozzo, G.; Masi, P.; Molinari, S.; Negri, V.; Olita, G.; Tiranti, B.; et al. Valorisation and certification of Italian bean agro-ecotypes (Phaseolus vulgaris). In Proceedings of the Ecotipi Vegetali Italiani: Una Preziosa Risorsa di Variabilità Genetica, Roma, Italy, 7 October 2004; p. 19. (In Italian). [Google Scholar]
- Beebe, S.E.; Rao, I.M.; Blair, M.W.; Acosta-Gallegos, J.A. Phenotyping common beans for adaptation to drought. Front. Physiol. 2013, 4, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broughton, W.J.; Hernández, G.; Blair, M.; Beebe, S.; Gepts, P.; Vanderleyden, J. Beans (Phaseolus spp.)—Model Food Legumes. Plant Soil 2003, 252, 55–128. [Google Scholar] [CrossRef] [Green Version]
- FAOSTAT Beans, Dry. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 7 January 2020).
- NIDIS Current Conditions | Global Drought Information System. Available online: https://www.drought.gov/gdm/current-conditions (accessed on 4 June 2020).
- USDA Risk of Human-Induced Desertification Map | NRCS Soils. Available online: https://www.nrcs.usda.gov/wps/portal/nrcs/detail/soils/survey/geo/?cid=nrcs142p2_054004 (accessed on 9 October 2020).
- Hendricks, T.; Franklin, D.; Dahal, S.; Hancock, D.; Stewart, L.; Cabrera, M.; Hawkins, G. Soil carbon and bulk density distribution within 10 Southern Piedmont grazing systems. J. Soil Water Conserv. 2019, 74, 323–333. [Google Scholar] [CrossRef]
- Singh, S.P. A Key for the Identification of Different Growth Habits of Phaseolus vulgaris; CIAT: Cali, Colombia, 1981. [Google Scholar]
- Feller, C.H.; Bleiholder, L.; Buhr, H.; Hack, M.; Hess, R.; Klose, U.; Meier, R.; Stauss, T.; Van Den Boom, E. Phenological stages of development of vegetables: II. Fruit vegetables and legumes. In Growth Stages of Mono-and Dicotyledonous Plants BBCH Monograph Federal Biological Research Centre for Agriculture and Forestry (2001); Meier, U., Ed.; BBA: Berlin, Germany, 1995; pp. 141–144. [Google Scholar]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar] [CrossRef]
- Fischer Scientific Folin and Ciocalteu’s Phenol Reagent, 2.0N, MP Biomedicals 500 mL: Chemicals | Fisher Scientific. Available online: https://www.fishersci.com/shop/products/folin-ciocalteu-s-phenol-reagent-2-0n-mp-biomedicals-500mL/icn19518690 (accessed on 5 September 2020).
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Hjalmarsson, S.; Akesson, R. Modern Kjeldahl Procedure. Int. Lab. 1983, 3, 70–76. [Google Scholar]
- Jirka, A.M.; Carter, M.J.; May, D.; Fuller, F.D. Ultramicro Semiautomated Method for Simultaneous Determination of Total Phosphorus and Total Kjeldahl Nitrogen in Wastewaters. Environ. Sci. Technol. 1976, 10, 1038–1044. [Google Scholar] [CrossRef]
- Giurcă, D.M. Morphological and phenological differences between the two species of the Phaselous genus (Phaseolus vulgaris and Phaseolus Coccineus). Cercet. Agron. în Mold. 2009, XLII, 39–45. [Google Scholar]
- Polania, J.; Rao, I.M.; Cajiao, C.; Rivera, M.; Raatz, B.; Beebe, S. Physiological traits associated with drought resistance in Andean and Mesoamerican genotypes of common bean (Phaseolus vulgaris L.). Euphytica 2016, 210, 17–29. [Google Scholar] [CrossRef]
- Dipp, C.C.; Marchese, J.A.; Woyann, L.G.; Bosse, M.A.; Roman, M.H.; Gobatto, D.R.; Paludo, F.; Fedrigo, K.; Kovali, K.K.; Finatto, T. Drought stress tolerance in common bean: What about highly cultivated Brazilian genotypes? Euphytica 2017, 213, 1–16. [Google Scholar] [CrossRef]
- Smith, M.R.; Veneklaas, E.; Polania, J.; Rao, I.M.; Beebe, S.E.; Merchant, A. Field drought conditions impact yield but not nutritional quality of the seed in common bean (Phaseolus vulgaris L.). PLoS ONE 2019, 14, e0217099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orto Quando Irrigare le Piante di Fagioli. Available online: https://www.ortodacoltivare.it/domande/irrigare-fagioli.html (accessed on 7 September 2020).
- Albanesi Coltivazione dei Fagioli—Coltivazione dei Borlotti-Albanesi.it. Available online: https://www.albanesi.it/ambiente/orto/coltivazione-fagioli.htm (accessed on 7 September 2020).
- Giardinaggio Fagiolo-Phaseolus vulgaris-Orto-Fagiolo-Phaseolus Vulgaris—Orto. Available online: https://www.giardinaggio.it/orto/singoleorticole/fagiolo/fagiolo.asp (accessed on 7 September 2020).
- Nicoletto, C.; Zanin, G.; Sambo, P.; Dalla Costa, L. Quality assessment of typical common bean genotypes cultivated in temperate climate conditions and different growth locations. Sci. Hortic. 2019, 256, 108599. [Google Scholar] [CrossRef]
- Rocha-Guzmán, N.E.; González-Laredo, R.F.; Ibarra-Pérez, F.J.; Nava-Berúmen, C.A.; Gallegos-Infante, J.A. Effect of pressure cooking on the antioxidant activity of extracts from three common bean (Phaseolus vulgaris L.) cultivars. Food Chem. 2007, 100, 31–35. [Google Scholar] [CrossRef]
- Boateng, J.; Verghese, M.; Walker, L.T.; Ogutu, S. Effect of processing on antioxidant contents in selected dry beans (Phaseolus spp. L.). LWT Food Sci. Technol. 2008, 41, 1541–1547. [Google Scholar] [CrossRef]
- Barroga, C.F.; Laurena, A.C.; Mendoza, E.M.T. Polyphenols in Mung Bean (Vigna radiata (L.) Wilczek): Determination and Removal. J. Agric. Food Chem. 1985, 33, 1006–1009. [Google Scholar] [CrossRef]
- Blair, M.W.; Astudillo, C.; Grusak, M.A.; Graham, R.; Beebe, S.E. Inheritance of seed iron and zinc concentrations in common bean (Phaseolus vulgaris L.). Mol. Breed. 2009, 23, 197–207. [Google Scholar] [CrossRef]
- Amongi, W.; Mukankusi, C.; Sebuliba, S.; Mukamuhirwa, F. Iron and zinc grain concentrations diversity and agronomic performance of common bean germplasm collected from East Africa. African J. Food Agric. Nutr. Dev. 2018, 18, 13717–13742. [Google Scholar] [CrossRef]
- Celmeli, T.; Sari, H.; Canci, H.; Sari, D.; Adak, A.; Eker, T.; Toker, C. The nutritional content of common bean (phaseolus vulgaris l.) landraces in comparison to modern varieties. Agronomy 2018, 8, 166. [Google Scholar] [CrossRef] [Green Version]
- NIH (National Institutes of Health) Zinc—Consumer. Available online: https://ods.od.nih.gov/factsheets/Zinc-Consumer/ (accessed on 13 October 2020).
- Paredes, M.; Becerra, V.; Tay, J. Inorganic Nutritional Composition of Common Bean (Phaseolus vulgaris L.) Genotypes Race Chile. Chil. J. Agric. Res. 2010, 69, 486–495. [Google Scholar] [CrossRef]
- Bulyaba, R.; Winham, D.M.; Lenssen, A.W.; Moore, K.J.; Kelly, J.D.; Brick, M.A.; Wright, E.M.; Ogg, J.B. Genotype by Location Effects on Yield and Seed Nutrient Composition of Common Bean. Agronomy 2020, 10, 347. [Google Scholar] [CrossRef] [Green Version]
- Kazai, P.; Noulas, C.; Khah, E.; Vlachostergios, D. Yield and seed quality parameters of common bean cultivars grown under water and heat stress field conditions. AIMS Agric. Food 2019, 4, 285–302. [Google Scholar] [CrossRef]
- Davis, D.R.; Epp, M.D.; Riordan, H.D. Changes in USDA Food Composition Data for 43 Garden Crops, 1950 to 1999. J. Am. Coll. Nutr. 2004, 23, 669–682. [Google Scholar] [CrossRef] [PubMed]
- Krüger, E.; Dietrich, H.; Hey, M.; Patz, C.D. Effects of cultivar, yield, berry weight, temperature and ripening stage on bioactive compounds of black currants. J. Appl. Bot. Food Qual. 2011, 84, 40–46. [Google Scholar]
- Ribeiro, N.D.; Mambrin, R.B.; Storck, L.; Prigol, M.; Nogueira, C.W. Combined selection for grain yield, cooking quality and minerals in the common bean. Rev. Ciência Agronômica 2013, 44, 869–877. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, N.D.; Jost, E.; Maziero, S.M.; Storck, L.; Rosa, D.P. Selection of common bean lines with high grain yield and high grain calcium and iron concentrations. Rev. Ceres 2014, 61, 77–83. [Google Scholar] [CrossRef] [Green Version]
- Domingues, L.d.S.; Ribeiro, N.D.; Andriolo, J.L.; Possobom, M.T.D.F.; Zemolin, A.E.M. Growth, grain yield and calcium, potassium and magnesium accumulation in common bean plants as related to calcium nutrition. Acta Sci. Agron. 2016, 38, 207–217. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Varieties | Treatments | Varieties × Treatments |
|---|---|---|---|
| Yield (g per plant) | <0.001 (11.71) | <0.001 (14.57) | 0.021 (2.56) |
| Total phenols (mg GAE kg−1 dw) | <0.001 (17.13) | 0.065 (2.87) | 0.798 (0.63) |
| Antioxidant capacity (mg Fe+2 kg−1 dw) | 0.006 (4.57) | 0.301 (1.22) | 0.296 (1.21) |
| Protein content (%) | 0.198 (1.64) | 0.057 (3.10) | 0.813 (0.60) |
| Total phosphorus content (mg kg−1) | 0.196 (1.69) | 0.698 (0.36) | 0.053 (2.21) |
| Mg content (mg kg−1) | 0.231 (1.49) | 0.489 (0.73) | 0.597 (0.85) |
| Ca content (mg kg−1) | <0.001 (8.72) | 0.111 (2.32) | 0.149 (1.43) |
| Zn content (mg kg−1) | <0.001 (30.43) | 0.070 (2.83) | 0.002 (4.33) |
| Degrees of freedom | 4 | 3 | 12 |
| Parameters | Gialet | Posenati | Secle | D’oro | Maron |
|---|---|---|---|---|---|
| Total phenols (mg GAE Kg−1 dw) | 2455 (76) c | 3368 (76) b | 3326 (52) b | 4288 (77) a | 4128 (67) a |
| Antioxidant capacity (mg Fe+2 kg−1 dw) | 2505 (36) c | 3718 (85) b | 3256 (88) bc | 3736 (154) b | 4545 (135) a |
| Protein content (% dw) n.s. | 30.9 (4.4) | 30.3 (2.4) | 28.2 (3.4) | 32.3 (2.8) | 27.1 (3.5) |
| Total phosphorus (mg kg−1 dw) n.s. | 1395 (36) | 1386 (54) | 1439 (81) | 1422 (28) | 1379 (55) |
| Mg content (mg kg−1 dw) n.s. | 297 (13.16) | 287.8 (14.7) | 292.7 (12.4) | 296.3 (6.5) | 281.3 (11.1) |
| Ca content (mg kg−1 dw) | 1280 (163) a | 1118 (193) a | 996 (194) b | 945 (133) b | 1081 (132) ab |
| Varieties | Treatment | Zn Content (mg kg−1) |
|---|---|---|
| Gialet | NES | 58.4 (1.9) abcd |
| ALS | 56.8 (2.3) abcde | |
| SBF | 61.8 (6.8) abc | |
| SAF | 65.6 (6.7) ab | |
| Posenati | NES | 43.9 (2.7) efg |
| ALS | 47.4 (3.6) defg | |
| SBF | 37.1 (5.2) g | |
| SAF | 38.7 (2.4) fg | |
| Secle | NES | 47.9 (3.0) cdefg |
| ALS | 49.2 (1.8) bcdefg | |
| SBF | 58.3 (8.6) abcd | |
| SAF | 50.9 (4.8) bcdefg | |
| D’oro | NES | 49.4 (0.3) bcdefg |
| ALS | 56.2 (1.1) abcdef | |
| SBF | 41.2 (1.6) fg | |
| SAF | 37.5 (2.7) fg | |
| Maron | NES | 69.2 (0.3) a |
| ALS | — | |
| SBF | 58.3 (7.1) abcd | |
| SAF | 51.4 (1.8) abcdefg |
| Yield | TP | AC | Protein | P | Zn | Mg | Ca | |
|---|---|---|---|---|---|---|---|---|
| Yield | 1 | |||||||
| TP | −0.5354 * | 1 | ||||||
| AC | −0.3431 * | 0.6675 | 1 | |||||
| Protein | −0.2955 * | 0.3749 * | −0.0078 | 1 | ||||
| P | −0.1147 | 0.1566 | 0.0458 | −0.1097 | 1 | |||
| Zn | 0.0198 | −0.2747 * | −0.2994 * | −0.1471 | −0.0765 | 1 | ||
| Mg | 0.0019 | −0.1941 | −0.2381 | −0.0455 | 0.0997 | −0.0504 | 1 | |
| Ca | 0.5062 * | −0.6477 * | −0.4166 * | −0.1714 | −0.1023 | 0.2366 | 0.2335 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sica, P.; Galvao, A.; Scariolo, F.; Maucieri, C.; Nicoletto, C.; Pilon, C.; Sambo, P.; Barcaccia, G.; Borin, M.; Cabrera, M.; et al. Effects of Drought on Yield and Nutraceutical Properties of Beans (Phaseolus spp.) Traditionally Cultivated in Veneto, Italy. Horticulturae 2021, 7, 17. https://doi.org/10.3390/horticulturae7020017
Sica P, Galvao A, Scariolo F, Maucieri C, Nicoletto C, Pilon C, Sambo P, Barcaccia G, Borin M, Cabrera M, et al. Effects of Drought on Yield and Nutraceutical Properties of Beans (Phaseolus spp.) Traditionally Cultivated in Veneto, Italy. Horticulturae. 2021; 7(2):17. https://doi.org/10.3390/horticulturae7020017
Chicago/Turabian StyleSica, Pietro, Aline Galvao, Francesco Scariolo, Carmelo Maucieri, Carlo Nicoletto, Cristiane Pilon, Paolo Sambo, Gianni Barcaccia, Maurizio Borin, Miguel Cabrera, and et al. 2021. "Effects of Drought on Yield and Nutraceutical Properties of Beans (Phaseolus spp.) Traditionally Cultivated in Veneto, Italy" Horticulturae 7, no. 2: 17. https://doi.org/10.3390/horticulturae7020017