Ecologically Based Management of Pineapple Mealybug Wilt: Controlling Dysmicoccus brevipes Mealybug Populations with Salicylic Acid Analogs and Plant Extracts




Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Mealybugs
2.3. Treatments
2.3.1. In Vitro Toxicity of Compounds against Mealybugs
2.3.2. Biological Assay in Culture Chambers
2.3.3. Biological Assay in the Greenhouse
2.3.4. Biological Assays on Daughter Plants from Treated Parental Plants in the Greenhouse
2.4. Statistics
3. Results
3.1. In Vitro Toxicity of Compounds against Mealybugs
3.2. Biological Efficacy of Soil Application of SA against Mealybug Populations in Controlled Conditions
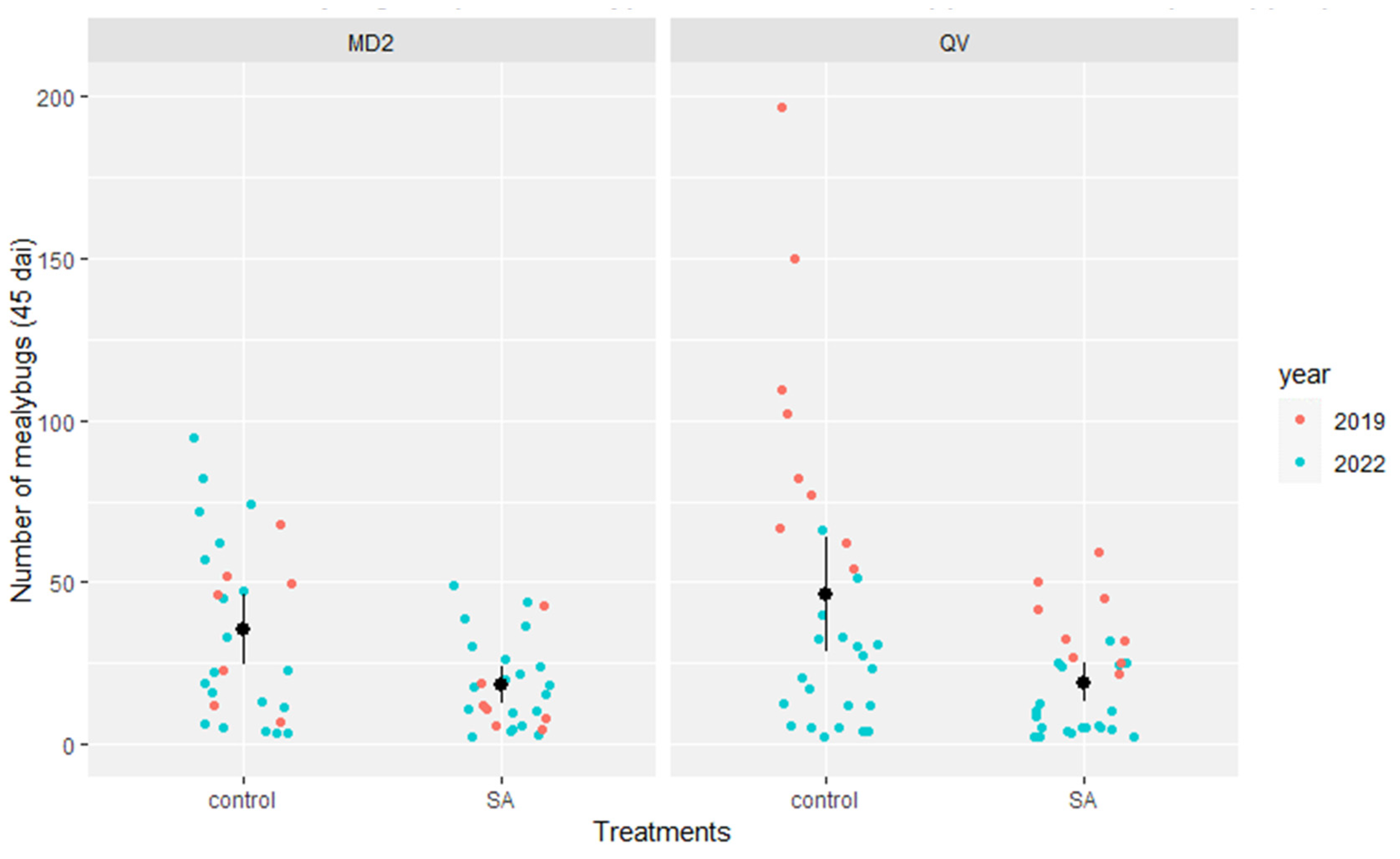
3.3. Biological Efficacy of Soil Application of SA, Natural Extracts, and Chemicals on Queen Victoria against Mealybug Populations in the Greenhouse
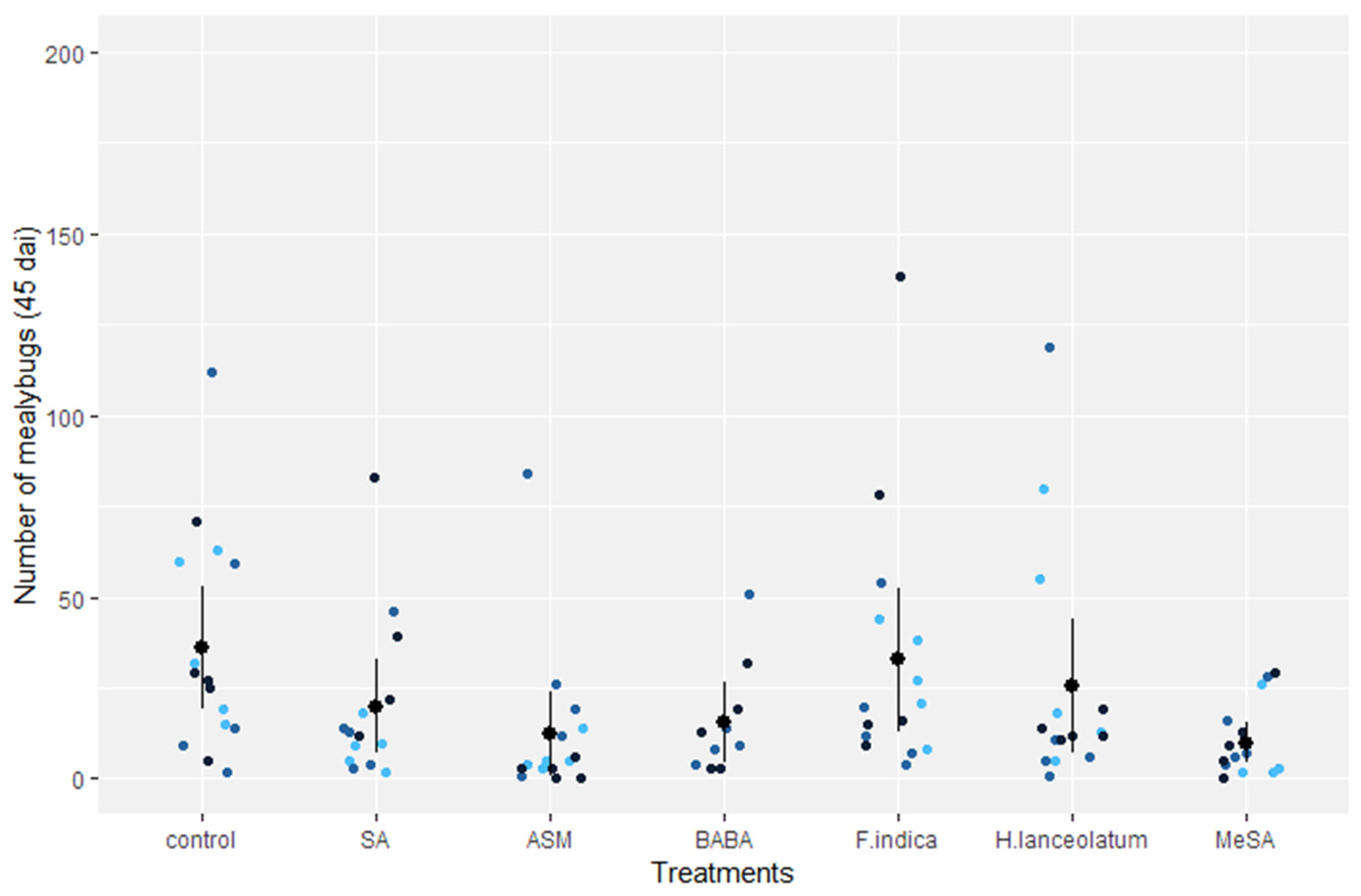
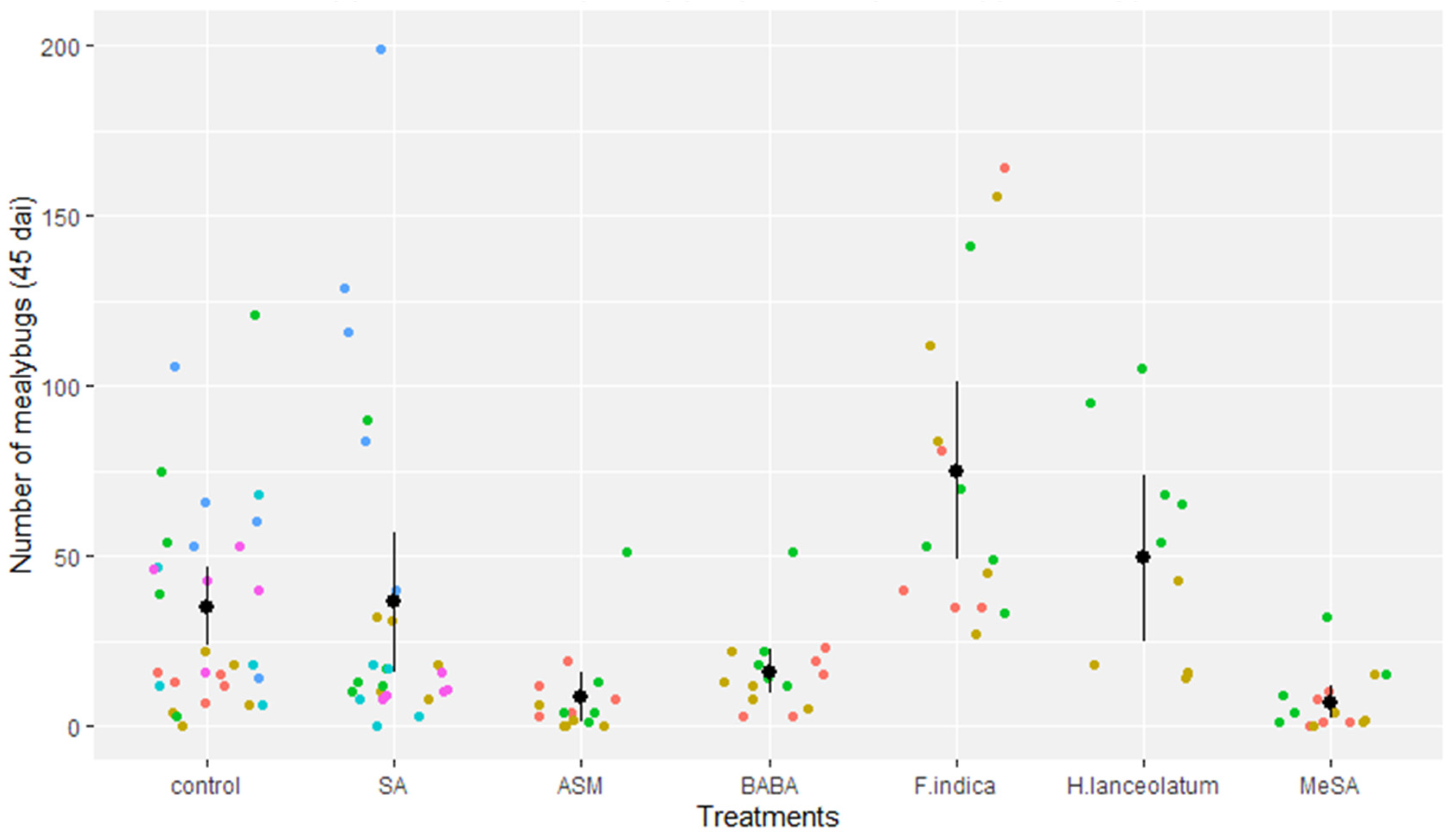
3.4. Biological Efficacy of Parental Application of SA, Natural Extracts, and Chemicals against Mealybug Populations on Daughter Plants in the Greenhouse
3.5. Effects of Treatments on Plant Physiology in the Greenhouse
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sether, D.M.; Hu, J.S. Yield Impact and Spread of Pineapple mealybug wilt associated virus-2 and Mealybug Wilt of Pineapple in Hawaii. Plant Dis. 2002, 86, 867–874. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.S.; Sether, D.M.; Metzer, M.J.; Pérez, E.; Gonsalves, A.; Karasev, A.V.; Nagai, C. Pineapple Mealybug Wilt Associated Virus and Mealybug Wilt of Pineapple. Acta Hortic. 2005, 666, 209–212. [Google Scholar] [CrossRef]
- Gambley, C.F.; Steele, V.; Geering, A.D.W.; Thomas, J.E. The Genetic Diversity of Ampeloviruses in Australian Pineapples and Their Association with Mealybug Wilt Disease. Australas. Plant Pathol. 2008, 37, 95. [Google Scholar] [CrossRef]
- Hu, J.S.; Sether, D.M.; Melzer, M.J.; Subere, C.V.; Cheah, K.; Chen, Y.; Li, Q.; Borth, W.; Wang, I.C.; Nagai, C.; et al. Characterization and Management of Pineapple Mealybug Wilt Associated Viruses. Acta Hortic. 2009, 822, 185–190. [Google Scholar] [CrossRef]
- Dey, K.; Green, J.; Melzer, M.; Borth, W.; Hu, J. Mealybug Wilt of Pineapple and Associated Viruses. Horticulturae 2018, 4, 52. [Google Scholar] [CrossRef]
- Nyarko, J.; Asare-Bediako, E. First Report of Pineapple Mealybug Wilt-associated Virus-1 and -3 in Ghanaian Pineapple. New Dis. Rep. 2019, 40, 18. [Google Scholar] [CrossRef]
- Sether, D.M.; Hu, J.S. Closterovirus Infection and Mealybug Exposure Are Necessary for the Development of Mealybug Wilt of Pineapple Disease. Phytopathology 2002, 92, 928–935. [Google Scholar] [CrossRef]
- Massé, D.; Cassam, N.; Hostachy, B.; Iskra-Caruana, M.-L.; Darnaudery, M.; Lefeuvre, P.; Lett, J.-M. First Report of Three Pineapple Mealybug Wilt-Associated Viruses in Queen Victoria Pineapples in Reunion Island. Plant Dis. 2021, 105, 715. [Google Scholar] [CrossRef]
- Gungoosingh-Bunwaree, A.; Maudarbaccus, F.; Knierim, D.; Margaria, P.; Winter, S.; Menzel, W. First Report of Pineapple Mealybug Wilt-associated Virus-1 and -2 Associated with Mealybug Wilt Disease of Pineapple in Mauritius. New Dis. Rep. 2021, 44, e12037. [Google Scholar] [CrossRef]
- Sether, D.M.; Melzer, M.J.; Busto, J.; Zee, F.; Hu, J.S. Diversity and Mealybug Transmissibility of Ampeloviruses in Pineapple. Plant Dis. 2005, 89, 450–456. [Google Scholar] [CrossRef] [PubMed]
- Darnaudery, M.; Fournier, P.; Léchaudel, M. Low-Input Pineapple Crops with High Quality Fruit: Promising Impacts of Locally Integrated and Organic Fertilisation Compared to Chemical Fertilisers. Exp. Agric. 2018, 54, 286–302. [Google Scholar] [CrossRef]
- Hossain, M.F. World Pineapple Production: An Overview. Afr. J. Food Agric. Nutr. Dev. 2016, 16, 11443–11456. [Google Scholar] [CrossRef]
- Yuan, M.; Ngou, B.P.M.; Ding, P.; Xin, X.-F. PTI-ETI Crosstalk: An Integrative View of Plant Immunity. Curr. Opin. Plant Biol. 2021, 62, 102030. [Google Scholar] [CrossRef]
- Komives, T. Vaccinating Plants. Ecocycles 2022, 8, 40–50. [Google Scholar] [CrossRef]
- Jones, J.D.G.; Dangl, J.L. The Plant Immune System. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Pruitt, R.N.; Gust, A.A.; Nürnberger, T. Plant Immunity Unified. Nat. Plants 2021, 7, 382–383. [Google Scholar] [CrossRef] [PubMed]
- Ngou, B.P.M.; Jones, J.D.G.; Ding, P. Plant Immune Networks. Trends Plant Sci. 2022, 27, 255–273. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.Q.; Dong, X. Systemic Acquired Resistance: Turning Local Infection into Global Defense. Annu. Rev. Plant Biol. 2013, 64, 839–863. [Google Scholar] [CrossRef] [PubMed]
- Klessig, D.F.; Choi, H.W.; Dempsey, D.A. Systemic Acquired Resistance and Salicylic Acid: Past, Present, and Future. Mol. Plant-Microbe Interact. 2018, 31, 871–888. [Google Scholar] [CrossRef]
- Park, S.-W.; Kaimoyo, E.; Kumar, D.; Mosher, S.; Klessig, D.F. Methyl Salicylate Is a Critical Mobile Signal for Plant Systemic Acquired Resistance. Science 2007, 318, 113–116. [Google Scholar] [CrossRef]
- Hilker, M.; Schwachtje, J.; Baier, M.; Balazadeh, S.; Bäurle, I.; Geiselhardt, S.; Hincha, D.K.; Kunze, R.; Mueller-Roeber, B.; Rillig, M.C.; et al. Priming and Memory of Stress Responses in Organisms Lacking a Nervous System. Biol. Rev. 2016, 91, 1118–1133. [Google Scholar] [CrossRef]
- Conrath, U.; Beckers, G.J.M.; Langenbach, C.J.G.; Jaskiewicz, M.R. Priming for Enhanced Defense. Annu. Rev. Phytopathol. 2015, 53, 97–119. [Google Scholar] [CrossRef]
- Martinez-Medina, A.; Flors, V.; Heil, M.; Mauch-Mani, B.; Pieterse, C.M.J.; Pozo, M.J.; Ton, J.; van Dam, N.M.; Conrath, U. Recognizing Plant Defense Priming. Trends Plant Sci. 2016, 21, 818–822. [Google Scholar] [CrossRef]
- Marolleau, B.; Gaucher, M.; Heintz, C.; Degrave, A.; Warneys, R.; Orain, G.; Lemarquand, A.; Brisset, M.-N. When a Plant Resistance Inducer Leaves the Lab for the Field: Integrating ASM into Routine Apple Protection Practices. Front. Plant Sci. 2017, 8, 1938. [Google Scholar] [CrossRef]
- De Matos, A.P.; Cabral, J.R.S.; Querino, C.M.B.; Caldas, R.C. Preliminary Report on Sytemic Acquired Resistance in Pineapple Plants to Control Fusarium Subglutinans. Acta Hortic. 2006, 702, 167–171. [Google Scholar] [CrossRef]
- Chinnasri, B.; Christopher, D.A.; Sipes, B.S. Evidence for the Induction of Systemic Acquired Resistance (SAR) by Acibenzolar in Cultivated Pineapple. Acta Hortic. 2006, 702, 151–156. [Google Scholar] [CrossRef]
- Chinnasri, B.; Borsics, T.; Christopher, D.A.; Sipes, B.S. Induction of Pathogenesis-Related Gene 1 (PR-1) by Acibenzolar-s-Methyl Application in Pineapple and Its Effect on Reniform Nematodes (Rotylenchulus reniformis). Agric. Nat. Resour. 2016, 50, 368–373. [Google Scholar] [CrossRef]
- Soler, A.; Marie-Alphonsine, P.-A.; Corbion, C.; Quénéhervé, P. Differential Response of Two Pineapple Cultivars (Ananas comosus (L.) Merr.) to SAR and ISR Inducers against the Nematode Rotylenchulus reniformis. Crop Prot. 2013, 54, 48–54. [Google Scholar] [CrossRef]
- Lu, X.; Sun, D.; Rookes, J.E.; Kong, L.; Zhang, X.; Cahill, D.M. Nanoapplication of a Resistance Inducer to Reduce Phytophthora Disease in Pineapple (Ananas comosus L.). Front. Plant Sci. 2019, 10, 1238. [Google Scholar] [CrossRef] [PubMed]
- Pandey, R.R.; Johnson, M.W. Enhanced Production of Pink Pineapple Mealybug, Dysmicoccus brevipes (Hemiptera: Pseudococcidae). Biocontrol Sci. Technol. 2006, 16, 389–401. [Google Scholar] [CrossRef]
- Joy, P.; Anjana, R.; Soumya, K. Insect Pests of pineapple and management. In Insect Pests Management of FRUIT CROPS; Pandey, A.K., Mall, P., Eds.; Biotech Books: Delhi, India, 2016; pp. 471–492. [Google Scholar]
- Momchev, P.; Ciganović, P.; Jug, M.; Marguí, E.; Jablan, J.; Zovko Končić, M. Comparison of Maceration and Ultrasonication for Green Extraction of Phenolic Acids from Echinacea Purpurea Aerial Parts. Molecules 2020, 25, 5142. [Google Scholar] [CrossRef]
- Joshi, S.; Subedi, P. Phytochemical and Biological Studies on Essential Oil and Leaf Extracts of Gaultheria fragrantissima Wall. Nepal J. Sci. Technol. 2014, 14, 59–64. [Google Scholar] [CrossRef]
- Balmer, D.; Planchamp, C.; Mauch-Mani, B. On the Move: Induced Resistance in Monocots. J. Exp. Bot. 2013, 64, 1249–1261. [Google Scholar] [CrossRef] [PubMed]
- Holeski, L.M.; Jander, G.; Agrawal, A.A. Transgenerational Defense Induction and Epigenetic Inheritance in Plants. Trends Ecol. Evol. 2012, 27, 618–626. [Google Scholar] [CrossRef] [PubMed]
- Conrath, U. Molecular aspects of defense priming. Trends Plant Sci. 2011, 16, 524–531. [Google Scholar] [CrossRef] [PubMed]
- Soler, A.; Pochat, C.; Perrin, M.; Mobarak, T.; N’Guessan, L.; Tullus, G. Control of Dysmicoccus brevipes Mealybugs Associated with Pineapple Wilt Disease Is Possible with Systemic Acquired Resistance (SAR) of the MD2 Variety. Presented at Acta Horticulturae, Proceedings of the X International Pineapple Symposium, Uvero Alto, Dominican Republic, 15–19 May 2023.
- Conrath, U.; Beckers, G.J.M.; Flors, V.; García-Agustín, P.; Jakab, G.; Mauch, F.; Newman, M.-A.; Pieterse, C.M.J.; Poinssot, B.; Pozo, M.J.; et al. Priming: Getting Ready for Battle. Mol. Plant-Microbe Interact. 2006, 19, 1062–1071. [Google Scholar] [CrossRef]
- Desmedt, W.; Vanholm, B.; Kyndt, T. Chapt 5—Plant Defense Priming in the Field: A Review. In Recent Highlights in the Discovery and Optimization of Crop Protection Products; Maienfisch, P., Mangelinckx, S., Eds.; Academic Press: Cambridge, MA, USA, 2021; pp. 87–124. [Google Scholar] [CrossRef]
- Parkinson, L.E.; Crew, K.S.; Thomas, J.E.; Dann, E.K. Efficacy of Acibenzolar-S-Methyl (Bion) Treatment of Australian Commercial Passionfruit, Passiflora Edulis f. Sp. Flavicarpa, on Resistance to Passionfruit Woodiness Virus (PWV) and Activities of Chitinase & β-1,3-Glucanase. Australas. Plant Pathol. 2015, 44, 311–318. [Google Scholar] [CrossRef]
- Jeon, H.W.; Park, A.R.; Sung, M.; Kim, N.; Mannaa, M.; Han, G.; Kim, J.; Koo, Y.; Seo, Y.-S.; Kim, J.-C. Systemic Acquired Resistance-Mediated Control of Pine Wilt Disease by Foliar Application With Methyl Salicylate. Front. Plant Sci. 2022, 12, 812414. [Google Scholar] [CrossRef]
- Gozzo, F.; Faoro, F. Systemic Acquired Resistance (50 Years after Discovery): Moving from the Lab to the Field. J. Agric. Food Chem. 2013, 61, 12473–12491. [Google Scholar] [CrossRef]
- Beckers, G.J.; Conrath, U. Priming for Stress Resistance: From the Lab to the Field. Curr. Opin. Plant Biol. 2007, 10, 425–431. [Google Scholar] [CrossRef]
- Luna, E.; Bruce, T.J.A.; Roberts, M.R.; Flors, V.; Ton, J. Next-Generation Systemic Acquired Resistance. Plant Physiol. 2012, 158, 844–853. [Google Scholar] [CrossRef]
- Gondor, O.K.; Pál, M.; Janda, T.; Szalai, G. The Role of Methyl Salicylate in Plant Growth under Stress Conditions. J. Plant Physiol. 2022, 277, 153809. [Google Scholar] [CrossRef] [PubMed]
- Kalaivani, K.; Maruthi-Kalaiselvi, M.; Senthil-Nathan, S. Seed Treatment and Foliar Application of Methyl Salicylate (MeSA) as a Defense Mechanism in Rice Plants against the Pathogenic Bacterium, Xanthomonas Oryzae Pv. Oryzae. Pestic. Biochem. Physiol. 2021, 171, 104718. [Google Scholar] [CrossRef]
- El-Solimany, E. The Impact of Faba Bean Seeds Soaking in Salicylic Acid, Acetyl-Salicylic Acid and Methyl Salicylate on Inducing Plant Resistance against the Cowpea Aphid, Aphis Craccivora Koch. J. Plant Prot. Pathol. 2020, 11, 243–247. [Google Scholar] [CrossRef]
- Hodge, S.; Thompson, G.A.; Powell, G. Application of DL-β-Aminobutyric Acid (BABA) as a Root Drench to Legumes Inhibits the Growth and Reproduction of the Pea Aphid Acyrthosiphon Pisum (Hemiptera: Aphididae). Bull. Entomol. Res. 2005, 95, 449–455. [Google Scholar] [CrossRef]
- Maerere, A.P. Axillary-Bud Development as It Determines Suckering in “Queen Victoria” and “Smooth Cayenne” Pineapples. Acta Hortic. 1997, 425, 309–320. [Google Scholar] [CrossRef]
- Avila, M.D.V.; Achimón, F.; Brito, V.D.; Aguilar, R.; Pizzolitto, R.P.; Zunino, M.P.; Peschiutta, M.L. Insecticidal Activity of Essential Oils against Mealybug Pests (Hemiptera: Pseudococcidae): A Systematic Review and Meta-Analysis. Plants 2022, 12, 109. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Treatment | Concentration | Source |
|---|---|---|
| SA (salicylic acid) | 1 mM | Sigma-Aldrich (St. Louis, MO, USA) |
| ASM (acibenzolar-S-methyl) | 1 mM | Bion 50WG Syngenta (Basel, Switzerland) |
| BABA (β-aminobutyric acid) | 1 mM | Sigma-Aldrich (St. Louis, MO, USA) |
| MeSA (essential oil of Gaultheria fragrantissima) | 1 mM | Pure Essential Bio (Washington, DC, USA) |
| Hypericum lanceolatum (herbal maceration) | 20 g/L | Tisane Bourbon (Le Tampon, France) |
| Flacourtia indica (herbal maceration) | 150 g/L | Collected in Réunion |
| Treatments | % Mortality of Mealybugs | |
|---|---|---|
| 1 Day | 8 Days | |
| SA (salicylic acid) | 0 | 0 |
| ASM (acibenzolar-S-methyl) | 0 | 0 |
| BABA (β-aminobutyric acid) | 0 | 0 |
| MeSA (essential oil of Gaultheria fragrantissima) | 0 | 0 |
| Hypericum lanceolatum (herbal maceration) | 0 | 0 |
| Flacourtia indica (herbal maceration) | 0 | 0 |
| Control (water) | 0 | 0 |
| Treatment | Mean Mealybugs/Plant | Percentage of Reduction Based on Control (%) | Risk of Plants Harboring More than 25 Mealybugs (%) |
|---|---|---|---|
| SA | 18.9 | 51.1% (p < 0.001) | 26.7% (* p = 0.001) |
| Control | 40.9 | - | 53.6% (* p = 0.002) |
| Treatment | Mean Mealybugs/Plant | Percentage of Reduction Based on Control (%) | Risk of Plants Harboring More than 25 Mealybugs (%) |
|---|---|---|---|
| SA (salicylic acid) | 18.1 | 51% | 27% |
| ASM (acibenzolar-S-methyl) | 11.9 | 67% (* p < 0.01) | 13% |
| BABA (β-aminobutyric acid) | 17.1 | 52% | 20% |
| Flacourtia indica (plant extract) | 31.9 | 11% | 40% |
| Hypericum lanceolatum (plant extract) | 23.4 | 35% | 20% |
| MeSA (essential oil of Gaultheria fragrantissima) | 9.9 | 72% (* p < 0.002) | 20% |
| Control | 35.9 | 0% | 53% |
| Treatment | Mean Mealybugs/Plant | Percentage of Reduction Based on Control (%) | Risk of Plants Harboring More than 25 Mealybugs (%) |
|---|---|---|---|
| SA (salicylic acid) | 36.4 | 0% | 32% |
| ASM (acibenzolar-S-methyl) | 8.5 | 76% (* p < 0.001) | 7% |
| BABA (β-aminobutyric acid) | 16.0 | 54% (* p = 0.01) | 7% |
| Flacourtia indica (plant extract) | 75.0 | - | 100% |
| Hypericum lanceolatum (plant extract) | 49.3 | - | 60% |
| MeSA (essential oil of Gaultheria fragrantissima) | 6.9 | 80% (* p < 0.001) | 7% |
| Control | 35.1 | 0% | 46% |
| Direct Soil Application | Transgenerational Application | ||||
|---|---|---|---|---|---|
| Treatment | n (Plants) | Plant Height (cm) | Chlorophyll Level (SPAD) | Plant Height (cm) | Chlorophyll Level (SPAD) |
| p < 0.001 ** | p = 0.28 | p = 0.09 | p < 0.001 ** | ||
| SA (salicylic acid) | 30 | 42 ± 2.2 | 72 | 24 ± 2.2 | 68 |
| ASM (Acibenzolar-S-Methyl) | 15 | 36 ± 2.2 | 71 | 20 ± 2.3 | 61 |
| BABA (β-Aminobutyric acid) | 15 | 42 ± 2.5 | 73 | 26 ± 3.2 | 68 |
| Flacourtia indica (plant extract) | 15 | 45 ± 2.2 * p = 0.002 | 68 | 22 ± 2.6 | 64 |
| Hypericum lanceolatum (plant extract) | 15 | 37 ± 2.2 | 69 | 29 ± 4.2 * | 78 * p = 0.007 |
| MeSA (essential oil of Gaultheria fragrantissima) | 15 | 39 ± 2.2 | 73 | 17 ± 2.0 * | 57 |
| Control | 30 | 36 ± 2.2 | 73 | 23 ±1.9 | 64 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
N’Guessan, L.; Chillet, M.; Chiroleu, F.; Soler, A. Ecologically Based Management of Pineapple Mealybug Wilt: Controlling Dysmicoccus brevipes Mealybug Populations with Salicylic Acid Analogs and Plant Extracts. Horticulturae 2024, 10, 227. https://doi.org/10.3390/horticulturae10030227
N’Guessan L, Chillet M, Chiroleu F, Soler A. Ecologically Based Management of Pineapple Mealybug Wilt: Controlling Dysmicoccus brevipes Mealybug Populations with Salicylic Acid Analogs and Plant Extracts. Horticulturae. 2024; 10(3):227. https://doi.org/10.3390/horticulturae10030227
Chicago/Turabian StyleN’Guessan, Lysa, Marc Chillet, Frédéric Chiroleu, and Alain Soler. 2024. "Ecologically Based Management of Pineapple Mealybug Wilt: Controlling Dysmicoccus brevipes Mealybug Populations with Salicylic Acid Analogs and Plant Extracts" Horticulturae 10, no. 3: 227. https://doi.org/10.3390/horticulturae10030227