
Underneath the Purple Stain
by
 , and
, and
Catarina Pinheiro
1,*,
Ana Zélia Miller
1,2,
Patrícia Vaz
3,
Ana Teresa Caldeira
1 and
Conceição Casanova
3,4 1
Laboratório HERCULES, Herança Cultural, Estudos e Salvaguarda, Largo Marquês de Marialva, 8, 7000-809 Évora, Portugal
2
Instituto de Recursos Naturales y Agrobiología de Sevilla (IRNAS-CSIC), Av. Reina Mercedes 10, 41012 Sevilla, Spain
3
Departamento de Conservação e Restauro, Faculdade de Ciências e Tecnologia, Universidade Nova de Lisboa, Campus Caparica, 2829-516 Caparica, Portugal
4
LAQV—REQUIMTE, Universidade Nova de Lisboa, Campus Caparica, 2829-516 Caparica, Portugal
*
Author to whom correspondence should be addressed.
Heritage 2022, 5(4), 4100-4113; https://doi.org/10.3390/heritage5040212
Submission received: 8 November 2022
/
Revised: 6 December 2022
/
Accepted: 7 December 2022
/
Published: 11 December 2022
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:The Alcobacenses are one of the most important Portuguese collections of Monastic codices. During a campaign study to further advance our knowledge on this important legacy and its state of conservation, a heavily deteriorated codex (ALC.338) stood out due to the presence of purple stains on the lower margins on virtually all folios. In some of them, the purple stains are accompanied by yellow and brown patches as well. Also noticed was the presence of these purple spots on the white tawed leather covering the codex. The codex was analyzed macro and microscopically, chemically and biologically using state-of-the-art technology such as next-generation sequencing. The biological deterioration associated with the appearance of these purple spots arises from the inside out and reveals significant abundance of the bacterial genus Saccharopolyspora sp., which accounted for 86% of the bacterial genera identified. For the fungal population, the most common genus was Malassezia sp. Visual inspection of the sample confirmed the presence of fungal and bacterial biofilms. Pressing questions, comparison to similar studies and future perspectives are also given.
1. Introduction
Founded in 1153 by the mother abbey of Clairvaux, the Alcobaça Monastery likely began producing its own illuminated manuscripts after 1175 and kept its activity until the sixteenth century [1,2]. The Alcobaça Monastery library offers one of the most important collections of illuminated manuscripts from medieval Portugal and shows a strong intertwining between French and Iberian artistic and textual traditions [1]. These were the most important manuscripts monasteries produced, indispensable to the celebration of the Daily Office, around which the lives of the monks were structured. Unlike other abbeys, a large number of these manuscripts have survived at Alcobaça, which presents the possibility of dating them more accurately and thus charting the evolution of materials and techniques of book production within the scriptorium for over more than four centuries. This is especially important in the case of the codices’ bookbinding, which represent one of the surviving collections, in Europe, which is closer to its pristine state and/ or showing several original binding features. It is believed that this will clarify uncertainties about provenance and dating, deepen our knowledge on the technologies applied and used during Middle Ages, as well as reveal specific degradation patterns in the codex which will contribute to define a further care policy for these National treasures currently kept at the National Library of Lisbon. Out of a total of 467 volumes [2] (named the Alcobacenses) opinions may diverge on how many of these still display their original features, as considered by Aires do Nascimento [3] but this number is situated between 64 [4,5] and 80 [3]. The Manuscript Alcobacense 338 (ALC.338), written in the XIII century, is considered one of these codices and it contains the Commentaries of St. Jerome to the Twelve Minor Prophets. It presents a relatively well-preserved white tawed leather cover enclosing 236 parchment folios. The text is written in two columns of 39 lines each, in Latin, with a few words in Greek. At the end of each 8-folium quire there is a Proclamation section. Figure 1, Figure 2 and Figure 3 show some of the details of this particular codex.
Parchment and leather are created from the same source: animal skin. After being removed (cows, calves, sheep, deer and goats were the most common source), the skin was treated with salt to prevent putrefaction [6,7]. The skins would remain in this brine state until they were ready for the next step, also common to both parchment and leather. Calcium hydroxide, in the form of a lime bath [7] was used to aid in the removal of the hair and the skins would stay there for at least eight days in summer and longer in the winter time. The skin would then be placed in a wooden beam and the hair would be scrubbed off mechanically with a blunt dehairing knife [7,8]. The process was performed twice [7] and the skin is left only with the pore mark, the follicle. The way these follicles form a pattern is a feature indispensable to visually decipher the animal origin, in the final product. The skin was then placed on the stretcher, or herse, where it was left to dry under tension [9]. While it was in the stretcher it would go through several dry and wet cycles. The water used to keep it moist was a fresh lime bath in an effort to remove excessive grease and, thus, prevent the ink from running when applied to the final product [10]. To improve its smoothness and whiteness, parchment would also be treated with other substances such as flour, milk, egg whites, and pumice [7,10].
Parchment has served as a writing material for centuries, and many examples from the Middle Ages, such as this one, have reached us. Despite its endurance, it is not immune to deterioration from exposure to environmental factors such as light, elevated temperatures, humidity or pollutants [10]. Because of its organic nature, microorganisms that cause both chemical and structural modifications are also frequently present and are held responsible for many of the chromatic changes parchment folios or codices display [10]. Purple stains, such as the ones depicted in Figure 1b and Figure 2, are examples of the effect that microorganisms can have on parchment. On the ALC.338, it is not just the parchment that is affected, as purple stains (albeit in much lesser extent) can also be observed on the white tawed leather protecting the codex (Figure 3).
The fact that microorganisms produce pigmented metabolites has led the research on chromatic changes observed in cultural heritage [11,12,13,14,15]. Throughout the years, explaining the purple stains was the driving force behind remarkable efforts [16,17].
However, because biodeterioration was essentially assessed by traditional culturing methods until 20 years ago, the purple stains' origin remained a mystery, as no common ground could be found among the different studies being performed. Even after the successful isolation of a possible causative agent, the production of pigments that resembled the ones observed on artefacts has fallen short of expectations [18].
A large step towards understanding this phenomenon and what causes it has been recently achieved through the studies of Piñar [10], Migliore [18], and Perini [19,20]. Next-generation sequencing technologies, such as the ones used in these studies, were able to provide information on the communities present on parchment and offer a possible common ground between purple stained parchment manuscripts and hence develop a theory on the appearance of these particular spots. Just like layers on top of each other, this theory presents an evolving biological community, one that thrives from the inside of the parchment towards the surface.
From the collection of Alcobacenses, ALC.338 is one of a few presenting this problem. To determine whether or not the succession theory proposed by previous studies was relatable to the communities encountered in this codex, we set out to discover what was underneath the exuberant purple stains displayed in this manuscript. A series of methodologies were applied and the results were discussed and compared with recent literature.
2. Materials and Methods
2.1. Visual Inspection
Visual inspection of the parchment and leather present in ALC.338 was performed by naked eye, with magnifying lenses, with a portable microscope (Pancellent, Inskam, Shenzhen, China) and using a stereomicroscope (Stereo microscope Leica M205C, Wetzlar, Germany). This first step is valuable to ascertain the quality of the materials used and the current state of conservation. Because the parchment appears severely damaged and even careful manipulation results in the detachment of tiny fragments, these were taken to the lab for scanning electron microscopy with energy dispersive X-ray spectroscopy, a valuable technique to identify current and former biological colonization. The SEM equipment used was a Hitachi 3700 N scanning electron microscope (Hitachi, Tokyo, Japan) interfaced with a Quantax Energy Dispersive X-Ray Spectroscopy (EDS) microanalysis system (Bruker AXS GmbH, Karlsruhe, Germany). This system is equipped with a Bruker AXS XFlash® Silicon Drift Detector (EM-EDS Hitachi S3700N).
2.2. Fungal Communities
Sterile swabs were used to sample the stained and unstained areas of ALC.338. The swabs were transported in a falcon tube and the cotton tips were cut, placed in an Eppendorf with saline solution, and vortexed vigorously. Two hundred and fifty microliters of the saline solution was pipetted into the culture medium used: Malt Extract Agar (MEA) and Red Bile (RB). The plates, in triplicate, were placed for 7 days at 27 °C and monitored. Fungal cultures were isolated for identification [21].
2.3. DNA Extraction and Sequencing
Parchment microsamples (two, 1.5 mm × 2 mm) were taken from the ALC.338 for microbiological content analysis. A swab was also used on the affected and unaffected parchment samples. The leather cover presenting purple stains presented a more cohesive state and only swab samples were taken from its affected and unaffected areas. Upon arrival at the lab, all samples were immediately frozen at −80 °C. The DNEasy PowerLyzer PowerSoil Kit from Qiagen (Hilden, Germany) was used as instructed but three freeze-thaw cycles (−80 °C/65 °C) were introduced after the C1 reagent addition. A fluorometer (Quantus Fluorometer, Promega, Walldorf, Germany) was used to determine the DNA amount retrieved after extraction. Positive results were sent to a specialized facility for Next Generation Sequencing (STABVIDA, Portugal) for both bacterial and fungal communities. For this purpose, the V3–V4 fragment of the 16S rRNA gene and the ITS1 region were amplified using region-specific primers and sequenced in an MiSeq platform to produce 2 × 300 bp paired-end reads. The raw reads were deposited into the NCBI Sequence Read Archive (SRA) database under accession number PRJNA899037.
3. Results and Discussion
3.1. General Quality and State of Conservation for Leather and Parchment
The leather bindings exhibit a greater cohesive state of conservation than the parchment folios. The white leather is presumed to be originated from calf and sheep (Figure 4a,b), [22] and presents marks that were probably endured during manufacture (Figure 1, the scratches on the lower part) and others during use (the loss of the bosses and the general staining from pollution and soot).
The leather binding displays stiffness in the lower and upper edges, a condition that often affects this type of material. When the codex is opened, the purple stains appear visible in the turn-ins both in the front and in the back of the codex. The stains visible in the leather are not a result of contamination from the adjacent parchment, as the stains are not always superimposable (see Figure 3).
As mentioned, the parchment is in much poorer condition. The folios are visibly deteriorating and this is particularly relevant in the lower section of each folio. As expected, the flesh side of each folio presents a much heavier degradation with a wider dissemination of purple stains. The folios are fragile and tiny fragments are easily detached, even with careful manipulation. As confirmed in previous studies [19], there is also an inhibitory effect from the inks used in the text and the purple stains seem to go around them. However, the authors also noticed a relatively sound and purple-free area going 2 mm–5 mm inwards from the lateral margins in a significant portion of the affected folia. Could a decrease in water activity (Aw) play a role here, as this area was in closer contact with the environment surrounding the codex and was relatively exempt from manipulation?
Unstained and stained areas of the parchment were observed under the stereomicroscope (Figure 5). These images show that even on the side less affected by the chromatic changes (left), the immediate layer below the surface looks tainted. Figure 5b shows this clearly: there seems to be a superficial, still whitish layer covering a much more degraded one. It is not clear, however, if all the chromatic changes are a result of exposing a tainted core or if they can also be the result of pigmented metabolites produced by superficial microbiological contaminants that have, in time, been advancing towards the core of the parchment. This hypothesis must be considered since there are top layers displaying purple hues.
3.2. Chemical Profile by X-ray Energy Dispersive Spectroscopy
Stained and unstained samples of parchment were analyzed by SEM-EDS. Figure 6 and Figure 7 display the results obtained in two of the samples areas—one less damaged than the other (respectively).
Comparing Figure 6 and Figure 7, the unstained area shows a more cohesive state than the purple stained area, as expected (see Figure 5). The fibers on these last samples show signs of breakage and disintegration. Figure 8, Figure 9 and Figure 10 demonstrate these differences in an even greater fashion.
Regarding EDS, and as has happened in previous studies [10], all samples displayed the following elements: Na, Mg, Al, Si, P, S, Cl and Ca. This last element can be attributed to the lime treatments to which parchment was submitted for the removal of the hair and also by the use of chalk to perfect the final product rendering it suppler and whiter. Sodium (Na) and chlorine (Cl) are related to the initial phases the skin goes through to prevent putrefaction. Magnesium (Mg) and potassium (K) can also be implicated in this first phase. Together with iron (Fe), their origin can, however, also be traced back to the skin itself. Sulphur (S) and phosphorous (P) are also traceable to the original skin. Unlike in previous studies [23] we did not notice an increment in the amounts registered for these two elements in the damaged areas. Si and Al are present in the pumice stones used to abrade both surfaces of the skin.
Despite the known fragilities of using EDS for quantification (given the heterogeneous density of the matrix and the use of un-polished samples) [23,24], damaged areas tend to show a general lower content of the elements, suggesting that the microbial communities may leach these components off. Iron, however, was only present in the unaffected area, a fact also noticed in other studies [10].
3.3. Biological Communities by Visual Inspection—Scanning Electron Microscopy
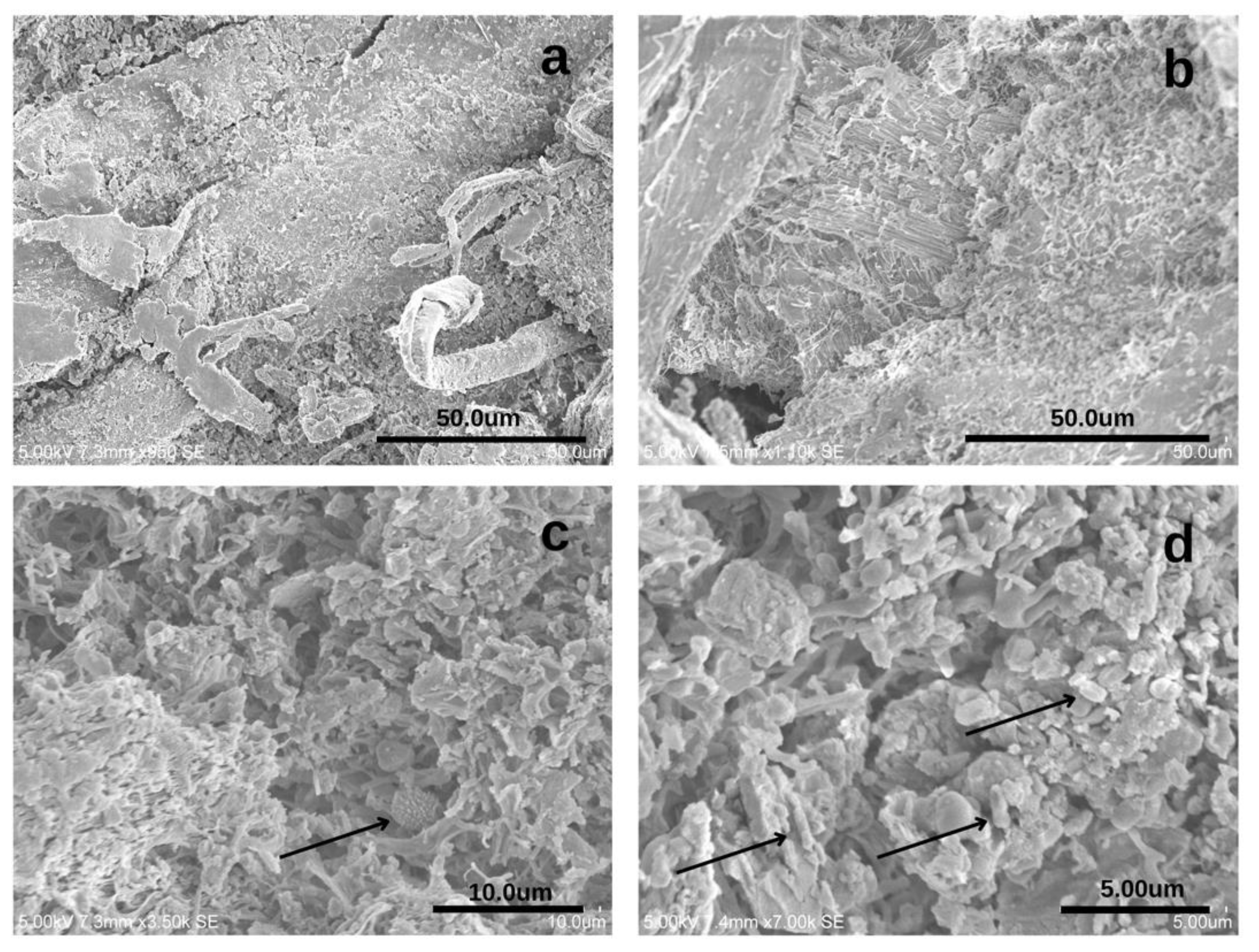
The gold coating performed on parchment samples increases the ability to identify biological markers such as spores and bacterial cells. Figure 8 compiles images taken from undamaged areas, and the difference between Figure 8a,b is clear. In Figure 8c, it is possible to detect a spiked structure resembling Cladosporium sp., a common airborne contaminant [15,21]. The lack of other defining structures makes it difficult to confirm this hypothesis. The size of the structures pointed out in Figure 8b is concurrent to the presence of bacterial cells. A DNA-based analysis (see Section 3.5) points to the presence of Saccharopolyspora spp. and Malassezia spp. (a yeast-like fungus) and both phenotypically fit within the identified structures. However, the lack of other defining structures makes it hard to confirm these assumptions.
3.4. Fungal Cultivated Communities
The samples were qualitatively assessed and no significant differences were encountered in the unstained and stained areas. Aspergillus spp. were present in both sample types, as was Fusarium sp. and Paecilomyces sp. Penicillium spp. and Ulocladium sp. were only identified in media culture inoculated with the unstained extract, and Cladosporium sp. was identified solely in the extract coming from the stained area.
3.5. Biological Communities by NGS
After extraction, only one sample (corresponding to a microfragment of the affected parchment) delivered enough DNA for sequencing and was sent for NGS. In fact, the amount of sample that it is possible to collect from this type of artifact is a limiting factor.
For bacteria, the sample generated 187,248 raw sequence reads. After denoising, a total of 90 unique features (OTUs) were identified and a taxonomical classification was performed, using the SILVA database (release 138 QIIME) trained using scikit-learn. For fungi, the sample generated 96,334 raw sequence reads and a total of 26 unique features (OTUs) were identified using the UNITE database for taxonomical identification.
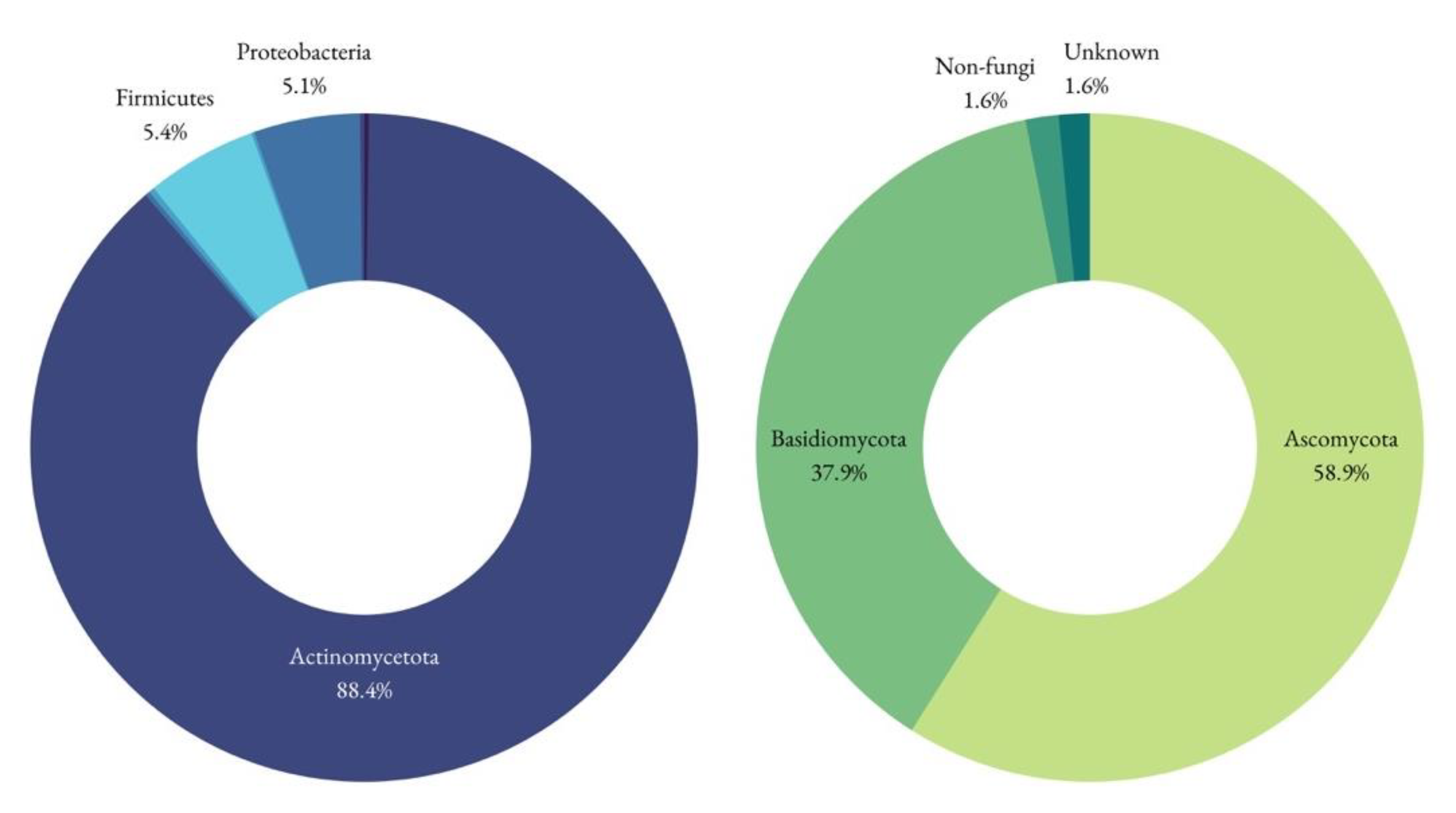
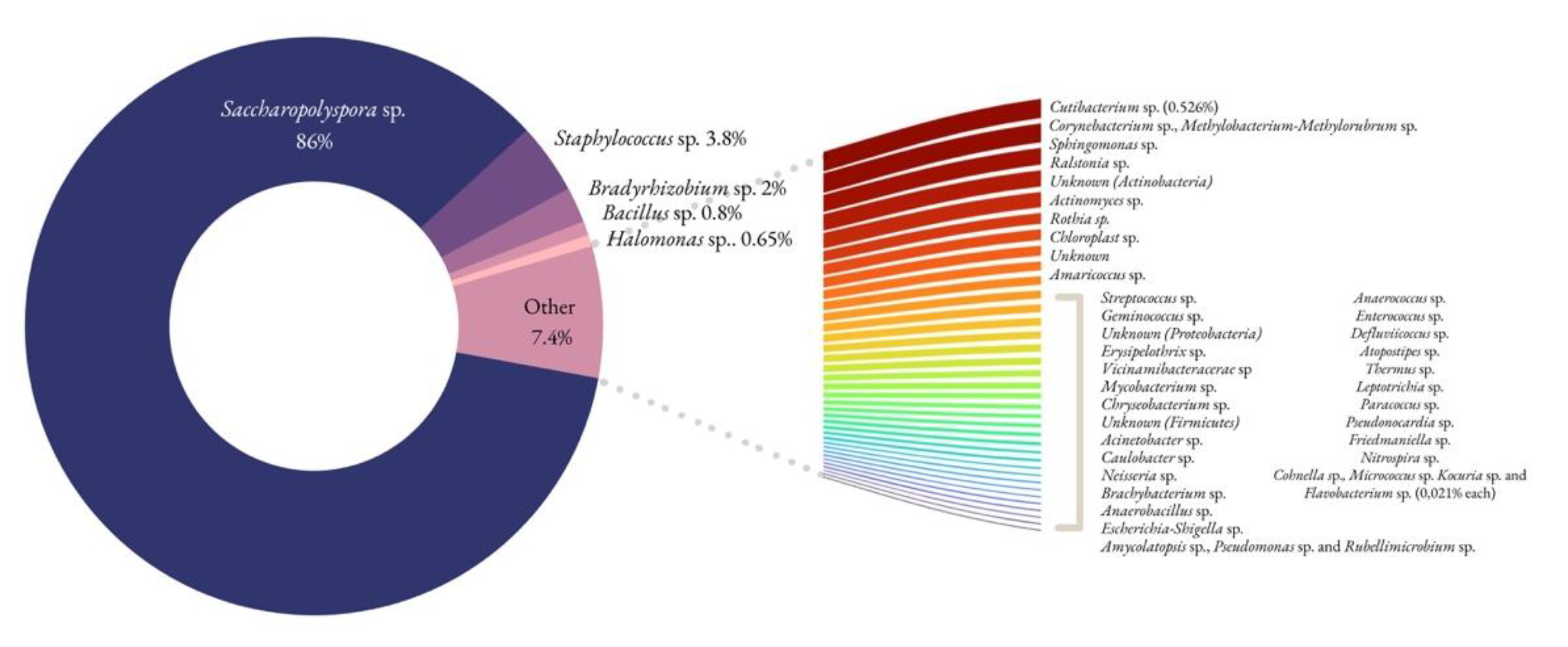
In the damaged area, the only one where DNA could be amplified, the analysis revealed the presence of Actinomycetota (88.4%), Firmicutes (5.4%), Proteobacteria (5.1%), Cyanobacteria, Acidobacteriota, Bacteroidota, and Plantomycetota. Actinomycetota were the predominant phylum present, as happens in similar studies [10]. Saccharopolyspora accounted for the majority of the identified genera (86%), and its presence is in accordance with previous studies [10,18,25] as well. This particular genus is known for its relevance in the pharmaceutical industry and is frequently reported in coastal sediments and marine organisms [26]. It grows well within 6–18% NaCl concentrations, and its absence in the medium inhibits the bacterial development [10]. Still in the Actinomycetota phylum, but lesser represented, are species belonging to the Cutibacterium and Corynebacterium genera, widely distributed in nature in the microbiota of animals, humans included. Hence, one cannot rule out the presence of human contamination, but this genus could very well be present on the animal skin transformed into parchment. The Corynebacteriales order, of which each Corynebacterium spp. and Mycobacterium spp. are part of, are known producers of bacterioruberin, a carotenoid pigment found in documentation affected by purple spots [27]. This order has, however, been found to be present in both damaged and undamaged areas [27]. The Micrococcales order (represented by Brachybacterium spp. and Rothia spp. in the present study) also produces bacterioruberin [27].
The Firmicutes are mainly represented by Staphylococcus spp. (4% of the total bacterial microbiota) and the sporulating Bacillus sp. Among this last genus are species with high proteolytic activity that may be responsible for the deterioration of parchment [10,28,29] and have been identified in similar studies [10]. This genus accounts for circa 1% of the total bacterial microbiota. Streptococcus spp. and Erysipelotrix spp. are also human and animal commensals and possible pathogens. These four genera can be attributed to human manipulation, and such natural contaminants have been identified in other studies [30].
The third largest phylum is the Proteobacteria, divided in alpha and gamma proteobacteria. Bradyrhizobium spp. and Methylobacterium spp. have been identified as contaminants of DNA extraction kit reagents and ultrapure water systems, which may lead to its erroneous appearance in microbiota or metagenomic datasets [23]. Due to the low OTUs obtained, this is a hypothesis that must be considered. Methylobacterium spp. has, however, appeared in similar studies [10], and the newly attributed genus Methylorubrum, to which some of the species of Methylobacterium have migrated [31,32], deserves further attention. The vast majority of species in both these genera are pink-pigmented, and the pigment is of carotenoid origin [32]. Some of them are inhibited by the presence of NaCl when above 2%, which may mean that these would only thrive in areas where the salt concentration was lower, probably the more superficial ones. Another alpha-proteobacteria is Sphingomonas spp., a Gram-negative, rod-shaped, chemoheterotrophic and strictly aerobic bacteria which typically produces yellow-pigmented colonies [33].
The gammaproteobacteria Halomonas spp. also benefits from a rich NaCl environment, as happens with the Saccharopolyspora spp. and has been identified in previous studies, although not all of them in parchment [10,34]. Both may be responsible for the purple stains as a result of the very common pigmentation of these species. Contamination by halophile bacteria such as these can come from salt used for preservation, [10,28] and their presence has been integrated in a microbial succession theory [19].
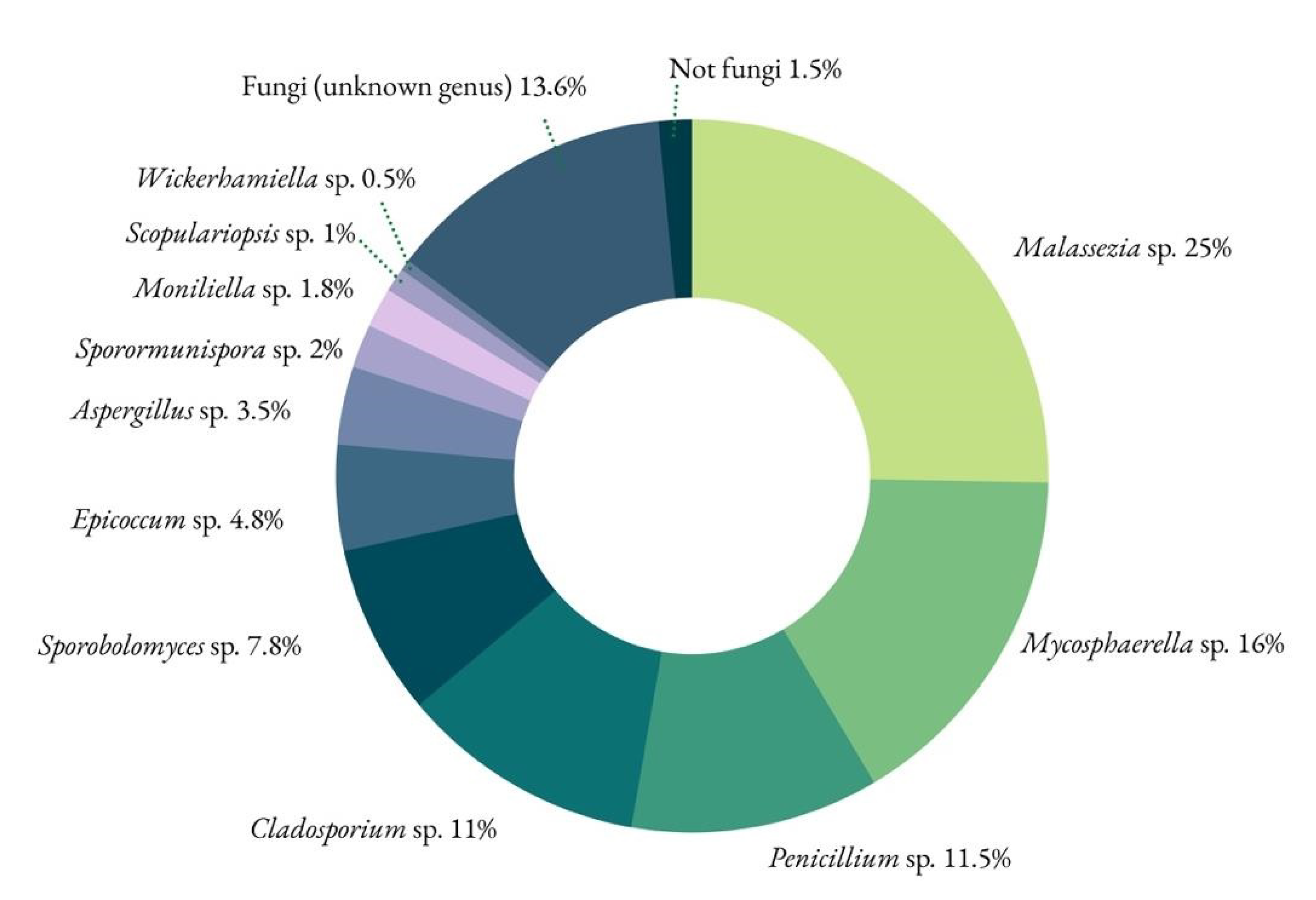
For fungi, the identified phyla were Basidiomycota and Ascomycota. The Viridiplantae kingdom was also identified using the ITS1 primers, but this can be considered a non-specific amplification, as the amplified region is somewhat similar between fungi and plants [35] (Figure 11).
Malassezia spp. yeasts belong to the resident microbiota of animal (and human) skin, and among the lipid-dependent Malassezia species, the synthesis of pigments and fluorochromes is a particular metabolic feature of Malassezia furfur when tryptophan is the main nitrogen source [36]. There is no tryptophan in the sequence of amino-acids that composes collagen, but there is plenty of tryptophan in the mixture added to the parchment as finishing touches to increase its quality (milk, eggs and flour). This genus has been identified recently in parchment [30].
The fungal genus Mycosphaerella has evolved as symbionts, endophytes and saprophytes, with particular implications for crops, being a foliicolous plant pathogen [37].
Penicillium sp., Cladosporium sp., Epicoccum sp. and Aspergillus sp. are ubiquitous environmental fungi and were expected to show up in such a study. Because they are present in so many cultural heritage studies in paper based, archival, and museum collections [15] and have not been associated to such a particular type of biodeterioration pattern, they are not the most likely causative agents.
Sporobolomyces sp.—as Malassezia sp.—is also a basidiomycetous yeast. It can produce carotenoid pigments ranging from yellowish to red that can be visualized with individual colonies [38,39].
The study performed on the ALC.338 and its purple stain afflicted parchment folia has identified a complex microbiologic profile. The NGS technique, despite delivering a relatively low number of OTUs, dramatically improves the results obtained through classic culturing methods, considered ineffective to display the full microbiological profile [19]. The fungal community identified through inoculation rendered only a display of common airborne fungi, although some of them, such as Fusarium sp., do have the ability to produce purple pigmentation [21]. In fact, many are the biological contaminants with metabolites capable of inducing chromatic changes. Some of them—in the Micrococcales and Corynebacteriales order, for instance, identified in the present study, can produce bacterioruberin, identified in previous studies to be one of the components of the purple stains [18,19]. Therefore, could there be more than one mechanism present in the formation of these purple spots? In a document profusely affected by several types of stains—varying in color and size—it is possible to have apparently similar purple hues coming from distinct sources [10].
Similar studies on purple stains have identified Archaeobacteria in parchment affected and unaffected by purple stains [27,40] but our study, as happened with others using molecular biology methodologies [10,18], did not deliver such results. This can be due to a true absence or the still very difficult process of acquiring a representative biological sample from tiny fragments of parchment. Halobacterium salinarum has been considered the original contaminant in this succession model leading to the development of the purple spots [19,27]. This particular Archae has also been identified in other cultural heritage settings, also associated with a rosy outcome [41]. Following H. salinarum are the marine bacteria [19,27] that are also brought along during the process of brining—a treatment used to prevent microbiological attacks before the hide could be transformed into leather or parchment [10,42]. These marine bacteria, Halomonas spp., a gamma-Proteobacteria, are confirmed in the present study. Finally, the third step in the ladder is the Actinobacteria, Saccharopolyspora sp., which has been present in several studies [10,18,25,30] and occupies a large proportion of the bacterial genera encountered in our sample.
There are still many questions to be answered. One of them refers to incidence. Assuming that a large proportion of hides were pre-treated with marine salt, why have the purple stains only appeared in very few codices (so far, we have identified seven out of about 60, as the work is ongoing) from the same time period? In general, the Alcobacenses have been kept as a whole for centuries, but between the extinction of the Religious Orders and the establishment of these codices in Lisbon, some may have been diverted from the group, as has sadly happened to other collections [43,44], and their whereabouts could be responsible for their present state. We do know that when in Lisbon, the collection was kept under strenuous conditions [43,45], but if this was the trigger, then only a few books have been contaminated with the original Bacteria and Archaeobacteria. Can alum tawed leather suffer from the same fate? The pre-treatment was similar [42], but the process of transformation may render it more stable and able to better hamper biological attack.
4. Conclusions
Although it is still not possible to determine the definitive origin of the purple stains, this particular study reinforces a succession theory: it all begins with the salt used to preserve the skins and with this salt come marine bacteria, such as halophilic Proteobacteria and even Actinobacteria. In some of the studies performed on stained parchment, Archaeobacteria would arrive and thrive prior to these two contaminants. Our study does not confirm their presence, but for the reasons presented before, one cannot discard them completely. The elements encountered in larger quantities are the ones that developed later on: Firmicutes and Actinobacteria (Saccharopolyspora sp.), followed by the fungal contaminants.
Further studies and improvements on the DNA extraction process are underway in order to determine if the first step of the ladder is, in fact, present or not, and if ALC.338 can be another confirmation of the model. The authors acknowledge, however, that the longevity of this particular document and its long history of use may translate into an irreversibly damaged Archae DNA or one that is so underrepresented that it will not show up despite future efforts. The study on the alum-tawed leather binding of ALC.338 is also underway.
Author Contributions
Conceptualization, C.P.; formal analysis, C.P., A.Z.M. and P.V.; methodology, C.P., A.T.C. and A.Z.M.; investigation, C.P., C.C. and P.V.; writing—original draft preparation, C.P.; writing—C.P., A.Z.M., C.C., A.T.C. and P.V.; visualization, C.P.; project administration, C.C.; funding acquisition, C.C. and A.T.C.; resources, A.T.C. All authors have read and agreed to the published version of the manuscript.
Funding
Catarina Pinheiro is under a researcher contract with the HERCULES Laboratory, Évora University (CEECIND/02598/2017) and gratefully acknowledges the FCT/MCTES national support through the UIDB/04449/2020 and UIDP/04449/2020. C.C. acknowledges FCT/MCTES national support through the UIDB/50006/2020 and UIDP/50006/2020. A.Z.M contribution is supported by the Spanish Ministry of Science and Innovation (MCIN) under the Ramón y Cajal contract (RYC2019-026885-I), the CEECIND/01147/2017 research contract from FCT and the intramural project PIE_20214AT021 from the Spanish National Research Council (CSIC). The project under which this work was developed was also funded by FCT through the grant PTDC/ART-HIS/29522/2017.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
References
- Miguélez Cavero, A.; Melo, M.J.; Miranda, M.A.; Castro, R.; Casanova, C. Beatus manuscripts under the microscope: The Alcobaça Beatus and the Iberian Cistercian tradition revisited. J. Mediev. Iber Stud. 2016, 8, 217–251. [Google Scholar] [CrossRef] [Green Version]
- Barreira, C. Approaches to the study of a fourteenth-century breviary from the cistercian abbey of Alcobaça. Citeaux 2017, 68, 249–276. [Google Scholar]
- Nascimento, A.A.; Diogo, A.D. Encadernação Portuguesa Medieval—Alcobaça [Medieval Portuguese Bookbinding: Alcobaça]; Imprensa Nacional da Casa da Moeda: Lisbon, Portugal, 1984; 109p. [Google Scholar]
- Tourais, A. Development of a Material Characterization Method for the Medieval Codices of the Collection from the Monastery of Alcobaça; Faculdade de Ciências e Tecnologia da Universidade Nova de Lisboa: Lisboa, Portugal, 2020; Available online: http://hdl.handle.net/10362/114642 (accessed on 17 October 2022).
- Tourais, A.; Casanova, C.; Barreira, C.F. Filling the gap: New approaches to medieval bookbinding studies. J. Mediev. Iber. Stud. 2022, 14, 109–126. [Google Scholar] [CrossRef]
- Larsen, R. Introduction to damage and damage assessment of parchment. In Improved Damage Assessment of Parchment (IDAP), Assessment, Data Collection and Sharing of Knowledge; Larsen, R., Ed.; Office for Official Publications of the European Communities: Luxembourg, 2007; pp. 17–21. [Google Scholar]
- Hallebeek, P.B. Guidelines for the Conservation of Leather and Parchment; Koninklijke Bibliotheek: Amsterdam, The Netherlands, 1997. [Google Scholar]
- Thomas, S. Leathermaking in the Middle Ages. In Leather Manufacture through the Ages; Thomas, S., Clarkson, L., Thomson, R., Eds.; Arkle Print Limited: Northampton, UK, 1983; pp. 1–10. [Google Scholar]
- Clarkson, C. Rediscovering parchment: The nature of the beast. Pap. Conserv. 1992, 16, 5–26. [Google Scholar] [CrossRef]
- Piñar, G.; Sterflinger, K.; Pinzari, F. Unmasking the measles-like parchment discoloration: Molecular and microanalytical approach. Environ. Microbiol. 2015, 17, 427–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vijayakumar, S. Role of Cyanobacteria in Biodeterioration of Historical Monuments—A Review. BMR Microbiol. 2014, 1, 1–13. [Google Scholar]
- Warscheid, T.; Braams, J. Biodeterioration of stone: A review. Int. Biodeterior. Biodegrad. 2000, 46, 343–368. [Google Scholar] [CrossRef]
- Sterflinger, K.; Piñar, G. Microbial deterioration of cultural heritage and works of art—Tilting at windmills? Appl. Microbiol. Biotechnol. 2013, 97, 9637–9646. [Google Scholar] [CrossRef] [Green Version]
- Villa, F.; Wu, Y.-L.; Zerboni, A.; Cappitelli, F. In Living Color: Pigment-Based Microbial Ecology at the Mineral–Air Interface. Bioscience 2022, 72, 1156–1175. [Google Scholar] [CrossRef]
- Pinheiro, A.C.; Sequeira, S.O.; Macedo, M.F. Fungi in archives, libraries, and museums: A review on paper conservation and human health. Crit. Rev. Microbiol. 2019, 45, 686–700. [Google Scholar] [CrossRef]
- Karbowska-Berent, J.; Strzelczyk, A. The Role of Streptomycetes in the Biodeterioration of Historic Parchment; Copernicus University Press: Torun, Poland, 2000. [Google Scholar]
- Gallo, F.; Strzelczyk, A. Indagine preliminare sulle alterazioni microbiche della pergamena. Bollett. Ist. Centr. Patologia Libro “Alfonso Gallo.” 1971, 30, 71–87. [Google Scholar]
- Migliore, L.; Thaller, M.C.; Vendittozzi, G.; Mejia, A.Y.; Mercuri, F.; Orlanducci, S.; Rubechini, A. Purple spot damage dynamics investigated by an integrated approach on a 1244 A.D. parchment roll from the Secret Vatican Archive. Sci. Rep. 2017, 7, 9521. [Google Scholar] [CrossRef] [PubMed]
- Perini, N.; Mercuri, F.; Orlanducci, S.; Thaller, M.C.; Migliore, L. The Integration of Metagenomics and Chemical Physical Techniques Biodecoded the Buried Traces of the Biodeteriogens of Parchment Purple Spots. Front. Microbiol. 2020, 11, 598945. [Google Scholar] [CrossRef] [PubMed]
- Perini, N.; Mercuri, F.; Thaller, M.C.; Orlanducci, S.; Castiello, D.; Talarico, V.; Migliore, L. The Stain of the Original Salt: Red Heats on Chrome Tanned Leathers and Purple Spots on Ancient Parchments Are Two Sides of the Same Ecological Coin. Front. Microbiol. 2019, 10, 2459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samson, R.A.; Hoekstra, E.S.; Frisvad, J.C.; Filtenborg, O. Introduction to Food- and Airborne Fungi, 7th ed.; Samson, R.A., Hoekstra, E.S., Frisvad, J.C., Filtenborg, O., Eds.; Centraalbureau Voor Schimmelculture: Utrecht, The Netherlands, 2010; 390p. [Google Scholar]
- Pinheiro, C.; Faustino, M.; Tourais, A.; Gonçalves, C.; Casanova, C. The Medieval Leather Bookbinding of Santa Maria de Alcobaça: Technological Approaches. In Medieval Europe in Motion V—Materialities and Devotion (5th–15th Centuries); Barreira, C., Martins, D., Fontes, J.L., Farelo, M., Eds.; 2022; in press. [Google Scholar]
- Pinzari, F.; Cialei, V.; Piñar, G. A case study of ancient parchment biodeterioration using variable pressure and high vacuum scanning electron microscopy. In Historical Technology, Materials and Conservation: SEM and Microanalysis; Archetype Books: London, UK, 2012; pp. 93–99. [Google Scholar]
- Cicero, C.; Pinzari, F.; Mercuri, F. 18th Century knowledge on microbial attacks on parchment: Analytical and historical evidence. Int. Biodeterior. Biodegrad. 2018, 134, 76–82. [Google Scholar] [CrossRef]
- Teasdale, M.D.; Fiddyment, S.; Vnouček, J.; Mattiangeli, V.; Speller, C.; Binois, A.; Carver, M.; Dand, C.; Newfield, T.P.; Webb, C.C.; et al. The York Gospels: A one thousand year biological palimpsest. R. Soc. Open Sci. 2017, 4, 170988. [Google Scholar] [CrossRef] [Green Version]
- Sayed, A.; Abdel-Wahab, N.M.; Hassan, H.; Abdelmohsen, U. Saccharopolyspora: An underexplored source for bioactive natural products. J. Appl. Microbiol. 2020, 128, 314–329. [Google Scholar] [CrossRef] [Green Version]
- Migliore, L.; Perini, N.; Mercuri, F.; Orlanducci, S.; Rubechini, A.; Thaller, M.C. Three ancient documents solve the jigsaw of the parchment purple spot deterioration and validate the microbial succession model. Sci. Rep. 2019, 9, 1623. [Google Scholar] [CrossRef] [Green Version]
- Falkiewicz-Dulík, M. Leather and Leather Products. In Handbook of Material Biodegradation, Biodeterioration, and Biostablization, 2nd ed.; ChemTec Publishing: Toronto, ON, Canada, 2015; pp. 133–256. [Google Scholar] [CrossRef]
- Lech, T. Evaluation of a Parchment Document, the 13th Century Incorporation Charter for the City of Krakow, Poland, for Microbial Hazards. Appl. Environ. Microbiol. 2016, 82, 2620–2631. [Google Scholar] [CrossRef] [Green Version]
- Cappa, F.; Piñar, G.; Brenner, S.; Frühmann, B.; Wetter, W.; Schreiner, M.; Engel, P.; Miklas, H.; Sterflinger, K. The Kiev Folia: An interdisciplinary approach to unravelling the past of an ancient Slavonic manuscript. Int. Biodeterior. Biodegrad. 2022, 167, 105342. [Google Scholar] [CrossRef]
- Green, P.N.; Ardley, J.K. Review of the genus Methylobacterium and closely related organisms: A proposal that some Methylobacterium species be reclassified into a new genus, Methylorubrum gen. nov. Int. J. Syst. Evol. Microbiol. 2018, 68, 2727–2748. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Wang, M.Y.; Khan, N.; Tan, L.L.; Yang, S. Potentials, Utilization, and Bioengineering of Plant Growth-Promoting Methylobacterium for Sustainable Agriculture. Sustainability 2021, 13, 3941. [Google Scholar] [CrossRef]
- Balkwill, D.L.; Fredrickson, J.K.; Romine, M.F. Sphingomonas and Related Genera; Pacific Northwest National Lab. (PNNL): Richland, WA, USA, 2003. [Google Scholar]
- Rölleke, S.; Muyzer, G.; Wawer, C.; Wanner, G.; Lubitz, W. Identification of bacteria in a biodegraded wall painting by denaturing gradient gel electrophoresis of PCR-amplified gene fragments coding for 16S rRNA. Appl. Environ. Microbiol. 1996, 62, 2059–2065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hollingsworth, P.M. Refining the DNA barcode for land plants. Proc. Natl. Acad. Sci. USA 2011, 108, 19451–19452. [Google Scholar] [CrossRef] [Green Version]
- Hossain, H.; Landgraf, V.; Weiss, R.; Mann, M.; Hayatpour, J.; Chakraborty, T.; Mayser, P. Genetic and biochemical characterization of Malassezia pachydermatis with particular attention to pigment-producing subgroups. Med. Mycol. 2007, 45, 41–49. [Google Scholar] [CrossRef] [Green Version]
- Crous, P.W.; Kang, J.-C.; Braun, U. A Phylogenetic Redefinition of Anamorph Genera in Mycosphaerella Based on ITS rDNA Sequence and Morphology. Mycologia 2001, 93, 1081. [Google Scholar] [CrossRef]
- Kot, A.M.; Kieliszek, M.; Piwowarek, K.; Błażejak, S.; Mussagy, C.U. Sporobolomyces and Sporidiobolus—Non-conventional yeasts for use in industries. Fungal Biol. Rev. 2021, 37, 41–58. [Google Scholar] [CrossRef]
- Pfaller, M.A.; Diekema, D.J.; Merz, W.G. Infections caused by non-Candida, non-Cryptococcus yeasts. Clin. Mycol. 2009, 10, 251–270. [Google Scholar] [CrossRef]
- Piñar, G.; Tafer, H.; Schreiner, M.; Miklas, H.; Sterflinger, K. Decoding the biological information contained in two ancient Slavonic parchment codices: An added historical value. Environ. Microbiol. 2020, 22, 3218–3233. [Google Scholar] [CrossRef]
- Rölleke, S.; Witte, A.; Wanner, G.; Lubitz, W. Medieval wall paintings—A habitat for archaea: Identification of archaea by denaturing gradient gel electrophoresis (DGGE) of PCR-amplified gene fragments coding for 16S rRNA in a medieval wall painting. Int. Biodeterior. Biodegrad. 1998, 41, 85–92. [Google Scholar] [CrossRef]
- Vest, M. White tawed leather—Aspects of conservation. In Proceedings of the 9th International Congress of IADA, Copenhagen, Denmark, 16–21 August 1999; pp. 67–72. [Google Scholar]
- Casanova, M.C. De Artífice a Cientista. Evolução da Conservação e do Estatuto Profissional do Conservador Restaurador de Documentos Gráficos no AHU (1926–2006); Faculdade de Ciências e Tecnologia, Universidade Nova de Lisboa: Lisboa, Portugal, 2012; Available online: http://hdl.handle.net/10362/8777 (accessed on 17 October 2022).
- Caldas, J. História de um Fogo-Morto. Subsídios para uma História Nacional. 1258–1848. Vianna do Castelo; Livraria Chardron: Porto, Portugal, 1903. [Google Scholar]
- Proença, R. A Biblioteca Nacional: Breves Noções Históricas e Descritivas; Publicações da Biblioteca Nacional: Lisbon, Portugal, 1918; Volume 1, pp. 7–57. [Google Scholar]
Figure 1.
ALC.338 white tawed leather. (a) Stains and scratches probably inflicted during the transformation of animal skin into leather. The codex’s cover is 330 × 450 × 145 mm and some of its original features, such as the copper alloy bosses, are missing. The dimensions of the core section are 307 × 425 mm. (b) Intense biodeterioration by insects and microorganisms observed after opening the codex. The lower sections of the parchment are the most affected and the corners must have been so damaged that they were eventually cut out.
Figure 1.
ALC.338 white tawed leather. (a) Stains and scratches probably inflicted during the transformation of animal skin into leather. The codex’s cover is 330 × 450 × 145 mm and some of its original features, such as the copper alloy bosses, are missing. The dimensions of the core section are 307 × 425 mm. (b) Intense biodeterioration by insects and microorganisms observed after opening the codex. The lower sections of the parchment are the most affected and the corners must have been so damaged that they were eventually cut out.

Figure 2.
(a) Development of microbial agents on the flesh side. The colours displayed are not just purple in nature, but pink, brown and yellow hues can also be found. In this folio the lead pen used to define the lines displays antiseptic activity, as microbial growth is far less exuberant over these lines. (b) Some of the stains display a nucleus with a halo.
Figure 2.
(a) Development of microbial agents on the flesh side. The colours displayed are not just purple in nature, but pink, brown and yellow hues can also be found. In this folio the lead pen used to define the lines displays antiseptic activity, as microbial growth is far less exuberant over these lines. (b) Some of the stains display a nucleus with a halo.

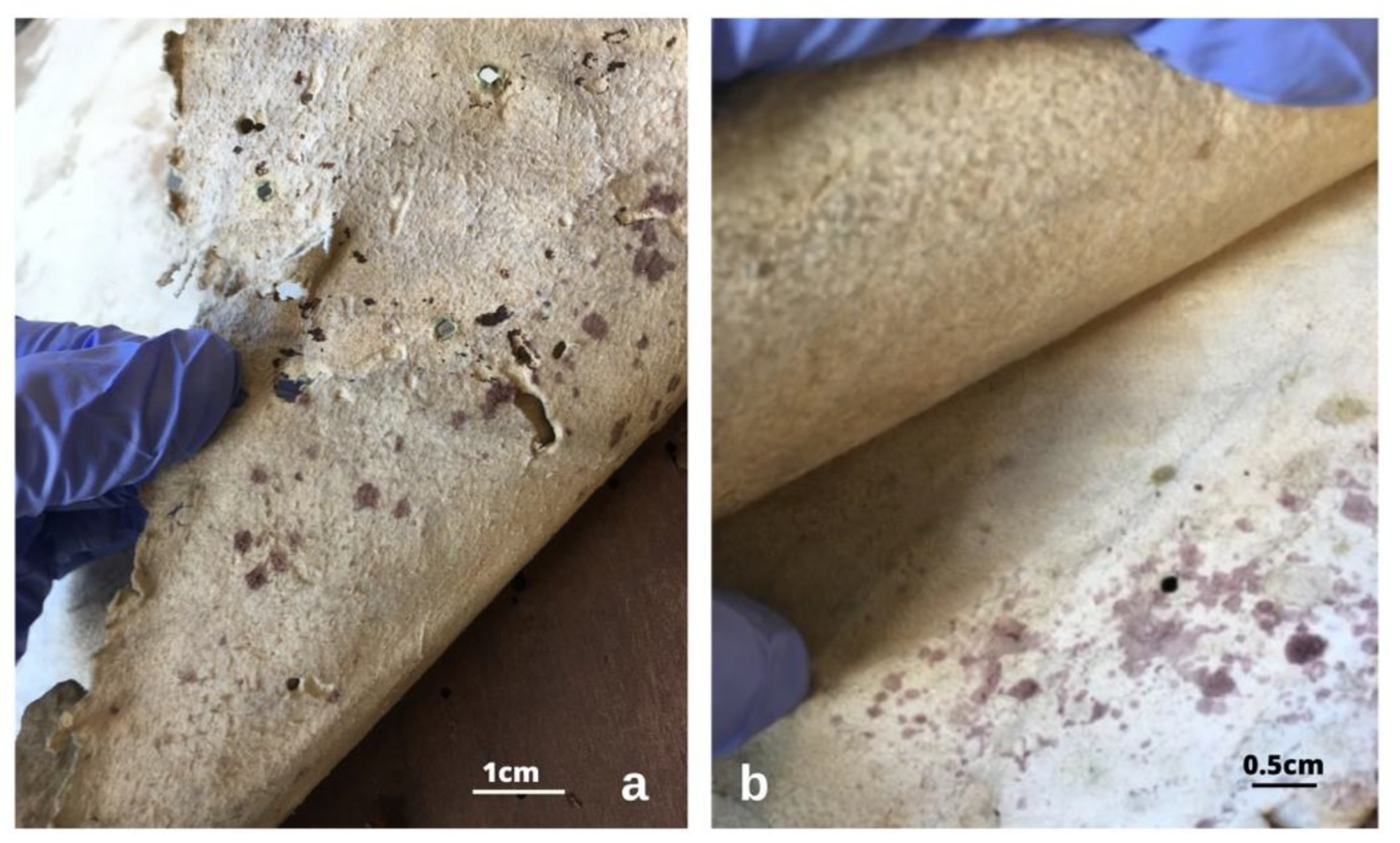
Figure 3.
Purple stains in the tawed white leather coverings, both on the flesh side (a) and in the grain side. The purple spots present different hues, and brownish stains are also visible (b).
Figure 3.
Purple stains in the tawed white leather coverings, both on the flesh side (a) and in the grain side. The purple spots present different hues, and brownish stains are also visible (b).

Figure 4.
Bovine leather in the cover (a), sheep leather in the linings and straps (b). Magnification: circa 50× [22]. Image by Catarina Pinheiro.
Figure 4.
Bovine leather in the cover (a), sheep leather in the linings and straps (b). Magnification: circa 50× [22]. Image by Catarina Pinheiro.

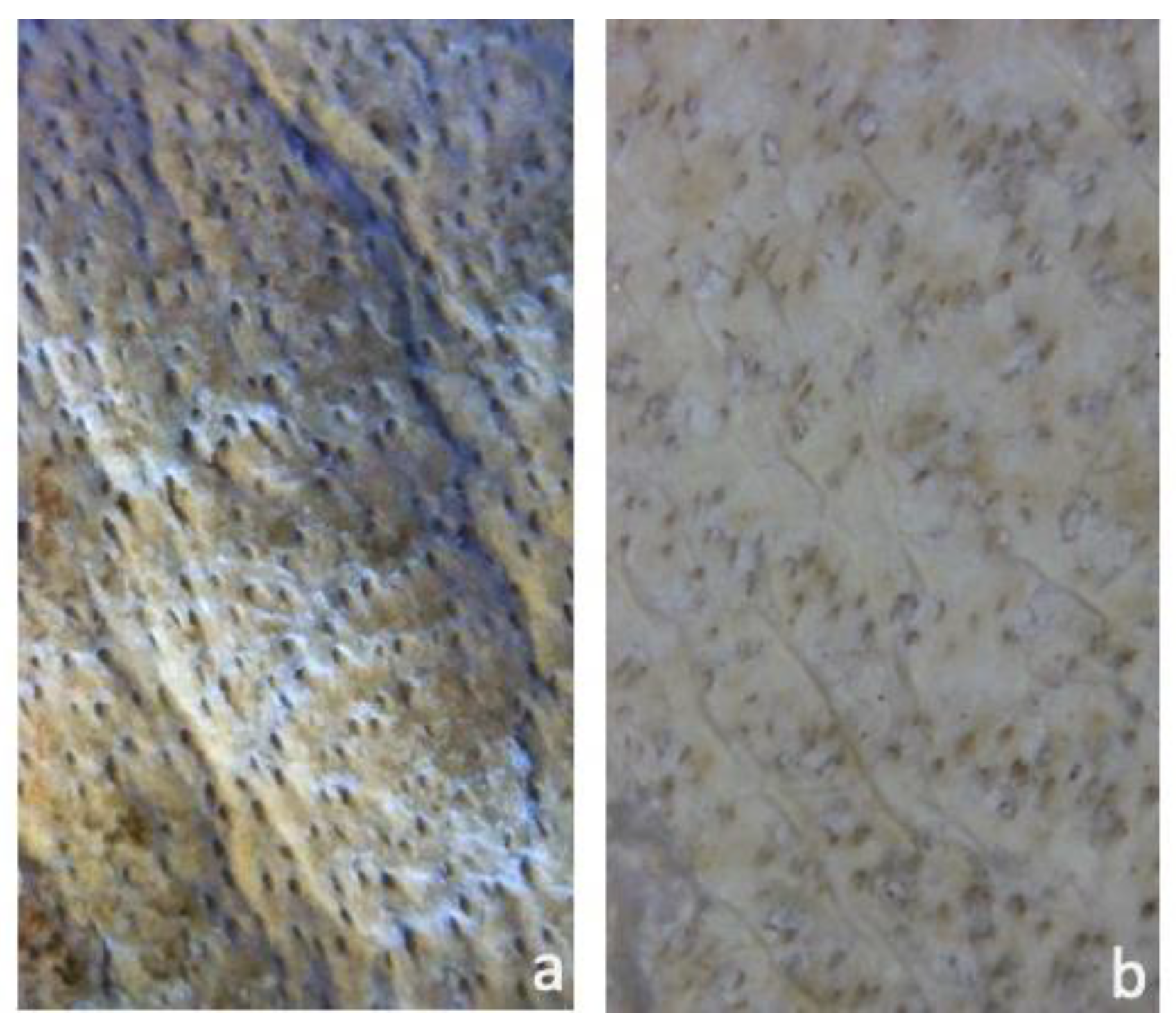
Figure 5.
Magnified fragment of deteriorated parchment at the stereomicroscope. The left section (a) shows less biodeterioration, but one can identify a darkish core underneath the still “undamaged” parchment. The right section (b) has lost nearly all of its top layer, exposing the purple core. Scale bar = 500 µm.
Figure 5.
Magnified fragment of deteriorated parchment at the stereomicroscope. The left section (a) shows less biodeterioration, but one can identify a darkish core underneath the still “undamaged” parchment. The right section (b) has lost nearly all of its top layer, exposing the purple core. Scale bar = 500 µm.

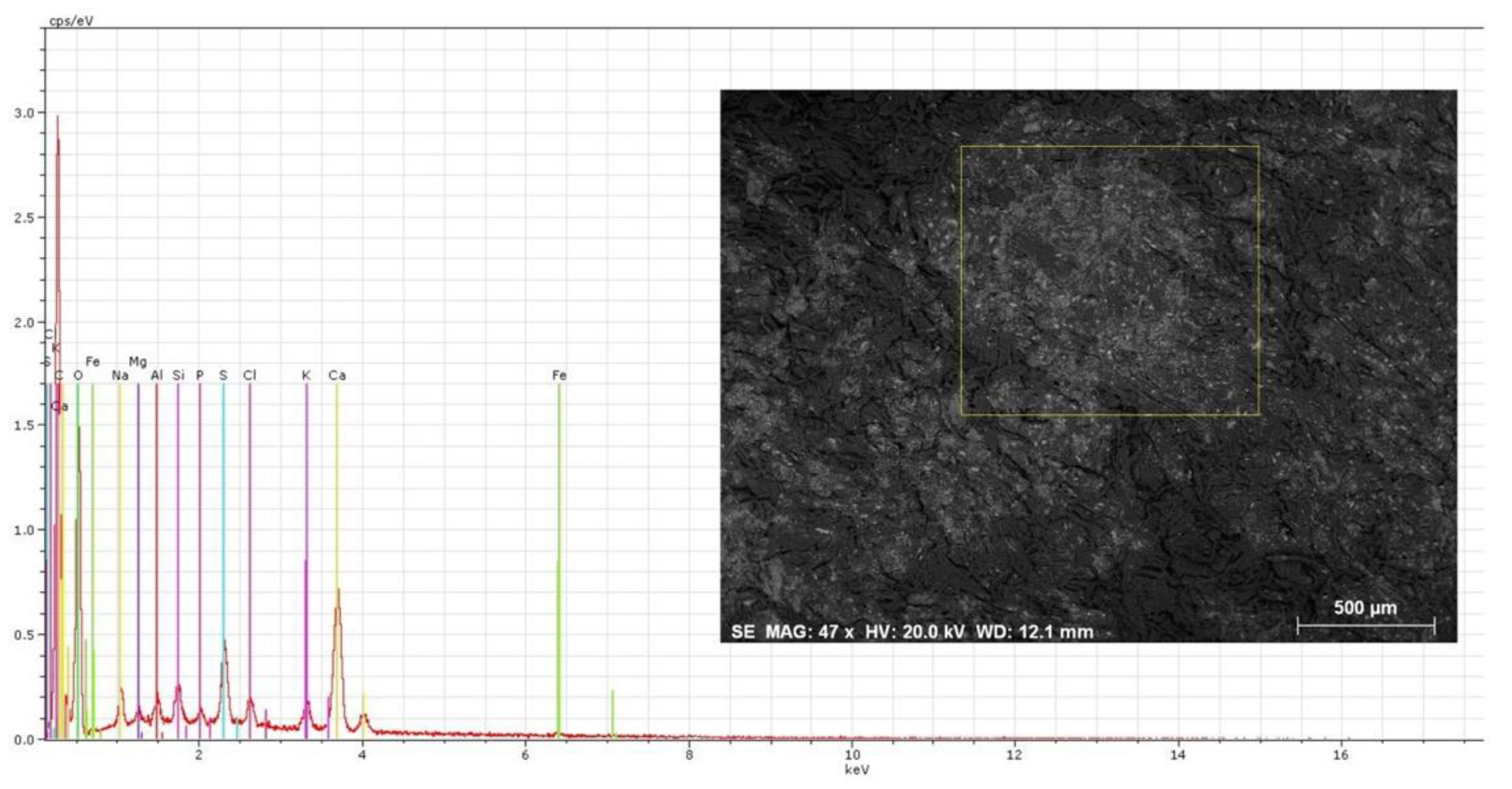
Figure 6.
SEM image of the unstained area. Next to this image is the EDS spectra obtained. Six areas were selected from within the unstained area of the parchment and each was analyzed three times for representativeness. Some alterations were noted on the heights of the peaks for each of the elements being displayed, but overall the results were similar, qualitatively and quantitatively. The spectrum displayed is an example of the obtained results. The operating conditions for EDS analysis were 20 kV accelerating voltage and 10–12 mm working distance.
Figure 6.
SEM image of the unstained area. Next to this image is the EDS spectra obtained. Six areas were selected from within the unstained area of the parchment and each was analyzed three times for representativeness. Some alterations were noted on the heights of the peaks for each of the elements being displayed, but overall the results were similar, qualitatively and quantitatively. The spectrum displayed is an example of the obtained results. The operating conditions for EDS analysis were 20 kV accelerating voltage and 10–12 mm working distance.

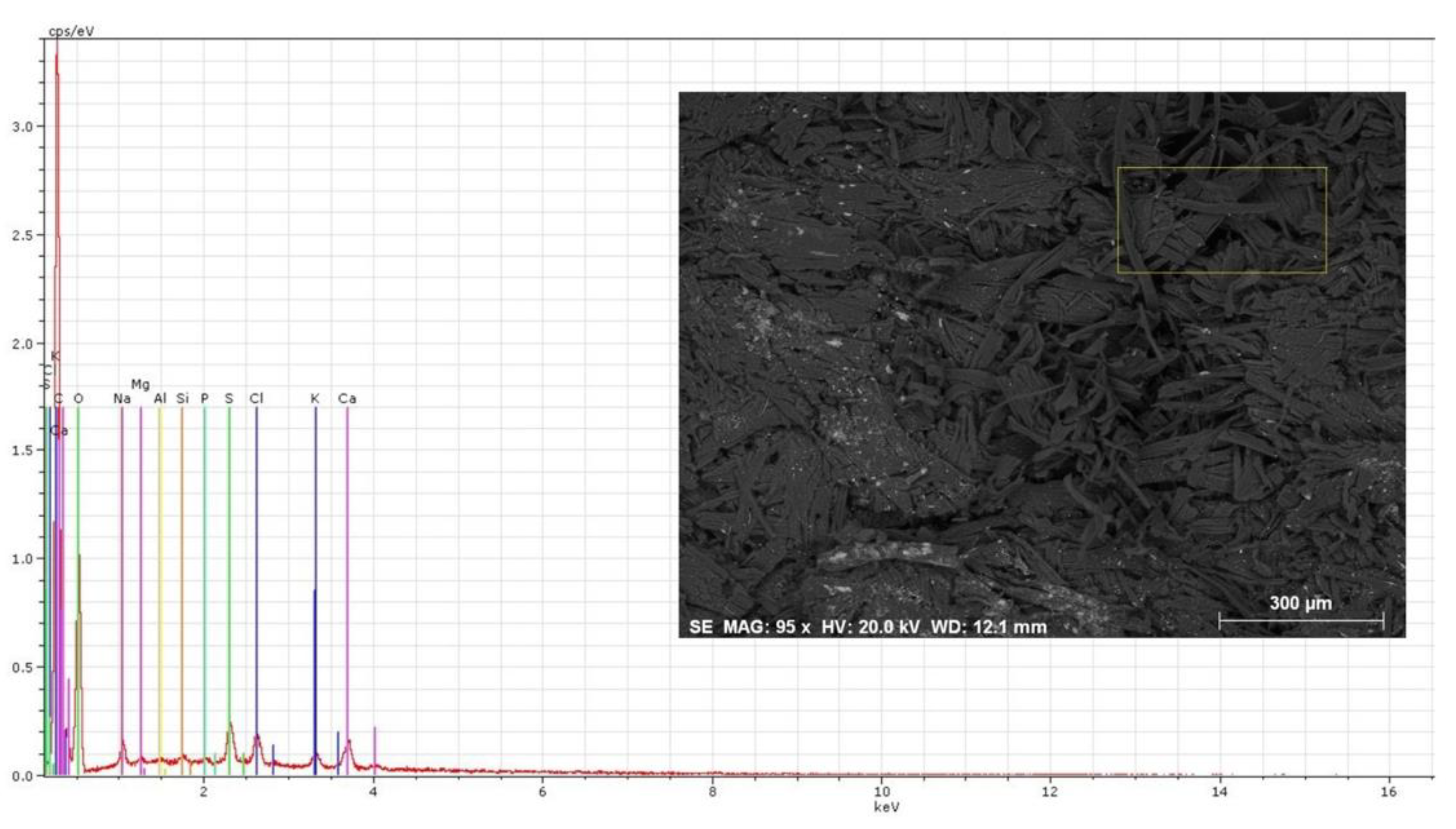
Figure 7.
SEM image of the stained area, visibly more deteriorated than the former one (Figure 6). Representative EDS spectrum obtained. Six areas were selected from within the stained area of the parchment and each was analyzed three times for representativeness. The operating conditions for EDS analysis were 20 kV accelerating voltage and 10–12 mm working distance.
Figure 7.
SEM image of the stained area, visibly more deteriorated than the former one (Figure 6). Representative EDS spectrum obtained. Six areas were selected from within the stained area of the parchment and each was analyzed three times for representativeness. The operating conditions for EDS analysis were 20 kV accelerating voltage and 10–12 mm working distance.

Figure 8.
Electron microscopy images from unstained (a) and purple-stained areas (b–d) from ALC.338 parchment folia. The more cohesive structure is visible in (a) when compared to image (b), where one can detect an intense net of biological structures. (c) the fungal spore is visible and bacterial cells are visible in image (d). Measurements were made under high-vacuum and low-pressure conditions, at an accelerating voltage of 5 kV. The sample was gold coated for better visualization.
Figure 8.
Electron microscopy images from unstained (a) and purple-stained areas (b–d) from ALC.338 parchment folia. The more cohesive structure is visible in (a) when compared to image (b), where one can detect an intense net of biological structures. (c) the fungal spore is visible and bacterial cells are visible in image (d). Measurements were made under high-vacuum and low-pressure conditions, at an accelerating voltage of 5 kV. The sample was gold coated for better visualization.

Figure 9.
Bacterial and Fungal phyla identified. In the Bacterial group (left) there was Acidobacteria, Bacteroidota, Cyanobacteria and Planctomycetota, but with very low relative frequencies (0.23% each). The most represented phyla for the fungi (right) was the Ascomycota, followed by the Basidiomycota.
Figure 9.
Bacterial and Fungal phyla identified. In the Bacterial group (left) there was Acidobacteria, Bacteroidota, Cyanobacteria and Planctomycetota, but with very low relative frequencies (0.23% each). The most represented phyla for the fungi (right) was the Ascomycota, followed by the Basidiomycota.

Figure 10.
Bacterial profile from the parchment sample affected by the purple stains. The genera grouped together display the same frequency (%).
Figure 10.
Bacterial profile from the parchment sample affected by the purple stains. The genera grouped together display the same frequency (%).

Figure 11.
Fungal profile from the parchment sample affected by the purple stains.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Pinheiro, C.; Miller, A.Z.; Vaz, P.; Caldeira, A.T.; Casanova, C. Underneath the Purple Stain. Heritage 2022, 5, 4100-4113. https://doi.org/10.3390/heritage5040212
AMA Style
Pinheiro C, Miller AZ, Vaz P, Caldeira AT, Casanova C. Underneath the Purple Stain. Heritage. 2022; 5(4):4100-4113. https://doi.org/10.3390/heritage5040212
Chicago/Turabian StylePinheiro, Catarina, Ana Zélia Miller, Patrícia Vaz, Ana Teresa Caldeira, and Conceição Casanova. 2022. "Underneath the Purple Stain" Heritage 5, no. 4: 4100-4113. https://doi.org/10.3390/heritage5040212