
Biological Deterioration of an Inca Monument at High Altitude in the Andean Range: A Case Study from Ingapirca’s Temple of the Sun (Ecuador)
 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
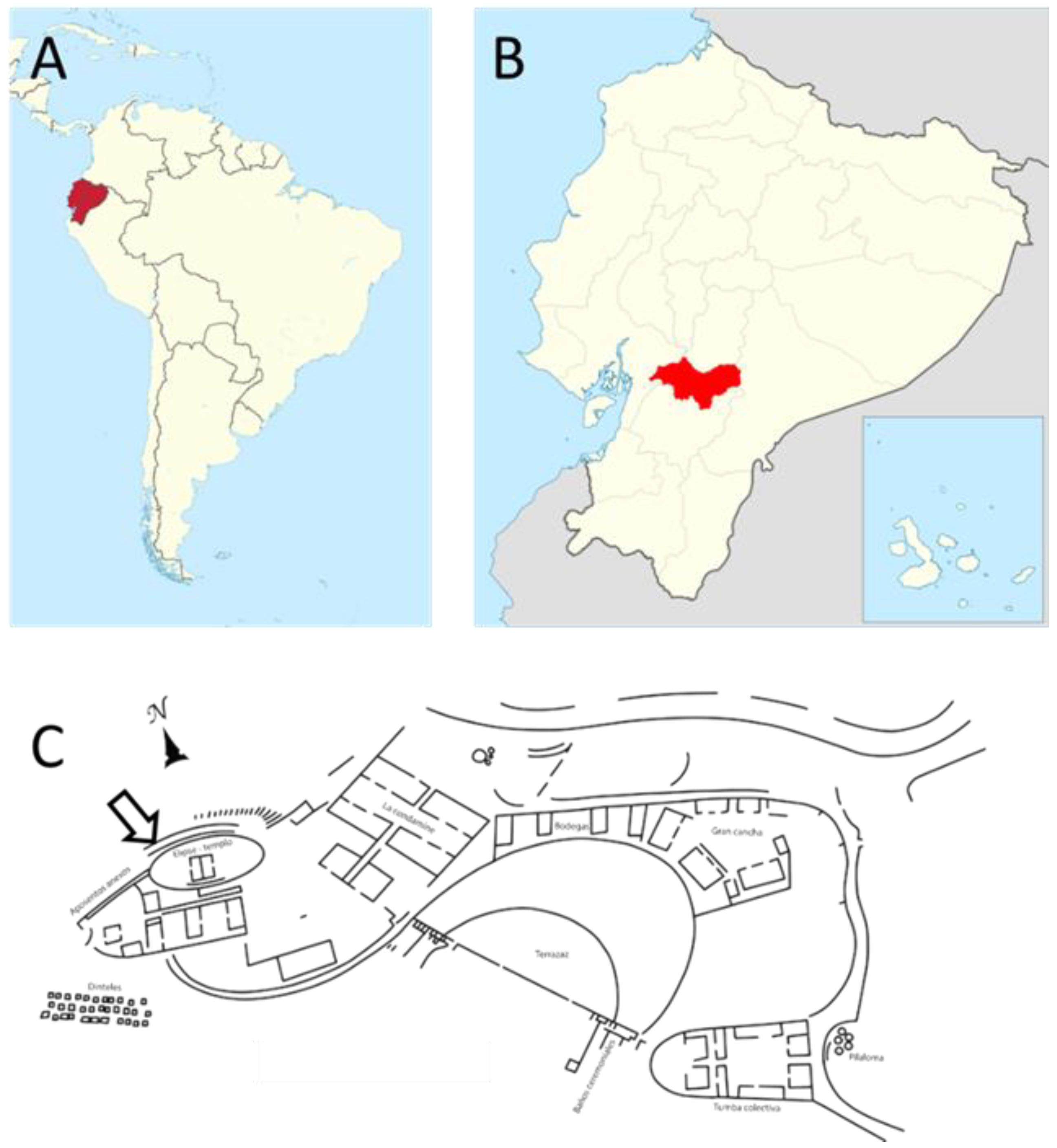
2.1. Field Location and Microclimate
2.2. Rock Sample Collection
2.3. Scanning Electron Microscopy Analysis of Stone Samples
2.4. Sampling and Isolation of Bacteria Colonizing Andesite Blocks’ Surface
2.5. Characterization of Bacterial Isolates
2.6. Identification of Bacterial Isolates
2.7. Screening of the Mineral-Solubilization Abilities of Bacterial Isolates
2.8. In Vitro Simulation of Green Andesite Deterioration Process
2.9. Flame Atomic Absorption Spectroscopy (FAAS) Analysis of Solubilized Minerals
2.10. Statistical Analysis
3. Results
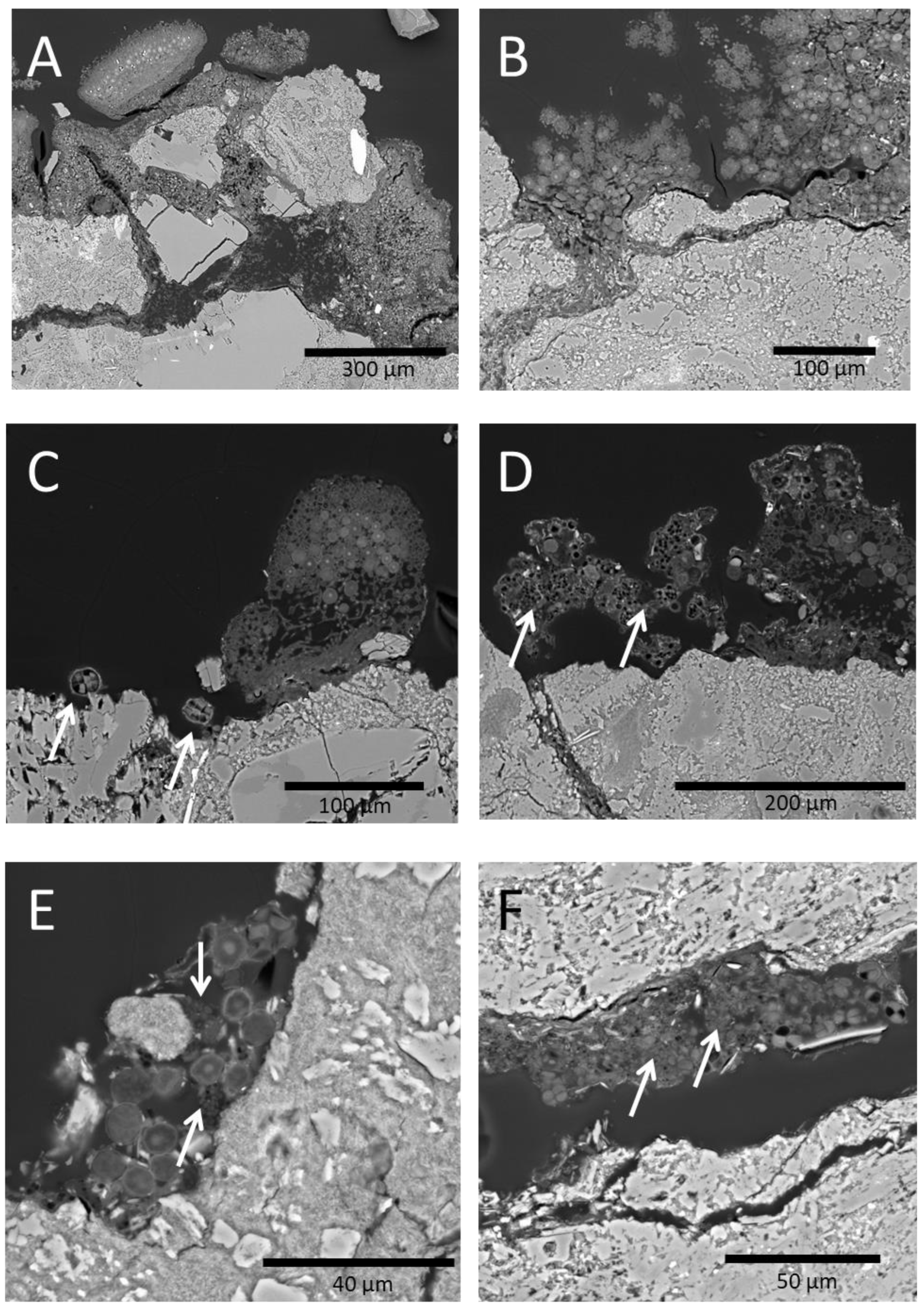
3.1. Biological Characterization of Andesite Blocks’ Surfaces
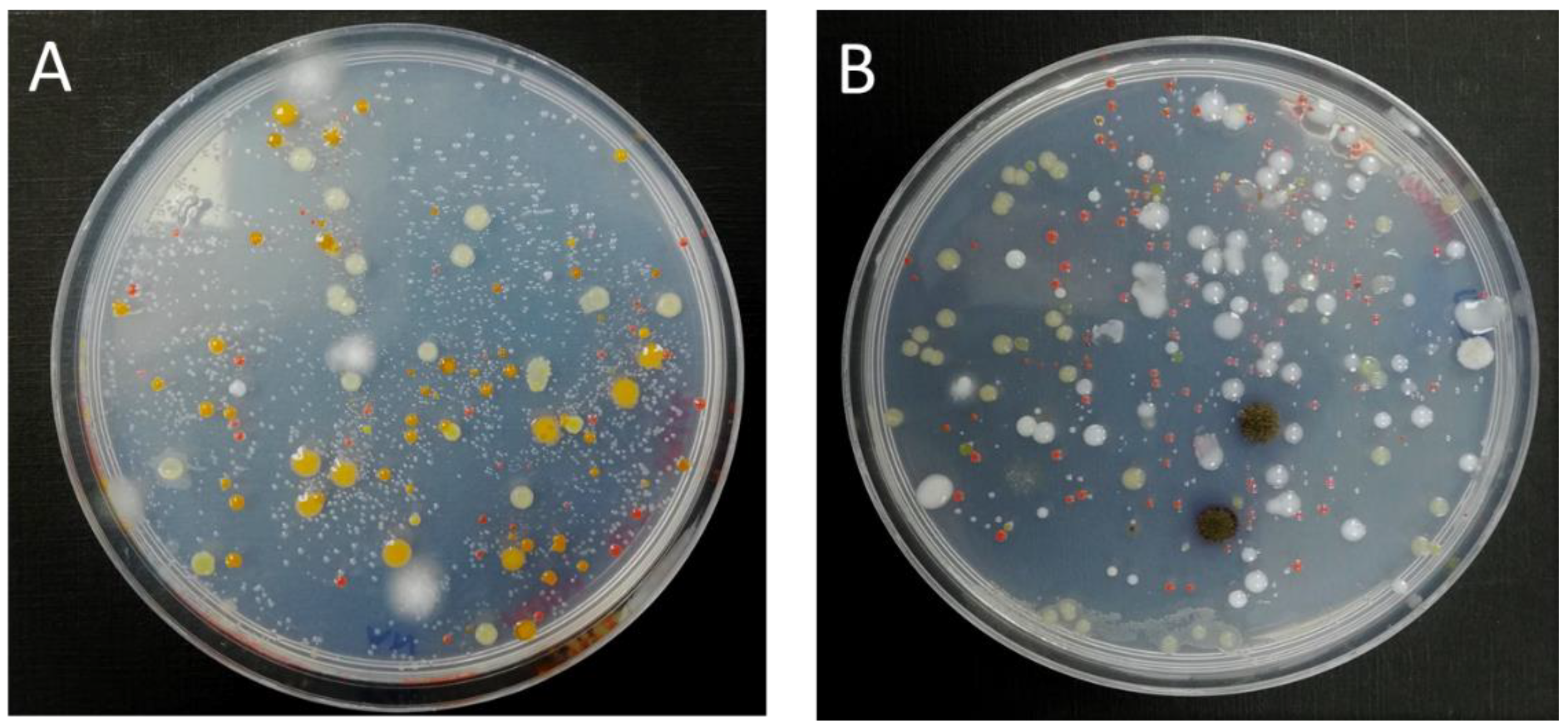
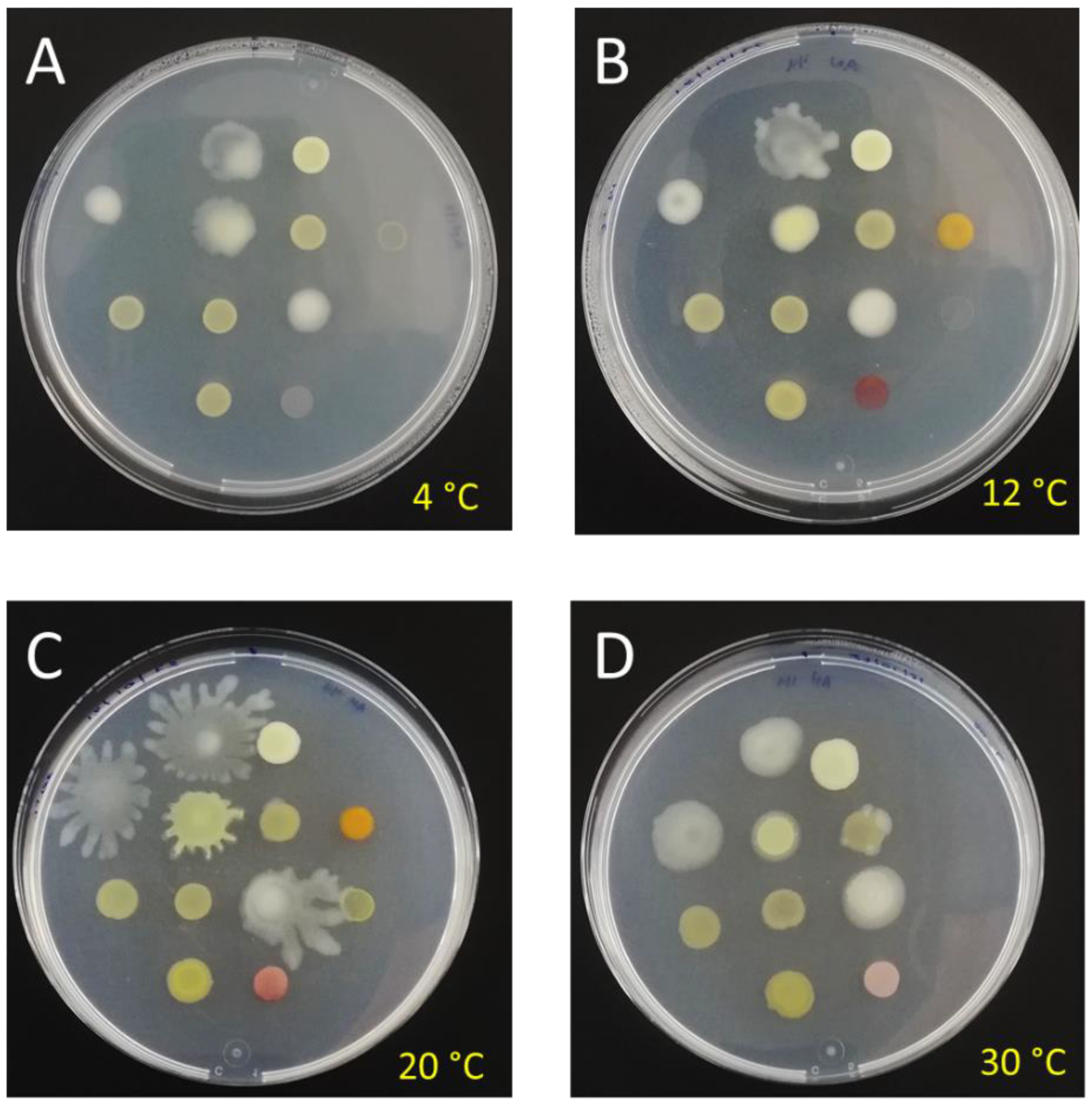
3.2. Morphological and Physiological Characterization of Bacterial Isolates
3.3. Mineral Solubilization Assays
3.4. Simulation of Green Andesite Bioweathering
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, X.; Koestler, R.J.; Warscheid, T.; Katayama, Y.; Gu, J.D. Microbial deterioration and sustainable conservation of stone monuments and buildings. Nat. Sustain. 2020, 3, 991–1004. [Google Scholar] [CrossRef]
- Scheerer, S.; Ortega-Morales, O.; Gaylarde, C. Microbial deterioration of stone monuments—An updated overview. Adv. Appl. Microbiol. 2009, 66, 97–139. [Google Scholar] [CrossRef] [PubMed]
- Schröer, L.; Boon, N.; De Kock, T.; Cnudde, V. The capabilities of bacteria and archaea to alter natural building stones—A review. Int. Biodeterior. Biodegrad. 2021, 165, 105329. [Google Scholar] [CrossRef]
- Sohrabi, M.; Favero-Longo, S.E.; Pérez-Ortega, S.; Ascaso, C.; Haghighat, Z.; Talebian, M.H.; Fadaei, H.; de los Ríos, A. Lichen colonization and associated deterioration processes in Pasargadae, UNESCO world heritage site, Iran. Int. Biodeterior. Biodegrad. 2017, 117, 171–182. [Google Scholar] [CrossRef]
- Tonon, C.; Favero-Longo, S.E.; Matteucci, E.; Piervittori, R.; Croveri, P.; Appolonia, L.; Meirano, V.; Serino, M.; Elia, D. Microenvironmental features drive the distribution of lichens in the House of the Ancient Hunt, Pompeii, Italy. Int. Biodeterior. Biodegrad. 2019, 136, 71–81. [Google Scholar] [CrossRef]
- Nir, I.; Barak, H.; Kramarsky-Winter, E.; Kushmaro, A.; de los Ríos, A. Microscopic and biomolecular complementary approaches to characterize bioweathering processes at petroglyph sites from the Negev Desert, Israel. Environ. Microbiol. 2022, 24, 967–980. [Google Scholar] [CrossRef]
- Ortega-Morales, O.; Montero-Munoz, J.L.; Baptista Neto, J.A.; Beech, I.B.; Sunner, J.; Gaylarde, C. Deterioration and microbial colonization of cultural heritage stone buildings in polluted and unpolluted tropical and subtropical climates: A metaanalysis. Int. Biodeterior. Biodegrad. 2019, 143, 104734. [Google Scholar] [CrossRef]
- Ortega-Morales, B.O.; Narváez-Zapata, J.A.; Schmalenberger, A.; Sosa-López, A.; Tebbe, C.C. Biofilms fouling ancient limestone Mayan monuments in Uxmal, Mexico: A cultivation-independent analysis. Biofilms 2004, 1, 79–90. [Google Scholar] [CrossRef]
- McNamara, C.J.; Perry, T.D.; Bearce, K.A.; Hernandez-Duque, G.; Mitchell, R. Epilithic and endolithic bacterial communities in limestone from a Maya archaeological site. Microb. Ecol. 2006, 51, 51–64. [Google Scholar] [CrossRef]
- Gaylarde, C.; Ogawa, A.; Beech, I.; Kowalski, M.; Baptista Neto, J.A. Analysis of dark crusts on the church of Nossa Senhora do Carmo in Rio de Janeiro, Brazil, using chemical, microscope and metabarcoding microbial identification techniques. Int. Biodeterior. Biodegrad. 2017, 117, 60–67. [Google Scholar] [CrossRef]
- Lemus-Espinoza, D.; Maniscalchi-Badaoui, M.T.; Quereguan, R.; Morales, Y. Biodeterioro del Monumento Religioso Arquitectónico “San Antonio de Padua”, Clarines, Anzoátegui, Venezuela. SABER 2012, 24, 98–102. Available online: https://www.redalyc.org/pdf/4277/427739447010.pdf (accessed on 28 August 2022).
- Gaylarde, C.C.; Gaylarde, P.M.; Neilan, B.A. Endolithic phototrophs in built and natural stone. Curr. Microbiol. 2012, 65, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Morillas, H.; Maguregui, M.; Gallego-Cartagena, E.; Huallparimachi, G.; Marcaida, I.; Salcedo, I.; Silva, L.F.O.; Astete, F. Evaluation of the role of biocolonizations in the conservation state of Machu Picchu (Peru): The Sacred Rock. Sci. Total Environ. 2019, 654, 1379–1388. [Google Scholar] [CrossRef]
- Instituto Nacional del Patrimonio Cultural (INPC) Report. Diagnóstico del Estado de Conservación. Complejo Arqueológico Ingapirca. Available online: https://nube.patrimoniocultural.gob.ec/owncloud/index.php/s/XX6fv76QqxZ6Eht#pdfviewer (accessed on 4 August 2022).
- Romero Bastidas, M.; Díaz Benalcázar, R.; Espinoza, F. Estado de Conservación del Complejo Arqueológico Ingapirca. PAQU Instituto Nacional del Patrimonio Cultural. 2020. Available online: https://paqu.patrimoniocultural.gob.ec/investigacion/diagnostico-estado-conservacion-ingapirca/ (accessed on 4 August 2022).
- Wierzchos, J.; Ascaso, C. Application of back-scattered electron imaging to study the lichen–rock interface. J. Microsc. 1994, 175, 54–59. [Google Scholar] [CrossRef]
- Brenner, D.J.; Krieg, N.R.; Staley, J.T.; Garrity, G.M. The Proteobacteria, Part A Introductory Essays. In Bergey’s Manual of Systematic Bacteriology; Williams & Wilkins: Baltimore, MD, USA; London, UK, 2012; Volume 2. [Google Scholar]
- Whitman, W.B.; Goodfellow, M.; Kämpfer, P.; Busse, H.J.; Trujillo, M.E.; Suzuki, K.I.; Ludwig, W.; Whitman, W.B. The Actinobacteria. In Bergey’s Manual of Systematic Bacteriology; Williams & Wilkins: Baltimore, MD, USA; London, UK, 2012; Volume 5. [Google Scholar]
- Nautiyal, C.S. An efficient microbiological growth medium for screening phosphate solubilizing microorganisms. FEMS Microbiol. Lett. 1999, 170, 265–270. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 11 July 2022).
- Torracchi, J.E.; Universidad Católica de Cuenca, Cuenca, Azuay, Ecuador. Personal communication, 2022.
- Wong, F.K.Y.; Lau, M.C.Y.; Lacap, D.C.; Aitchison, J.C.; Cowan, D.A.; Pointing, S.B. Endolithic microbial colonization of limestone in a high-altitude arid environment. Microb. Ecol. 2010, 59, 689–699. [Google Scholar] [CrossRef]
- Choe, Y.H.; Kim, M.; Woo, J.; Lee, M.J.; Lee, J.I.; Lee, E.J.; Lee, Y.K. Comparing rock-inhabiting microbial communities in different rock types from a high arctic polar desert. FEMS Microbiol. Ecol. 2018, 94, fiy070. [Google Scholar] [CrossRef]
- de Los Ríos, A.; Wierzchos, J.; Ascaso, C. The lithic microbial ecosystems of Antarctica’s McMurdo Dry Valleys. Antarct. Sci. 2014, 26, 459–477. [Google Scholar] [CrossRef]
- Garrido-Benavent, I.; Pérez-Ortega, S.; Durán, J.; Ascaso, C.; Pointing, S.B.; Rodríguez-Cielos, R.; Navarro, F.; de Los Ríos, A. Differential Colonization and Succession of Microbial Communities in Rock and Soil Substrates on a Maritime Antarctic Glacier Forefield. Front. Microbiol. 2020, 11, 126. [Google Scholar] [CrossRef]
- Cavicchioli, R. On the concept of a psychrophile. ISME J. 2016, 10, 793–795. [Google Scholar] [CrossRef]
- Feller, G.; Gerday, C. Psychrophilic enzymes: Hot topics in cold adaptation. Nat. Rev. Microbiol. 2003, 1, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.; Yarzábal, L.A. Bioprospecting cold-adapted plant growth promoting microorganisms from mountain environments. Appl. Microbiol. Biotechnol. 2019, 103, 643–657. [Google Scholar] [CrossRef]
- Yarzábal, L.A. Perspectives for using glacial and periglacial microorganisms for plant growth promotion at low temperatures. Appl. Microbiol. Biotechnol. 2020, 104, 3267–3278. [Google Scholar] [CrossRef] [PubMed]
- Lessie, T.G.; Phibbs, P.V. Alternative pathways of carbohydrate utilization in pseudomonads. Annu. Rev. Microbiol. 1984, 38, 359–388. [Google Scholar] [CrossRef]
- Goldstein, A.H. Future trends in research on microbial phosphate solubilization: 100 years of insolubility. In First International Meeting on Microbial Phosphate Solubilization; Velazquez, E., Rodrıguez-Barrueco, C., Eds.; Springer: Dordrhecht, The Netherlands, 2007; pp. 91–96. [Google Scholar] [CrossRef]
- Rodríguez, H.; Fraga, R. Phosphate solubilizing bacteria and their role in plant growth promotion. Biotechnol. Adv. 1999, 17, 319–339. [Google Scholar] [CrossRef]
- Warscheid, T.; Braams, J. Biodeterioration of stone: A review. Int. Biodeterior. Biodegrad. 2000, 46, 343–368. [Google Scholar] [CrossRef]
- da Silva, A.V.; de Oliveira, A.J.; Tanabe, I.S.B.; Silva, J.V.; da Silva Barros, T.W.; da Silva, M.K.; França, P.H.B.; Leite, J.; Putzke, J.; Montone, R.; et al. Antarctic lichens as a source of phosphate-solubilizing bacteria. Extremophiles 2021, 25, 181–191. [Google Scholar] [CrossRef]
- Becerra-Castro, C.; Kidd, P.; Kuffner, M.; Prieto-Fernández, Á.; Hann, S.; Monterroso, C.; Sessitsch, A.; Wenzel, W.; Puschenreiter, M. Bacterially induced weathering of ultramafic rock and its implications for phytoextraction. Appl. Env. Microbiol. 2013, 79, 5094–5103. [Google Scholar] [CrossRef]
- De Maayer, P.; Anderson, D.; Cary, C.; Cowan, D.A. Some like it cold: Understanding the survival strategies of psychrophiles. EMBO Rep. 2014, 15, 508–517. [Google Scholar] [CrossRef]
- de los Ríos, A.; Wierzchos, J.; Sancho, L.G.; Ascaso, C. Exploring the physiological state of continental Antarctic endolithic microorganisms by microscopy. FEMS Microbiol. Ecol. 2004, 50, 143–152. [Google Scholar] [CrossRef] [Green Version]
- Nichols, C.A.M.; Guezennec, J.; Bowman, J.P. Bacterial exopolysaccharides from extreme marine environments with special consideration of the Southern Ocean, sea ice, and deep-sea hydrothermal vents: A review. Mar. Biotechnol. 2005, 7, 253–271. [Google Scholar] [CrossRef] [PubMed]
- Männik, J.; Driessen, R.; Galajda, P.; Keymer, J.E.; Dekker, C. Bacterial growth and motility in sub-micron constrictions. Proc. Natl. Acad. Sci. USA 2009, 106, 14861–14866. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
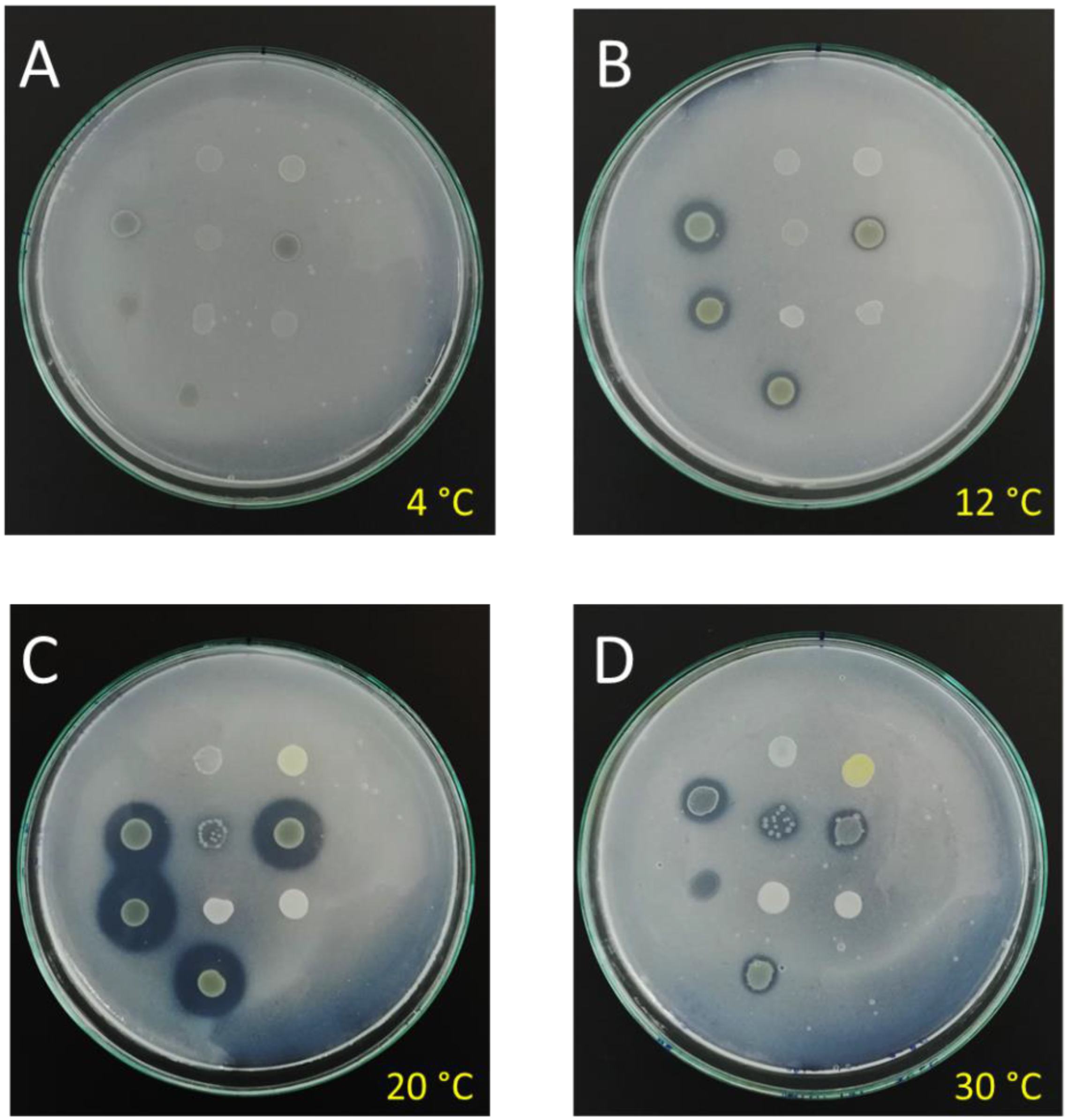{kind=link}
| Origin 1 | Colony Description 2 | Growth Temp. Range | NBRIP 3 | Identification | |
|---|---|---|---|---|---|
| IP001.1 | NA | WsRS | 4–30 | ++ | Pseudarthrobacter sp. |
| IP001.4 | NA | WsRS | 4–30 | ++ | Pseudomonas sp. |
| IP001.5 | NA | YsRS | 4–30 | - | Paenarthrobacter sp. |
| IP001.7 | NA | CsRS | 4–30 | +++ | Pseudomonas sp. |
| IP001.8.2 | NA | YsRS | 4–30 | ++ | Frigoribacterium sp. |
| IP001.11 | PDA | CsRS | 4–30 | +++ | Pseudomonas sp. |
| IP001.21 | Cz | WoRM | 4–30 | +++ | Curtobacterium sp. |
| IP001.23 | Cz | WoRS | 4–30 | +++ | Pseudomonas sp. |
| IP001.25 | NA | WsRM | 4–30 | +++ | Arthrobacter sp. |
| IP001.28 | NA | YsRM | 4–30 | - | Curtobacterium sp. |
| IP002.4.1 | NA | TsRS | 4–30 | +++ | Pseudomonas sp. |
| IP002.6.1 | NA | TsRS | 4–30 | +++ | Pseudomonas sp. |
| IP002.9 | PDA | CmRM | 4–30 | - | Pantoea sp. |
| IP002.20 | NA | YsIM | 4–30 | - | Curtobacterium sp. |
| IP002.21 | Cz | CsRS | 4–30 | +++ | Pseudomonas sp. |
| IP003.3 | NA | OsRS | 4–20 | - | Sphingomonas sp. |
| IP003.5 | NA | CsRS | 4–30 | +++ | Pseudomonas sp. |
| IP003.6 | NA | WsRS | 4–30 | - | Pseudomonas sp. |
| IP003.11 | PDA | CsRS | 4–30 | - | Pseudomonas sp. |
| IP003.15 | PDA | YmRS | 4–30 | + | Pseudomonas sp. |
| IP003.18 | NA | OsRS | 4–20 | - | Sphingomonas sp. |
| IP003.19 | PDA | YsRS | 4–30 | - | Pseudomonas sp. |
| IP003.21 | NA | CsRS | 4–30 | - | Plantibacter sp. |
| IP004.4 | NA | CsRS | 4–30 | ++ | Pseudomonas sp. |
| IP004.5 | NA | WmIL | 4–30 | - | Pseudarthrobacter sp. |
| IP004.8 | NA | WsRS | 4–30 | +++ | Pseudarthrobacter sp. |
| IP004.22 | PDA | YsRS | 12–20 | - | Frondihabitans sp. |
| IP004.24 | NA | CsRS | 4–30 | - | Microbacterium sp. |
| IP004.27 | NA | YsRM | 4–30 | - | Curtobacterium sp. |
| IP004.29 | NA | PsRS | 4–30 | - | Pseudarthrobacter sp. |
| Treatment | Iron (ppm) | Calcium (ppm) | Sodium (ppm) | Magnesium (ppm) |
|---|---|---|---|---|
| Control | 0.000 | 35.384 ± 1.633 | 6.424 ± 0.062 | 22.415 ± 0.032 |
| Isolate IP001.1 | 0.000 | 36.020 ± 2.098 | 5.851 ± 0.068 | 22.509 ± 0.169 |
| Isolate IP001.4 | 76.025 ± 1.532 ** | 34.687 ± 1.690 | 6.513 ± 0.341 | 22.411 ± 0.131 |
| Isolate IP001.11 | 79.756 ± 1.856 ** | 34.672 ± 1.590 | 6.543 ± 0.357 | 22.512 ± 0.069 |
| Isolate IP001.8.2 | 69.205 ± 9.595 ** | 34.990 ± 1.211 | 6.632 ± 0.454 | 22.415 ± 0.094 |
| Isolate IP001.28 | 78.013 ± 2.046 ** | 34.869 ± 0.584 | 6.404 ± 0.286 | 22.460 ± 0.151 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yarzábal, L.A.; Buela, L.; de los Ríos, A.; Peláez, D.; Romero, M.; Espinoza, F.; Torres, A.S.; Medina, G.M.; Landi, J.G.; Tapia, M.V. Biological Deterioration of an Inca Monument at High Altitude in the Andean Range: A Case Study from Ingapirca’s Temple of the Sun (Ecuador). Heritage 2022, 5, 2504-2518. https://doi.org/10.3390/heritage5030130
Yarzábal LA, Buela L, de los Ríos A, Peláez D, Romero M, Espinoza F, Torres AS, Medina GM, Landi JG, Tapia MV. Biological Deterioration of an Inca Monument at High Altitude in the Andean Range: A Case Study from Ingapirca’s Temple of the Sun (Ecuador). Heritage. 2022; 5(3):2504-2518. https://doi.org/10.3390/heritage5030130
Chicago/Turabian StyleYarzábal, Luis Andrés, Lenys Buela, Asunción de los Ríos, Diana Peláez, Martha Romero, Fernando Espinoza, Alisson Samantha Torres, Gina Maritza Medina, Jaqueline Gabriela Landi, and Marco Vinicio Tapia. 2022. "Biological Deterioration of an Inca Monument at High Altitude in the Andean Range: A Case Study from Ingapirca’s Temple of the Sun (Ecuador)" Heritage 5, no. 3: 2504-2518. https://doi.org/10.3390/heritage5030130