
Exploring the Relationship between CLPTM1L-MS2 Variants and Susceptibility to Bladder Cancer

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
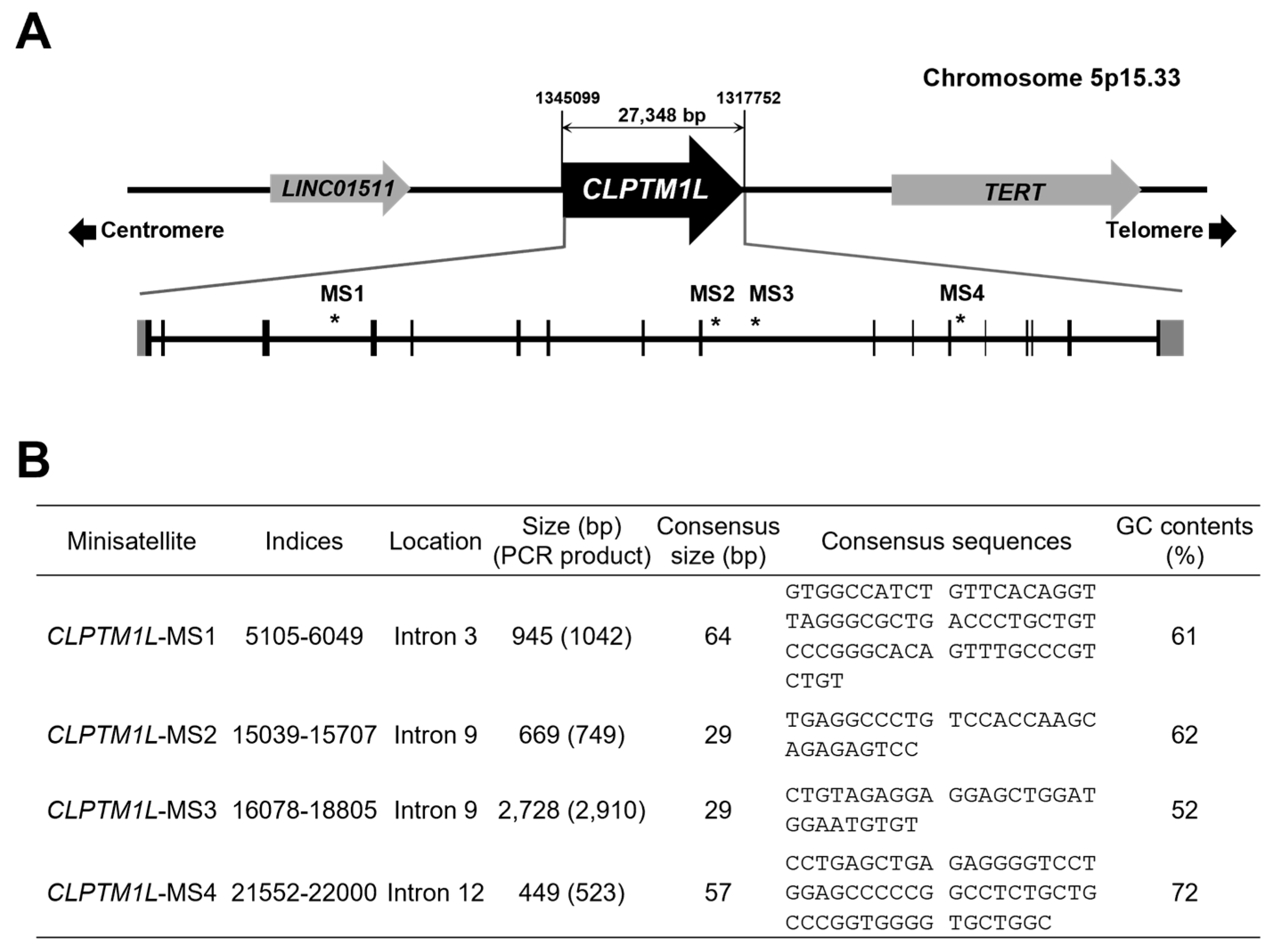
2.1. Structural Analysis and Primer Construction for VNTR Regions of CLPTM1L
2.2. Population for Case–Control Study
2.3. Polymerase Chain Reaction (PCR) Amplification of the VNTR Regions
2.4. Analysis of VNTR Polymorphism through Electrophoresis
2.5. Construction of CLPTM1L Luciferase Reporter Vector Containing CLPTM1L-MS2
2.6. Cell Culture and Luciferase Assay
2.7. Statistical Analysis
3. Results
3.1. Selection of CLPTM1L VNTR Region
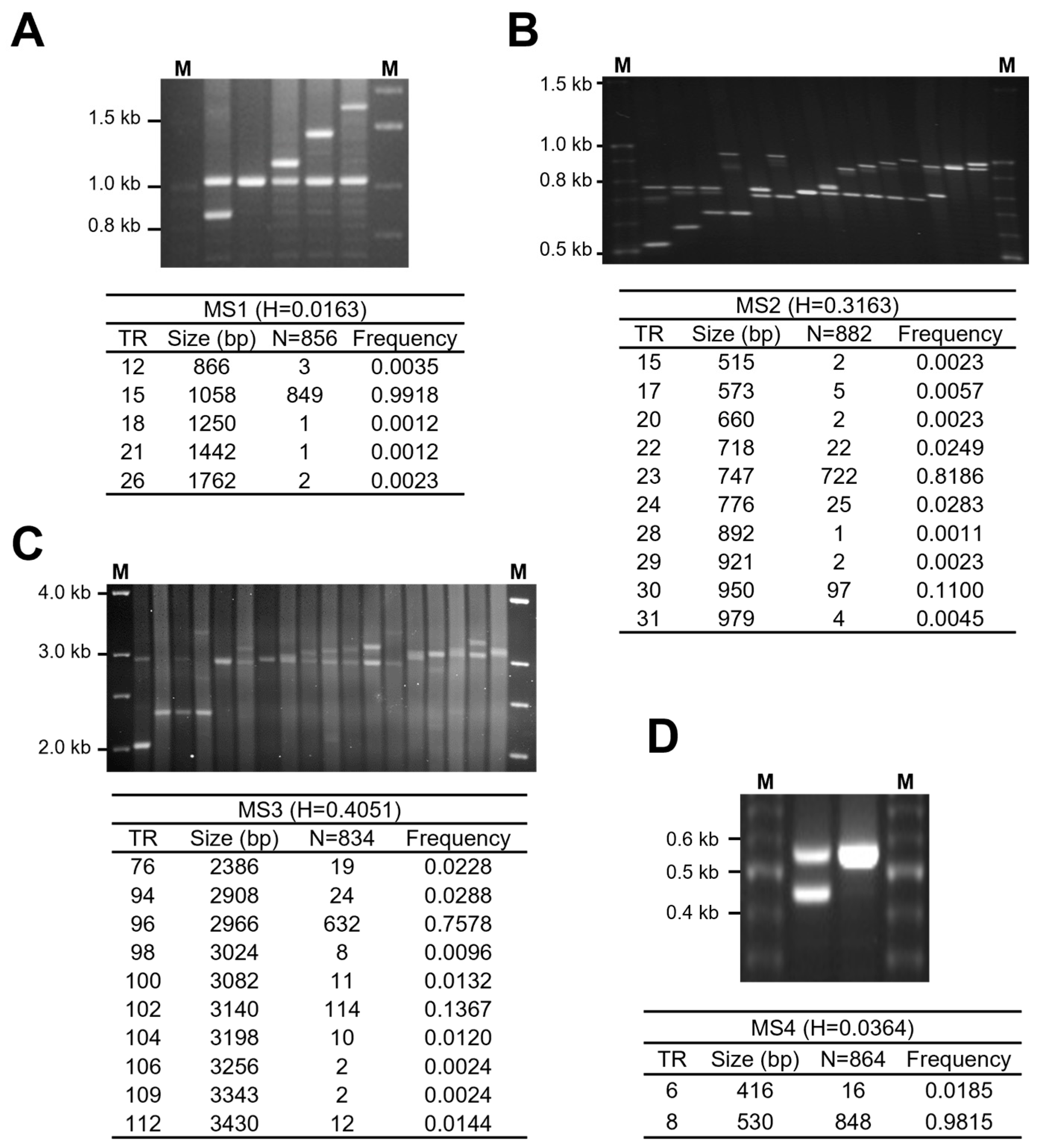
3.2. Polymorphic Analysis of CLPTM1L VNTR Regions
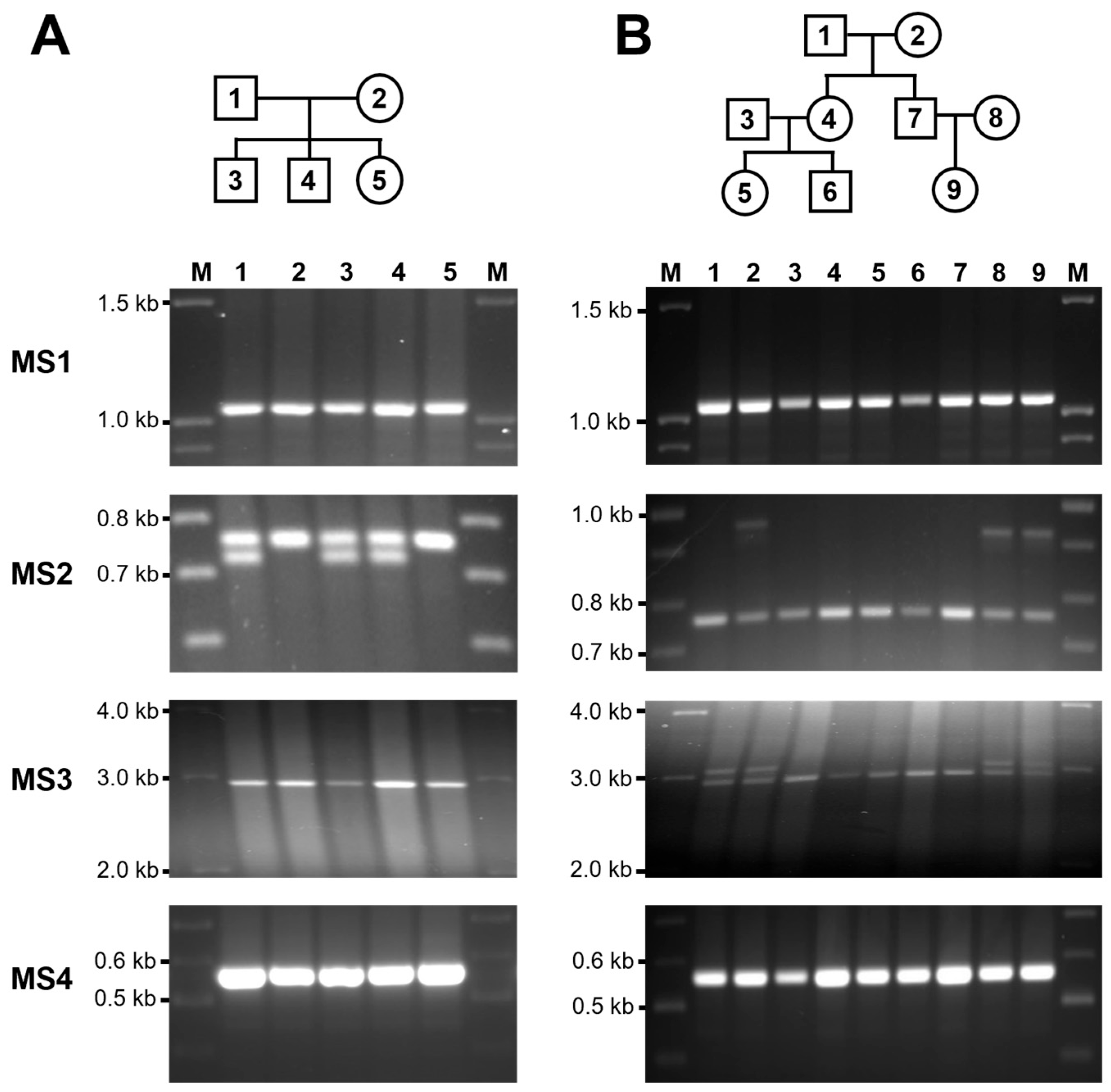
3.3. Mendelian Inheritance of CLPTM1L VNTR
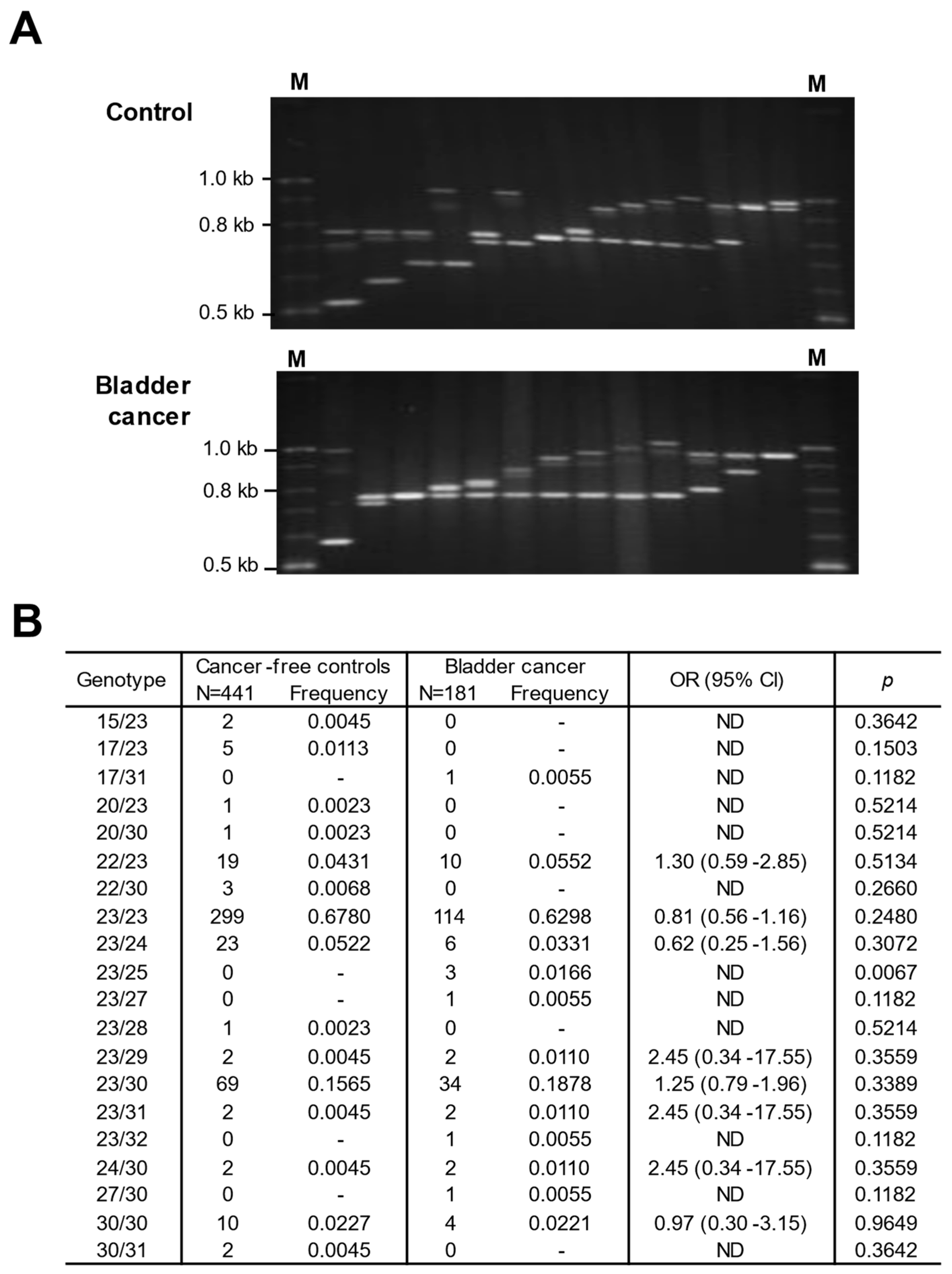
3.4. Association of CLPTM1L VNTR with Allelic Variation and Bladder Cancer
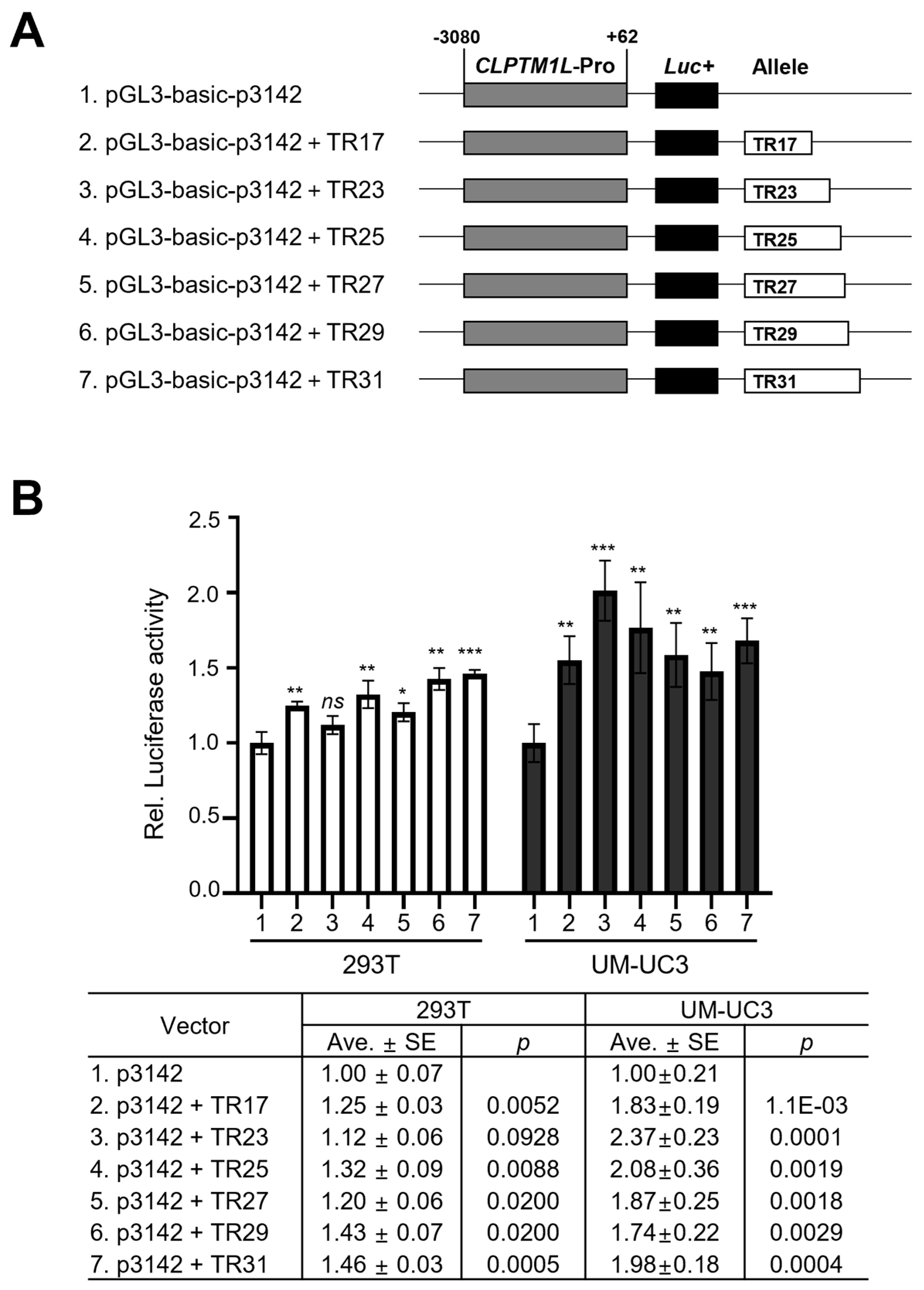
3.5. The Potential Effect of CLPTM1L-MS2 on the Gene Expression
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Al-Zalabani, A.H.; Stewart, K.F.; Wesselius, A.; Schols, A.M.; Zeegers, M.P. Modifiable risk factors for the prevention of bladder cancer: A systematic review of meta-analyses. Eur. J. Epidemiol. 2016, 31, 811–851. [Google Scholar] [CrossRef] [PubMed]
- Knowles, M.A.; Hurst, C.D. Molecular biology of bladder cancer: New insights into pathogenesis and clinical diversity. Nat. Rev. Cancer 2015, 15, 25–41. [Google Scholar] [CrossRef] [PubMed]
- Dudek, A.M.; Grotenhuis, A.J.; Vermeulen, S.H.; Kiemeney, L.A.; Verhaegh, G.W. Urinary bladder cancer susceptibility markers. What do we know about functional mechanisms? Int. J. Mol. Sci. 2013, 14, 12346–12366. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, K.; Okamoto, A.; Isonishi, S.; Ochiai, K.; Ohtake, Y. A novel gene, CRR9, which was up-regulated in CDDP-resistant ovarian tumor cell line, was associated with apoptosis. Biochem. Biophys. Res. Commun. 2001, 280, 1148–1154. [Google Scholar] [CrossRef] [PubMed]
- James, M.A.; Wen, W.D.; Wang, Y.A.; Byers, L.A.; Heymach, J.V.; Coombes, K.R.; Girard, L.; Minna, J.; You, M. Functional Characterization of CLPTM1L as a Lung Cancer Risk Candidate Gene in the 5p15.33 Locus. PLoS ONE 2012, 7, 9. [Google Scholar] [CrossRef] [PubMed]
- Ni, Z.H.; Tao, K.; Chen, G.; Chen, Q.G.; Tang, J.M.; Luo, X.M.; Yin, P.H.; Tang, J.H.; Wang, X.B.A. CLPTM1L Is Overexpressed in Lung Cancer and Associated with Apoptosis. PLoS ONE 2012, 7, 8. [Google Scholar] [CrossRef]
- Zhang, M.; Wu, X.; Lu, W.; Ge, Y.K.; Wang, X.; Cai, Z.M.; Wu, S. Rs401681 polymorphism in TERT-CLPTM1L was associated with bladder cancer risk: A meta-analysis. Iran J. Basic. Med. Sci. 2015, 18, 1130–1136. [Google Scholar]
- Haiman, C.A.; Chen, G.K.; Vachon, C.M.; Canzian, F.; Dunning, A.; Millikan, R.C.; Wang, X.S.; Ademuyiwa, F.; Ahmed, S.; Ambrosone, C.B.; et al. A common variant at the TERT-CLPTM1L locus is associated with estrogen receptor-negative breast cancer. Nat. Genet. 2011, 43, 1210–1214. [Google Scholar] [CrossRef]
- Rafnar, T.; Sulem, P.; Stacey, S.N.; Geller, F.; Gudmundsson, J.; Sigurdsson, A.; Jakobsdottir, M.; Helgadottir, H.; Thorlacius, S.; Aben, K.K.H.; et al. Sequence variants at the TERT-CLPTM1L locus associate with many cancer types. Nat. Genet. 2009, 41, 221–227. [Google Scholar] [CrossRef]
- Petersen, G.M.; Amundadottir, L.; Fuchs, C.S.; Kraft, P.; Stolzenberg-Solomon, R.Z.; Jacobs, K.B.; Arslan, A.A.; Bueno-de-Mesquita, H.B.; Gallinger, S.; Gross, M.; et al. A genome-wide association study identifies pancreatic cancer susceptibility loci on chromosomes 13q22.1, 1q32.1 and 5p15.33. Nat. Genet. 2010, 42, 224–228. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Li, Y.; Yin, M.; Sun, J.; Liu, L.; Qin, Q.; Li, X.; Long, L.; Nie, S.; Wei, S. TERT-CLPTM1L polymorphism rs401681 contributes to cancers risk: Evidence from a meta-analysis based on 29 publications. PLoS ONE 2012, 7, e50650. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Wang, Y.; Dong, Y.; Chang, J.; Wu, Y.; Chang, S.; Che, G. Cumulative Evidence for Relationships Between Multiple Variants in the TERT and CLPTM1L Region and Risk of Cancer and Non-Cancer Disease. Front. Oncol. 2022, 12, 946039. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.-L.; Jung, S.-I.; Do, E.-J.; Lee, S.-R.; Lee, S.-Y.; Chu, I.-S.; Kim, W.-J.; Jung, J.; Kim, C.S.; Cheon, S.-H.; et al. Short rare hTERT-VNTR2-2nd alleles are associated with prostate cancer susceptibility and influence gene expression. BMC Cancer 2010, 10, 393. [Google Scholar] [CrossRef] [PubMed]
- Hofer, P.; Zerelles, J.; Baierl, A.; Madersbacher, S.; Schatzl, G.; Maj-Hes, A.; Sutterlüty-Fall, H.; Gsur, A. MNS16A tandem repeat minisatellite of human telomerase gene and prostate cancer susceptibility. Mutagenesis 2013, 28, 301–306. [Google Scholar] [CrossRef] [PubMed]
- Kwon, J.A.; Jeong, M.S.; Yoon, S.L.; Mun, J.Y.; Kim, M.H.; Yang, G.E.; Park, S.H.; Chung, J.W.; Choi, Y.H.; Cha, H.J.; et al. The hTERT-VNTR2-2(nd) alleles are involved in genomic stability in gastrointestinal cancer. Genes Genom. 2019, 41, 1517–1525. [Google Scholar] [CrossRef]
- Diler, S.B.; Polat, F.; Bingöl, G. The MNS16A VNTR polymorphism of the TERT gene in bladder cancer. Turk. J. Urol. 2019, 46, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Benson, G. Tandem repeats finder: A program to analyze DNA sequences. Nucleic Acids Res. 1999, 27, 573–580. [Google Scholar] [CrossRef]
- Seol, S.-Y.; Lee, S.-Y.; Kim, Y.-D.; Do, E.-J.; Kwon, J.-A.; Kim, S.I.; Chu, I.-S.; Leem, S.-H. Minisatellite polymorphisms of the SLC6A19: Susceptibility in hypertension. Biochem. Biophys. Res. Commun. 2008, 374, 714–719. [Google Scholar] [CrossRef]
- Yoon, Y.-H.; Seol, S.-Y.; Heo, J.; Chung, C.-N.; Park, I.-H.; Leem, S.-H. Analysis of VNTRs in the Solute Carrier Family 6, Member 18 (SLC6A18) and Lack of Association with Hypertension. DNA Cell Biol. 2008, 27, 559–567. [Google Scholar] [CrossRef]
- Yoon, S.-L.; Kim, D.C.; Cho, S.H.; Lee, S.-Y.; Chu, I.-S.; Heo, J.; Leem, S.-H. Susceptibility for breast cancer in young patients with short rare minisatellite alleles of BORIS. BMB Rep. 2010, 43, 698–703. [Google Scholar] [CrossRef] [PubMed]
- Nei, M.; Roychoudhury, A.K.J.G. Sampling variances of heterozygosity and genetic distance. Genetics 1974, 76, 379–390. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, Y.; Koyama, K.; Matsushima, M. VNTR (variable number of tandem repeat) sequences as transcriptional, translational, or functional regulators. J. Human Genet. 1998, 43, 149–152. [Google Scholar] [CrossRef]
- Greenwood, T.A.; Kelsoe, J.R. Promoter and intronic variants affect the transcriptional regulation of the human dopamine transporter gene. Genomics 2003, 82, 511–520. [Google Scholar] [CrossRef] [PubMed]
- De Roeck, A.; Duchateau, L.; Van Dongen, J.; Cacace, R.; Bjerke, M.; Van den Bossche, T.; Cras, P.; Vandenberghe, R.; De Deyn, P.P.; Engelborghs, S.; et al. An intronic VNTR affects splicing of ABCA7 and increases risk of Alzheimer’s disease. Acta Neuropathol. 2018, 135, 827–837. [Google Scholar] [CrossRef] [PubMed]
- Brookes, K.J. The VNTR in complex disorders: The forgotten polymorphisms? A functional way forward? Genomics 2013, 101, 273–281. [Google Scholar] [CrossRef] [PubMed]
- Jeong, Y.H.; Kim, M.C.; Ahn, E.K.; Seol, S.Y.; Do, E.J.; Choi, H.J.; Chu, I.S.; Kim, W.J.; Kim, W.J.; Sunwoo, Y.; et al. Rare Exonic Minisatellite Alleles in MUC2 Influence Susceptibility to Gastric Carcinoma. PLoS ONE 2007, 2, 10. [Google Scholar] [CrossRef]
- Kwon, J.A.; Lee, S.Y.; Ahn, E.K.; Seol, S.Y.; Kim, M.C.; Kim, S.J.; Kim, S.I.; Chu, I.S.; Leem, S.H. Short Rare MUC6 Minisatellites-5 Alleles Influence Susceptibility to Gastric Carcinoma by Regulating Gene Expression. Hum. Mutat. 2010, 31, 942–949. [Google Scholar] [CrossRef]
- Karami, S.; Han, Y.; Pande, M.; Cheng, I.; Rudd, J.; Pierce, B.L.; Nutter, E.L.; Schumacher, F.R.; Kote-Jarai, Z.; Lindstrom, S.; et al. Telomere structure and maintenance gene variants and risk of five cancer types. Int. J. Cancer 2016, 139, 2655–2670. [Google Scholar] [CrossRef]
- Jia, J.P.; Bosley, A.D.; Thompson, A.; Hoskins, J.W.; Cheuk, A.; Collins, I.; Parikh, H.; Xiao, Z.; Ylaya, K.; Dzyadyk, M.; et al. CLPTM1L Promotes Growth and Enhances Aneuploidy in Pancreatic Cancer Cells. Cancer Res. 2014, 74, 2785–2795. [Google Scholar] [CrossRef]
- Puskas, L.G.; Man, I.; Szebeni, G.; Tiszlavicz, L.; Tsai, S.; James, M.A. Novel Anti-CRR9/CLPTM1L Antibodies with Antitumorigenic Activity Inhibit Cell Surface Accumulation, PI3K Interaction, and Survival Signaling. Mol. Cancer Ther. 2016, 15, 985–997. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Zou, X.; Wang, K.; Wang, Y.; Wang, M.; Guo, T.; Zhong, B.; Jiang, N. Comprehensive analysis of Transcription Factors identified novel prognostic biomarker in human bladder cancer. J. Cancer 2021, 12, 5605–5621. [Google Scholar] [CrossRef]
- Gerber, A.N.; Newton, R.; Sasse, S.K. Repression of transcription by the glucocorticoid receptor: A parsimonious model for the genomics era. J. Biol. Chem. 2021, 296, 100687. [Google Scholar] [CrossRef] [PubMed]
- McBeth, L.; Grabnar, M.; Selman, S.; Hinds, T.D., Jr. Involvement of the Androgen and Glucocorticoid Receptors in Bladder Cancer. Int. J. Endocrinol. 2015, 2015, 384860. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, H.; Kawasawa, Y.I.; Shuman, L.; Zheng, Z.; Tran, T.; Walter, V.; Warrick, J.I.; Chen, G.; Al-Ahmadie, H.; Kaag, M.; et al. Repression of transcription factor AP-2 α by PPARgamma reveals a novel transcriptional circuit in basal-squamous bladder cancer. Oncogenesis 2019, 8, 69. [Google Scholar] [CrossRef]
- McEwan, I.J.; Gustafsson, J. Interaction of the human androgen receptor transactivation function with the general transcription factor TFIIF. Proc. Natl. Acad. Sci. USA 1997, 94, 8485–8490. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Rojo, E.; Berumen, L.C.; Garcia-Alcocer, G.; Escobar-Cabrera, J. The Role of Androgens and Androgen Receptor in Human Bladder Cancer. Biomolecules 2021, 11, 594. [Google Scholar] [CrossRef] [PubMed]
- Roy, A.L. Biochemistry and biology of the inducible multifunctional transcription factor TFII-I: 10 years later. Gene 2012, 492, 32–41. [Google Scholar] [CrossRef]
- Sharron Lin, X.; Hu, L.; Sandy, K.; Correll, M.; Quackenbush, J.; Wu, C.L.; Scott McDougal, W. Differentiating progressive from nonprogressive T1 bladder cancer by gene expression profiling: Applying RNA-sequencing analysis on archived specimens. Urol. Oncol. 2014, 32, 327–336. [Google Scholar] [CrossRef]
- Kashiwagi, E.; Izumi, H.; Yasuniwa, Y.; Baba, R.; Doi, Y.; Kidani, A.; Arao, T.; Nishio, K.; Naito, S.; Kohno, K. Enhanced expression of nuclear factor I/B in oxaliplatin-resistant human cancer cell lines. Cancer Sci. 2011, 102, 382–386. [Google Scholar] [CrossRef]
- Adshead, J.M.; Ogden, C.W.; Penny, M.A.; Stuart, E.T.; Kessling, A.M. The expression of PAX5 in human transitional cell carcinoma of the bladder: Relationship with de-differentiation. BJU Int. 1999, 83, 1039–1044. [Google Scholar] [CrossRef] [PubMed]
- Esrig, D.; Elmajian, D.; Groshen, S.; Freeman, J.A.; Stein, J.P.; Chen, S.C.; Nichols, P.W.; Skinner, D.G.; Jones, P.A.; Cote, R.J. Accumulation of nuclear p53 and tumor progression in bladder cancer. N. Engl. J. Med. 1994, 331, 1259–1264. [Google Scholar] [CrossRef] [PubMed]
- Huh, H.D.; Kim, D.H.; Jeong, H.S.; Park, H.W. Regulation of TEAD Transcription Factors in Cancer Biology. Cells 2019, 8, 600. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Yoon, K.; Sterneck, E.; Johnson, P.F.; Smart, R.C. CCAAT/enhancer binding protein-β is a mediator of keratinocyte survival and skin tumorigenesis involving oncogenic Ras signaling. Proc. Natl. Acad. Sci. USA 2002, 99, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Kiuru, M.; Busam, K.J. The NF1 gene in tumor syndromes and melanoma. Lab. Investig. 2017, 97, 146–157. [Google Scholar] [CrossRef]
- Kim, W.G.; Cheng, S.Y. Thyroid hormone receptors and cancer. Biochim. Biophys. Acta 2013, 1830, 3928–3936. [Google Scholar] [CrossRef]
- Liu, Z.; Zhu, Y.; Li, F.; Xie, Y. GATA1-regulated JAG1 promotes ovarian cancer progression by activating Notch signal pathway. Protoplasma 2020, 257, 901–910. [Google Scholar] [CrossRef]
- Yu, J.; Liu, M.; Liu, H.; Zhou, L. GATA1 promotes colorectal cancer cell proliferation, migration and invasion via activating AKT signaling pathway. Mol. Cell. Biochem. 2019, 457, 191–199. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Minisatellite | Forward Primer Sequence | Reverse Primer Sequence |
|---|---|---|
| MS1 | CCCTCTCTGCTGGGCTCTCC | CTCCGCCTCGATCTGCTGTT |
| MS2 | TGAACAAGTGGAGAGCAAGGAAA | ACATGGTGGGATTTCTCAAGCAC |
| MS3 | GTCTCTGCAGTTGGTGGCCC | CTGAGAGCCGCCACCTCATG |
| MS4 | CCACCAGGCTTCATGGGAGG | AGTGCTGAGCCTGGTTCTC |
| Characteristic | Cancer-Free Controls, N (%) | Bladder Cancer Cases, N (%) | p | |
|---|---|---|---|---|
| Age | 50–59 | 128 (29.02) | 46 (25.41) | 0.935 |
| 60–69 | 164 (37.19) | 68 (37.57) | ||
| 70–79 | 127 (28.80) | 56 (30.94) | ||
| ≥80 | 22 (4.99) | 11 (6.08) | ||
| Average | 65.25 | 66.52 | ||
| Median | 65 | 67 | ||
| N | 441 | 181 | ||
| TR Group | Repeats | Size (bp) | Cancer-Free Controls | Bladder Cancer | OR (95% CI) | p | ||
|---|---|---|---|---|---|---|---|---|
| N = 882 | Frequency | N = 362 | Frequency | |||||
| SR | 15 | 515 | 2 | 0.0023 | 0 | - | ND | 0.3645 |
| 17 | 573 | 5 | 0.0057 | 1 | 0.0028 | 0.49 (0.06–4.17) | 0.5020 | |
| 20 | 660 | 2 | 0.0023 | 0 | - | ND | 0.3645 | |
| Total | 9 | 0.0102 | 1 | 0.0028 | 0.27 (0.03–2.13) | 0.1820 | ||
| SC | 22 | 718 | 22 | 0.0249 | 10 | 0.0276 | 1.11 (0.52–2.37) | 0.7862 |
| 23 | 747 | 722 | 0.8186 | 287 | 0.7928 | 0.85 (0.62–1.15) | 0.2914 | |
| 24 | 776 | 25 | 0.0283 | 8 | 0.0221 | 0.77 (0.35–1.73) | 0.5335 | |
| Total | 769 | 0.8719 | 305 | 0.8425 | 0.79 (0.56–1.11) | 0.1710 | ||
| MR | 25 | 805 | 0 | - | 3 | 0.0083 | ND | 0.0068 * |
| 27 | 863 | 0 | - | 2 | 0.0055 | ND | 0.0272 * | |
| 28 | 892 | 1 | 0.0011 | 0 | - | ND | 0.5216 | |
| 29 | 921 | 2 | 0.0023 | 2 | 0.0055 | 2.44 (0.34–17.42) | 0.3567 | |
| Total | 3 | 0.0034 | 7 | 0.0110 | 5.78 (1.49–22.47) | 0.0040 * | ||
| LC | 30 | 950 | 97 | 0.1100 | 45 | 0.1243 | 1.15 (0.79–1.67) | 0.4700 |
| Total | 97 | 0.1156 | 45 | 0.1409 | 1.15 (0.79–1.67) | 0.4700 | ||
| LR | 31 | 979 | 4 | 0.0045 | 3 | 0.0083 | 1.83 (0.41–8.24) | 0.4216 |
| 32 | 1008 | 0 | - | 1 | 0.0028 | ND | 0.1184 | |
| Total | 4 | 0.1202 | 4 | 0.1519 | 2.45 (0.61–9.86) | 0.1920 | ||
| Genotype Group | Genotype | Cancer-Free Controls | Bladder Cancer | OR (95% CI) | p | ||
|---|---|---|---|---|---|---|---|
| N = 441 | Frequency | N = 181 | Frequency | ||||
| SR/– | 15/23 | 2 | 0.0045 | 0 | - | ND | 0.3642 |
| 17/23 | 5 | 0.0113 | 0 | - | ND | 0.1503 | |
| 17/31 | 0 | - | 1 | 0.0055 | ND | 0.1182 | |
| 20/23 | 1 | 0.0023 | 0 | - | ND | 0.5214 | |
| 20/30 | 1 | 0.0023 | 0 | - | ND | 0.5214 | |
| Total | 9 | 0.0204 | 1 | 0.0055 | 0.27 (0.03–2.12) | 0.1811 | |
| MR/– | 23/25 | 0 | - | 3 | 0.0166 | ND | 0.0067 * |
| 23/27 | 0 | - | 1 | 0.0055 | ND | 0.1182 | |
| 23/28 | 1 | 0.0023 | 0 | - | ND | 0.5214 | |
| 23/29 | 2 | 0.0045 | 2 | 0.0110 | 2.45 (0.34–17.55) | 0.3559 | |
| 27/30 | 0 | - | 1 | 0.0055 | ND | 0.1182 | |
| Total | 3 | 0.0068 | 7 | 0.0367 | 5.87 (1.50–22.97) | 0.0041 * | |
| LR/– | 17/31 | 0 | - | 1 | 0.0055 | ND | 0.1182 |
| 23/31 | 2 | 0.0045 | 2 | 0.0110 | 2.45 (0.34–17.55) | 0.3559 | |
| 23/32 | 0 | - | 1 | 0.0055 | ND | 0.1182 | |
| 30/31 | 2 | 0.0045 | 0 | - | ND | 0.3642 | |
| Total | 4 | 0.0091 | 4 | 0.2210 | 2.47 (0.61–9.98) | 0.1902 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, M.-S.; Mun, J.-Y.; Yang, G.-E.; Kim, M.-H.; Lee, S.-Y.; Choi, Y.H.; Kim, H.S.; Nam, J.-K.; Kim, T.N.; Leem, S.-H. Exploring the Relationship between CLPTM1L-MS2 Variants and Susceptibility to Bladder Cancer. Genes 2024, 15, 50. https://doi.org/10.3390/genes15010050
Jeong M-S, Mun J-Y, Yang G-E, Kim M-H, Lee S-Y, Choi YH, Kim HS, Nam J-K, Kim TN, Leem S-H. Exploring the Relationship between CLPTM1L-MS2 Variants and Susceptibility to Bladder Cancer. Genes. 2024; 15(1):50. https://doi.org/10.3390/genes15010050
Chicago/Turabian StyleJeong, Mi-So, Jeong-Yeon Mun, Gi-Eun Yang, Min-Hye Kim, Sang-Yeop Lee, Yung Hyun Choi, Heui Soo Kim, Jong-Kil Nam, Tae Nam Kim, and Sun-Hee Leem. 2024. "Exploring the Relationship between CLPTM1L-MS2 Variants and Susceptibility to Bladder Cancer" Genes 15, no. 1: 50. https://doi.org/10.3390/genes15010050