
Inheritance and Quantitative Trait Loci Mapping of Aromatic Compounds from Clementine (Citrus × clementina Hort. ex Tan.) and Sweet Orange (C. × sinensis (L.) Osb.) Fruit Essential Oils
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Biological Material
2.2. Essential Oil Extraction
2.3. Chemical Composition Analysis
2.4. Sensorial Analysis
2.5. Statistical Analysis
2.6. GBS and Marker Filtering
2.7. Construction of the Consensus Genetic Map
2.8. QTL Detection
3. Results
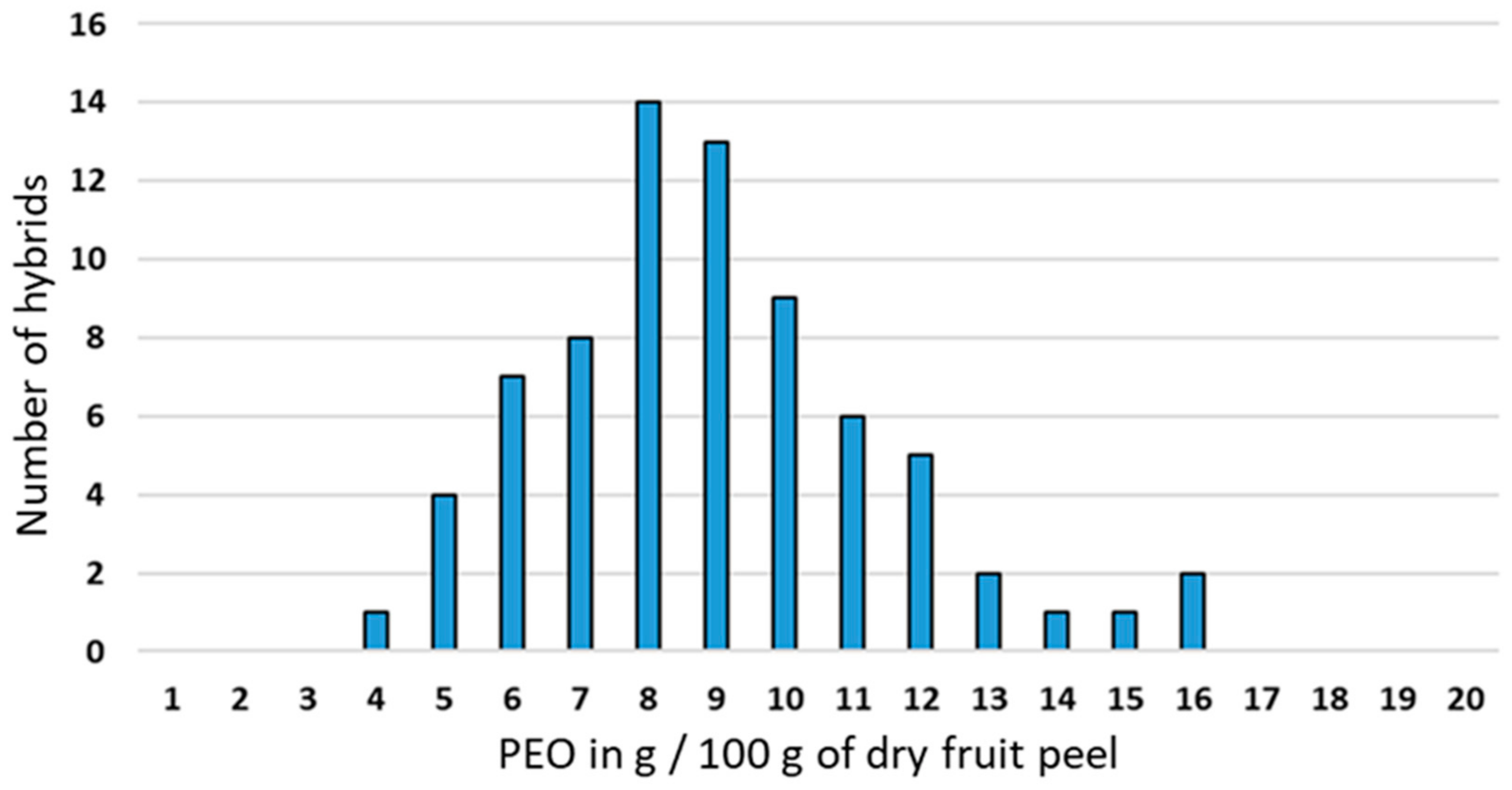
3.1. Essential Oil Yield of the Segregating Population
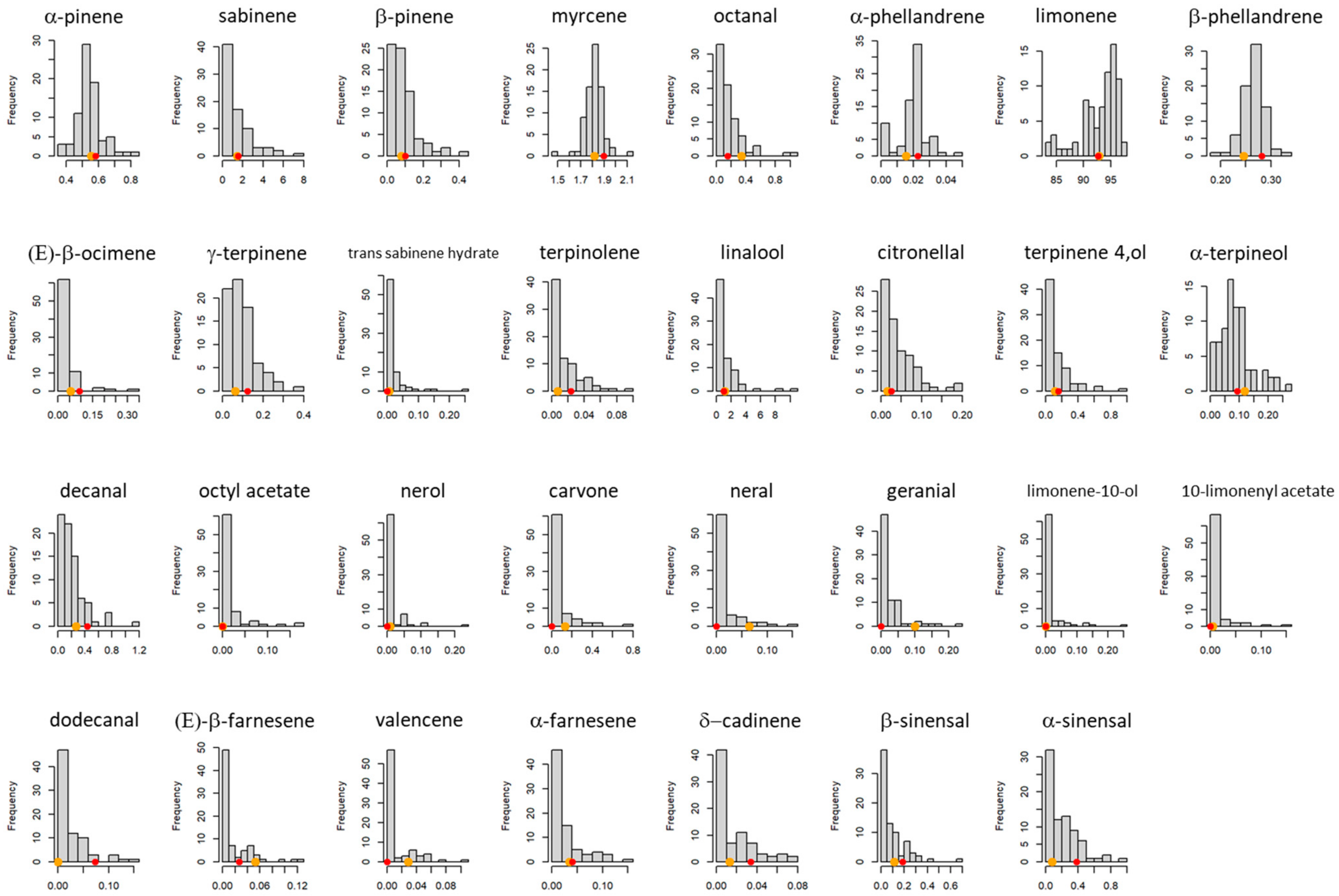
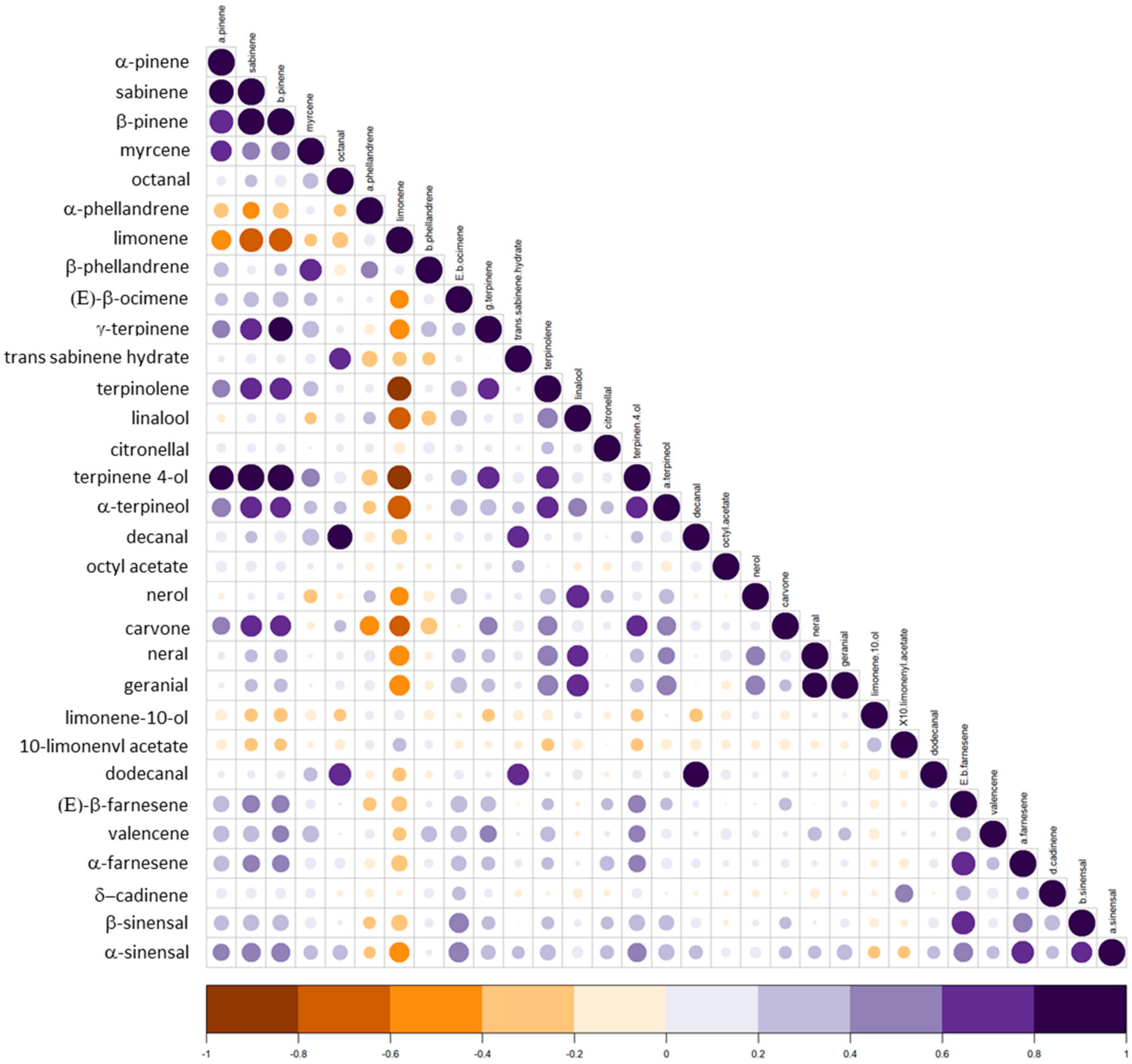
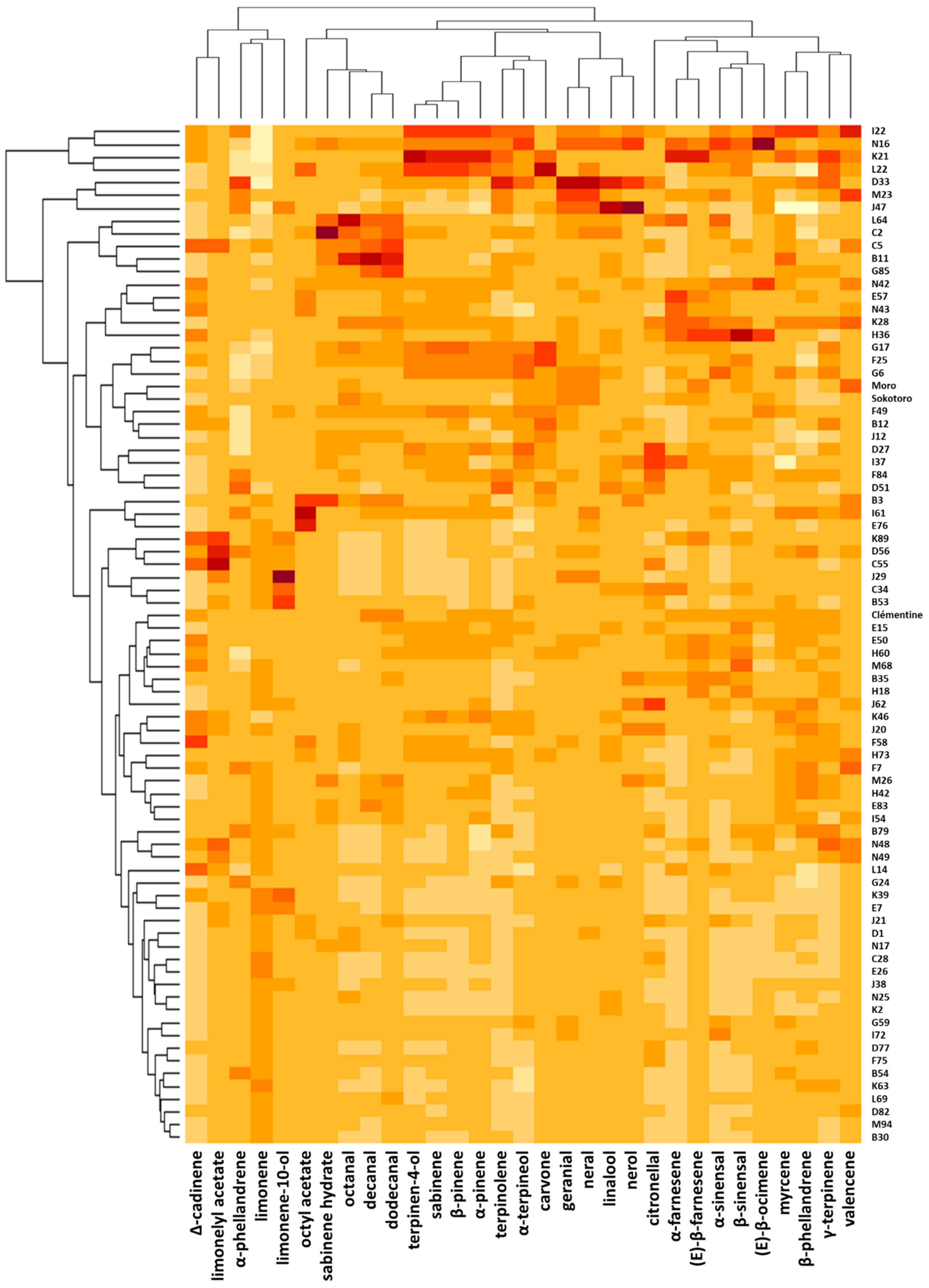
3.2. Composition, Segregation and Variation of PEO Compounds in the Progeny and Their Parents
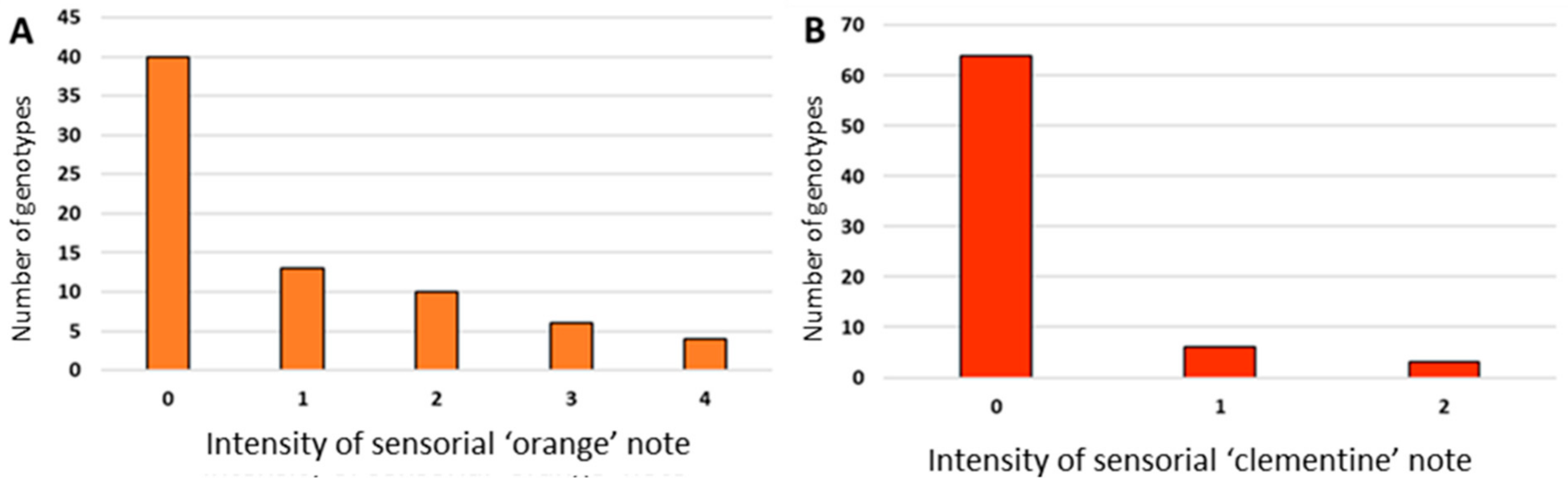
3.3. Sensorial Profile of the Hybrids
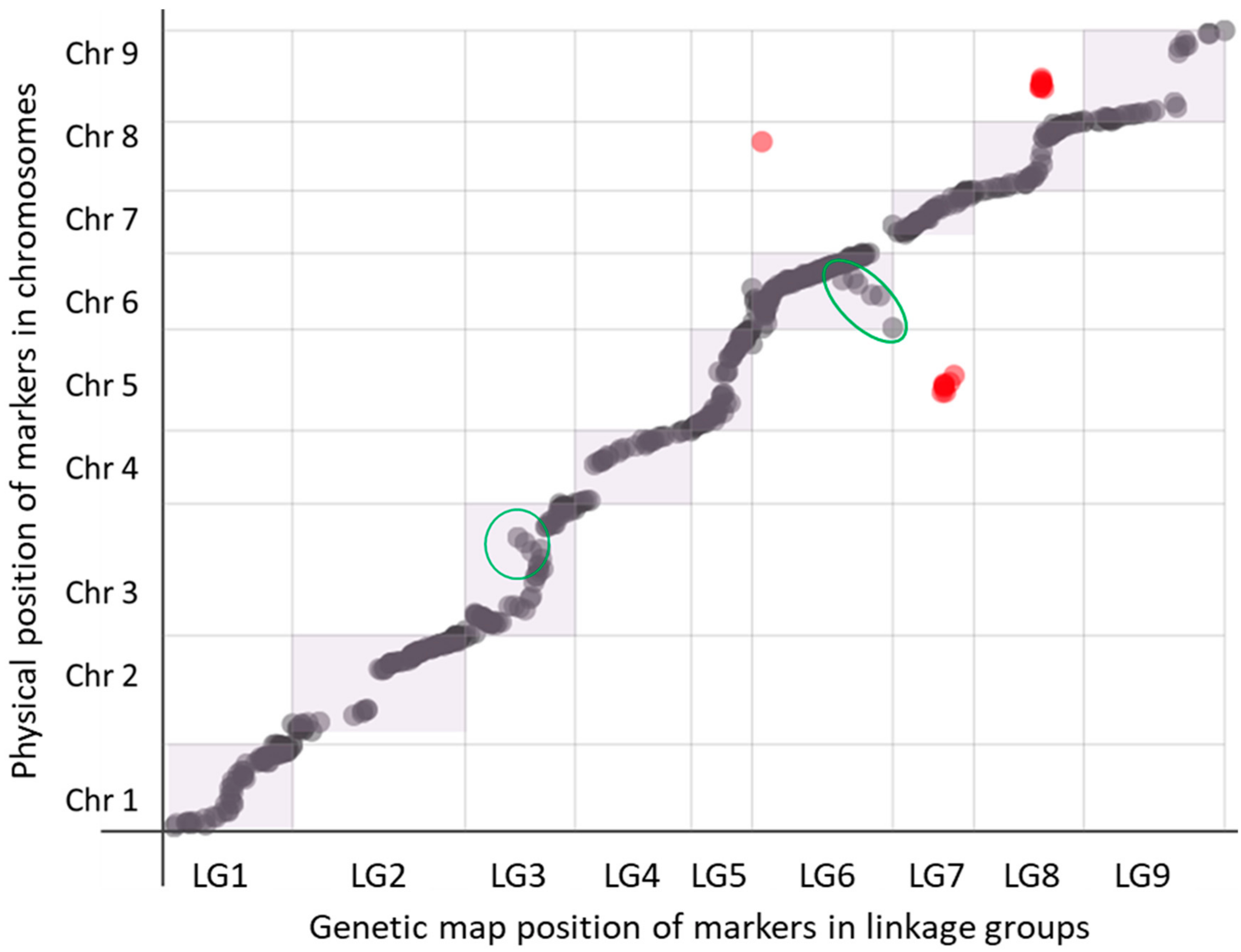
3.4. Genetic Map
3.5. Inheritance of Volatile Compounds and Oil Yield
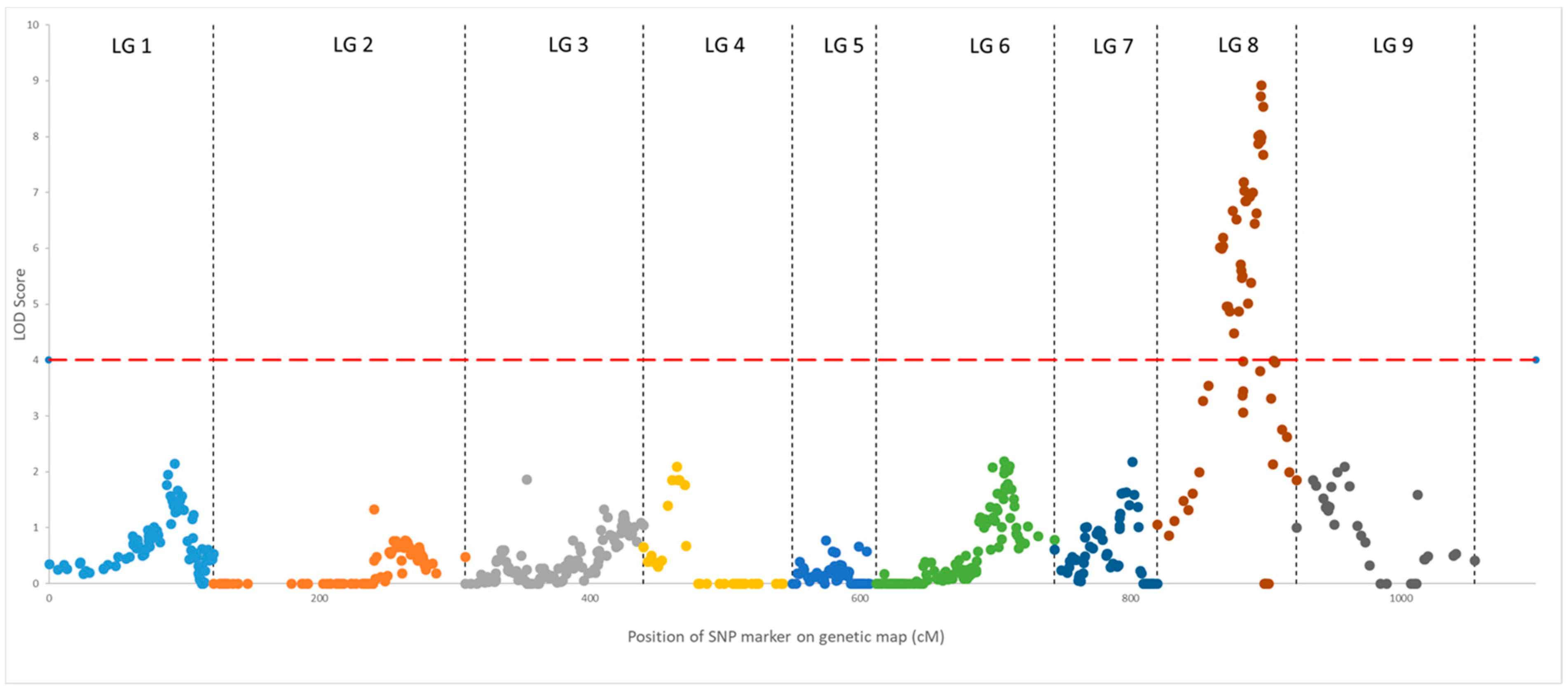
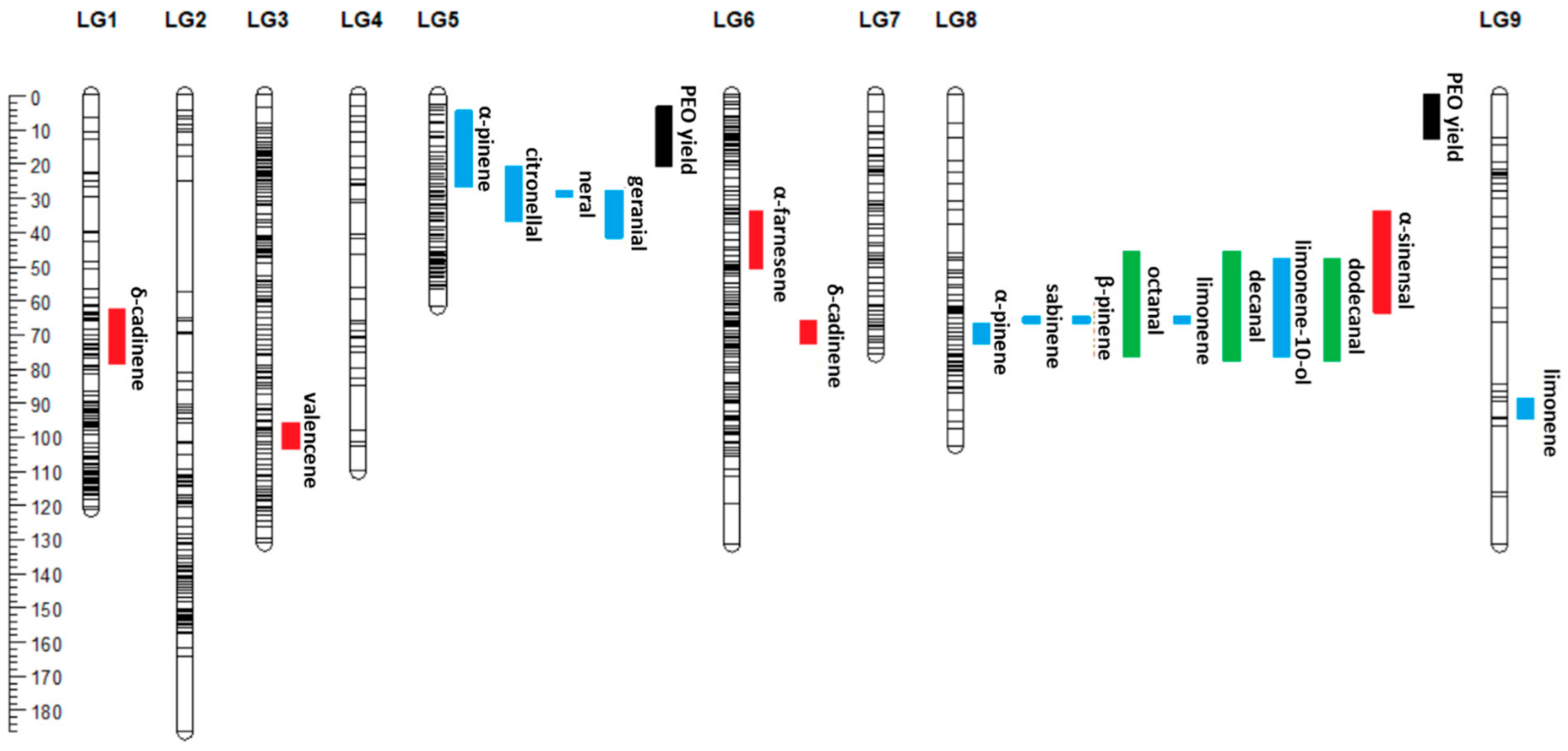
3.6. Research on the QTLs and Genes Associated with Terpenoid Biosynthesis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Teigiserova, D.A.; Tiruta-Barna, L.; Ahmadi, A.; Hamelin, L.; Thomsen, M.A. Step Closer to Circular Bioeconomy for Citrus Peel Waste: A Review of Yields and Technologies for Sustainable Management of Essential Oils. J. Environ. Manag. 2021, 280, 111832. [Google Scholar] [CrossRef] [PubMed]
- Ferrer, V.; Paymal, N.; Costantino, G.; Paoli, M.; Quinton, C.; Tomi, F.; Luro, F. Correspondence between the compositional and aromatic diversity of leaf and fruit essential oils and pomological diversity of 43 sweet oranges (Citrus x aurantium var sinensis L.). Plants 2023, 12, 990. [Google Scholar] [CrossRef] [PubMed]
- González-Mas, M.C.; Rambla, J.L.; López-Gresa, M.P.; Amparo Blázquez, M.; Granell, A. Volatile Compounds in Citrus Essential Oils: A Comprehensive Review. Front. Plant Sci. 2019, 10, 12. [Google Scholar] [CrossRef] [PubMed]
- Gershenzon, J.; Dudareva, N. The Function of Terpene Natural Products in the Natural World. Nat. Chem. Biol. 2007, 3, 408–414. [Google Scholar] [CrossRef] [PubMed]
- Tholl, D. Terpene Synthases and the Regulation, Diversity and Biological Roles of Terpene Metabolism. Curr. Opin. Plant Biol. 2006, 9, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Degenhardt, J.; Köllner, T.G.; Gershenzon, J. Monoterpene and Sesquiterpene Synthases and the Origin of Terpene Skeletal Diversity in Plants. Phytochemistry 2009, 70, 1621–1637. [Google Scholar] [CrossRef] [PubMed]
- Dudareva, N.; Klempien, A.; Muhlemann, J.K.; Kaplan, I. Biosynthesis, Function and Metabolic Engineering of Plant Volatile Organic Compounds. New Phytol. 2013, 198, 16–32. [Google Scholar] [CrossRef] [PubMed]
- Bohlmann, J.; Meyer-Gauen, G.; Croteau, R. Plant Terpenoid Synthases: Molecular Biology and Phylogenetic Analysis. Proc. Natl. Acad. Sci. USA 1998, 95, 4126–4133. [Google Scholar] [CrossRef]
- Maruyama, T.; Ito, M.; Honda, G. Molecular Cloning, Functional Expression and Characterization of (E)-b-Farnesene Synthase from Citrus junos. Biol. Pharm. Bull. 2001, 24, 1171–1175. [Google Scholar] [CrossRef]
- Lücker, J.; El Tamer, M.K.; Schwab, W.; Verstappen, F.W.A.; Van der Plas, L.H.W.; Bouwmeester, H.J.; Verhoeven, H.A. Monoterpene Biosynthesis in Lemon (Citrus limon): CDNA Isolation and Functional Analysis of Four Monoterpene Synthases. Eur. J. Biochem. 2002, 269, 3160–3171. [Google Scholar] [CrossRef]
- Sharon-Asa, L.; Shalit, M.; Frydman, A.; Bar, E.; Hollandetti, O.D.; Lavi, U.; Lewinsohn, E.; Eyal, Y. Citrus Fruit Flavor and Aroma Biosynthesis: Isolation, Functional Characterization, and Developmental Regulation of Cstps1, a Key Gene in the Production of the Sesquiterpene Aroma Compound Valencene: Characterization and Regulation of Valencene Synthase. Plant J. 2003, 36, 664–674. [Google Scholar] [CrossRef] [PubMed]
- Shimada, T.; Endo, T.; Fujii, H.; Omura, M. Isolation and Characterization of a New D-Limonene Synthase Gene with a Different Expression Pattern in Citrus unshiu Marc. Sci. Hortic. 2005, 105, 507–512. [Google Scholar] [CrossRef]
- Shimada, T.; Endo, T.; Fujii, H.; Hara, M.; Ueda, T.; Kita, M.; Omura, M. Molecular Cloning and Functional Characterization of Four Monoterpene Synthase Genes from Citrus unshiu Marc. Plant Sci. 2004, 166, 49–58. [Google Scholar] [CrossRef]
- Dornelas, M.C.; Mazzafera, P.A. Genomic Approach to Characterization of the Citrus Terpene Synthase Gene Family. Genet. Mol. Biol. 2007, 30, 832–840. [Google Scholar] [CrossRef]
- Kohzaki, K.; Gomi, K.; Yamasaki-Kokudo, Y.; Ozawa, R.; Takabayashi, J.; Akimitsu, K. Characterization of a Sabinene Synthase Gene from Rough Lemon (Citrus jambhiri). J. Plant Physiol. 2009, 166, 1700–1704. [Google Scholar] [CrossRef]
- Rodríguez, A.; San Andrés, V.; Cervera, M.; Redondo, A.; Alquézar, B.; Shimada, T.; Gadea, J.; Rodrigo, M.; Zacarías, L.; Palou, L.; et al. The Monoterpene Limonene in Orange Peels Attracts Pests and Microorganisms. Plant Signal. Behav. 2011, 6, 1820–1823. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, A.; Shimada, T.; Cervera, M.; Redondo, A.; Alquézar, B.; Rodrigo, M.J.; Zacarías, L.; Palou, L.; López, M.M.; Peña, L. Resistance to Pathogens in Terpene Down-Regulated Orange Fruits Inversely Correlates with the Accumulation of D-Limonene in Peel Oil Glands. Plant Signal. Behav. 2015, 10, e1028704. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, A.; Peris, J.E.; Redondo, A.; Shimada, T.; Costell, E.; Carbonell, I.; Rojas, C.; Peña, L. Impact of D-Limonene Synthase up- or down-Regulation on Sweet Orange Fruit and Juice Odor Perception. Food Chem. 2017, 217, 139–150. [Google Scholar] [CrossRef]
- Wu, G.A.; Prochnik, S.; Jenkins, J.; Salse, J.; Hellsten, U.; Murat, F.; Perrier, X.; Ruiz, M.; Scalabrin, S.; Terol, J.; et al. Sequencing of Diverse Mandarin, Pummelo and Orange Genomes Reveals Complex History of Admixture during Citrus Domestication. Nat. Biotechnol. 2014, 32, 656–662. [Google Scholar] [CrossRef]
- Xu, Q.; Chen, L.-L.; Ruan, X.; Chen, D.; Zhu, A.; Chen, C.; Bertrand, D.; Jiao, W.-B.; Hao, B.-H.; Lyon, M.P.; et al. The Draft Genome of Sweet Orange (Citrus sinensis). Nat. Genet. 2013, 45, 59–66. [Google Scholar] [CrossRef]
- Luro, F.; Curk, F.; Froelicher, Y.; Ollitrault, P. Recent Insights on Citrus Diversity and Phylogeny. In AGRUMED: Archaeology and History of Citrus Fruit in the Mediterranean: Acclimatization, Diversifications, Uses; Zech-Matterne, V., Girolamo, F., Eds.; Centre Jean Bérard: Naples, Italy, 2017; pp. 16–28. ISBN 9782918887775. [Google Scholar]
- Alquézar, B.; Rodríguez, A.; de la Peña, M.; Peña, L. Genomic Analysis of Terpene Synthase Family and Functional Characterization of Seven Sesquiterpene Synthases from Citrus sinensis. Front. Plant Sci. 2017, 8, 1481. [Google Scholar] [CrossRef] [PubMed]
- Kerbiriou, P.; Plotto, A.; Goodner, K.; Baldwin, E.; Gmitter, F.G. Distribution of Aroma Volatiles in a Population of Tangerine Hybrids. Proc. Fla. State Hort. Soc. 2007, 120, 267–275. [Google Scholar]
- Barboni, T.; Luro, F.; Chiaramonti, N.; Desjobert, J.-M.; Muselli, A.; Costa, J. Volatile Composition of Hybrids Citrus Juices by Headspace Solid Phase Micro Extraction/Gas Chromatography/Mass Spectrometry. Food Chem. 2009, 116, 382–390. [Google Scholar] [CrossRef]
- Miyazaki, T.; Plotto, A.; Goodner, K.; Gmitter, F.G. Distribution of Aroma Volatile Compounds in Tangerine Hybrids and Proposed Inheritance. J. Sci. Food Agric. 2011, 91, 449–460. [Google Scholar] [CrossRef] [PubMed]
- Ruberto, G.; Biondi, D.; Piattelli, M.; Rapisarda, P.; Starrantino, A. Profiles of Essential Oils of New Citrus Hybrids. Flavour Fragr. J. 1993, 8, 179–184. [Google Scholar] [CrossRef]
- Ruberto, G.; Rapisarda, P. Essential Oils of New Pigmented Citrus Hybrids: Citrus sinensis L. Osbeck x C clementina Hort. Ex Tanaka. J. Food Sci. 2002, 67, 2778–2780. [Google Scholar] [CrossRef]
- Verzera, A.; Trozzi, A.; Zappalá, M.; Condurso, C.; Cotroneo, A. Essential Oil Composition of Citrus meyerii Y. Tan. and Citrus medica L. Cv. Diamante and Their Lemon Hybrids. J. Agric. Food Chem. 2005, 53, 4890–4894. [Google Scholar] [CrossRef]
- Verzera, A.; Tripodi, G.; Cotroneo, A. Characteristics of a New Citrus Hybrid Essential Oil, Citrus clementina Cv. Nules x Citrus limon Cv. Cavone. J. Essent. Oil Bear. Plants 2009, 12, 293–299. [Google Scholar] [CrossRef]
- Fabroni, S.; Ruberto, G.; Rapisarda, P. Essential Oil Profiles of New Citrus Hybrids, a Tool for Genetic Citrus Improvement. J. Essent. Oil Res. 2012, 24, 159–169. [Google Scholar] [CrossRef]
- Tomi, F.; Barzalona, M.; Casanova, J.; Luro, F. Chemical Variability of the Leaf Oil of 113 Hybrids from Citrus clementina (Commun) × Citrus deliciosa (Willow Leaf). Flavour Fragr. J. 2008, 23, 152–163. [Google Scholar] [CrossRef]
- Yu, Y.; Chen, C.; Gmitter, F.G. QTL Mapping of Mandarin (Citrus reticulata) Fruit Characters Using High-Throughput SNP Markers. Tree Genet. Genomes 2016, 12, 77. [Google Scholar] [CrossRef]
- Imai, A.; Yoshioka, T.; Hayashi, T. Quantitative Trait Locus (QTL) Analysis of Fruit Quality Traits for Mandarin Breeding in Japan. Tree Genet. Genomes 2017, 13, 79. [Google Scholar] [CrossRef]
- Curtolo, M.; Cristofani-Yaly, M.; Gazaffi, R.; Takita, M.A.; Figueira, A.; Machado, M.A. QTL Mapping for Fruit Quality in Citrus Using DArTseq Markers. BMC Genom. 2017, 18, 289. [Google Scholar] [CrossRef] [PubMed]
- Khefifi, H.; Dumont, D.; Costantino, G.; Doligez, A.; Anna Carla, B.; Bérard, A.; Morillon, R.; Ollitrault, P.; Luro, F. Mapping of QTLs for citrus quality traits throughout the fruit maturation process on clementine (Citrus reticulata x C. sinensis) and mandarin (C. reticulata Blanco) genetic maps. TGG 2022, 18, 40. [Google Scholar] [CrossRef]
- Yu, Y.; Bai, J.; Chen, C.; Plotto, A.; Yu, Q.; Baldwin, E.A.; Gmitter, F.G. Identification of QTLs Controlling Aroma Volatiles Using a ‘Fortune’ x ‘Murcott’ (Citrus reticulata) Population. BMC Genom. 2017, 18, 646. [Google Scholar] [CrossRef] [PubMed]
- Shimada, T.; Endo, T.; Fujii, H.; Rodríguez, A.; Yoshioka, T.; Peña, L.; Omura, M. Biological and Molecular Characterization of Linalool-Mediated Field Resistance against Xanthomonas citri Subsp. citri in Citrus Trees. Tree Physiol. 2021, 41, 2171–2188. [Google Scholar] [CrossRef] [PubMed]
- Butelli, E.; Licciardello, C.; Zhang, Y.; Liu, J.; Mackay, S.; Bailey, P.; Reforgiato-Recupero, G.; Martin, C. Retrotransposons Control Fruit-Specific, Cold-Dependent Accumulation of Anthocyanins in Blood Oranges. Plant Cell 2012, 24, 242–1255. [Google Scholar] [CrossRef]
- Luro, F.; Bloquel, E.; Tomu, B.; Costantino, G.; Tur, I.; Riolacci, S.; Varamo, F.; Ollitrault, P.; Froelicher, Y.; Curk, F.; et al. The INRA-CIRAD citrus germplasm collection of San Giuliano, Corsica. In AGRUMED: Archaeology and History of Citrus Fruit in the Mediterranean: Acclimatization, Diversifications, Uses; Zech-Matterne, V., Girolamo, F., Eds.; Centre Jean Bérard: Naples, Italy, 2017; pp. 243–261. ISBN 9782918887775. [Google Scholar]
- Bicchi, C.; Liberto, E.; Matteodo, M.; Sgorbini, B.; Mondello, L.; d’Acampora Zellner, B.; Costa, R.; Rubiolo, P. Quantitative Analysis of Essential Oils: A Complex Task. Flavour Fragr. J. 2008, 23, 382–391. [Google Scholar] [CrossRef]
- Oueslati, A.; Salhi-Hannachi, A.; Luro, F.; Vignes, H.; Mournet, P.; Ollitrault, P. Genotyping by Sequencing Reveals the Interspecific C. maxima/C. reticulata Admixture along the Genomes of Modern Citrus Varieties of Mandarins, Tangors, Tangelos, Orangelos and Grapefruits. PLoS ONE 2017, 12, e0185618. [Google Scholar] [CrossRef]
- Sonah, H.; Bastien, M.; Iquira, E.; Tardivel, A.; Légaré, G.; Boyle, B.; Normandeau, E.; Laroche, J.; LaRose, S.; Jean, M.; et al. An Improved Genotyping by Sequencing (GBS) Approach Offering Increased Versatility and Efficiency of SNP Discovery and Genotyping. PLoS ONE 2013, 8, e54603. [Google Scholar] [CrossRef]
- Turner, S.D. qqman: An R package for visualizing GWAS results using Q-Q and manhattan plots. J. Open Source Softw. 2018, 3, 731. [Google Scholar] [CrossRef]
- Luro, F.; Baccati, C.; Paoli, M.; Marchi, E.; Costantino, G.; Gibernau, M.; Ollitrault, P.; Tomi, F. Phylogenetic and taxonomic status of Citrus halimii B.C. Stone determined by genotyping complemented by chemical analysis of leaf and fruit rind essential oils. Sci. Hortic. 2022, 299, 111018. [Google Scholar] [CrossRef]
- Lota, M.-L.; de Rocca Serra, D. Chemical Composition of Peel and Leaf Essential Oils of Citrus medica L. and C. limonimedica Lush. Flavour Fragr. J. 1999, 14, 161–166. [Google Scholar] [CrossRef]
- Lota, M.-L.; de Rocca Serra, D.; Tomi, F.; Casanova, J. Chemical Variability of Peel and Leaf Essential Oils of Mandarins from Citrus reticulata Blanco. Biochem. Syst. Ecol. 2000, 28, 61–78. [Google Scholar] [CrossRef]
- Lota, M.-L.; de Rocca Serra, D.; Tomi, F.; Jacquemond, C.; Casanova, J. Volatile Components of Peel and Leaf Oils of Lemon and Lime Species. J. Agric. Food Chem. 2002, 50, 796–805. [Google Scholar] [CrossRef] [PubMed]
- Ferrer, V.; Paymal, N.; Quinton, C.; Tomi, F.; Luro, F. Investigations of the Chemical Composition and Aromatic Properties of Peel Essential Oils throughout the Complete Phase of Fruit Development for Two Cultivars of Sweet Orange (Citrus sinensis (L.) Osb.). Plants 2022, 11, 2747. [Google Scholar] [CrossRef] [PubMed]
- Paun, O.; Fay, M.F.; Soltis, D.E.; Chase, M.W. Genetic and Epigenetic Alterations after Hybridization and Genome Doubling. TAXON 2007, 56, 649–656. [Google Scholar] [CrossRef]
- Miyazaki, T.; Plotto, A.; Baldwin, E.A.; Reyes-De-Corcuera, J.I.; Gmitter, F.G., Jr. Aroma Characterization of Tangerine Hybrids by Gas-Chromatography Olfactometry and Sensory Evaluation. J. Sci. Food Agric. 2012, 92, 727–735. [Google Scholar] [CrossRef]
- Perez-Cacho, P.R.; Rouseff, R.L. Fresh Squeezed Orange Juice Odor: A Review. Food Sci. Nutr. 2008, 48, 681–695. [Google Scholar] [CrossRef]
- Eduardo, I.; Chietera, G.; Pirona, R.; Pacheco, I.; Troggio, M.; Banchi, E.; Bassi, D.; Rossini, L.; Vecchietti, A.; Pozzi, C. Genetic Dissection of Aroma Volatile Compounds from the Essential Oil of Peach Fruit: QTL Analysis and Identification of Candidate Genes Using Dense SNP Maps. Tree Genet. Genomes 2013, 9, 189–204. [Google Scholar] [CrossRef]
- Souleyre, E.J.F.; Chagné, D.; Chen, X.; Tomes, S.; Turner, R.M.; Wang, M.Y.; Maddumage, R.; Hunt, M.B.; Winz, R.A.; Wiedow, C.; et al. The AAT1 Locus Is Critical for the Biosynthesis of Esters Contributing to ‘Ripe Apple’ Flavour in ‘Royal Gala’ and ‘Granny Smith’ Apples. Plant J. 2014, 78, 903–915. [Google Scholar] [CrossRef] [PubMed]
- Shimada, T.; Endo, T.; Fujii, H.; Hara, M.; Omura, M. Isolation and Characterization of (E)-Beta-Ocimene and 1,8 Cineole Synthases in Citrus unshiu Marc. Plant Sci. 2005, 168, 987–995. [Google Scholar] [CrossRef]
- De los Reyes, B.G. Genomic and Epigenomic Bases of Transgressive Segregation—New Breeding Paradigm for Novel Plant Phenotypes. Plant Sci. 2019, 288, 110213. [Google Scholar] [CrossRef] [PubMed]
- Högnadóttir, Á.; Rouseff, R.L. Identification of Aroma Active Compounds in Orange Essence Oil Using Gas Chromatography–Olfactometry and Gas Chromatography–Mass Spectrometry. J. Chromatogr. 2003, 998, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Elston, A.; Lin, J.; Rouseff, R. Determination of the Role of Valencene in Orange Oil as a Direct Contributor to Aroma Quality. Flavour Fragr. J. 2005, 20, 381–386. [Google Scholar] [CrossRef]
- Qiao, Y.; Xie, B.; Zhang, Y.; Zhang, Y.; Fan, G.; Yao, X.; Pan, S. Characterization of Aroma Active Compounds in Fruit Juice and Peel Oil of Jinchen Sweet Orange Fruit (Citrus sinensis (L.) Osbeck) by GC-MS and GC-O. Molecules 2008, 13, 1333–1344. [Google Scholar] [CrossRef] [PubMed]
- Gaffney, B.M.; Havekotte, M.; Jacobs, B.; Costa, L.; Ho, C.-T.H. CharmAnalysis of Two Citrus sinensis Peel Oil Volatiles. Perfum. Flavorist 1996, 21, 1–6. [Google Scholar]
- Buettner, A.; Mestres, M.; Fischer, A.; Guasch, J.; Schieberle, P. Evaluation of the Most Odour-Active Compounds in the Peel Oil of Clementines (Citrus reticulata Blanco Cv. Clementine). Eur. Food Res. Technol. 2003, 216, 11–14. [Google Scholar] [CrossRef]
- Deterre, S.; Leclair, C.; Bai, J.; Baldwin, E.A.; Narciso, J.A.; Plotto, A. Chemical and Sensory Characterization of Orange (Citrus sinensis) Pulp, a by Product of Orange Juice Processing Using Gas-Chromatography-Olfactometry. J. Food Qual. 2016, 39, 826–838. [Google Scholar] [CrossRef]
- Cuenca, J.; Aleza, P.; Garcia-Lor, A.; Ollitrault, P.; Navarro, L. Fine mapping for identification of citrus alternaria brown spot candidate resistance genes and development of new SNP markers for marker-assisted selection. Front. Plant Sci. 2016, 7, 13. [Google Scholar] [CrossRef]
- Montalt, R.; Cuenca, J.; Vives, M.C.; Mournet, P.; Navarro, L.; Ollitrault, P.; Aleza, P. Genotyping by sequencing for SNP-Based linkage analysis and the development of KASPar markers for male sterility and polyembryony in citrus. Plants Adv. Breed. Genet. Genom. Citrus 2023, 12, 1567. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LG | Markers | Size (cM) | Mean Distance between SNPs (cM) | Max Distance between 2 SNP (cM)s |
|---|---|---|---|---|
| 1 | 99 | 121 | 1.3 | 10.3 |
| 2 | 98 | 187 | 2.0 | 32.1 |
| 3 | 132 | 131 | 1.0 | 4.4 |
| 4 | 32 | 110 | 3.6 | 12.8 |
| 5 | 69 | 62 | 0.9 | 5.1 |
| 6 | 127 | 132 | 1.0 | 11.9 |
| 7 | 60 | 76 | 1.3 | 5.1 |
| 8 | 63 | 103 | 1.7 | 8.6 |
| 9 | 30 | 132 | 4.5 | 19.1 |
| Total | 710 | 1054 | - | - |
| Compound | LG 1 | QTL Map Position 2 | Physical Position 3 | Max LOD 4 | LOD Threshold 5 | Var (%) 6 | K-W 7 |
|---|---|---|---|---|---|---|---|
| α-pinene | 5 | 5–21–27 | scaff_5_1891859–scaff_5_7341652–scaff_5_8671671 | 4.3 | 3.5 | 25.8 | |
| 8 | 67–67–73 | scaff_8_19483932–scaff_8_19483932–scaff_8_18742028 | 4.3 | 3.6 | 25.6 | ***** | |
| sabinene | 8 | 65–67–67 | scaff_8_17641584–scaff_8_19483932–scaff_8_19483932 | 5.4 | 4.4 | 31.0 | ******* |
| β-pinene | 8 | 65–67–67 | scaff_8_17641584–scaff_8_19483932–scaff_8_19483932 | 4.8 | 4.1 | 28.0 | ****** |
| octanal | 8 | 46–75–77 | scaff_8_2499192–scaff_8_19803231–scaff_8_20270007 | 5.6 | 4.3 | 31.9 | ******* |
| limonene | 9 | 89–90–95 | scaff_9_24887259–scaff_9_24887259–scaff_9_27099604 | 4.8 | 3.3 | 27.9 | ******* |
| 8 | 65–67–67 | scaff_8_17641584–scaff_8_19483932–scaff_8_19483932 | 3.7 | 3.7 | 22.5 | ***** | |
| citronellal | 5 | 21–37–37 | scaff_5_7341652–scaff_5_24327337–scaff_5_24327337 | 5.9 | 3.4 | 33.2 | ****** |
| decanal | 8 | 46–77–78 | scaff_8_2499192–scaff_8_20270007–scaff_8_20292393 | 8.9 | 4.0 | 45.8 | ******* |
| neral | 5 | 28–30–30 | scaff_5_11337482–scaff_5_12169332–scaff_5_12169332 | 3.8 | 3.7 | 22.8 | ** |
| geranial | 5 | 28–30–42 | scaff_5_11337482–scaff_5_12169332–scaff_5_27088110 | 5.3 | 3.7 | 30.7 | ****** |
| limonene-10-ol | 8 | 48–52–77 | scaff_8_2628498–scaff_8_4208594–scaff_8_20270007 | 11.6 | 4.7 | 55.0 | ******* |
| dodecanal | 8 | 48–77–78 | scaff_8_2359199–scaff_8_20270007–scaff_8_20292393 | 6.7 | 4.0 | 37.1 | ******* |
| valencene | 3 | 96–104–104 | scaff_3_19904227–scaff_3_21279770–scaff_3_21279770 | 5.6 | 4.7 | 32.0 | *** |
| α-farnesene | 6 | 34–51–51 | scaff_6_15868752–scaff_6_17711788–scaff_6_17893337 | 3.8 | 3.6 | 23.0 | ****** |
| δ-cadinene | 1 | 63–79–79 | scaff_1_10400304–scaff_1_20460221–scaff_1_219122840 | 5.9 | 3.5 | 33.5 | ******* |
| 6 | 66–73–73 | scaff_6_19700867–scaff_6_20697959–scaff_6_20697959 | 4.3 | 3.4 | 25.4 | ***** | |
| α-sinensal | 8 | 34–63–64 | scaff_8_2413159–scaff_8_13317923–scaff_8_17514156 | 5.6 | 3.7 | 33.3 | ******* |
| PEO yield | 5 | 3–8–21 | scaff_5_1134031–scaff_5_2137822–scaff_5_3423139 | 4.9 | 3.4 | 28.7 | ****** |
| 8 | 0–9–13 | scaff_8_68590–scaff_8_33631–scaff_8_341083 | 3.3 | 3.0 | 20.5 | ****** |
| QTL | Scaf. | Gene | Start | End | Protein Name | Pathway |
|---|---|---|---|---|---|---|
| δ-cadinene | 1 | CICLE_v10008448mg | 16657241 | 16660189 | tyrosine transaminase family protein | Ubiquinone and other terpenoid–quinone biosynthesis |
| δ-cadinene | 1 | CICLE_v10008222mg | 15860208 | 15866603 | tRNA dimethylallyltransferase | Zeatin biosynthesis |
| δ-cadinene | 6 | CICLE_v10012247mg | 20655671 | 20659398 | preny protein peptidase | Terpenoid backbone biosynthesis |
| α-pinene | 5 | CICLE_v10000884mg | 4487163 | 4490184 | GA requiring 3 | Isoprenoid biosynthetic process |
| CICLE_v10000902mg | 4498792 | 4501743 | ||||
| CICLE_V10001179MG | 4550039 | 4553132 | ||||
| α-pinene, citronellal | 5 | CICLE_v10001398mg | 9330188 | 9333235 | geranylgeranyl transferase | Ubiquinone and other terpenoid–quinone biosynthesis |
| 5 | CICLE_v10003969mg | 8303490 | 8304788 | geranylgeranyl pyrophosphate synthase | Terpenoid backbone biosynthesis | |
| CICLE_v10002201mg | 8359634 | 8360521 | ||||
| CICLE_v10002259mg | 8409347 | 8410432 | ||||
| α-pinene, citronellal, geranial | 5 | CICLE_v10000921mg | 15445701 | 15450041 | trans-cinnamate 4-monooxygenase | Ubiquinone and other terpenoid–quinone biosynthesis |
| 5 | CICLE_v10000962mg | 16114444 | 16118985 | prenylcysteine oxidase/farnesylcysteine lyase | Terpenoid backbone biosynthesis | |
| geranial | 5 | CICLE_v10000735mg | 25965963 | 25969199 | 4-coumarate--CoA ligase | Ubiquinone and other terpenoid–quinone biosynthesis |
| 5 | CICLE_v10000923mg | 26496115 | 26502834 | phosphomevalonate kinase | Terpenoid backbone biosynthesis (mevalonic pathway) | |
| α-farnesene | 6 | CICLE_v10012198mg | 16230126 | 16231785 | undecaprenyl pyrophosphate synthetase family protein | Terpenoid backbone biosynthesis |
| dodecanal, octanal, decanal, limonene-10-ol = (dodl) | 8 | CICLE_v10028350mg | 4340444 | 4344926 | 4-hydroxy-3-methylbut-2-enyl diphosphate reductase | Terpenoid backbone biosynthesis (non-mevaloic pathway) |
| 8 | CICLE_v10028537mg | 6315013 | 6322458 | farnesyl-diphosphate farnesyltransferase | Sesquiterpenoid and triterpenoid biosynthesis | |
| 8 | CICLE_v10029717mg | 15186679 | 15187280 | NADH-Ubiquinone/plastoquinone (complex I) protein | Ubiquinone and other terpenoid–quinone biosynthesis | |
| (dodl), sabinene, limonene, β-pinene | 8 | CICLE_v10030056mg | 17909793 | 17910005 | terpene synthase 14 | Terpenoid backbone biosynthesis |
| α-sinensal | 9 | CICLE_v10005360mg | 13264152 | 13266718 | γ-tocopherol methyltransferase | Ubiquinone and other terpenoid–quinone biosynthesis |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferrer, V.; Costantino, G.; Paymal, N.; Quinton, C.; Perdomo, E.C.; Paoli, M.; Mournet, P.; Ollitrault, P.; Tomi, F.; Luro, F. Inheritance and Quantitative Trait Loci Mapping of Aromatic Compounds from Clementine (Citrus × clementina Hort. ex Tan.) and Sweet Orange (C. × sinensis (L.) Osb.) Fruit Essential Oils. Genes 2023, 14, 1800. https://doi.org/10.3390/genes14091800
Ferrer V, Costantino G, Paymal N, Quinton C, Perdomo EC, Paoli M, Mournet P, Ollitrault P, Tomi F, Luro F. Inheritance and Quantitative Trait Loci Mapping of Aromatic Compounds from Clementine (Citrus × clementina Hort. ex Tan.) and Sweet Orange (C. × sinensis (L.) Osb.) Fruit Essential Oils. Genes. 2023; 14(9):1800. https://doi.org/10.3390/genes14091800
Chicago/Turabian StyleFerrer, Vincent, Gilles Costantino, Noémie Paymal, Carole Quinton, Estefania Carrillo Perdomo, Mathieu Paoli, Pierre Mournet, Patrick Ollitrault, Félix Tomi, and François Luro. 2023. "Inheritance and Quantitative Trait Loci Mapping of Aromatic Compounds from Clementine (Citrus × clementina Hort. ex Tan.) and Sweet Orange (C. × sinensis (L.) Osb.) Fruit Essential Oils" Genes 14, no. 9: 1800. https://doi.org/10.3390/genes14091800