
Candidate Gene Association Studies in Atopic Dermatitis in Participants of European and Asian Ancestry: A Systematic Review and Meta-Analysis
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
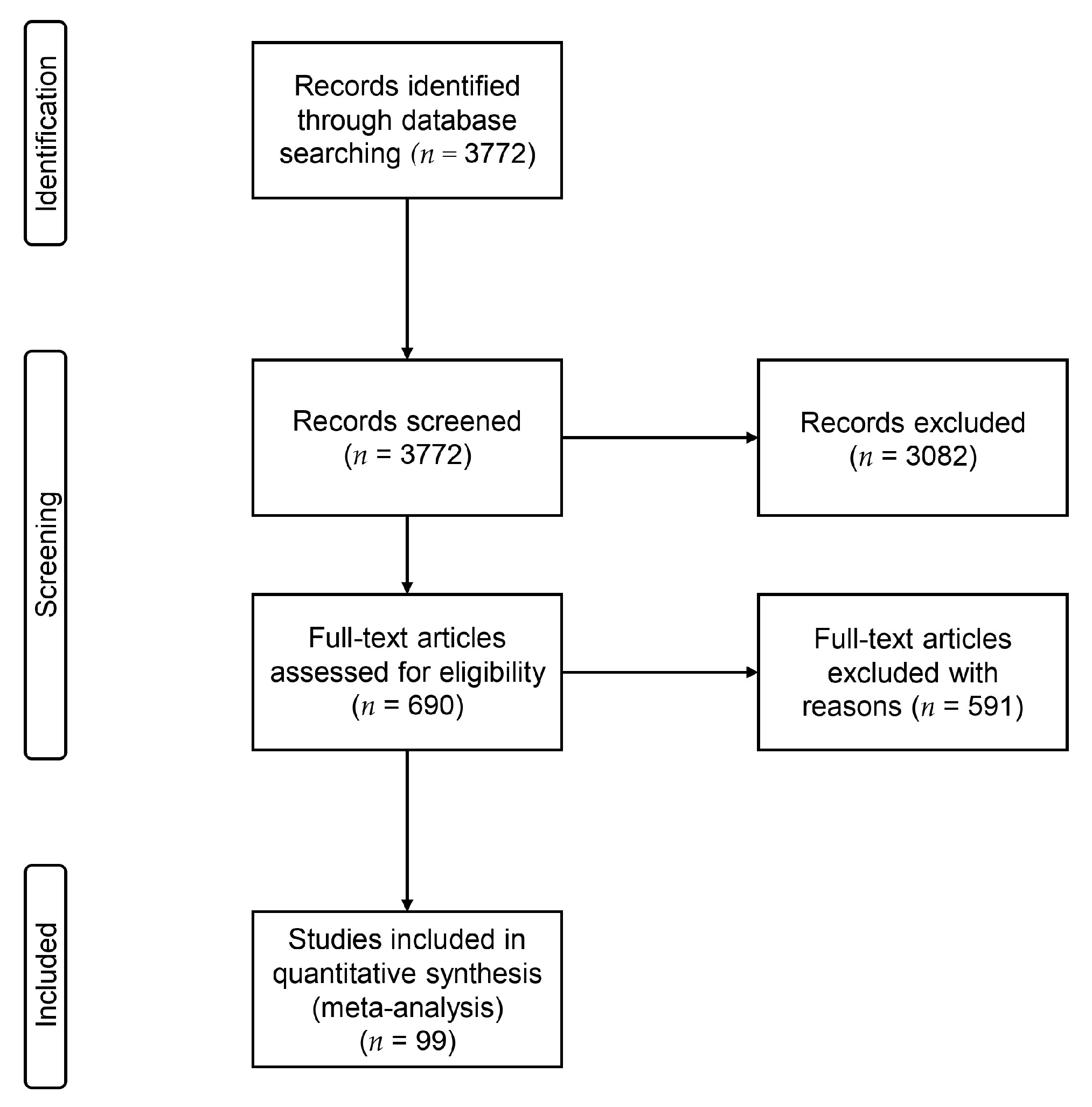
2.1. Search Strategy and Selection Criteria
2.2. Data Extraction
2.3. Statistical Analysis
3. Results
3.1. Studies Included in Our Analysis
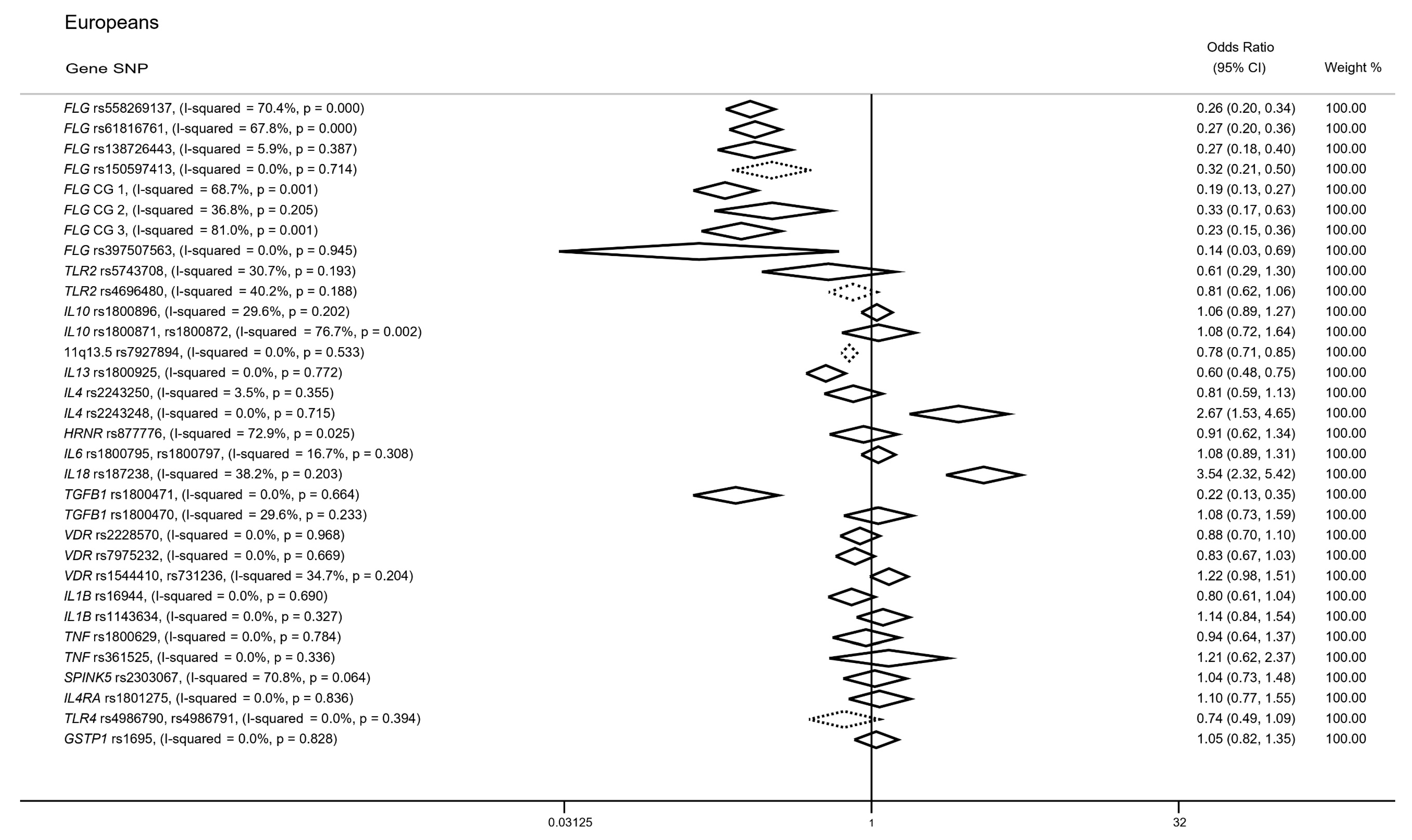
3.2. Candidate Gene Approaches in Cases of European Ancestry
3.2.1. FLG Gene
3.2.2. TLR2 Gene
3.2.3. IL10 Gene
3.2.4. 11. q13.5 Locus
3.2.5. IL13 Gene
3.2.6. SNPs in IL4, IL18 and TGFB1 Genes
3.2.7. SNPs in IL6, HRNR, VDR, IL1B, TNF, SPINK5, IL4RA, TLR4 and GSTP1 Genes
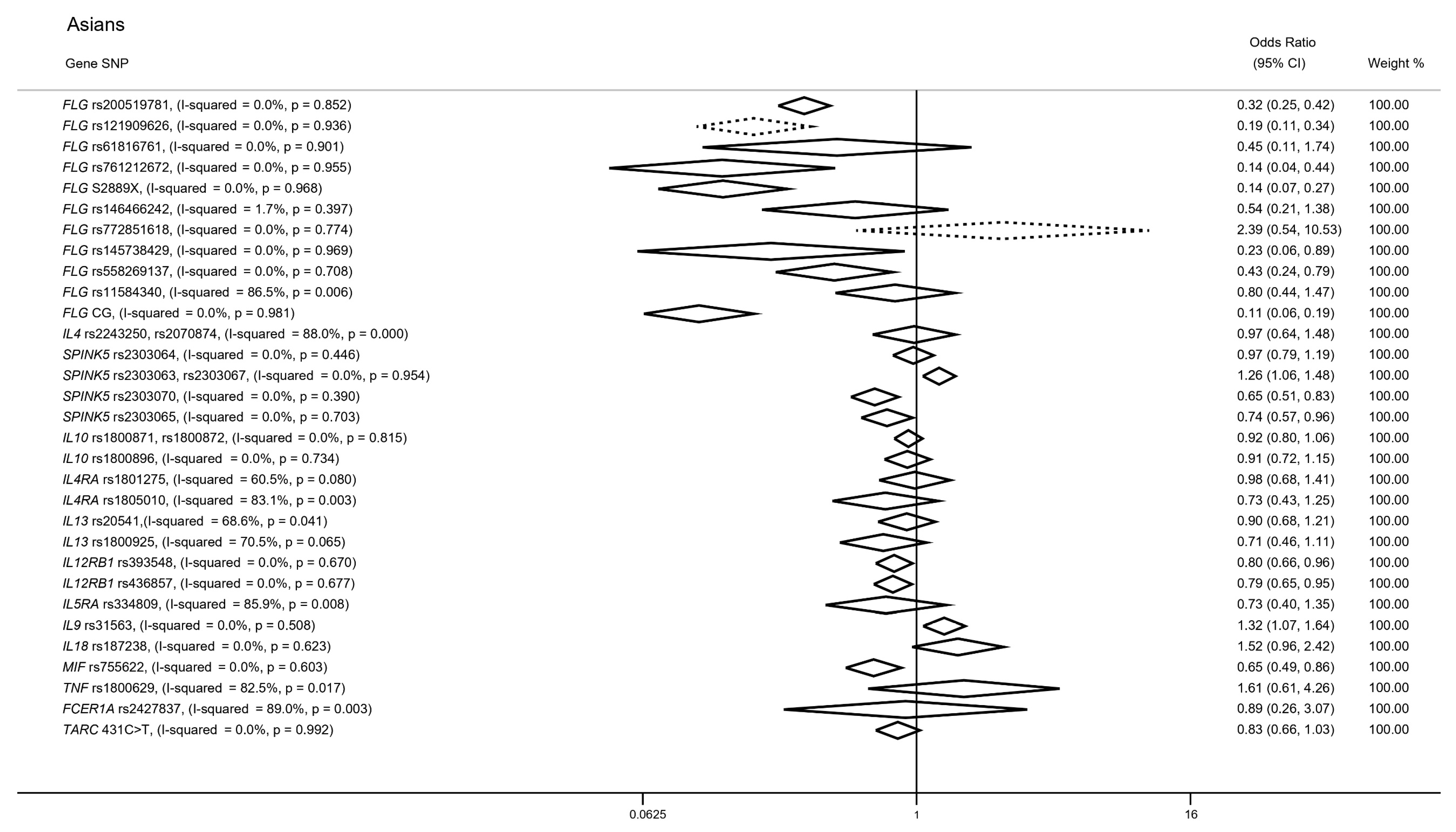
3.3. Candidate Gene Approaches in Cases of Asian Ancestry
3.3.1. FLG Gene
3.3.2. IL4 Gene
3.3.3. SPINK5 Gene
3.3.4. SNPs in IL10, IL4RA and IL13 Genes
3.3.5. SNPs in IL12RB1, IL9 and MIF Genes
3.3.6. SNPs in IL5RA, IL18, TNF, FCER1A and TARC Genes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Langan, S.M.; Irvine, A.D.; Weidinger, S. Atopic Dermatitis. Lancet 2020, 396, 345–360. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, S.F.; Ulrik, C.S.; Kyvik, K.O.; Hjelmborg, J.V.B.; Skadhauge, L.R.; Steffensen, I.; Backer, V. Importance of Genetic Factors in the Etiology of Atopic Dermatitis: A Twin Study. Allergy Asthma Proc. 2007, 28, 535–539. [Google Scholar] [CrossRef] [PubMed]
- Paternoster, L.; Standl, M.; Waage, J.; Baurecht, H.; Hotze, M.; Strachan, D.P.; Curtin, J.A.; Bønnelykke, K.; Tian, C.; Takahashi, A.; et al. Multi-Ancestry Genome-Wide Association Study of 21,000 Cases and 95,000 Controls Identifies New Risk Loci for Atopic Dermatitis. Nat. Genet. 2015, 47, 1449–1456. [Google Scholar] [CrossRef] [PubMed]
- Mucha, S.; Baurecht, H.; Novak, N.; Rodríguez, E.; Bej, S.; Mayr, G.; Emmert, H.; Stölzl, D.; Gerdes, S.; Jung, E.S.; et al. Protein-coding variants contribute to the risk of atopic dermatitis and skin-specific gene expression. J. Allergy Clin. Immunol. 2020, 145, 1208–1218. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, M.A.R.; Vonk, J.M.; Baurecht, H.; Marenholz, I.; Tian, C.; Hoffman, J.D.; Helmer, Q.; Tillander, A.; Ullemar, V.; Lu, Y.; et al. Eleven Loci with New Reproducible Genetic Associations with Allergic Disease Risk. J. Allergy Clin. Immunol. 2019, 143, 691–699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoober, J.K.; Eggink, L.L. The Discovery and Function of Filaggrin. Int. J. Mol. Sci. 2022, 23, 1455. [Google Scholar] [CrossRef]
- Kypriotou, M.; Huber, M.; Hohl, D. The Human Epidermal Differentiation Complex: Cornified Envelope Precursors, S100 Proteins and the “fused Genes” Family. Exp. Dermatol. 2012, 21, 643–649. [Google Scholar] [CrossRef]
- Wynn, T.A. Type 2 Cytokines: Mechanisms and Therapeutic Strategies. Nat. Rev. Immunol. 2015, 15, 271–282. [Google Scholar] [CrossRef]
- Amirzargar, A.A.; Movahedi, M.; Rezaei, N.; Moradi, B.; Dorkhosh, S.; Mahloji, M.; Mahdaviani, S.A. Polymorphisms in IL4 and ILARA Confer Susceptibility to Asthma. J. Investig. Allergol. Clin. Immunol. 2009, 19, 433–438. [Google Scholar]
- Cameron, L.; Webster, R.B.; Strempel, J.M.; Kiesler, P.; Kabesch, M.; Ramachandran, H.; Yu, L.; Stern, D.A.; Graves, P.E.; Lohman, I.C.; et al. Th2 Cell-Selective Enhancement of Human IL13 Transcription by IL13-1112C>T, a Polymorphism Associated with Allergic Inflammation. J. Immunol. 2006, 177, 8633–8642. [Google Scholar] [CrossRef]
- Gooderham, M.J.; Hong, H.C.-H.; Eshtiaghi, P.; Papp, K.A. Dupilumab: A Review of Its Use in the Treatment of Atopic Dermatitis. J. Am. Acad. Dermatol. 2018, 78, S28–S36. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, N.; Koido, M.; Suzuki, A.; Otomo, N.; Suetsugu, H.; Kochi, Y.; Tomizuka, K.; Momozawa, Y.; Kamatani, Y.; Ikegawa, S.; et al. Eight Novel Susceptibility Loci and Putative Causal Variants in Atopic Dermatitis. J. Allergy Clin. Immunol. 2021, 148, 1293–1306. [Google Scholar] [CrossRef]
- Brown, S.J. What Have We Learned from GWAS for Atopic Dermatitis? J. Investig. Dermatol. 2021, 141, 19–22. [Google Scholar] [CrossRef]
- Vardar Acar, N.; Cavkaytar, Ö.; Arik Yilmaz, E.; Büyüktiryaki, A.B.; Uysal Soyer, Ö.; Şahiner, Ü.M.; Şekerel, B.E.; Karaaslan, I.Ç.; Saçkesen, C. Rare Occurrence of Common Filaggrin Mutations in Turkish Children with Food Allergy and Atopic Dermatitis. Turk. J. Med. Sci. 2020, 50, 1865–1871. [Google Scholar] [CrossRef] [PubMed]
- Jurakic Toncic, R.; Kezic, S.; Jakasa, I.; Ljubojevic Hadzavdic, S.; Balic, A.; Petkovic, M.; Pavicic, B.; Zuzul, K.; Marinovic, B. Filaggrin Loss-of-Function Mutations and Levels of Filaggrin Degradation Products in Adult Patients with Atopic Dermatitis in Croatia. J. Eur. Acad. Dermatol. Venereol. 2020, 34, 1789–1794. [Google Scholar] [CrossRef] [PubMed]
- González-Tarancón, R.; Sanmartín, R.; Lorente, F.; Salvador-Rupérez, E.; Hernández-Martín, A.; Rello, L.; Puzo, J.; Gilaberte, Y. Prevalence of FLG Loss-of-Function Mutations R501X, 2282del4, and R2447X in Spanish Children with Atopic Dermatitis. Pediatr. Dermatol. 2020, 37, 98–102. [Google Scholar] [CrossRef] [PubMed]
- Gimalova, G.F.; Karunas, A.S.; Fedorova, Y.Y.; Khusnutdinova, E.K. The Study of Filaggrin Gene Mutations and Copy Number Variation in Atopic Dermatitis Patients from Volga-Ural Region of Russia. Gene 2016, 591, 85–89. [Google Scholar] [CrossRef]
- Woźniak, M.; Kaczmarek-Skamira, E.; Romańska-Gocka, K.; Czajkowski, R.; Kałużna, L.; Zegarska, B. The Prevalence of Mutations in the Gene Encoding Filaggrin in the Population of Polish Patients with Atopic Dermatitis. Postepy Dermatol. Alergol. 2016, 33, 128–133. [Google Scholar] [CrossRef]
- Trzeciak, M.; Gleń, J.; Rębała, K.; Bandurski, T.; Sikorska, M.; Nowicki, R. Coexistence of 2282del4 FLG Gene Mutation and IL-18 -137G/C Gene Polymorphism Enhances the Risk of Atopic Dermatitis. Postepy Dermatol. Alergol. 2016, 33, 57–62. [Google Scholar] [CrossRef] [Green Version]
- Trzeciak, M.; Wesserling, M.; Bandurski, T.; Glen, J.; Nowicki, R.; Pawelczyk, T. Association of a Single Nucleotide Polymorphism in a Late Cornified Envelope-like Proline-Rich 1 Gene (LELP1) with Atopic Dermatitis. Acta Derm. Venereol. 2016, 96, 459–463. [Google Scholar] [CrossRef] [Green Version]
- Komova, E.G.; Shintyapina, A.B.; Makarova, S.I.; Ivanov, M.K.; Chekryga, E.A.; Kaznacheeva, L.F.; Vavilin, V.A. Filaggrin Mutations in a Western Siberian Population and Their Association with Atopic Dermatitis in Children. Genet. Test Mol. Biomark. 2014, 18, 791–796. [Google Scholar] [CrossRef] [PubMed]
- Ballardini, N.; Kull, I.; Söderhäll, C.; Lilja, G.; Wickman, M.; Wahlgren, C.F. Eczema Severity in Preadolescent Children and Its Relation to Sex, Filaggrin Mutations, Asthma, Rhinitis, Aggravating Factors and Topical Treatment: A Report from the BAMSE Birth Cohort. Br. J. Dermatol. 2013, 168, 588–594. [Google Scholar] [CrossRef] [PubMed]
- Ercan, H.; Ispir, T.; Kirac, D.; Baris, S.; Ozen, A.; Oztezcan, S.; Cengizlier, M.R. Predictors of Atopic Dermatitis Phenotypes and Severity: Roles of Serum Immunoglobulins and Filaggrin Gene Mutation R501X. Allergol. Immunopathol. 2013, 41, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Mlitz, V.; Latreille, J.; Gardinier, S.; Jdid, R.; Drouault, Y.; Hufnagl, P.; Eckhart, L.; Guinot, C.; Tschachler, E. Impact of Filaggrin Mutations on Raman Spectra and Biophysical Properties of the Stratum Corneum in Mild to Moderate Atopic Dermatitis. J. Eur. Acad. Dermatol. Venereol. 2012, 26, 983–990. [Google Scholar] [CrossRef]
- O’Regan, G.M.; Campbell, L.E.; Cordell, H.J.; Irvine, A.D.; McLean, W.H.I.; Brown, S.J. Chromosome 11q13.5 Variant Associated with Childhood Eczema: An Effect Supplementary to Filaggrin Mutations. J. Allergy Clin. Immunol. 2010, 125, 170–174.e2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greisenegger, E.; Novak, N.; Maintz, L.; Bieber, T.; Zimprich, F.; Haubenberger, D.; Gleiss, A.; Stingl, G.; Kopp, T.; Zimprich, A. Analysis of Four Prevalent Filaggrin Mutations (R501X, 2282del4, R2447X and S3247X) in Austrian and German Patients with Atopic Dermatitis. J. Eur. Acad. Dermatol. Venereol. 2010, 24, 607–610. [Google Scholar] [CrossRef]
- Gao, P.-S.; Rafaels, N.M.; Hand, T.; Murray, T.; Boguniewicz, M.; Hata, T.; Schneider, L.; Hanifin, J.M.; Gallo, R.L.; Gao, L.; et al. Filaggrin Mutations That Confer Risk of Atopic Dermatitis Confer Greater Risk for Eczema Herpeticum. J. Allergy Clin. Immunol. 2009, 124, 507–513.e7. [Google Scholar] [CrossRef] [Green Version]
- Brown, S.J.; Relton, C.L.; Liao, H.; Zhao, Y.; Sandilands, A.; Wilson, I.J.; Burn, J.; Reynolds, N.J.; McLean, W.H.I.; Cordell, H.J. Filaggrin Null Mutations and Childhood Atopic Eczema: A Population-Based Case-Control Study. J. Allergy Clin. Immunol. 2008, 121, 940–946.e3. [Google Scholar] [CrossRef]
- Brown, S.J.; Sandilands, A.; Zhao, Y.; Liao, H.; Relton, C.L.; Meggitt, S.J.; Trembath, R.C.; Barker, J.N.W.N.; Reynolds, N.J.; Cordell, H.J.; et al. Prevalent and Low-Frequency Null Mutations in the Filaggrin Gene Are Associated with Early-Onset and Persistent Atopic Eczema. J. Investig. Dermatol. 2008, 128, 1591–1594. [Google Scholar] [CrossRef] [Green Version]
- Giardina, E.; Paolillo, N.; Sinibaldi, C.; Novelli, G. R501X and 2282del4 Filaggrin Mutations Do Not Confer Susceptibility to Psoriasis and Atopic Dermatitis in Italian Patients. Dermatology 2008, 216, 83–84. [Google Scholar] [CrossRef]
- Weidinger, S.; O’Sullivan, M.; Illig, T.; Baurecht, H.; Depner, M.; Rodriguez, E.; Ruether, A.; Klopp, N.; Vogelberg, C.; Weiland, S.K.; et al. Filaggrin Mutations, Atopic Eczema, Hay Fever, and Asthma in Children. J. Allergy Clin. Immunol. 2008, 121, 1203–1209.e1. [Google Scholar] [CrossRef] [PubMed]
- Rogers, A.J.; Celedón, J.C.; Lasky-Su, J.A.; Weiss, S.T.; Raby, B.A. Filaggrin Mutations Confer Susceptibility to Atopic Dermatitis but Not to Asthma. J. Allergy Clin. Immunol. 2007, 120, 1332–1337. [Google Scholar] [CrossRef] [PubMed]
- Lerbaek, A.; Bisgaard, H.; Agner, T.; Ohm Kyvik, K.; Palmer, C.N.A.; Menné, T. Filaggrin Null Alleles Are Not Associated with Hand Eczema or Contact Allergy. Br. J. Dermatol. 2007, 157, 1199–1204. [Google Scholar] [CrossRef]
- Sandilands, A.; Terron-Kwiatkowski, A.; Hull, P.R.; O’Regan, G.M.; Clayton, T.H.; Watson, R.M.; Carrick, T.; Evans, A.T.; Liao, H.; Zhao, Y.; et al. Comprehensive Analysis of the Gene Encoding Filaggrin Uncovers Prevalent and Rare Mutations in Ichthyosis Vulgaris and Atopic Eczema. Nat. Genet. 2007, 39, 650–654. [Google Scholar] [CrossRef]
- Weidinger, S.; Rodríguez, E.; Stahl, C.; Wagenpfeil, S.; Klopp, N.; Illig, T.; Novak, N. Filaggrin Mutations Strongly Predispose to Early-Onset and Extrinsic Atopic Dermatitis. J. Investig. Dermatol. 2007, 127, 724–726. [Google Scholar] [CrossRef] [Green Version]
- Marenholz, I.; Nickel, R.; Rüschendorf, F.; Schulz, F.; Esparza-Gordillo, J.; Kerscher, T.; Grüber, C.; Lau, S.; Worm, M.; Keil, T.; et al. Filaggrin Loss-of-Function Mutations Predispose to Phenotypes Involved in the Atopic March. J. Allergy Clin. Immunol. 2006, 118, 866–871. [Google Scholar] [CrossRef] [PubMed]
- Stemmler, S.; Parwez, Q.; Petrasch-Parwez, E.; Epplen, J.T.; Hoffjan, S. Two Common Loss-of-Function Mutations within the Filaggrin Gene Predispose for Early Onset of Atopic Dermatitis. J. Investig. Dermatol. 2007, 127, 722–724. [Google Scholar] [CrossRef]
- Barker, J.N.W.N.; Palmer, C.N.A.; Zhao, Y.; Liao, H.; Hull, P.R.; Lee, S.P.; Allen, M.H.; Meggitt, S.J.; Reynolds, N.J.; Trembath, R.C.; et al. Null Mutations in the Filaggrin Gene (FLG) Determine Major Susceptibility to Early-Onset Atopic Dermatitis That Persists into Adulthood. J. Investig. Dermatol. 2007, 127, 564–567. [Google Scholar] [CrossRef] [Green Version]
- Palmer, C.N.A.; Irvine, A.D.; Terron-Kwiatkowski, A.; Zhao, Y.; Liao, H.; Lee, S.P.; Goudie, D.R.; Sandilands, A.; Campbell, L.E.; Smith, F.J.D.; et al. Common Loss-of-Function Variants of the Epidermal Barrier Protein Filaggrin Are a Major Predisposing Factor for Atopic Dermatitis. Nat. Genet. 2006, 38, 441–446. [Google Scholar] [CrossRef]
- Dêbiñska, A.; Danielewicz, H.; Drabik-Chamerska, A.; Kalita, D.; Boznañski, A. Chromosome 11q13.5 Variant as a Risk Factor for Atopic Dermatitis in Children. Postepy Dermatol. Alergol. 2020, 37, 103–110. [Google Scholar] [CrossRef] [Green Version]
- Can, C.; Yazıcıoğlu, M.; Gürkan, H.; Tozkır, H.; Görgülü, A.; Süt, N.H. Lack of Association between Toll-like Receptor 2 Polymorphisms (R753Q and A-16934T) and Atopic Dermatitis in Children from Thrace Region of Turkey. Balkan Med. J. 2017, 34, 232–238. [Google Scholar] [CrossRef] [PubMed]
- Salpietro, C.; Rigoli, L.; Miraglia Del Giudice, M.; Cuppari, C.; Di Bella, C.; Salpietro, A.; Maiello, N.; La Rosa, M.; Marseglia, G.L.; Leonardi, S.; et al. TLR2 and TLR4 Gene Polymorphisms and Atopic Dermatitis in Italian Children: A Multicenter Study. Int. J. Immunopathol. Pharmacol. 2011, 24, 33–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galli, E.; Ciucci, A.; Cersosimo, S.; Pagnini, C.; Avitabile, S.; Mancino, G.; Delle Fave, G.; Corleto, V.D. Eczema and Food Allergy in an Italian Pediatric Cohort: No Association with TLR-2 and TLR-4 Polymorphisms. Int. J. Immunopathol. Pharmacol. 2010, 23, 671–675. [Google Scholar] [CrossRef] [Green Version]
- Oh, D.-Y.; Schumann, R.R.; Hamann, L.; Neumann, K.; Worm, M.; Heine, G. Association of the Toll-like Receptor 2 A-16934T Promoter Polymorphism with Severe Atopic Dermatitis. Allergy 2009, 64, 1608–1615. [Google Scholar] [CrossRef] [PubMed]
- Niebuhr, M.; Langnickel, J.; Draing, C.; Renz, H.; Kapp, A.; Werfel, T. Dysregulation of Toll-like Receptor-2 (TLR-2)-Induced Effects in Monocytes from Patients with Atopic Dermatitis: Impact of the TLR-2 R753Q Polymorphism. Allergy 2008, 63, 728–734. [Google Scholar] [CrossRef]
- Levchenko, L.Y.; Izmailova, O.V.; Shlykova, O.A.; Kaidashev, I.P. TLR4 896A/G Gene Polymorphism, Rather than the TLR4 1196C/T and TLR2 2258G/A Gene Polymorphisms, Determines the Severe and Aggravated Course of Atopic Dermatitis in Children. Cytol. Genet. 2013, 47, 167–173. [Google Scholar] [CrossRef] [Green Version]
- Ahmad-Nejad, P.; Mrabet-Dahbi, S.; Breuer, K.; Klotz, M.; Werfel, T.; Herz, U.; Heeg, K.; Neumaier, M.; Renz, H. The Toll-like Receptor 2 R753Q Polymorphism Defines a Subgroup of Patients with Atopic Dermatitis Having Severe Phenotype. J. Allergy Clin. Immunol. 2004, 113, 565–567. [Google Scholar] [CrossRef]
- Esposito, S.; Patria, M.F.; Spena, S.; Codecà, C.; Tagliabue, C.; Zampiero, A.; Lelii, M.; Montinaro, V.; Pelucchi, C.; Principi, N. Impact of Genetic Polymorphisms on Paediatric Atopic Dermatitis. Int. J. Immunopathol. Pharmacol. 2015, 28, 286–295. [Google Scholar] [CrossRef] [Green Version]
- Lesiak, A.; Zakrzewski, M.; Przybyłowska, K.; Rogowski-Tylman, M.; Wozniacka, A.; Narbutt, J. Atopic Dermatitis Patients Carrying G Allele in -1082 G/A IL-10 Polymorphism Are Predisposed to Higher Serum Concentration of IL-10. Arch. Med. Sci. 2014, 10, 1239–1243. [Google Scholar] [CrossRef] [Green Version]
- Kayserova, J.; Sismova, K.; Zentsova-Jaresova, I.; Katina, S.; Vernerova, E.; Polouckova, A.; Capkova, S.; Malinova, V.; Striz, I.; Sediva, A. A Prospective Study in Children with a Severe Form of Atopic Dermatitis: Clinical Outcome in Relation to Cytokine Gene Polymorphisms. J. Investig. Allergol. Clin. Immunol. 2012, 22, 92–101. [Google Scholar]
- Stavric, K.; Peova, S.; Trajkov, D.; Spiroski, M. Gene Polymorphisms of 22 Cytokines in Macedonian Children with Atopic Dermatitis. Iran. J. Allergy Asthma Immunol. 2012, 11, 37–50. [Google Scholar] [PubMed]
- Lesiak, A.; Kuna, P.; Zakrzewski, M.; van Geel, M.; Bladergroen, R.S.; Przybylowska, K.; Stelmach, I.; Majak, P.; Hawro, T.; Sysa-Jedrzejowska, A.; et al. Combined Occurrence of Filaggrin Mutations and IL-10 or IL-13 Polymorphisms Predisposes to Atopic Dermatitis. Exp. Dermatol. 2011, 20, 491–495. [Google Scholar] [CrossRef] [PubMed]
- Reich, K.; Westphal, G.; König, I.R.; Mössner, R.; Schupp, P.; Gutgesell, C.; Hallier, E.; Ziegler, A.; Neumann, C. Cytokine Gene Polymorphisms in Atopic Dermatitis. Br. J. Dermatol. 2003, 148, 1237–1241. [Google Scholar] [CrossRef]
- Arkwright, P.D.; Chase, J.M.; Babbage, S.; Pravica, V.; David, T.J.; Hutchinson, I.V. Atopic Dermatitis Is Associated with a Low-Producer Transforming Growth Factor β1 Cytokine Genotype. J. Allergy Clin. Immunol. 2001, 108, 281–284. [Google Scholar] [CrossRef] [PubMed]
- Ponińska, J.K.; Samoliński, B.; Tomaszewska, A.; Raciborski, F.; Samel-Kowalik, P.; Walkiewicz, A.; Lipiec, A.; Piekarska, B.; Krzych-Fałta, E.; Namysłowski, A.; et al. Haplotype Dependent Association of Rs7927894 (11q13.5) with Atopic Dermatitis and Chronic Allergic Rhinitis: A Study in ECAP Cohort. PLoS ONE 2017, 12, e0183922. [Google Scholar] [CrossRef] [Green Version]
- Greisenegger, E.K.; Zimprich, F.; Zimprich, A.; Gleiss, A.; Kopp, T. Association of the Chromosome 11q13.5 Variant with Atopic Dermatitis in Austrian Patients. Eur. J. Dermatol. 2013, 23, 142–145. [Google Scholar] [CrossRef]
- Marenholz, I.; Bauerfeind, A.; Esparza-Gordillo, J.; Kerscher, T.; Granell, R.; Nickel, R.; Lau, S.; Henderson, J.; Lee, Y.-A. The Eczema Risk Variant on Chromosome 11q13 (Rs7927894) in the Population-Based ALSPAC Cohort: A Novel Susceptibility Factor for Asthma and Hay Fever. Hum. Mol. Genet. 2011, 20, 2443–2449. [Google Scholar] [CrossRef] [Green Version]
- Hummelshoj, T.; Bodtger, U.; Datta, P.; Malling, H.J.; Oturai, A.; Poulsen, L.K.; Ryder, L.P.; Sorensen, P.S.; Svejgaard, E.; Svejgaard, A. Association between an Interleukin-13 Promoter Polymorphism and Atopy. Eur. J. Immunogenet. 2003, 30, 355–359. [Google Scholar] [CrossRef]
- Dębińska, A.; Danielewicz, H.; Sozańska, B. Genetic Variants in Epidermal Differentiation Complex Genes as Predictive Biomarkers for Atopic Eczema, Allergic Sensitization, and Eczema-Associated Asthma in a 6-Year Follow-Up Case-Control Study in Children. J. Clin. Med. 2022, 11, 4865. [Google Scholar] [CrossRef]
- Trzeciak, M.; Gleń, J.; Roszkiewicz, J.; Nedoszytko, B. Association of Single Nucleotide Polymorphism of Interleukin-18 with Atopic Dermatitis. J. Eur. Acad. Dermatol. Venereol. 2010, 24, 78–79. [Google Scholar] [CrossRef]
- Kılıç, S.; Sılan, F.; Hız, M.M.; Işık, S.; Ögretmen, Z.; Özdemir, Ö. Vitamin D Receptor Gene BSMI, FOKI, APAI, and TAQI Polymorphisms and the Risk of Atopic Dermatitis. J. Investig. Allergol. Clin. Immunol. 2016, 26, 106–110. [Google Scholar] [CrossRef] [PubMed]
- Heine, G.; Hoefer, N.; Franke, A.; Nöthling, U.; Schumann, R.R.; Hamann, L.; Worm, M. Association of Vitamin D Receptor Gene Polymorphisms with Severe Atopic Dermatitis in Adults. Br. J. Dermatol. 2013, 168, 855–858. [Google Scholar] [CrossRef]
- Dežman, K.; Korošec, P.; Rupnik, H.; Rijavec, M. SPINK5 Is Associated with Early-Onset and CHI3L1 with Late-Onset Atopic Dermatitis. Int. J. Immunogenet. 2017, 44, 212–218. [Google Scholar] [CrossRef] [PubMed]
- Fölster-Holst, R.; Stoll, M.; Koch, W.A.; Hampe, J.; Christophers, E.; Schreiber, S. Lack of Association of SPINK5 Polymorphisms with Nonsyndromic Atopic Dermatitis in the Population of Northern Germany. Br. J. Dermatol. 2005, 152, 1365–1367. [Google Scholar] [CrossRef] [PubMed]
- Vavilin, V.A.; Safronova, O.G.; Lyapunova, A.A.; Lyakhovich, V.V.; Kaznacheeva, L.F.; Manankin, N.A.; Molokova, A.V. Interaction of GSTM1, GSTT1, and GSTP1 Genotypes in Determination of Predisposition to Atopic Dermatitis. Bull. Exp. Biol. Med. 2003, 136, 388–391. [Google Scholar] [CrossRef]
- Safronova, O.G.; Vavilin, V.A.; Lyapunova, A.A.; Makarova, S.I.; Lyakhovich, V.V.; Kaznacheeva, L.F.; Manankin, N.A.; Batychko, O.A.; Gavalov, S.M. Relationship between Glutathione S-Transferase P1 Polymorphism and Bronchial Asthma and Atopic Dermatitis. Bull. Exp. Biol. Med. 2003, 136, 73–75. [Google Scholar] [CrossRef]
- Kim, B.J.; Wang, H.-Y.; Lee, H.; Lee, S.-Y.; Hong, S.-J.; Choi, E.H. Clinical Characteristics and Genetic Variations in Early-Onset Atopic Dermatitis Patients. Ann. Dermatol. 2019, 31, 286–293. [Google Scholar] [CrossRef]
- Sasaki, T.; Furusyo, N.; Shiohama, A.; Takeuchi, S.; Nakahara, T.; Uchi, H.; Hirota, T.; Tamari, M.; Shimizu, N.; Ebihara, T.; et al. Filaggrin Loss-of-Function Mutations Are Not a Predisposing Factor for Atopic Dermatitis in an Ishigaki Island under Subtropical Climate. J. Dermatol. Sci. 2014, 76, 10–15. [Google Scholar] [CrossRef]
- Meng, L.; Wang, L.; Tang, H.; Tang, X.; Jiang, X.; Zhao, J.; Gao, J.; Li, B.; Fu, X.; Chen, Y.; et al. Filaggrin Gene Mutation c.3321delA Is Associated with Various Clinical Features of Atopic Dermatitis in the Chinese Han Population. PLoS ONE 2014, 9, e98235. [Google Scholar] [CrossRef]
- Lee, D.-E.; Park, S.-Y.; Han, J.-Y.; Ryu, H.-M.; Lee, H.-C.; Han, Y.S. Association between Filaggrin Mutations and Atopic Dermatitis in Korean Pregnant Women. Int. J. Dermatol. 2013, 52, 772–773. [Google Scholar] [CrossRef]
- Chen, H.; Common, J.E.A.; Haines, R.L.; Balakrishnan, A.; Brown, S.J.; Goh, C.S.M.; Cordell, H.J.; Sandilands, A.; Campbell, L.E.; Kroboth, K.; et al. Wide Spectrum of Filaggrin-Null Mutations in Atopic Dermatitis Highlights Differences between Singaporean Chinese and European Populations. Br. J. Dermatol. 2011, 165, 106–114. [Google Scholar] [CrossRef]
- Zhang, H.; Guo, Y.; Wang, W.; Shi, M.; Chen, X.; Yao, Z. Mutations in the Filaggrin Gene in Han Chinese Patients with Atopic Dermatitis. Allergy 2011, 66, 420–427. [Google Scholar] [CrossRef] [PubMed]
- Osawa, R.; Konno, S.; Akiyama, M.; Nemoto-Hasebe, I.; Nomura, T.; Nomura, Y.; Abe, R.; Sandilands, A.; McLean, W.H.I.; Hizawa, N.; et al. Japanese-Specific Filaggrin Gene Mutations in Japanese Patients Suffering from Atopic Eczema and Asthma. J. Investig. Dermatol. 2010, 130, 2834–2836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nomura, Y.; Akiyama, M.; Nomura, T.; Nemoto-Hasebe, I.; Abe, R.; McLean, W.H.I.; Shimizu, H. Chromosome 11q13.5 Variant: No Association with Atopic Eczema in the Japanese Population. J. Dermatol. Sci. 2010, 59, 210–212. [Google Scholar] [CrossRef]
- Ma, L.; Zhang, L.; Di, Z.-H.; Zhao, L.-P.; Lu, Y.-N.; Xu, J.; Chen, H.-D.; Gao, X.-H. Association Analysis of Filaggrin Gene Mutations and Atopic Dermatitis in Northern China. Br. J. Dermatol. 2010, 162, 225–227. [Google Scholar] [CrossRef]
- Nemoto-Hasebe, I.; Akiyama, M.; Nomura, T.; Sandilands, A.; McLean, W.H.I.; Shimizu, H. FLG Mutation p.Lys4021X in the C-terminal Imperfect Filaggrin Repeat in Japanese Patients with Atopic Eczema. Br. J. Dermatol. 2009, 161, 1387–1390. [Google Scholar] [CrossRef] [PubMed]
- Nomura, T.; Akiyama, M.; Sandilands, A.; Nemoto-Hasebe, I.; Sakai, K.; Nagasaki, A.; Palmer, C.N.A.; Smith, F.J.D.; McLean, W.H.I.; Shimizu, H. Prevalent and Rare Mutations in the Gene Encoding Filaggrin in Japanese Patients with Ichthyosis Vulgaris and Atopic Dermatitis. J. Investig. Dermatol. 2009, 129, 1302–1305. [Google Scholar] [CrossRef] [Green Version]
- Nomura, T.; Akiyama, M.; Sandilands, A.; Nemoto-Hasebe, I.; Sakai, K.; Nagasaki, A.; Ota, M.; Hata, H.; Evans, A.T.; Palmer, C.N.A.; et al. Specific Filaggrin Mutations Cause Ichthyosis Vulgaris and Are Significantly Associated with Atopic Dermatitis in Japan. J. Investig. Dermatol. 2008, 128, 1436–1441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enomoto, H.; Hirata, K.; Otsuka, K.; Kawai, T.; Takahashi, T.; Hirota, T.; Suzuki, Y.; Tamari, M.; Otsuka, F.; Fujieda, S.; et al. Filaggrin Null Mutations Are Associated with Atopic Dermatitis and Elevated Levels of IgE in the Japanese Population: A Family and Case-Control Study. J. Hum. Genet. 2008, 53, 615. [Google Scholar] [CrossRef] [Green Version]
- Nomura, T.; Sandilands, A.; Akiyama, M.; Liao, H.; Evans, A.T.; Sakai, K.; Ota, M.; Sugiura, H.; Yamamoto, K.; Sato, H.; et al. Unique Mutations in the Filaggrin Gene in Japanese Patients with Ichthyosis Vulgaris and Atopic Dermatitis. J. Allergy Clin. Immunol. 2007, 119, 434–440. [Google Scholar] [CrossRef]
- Ching, G.K.; Hon, K.-L.; Ng, P.-C.; Leung, T.-F. Filaggrin Null Mutations in Childhood Atopic Dermatitis among the Chinese. Int. J. Immunogenet. 2009, 36, 251–254. [Google Scholar] [CrossRef] [PubMed]
- Zhong, W.-L.; Wu, X.; Yu, B.; Zhang, J.; Zhang, W.; Xu, N.; Zhou, J.; Zheng, J.-C.; Chen, X.-F.; Dou, X. Filaggrin Gene Mutation c.3321delA Is Associated with Dry Phenotypes of Atopic Dermatitis in the Chinese Han Population. Chin. Med. J. 2016, 129, 1498–1500. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-Y.; Yang, S.W.; Kim, H.-L.; Kim, S.-H.; Kim, S.J.; Park, S.-M.; Son, M.; Ryu, S.; Pyo, Y.-S.; Lee, J.-S.; et al. Association between P478S Polymorphism of the Filaggrin Gene & Atopic Dermatitis. Indian J. Med. Res. 2013, 138, 922–927. [Google Scholar]
- Wang, I.J.; Lin, T.J.; Kuo, C.F.; Lin, S.L.; Lee, Y.L.; Chen, P.C. Filaggrin Polymorphism P478S, IgE Level, and Atopic Phenotypes. Br. J. Dermatol. 2011, 164, 791–796. [Google Scholar] [CrossRef]
- Hua, L.; Chen, Q.; Liu, Q.-H.; Guo, Y.-F.; Cheng, R.-H.; Zhang, J.; Zhang, J.-H.; Wang, L.-W.; Ji, R.-X. Interaction between Antibiotic Use and MS4A2 Gene Polymorphism on Childhood Eczema: A Prospective Birth Cohort Study. BMC Pediatr. 2021, 21, 314. [Google Scholar] [CrossRef]
- Shang, H.; Cao, X.-L.; Wan, Y.-J.; Meng, J.; Guo, L.-H. IL-4 Gene Polymorphism May Contribute to an Increased Risk of Atopic Dermatitis in Children. Dis. Markers 2016, 2016, 1021942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussein, Y.M.; Alzahrani, S.S.; Alharthi, A.A.; Alhazmi, A.S.; Ghonaim, M.M.; Alghamdy, A.A.N.; El Askary, A. Gene Polymorphism of Interleukin-4, Interleukin-4 Receptor and STAT6 in Children with Atopic Dermatitis in Taif, Saudi Arabia. Immunol. Investig. 2016, 45, 223–234. [Google Scholar] [CrossRef]
- Gharagozlou, M.; Behniafard, N.; Amirzargar, A.A.; Hosseinverdi, S.; Sotoudeh, S.; Farhadi, E.; Khaledi, M.; Aryan, Z.; Moghaddam, Z.G.; Mahmoudi, M.; et al. Association between Single Nucleotide Polymorphisms of the Interleukin-4 Gene and Atopic Dermatitis. Acta Dermatovenerol. Croat. 2015, 23, 96–100. [Google Scholar]
- Hussein, Y.M.; Shalaby, S.M.; Nassar, A.; Alzahrani, S.S.; Alharbi, A.S.; Nouh, M. Association between Genes Encoding Components of the IL-4/IL-4 Receptor Pathway and Dermatitis in Children. Gene 2014, 545, 276–281. [Google Scholar] [CrossRef]
- Tanaka, K.; Sugiura, H.; Uehara, M.; Hashimoto, Y.; Donnelly, C.; Montgomery, D.S. Lack of Association between Atopic Eczema and the Genetic Variants of Interleukin-4 and the Interleukin-4 Receptor α Chain Gene: Heterogeneity of Genetic Backgrounds on Immunoglobulin E Production in Atopic Eczema Patients. Clin. Exp. Allergy 2001, 31, 1522–1527. [Google Scholar] [CrossRef]
- Kawashima, T.; Noguchi, E.; Arinami, T.; Yamakawa-Kobayashi, K.; Nakagawa, H.; Otsuka, F.; Hamaguchi, H. Linkage and Association of an Interleukin 4 Gene Polymorphism with Atopic Dermatitis in Japanese Families. J. Med. Genet. 1998, 35, 502–504. [Google Scholar] [CrossRef] [Green Version]
- Morizane, S.; Ouchida, M.; Sunagawa, K.; Sugimoto, S.; Kobashi, M.; Sugihara, S.; Nomura, H.; Tsuji, K.; Sato, A.; Miura, Y.; et al. Analysis of All 34 Exons of the SPINK5 Gene in Japanese Atopic Dermatitis Patients. Acta Med. Okayama 2018, 72, 275–282. [Google Scholar] [CrossRef]
- Zhao, L.P.; Di, Z.; Zhang, L.; Wang, L.; Ma, L.; Lv, Y.; Hong, Y.; Wei, H.; Chen, H.D.; Gao, X.H. Association of SPINK5 Gene Polymorphisms with Atopic Dermatitis in Northeast China. J. Eur. Acad. Dermatol. Venereol. 2012, 26, 572–577. [Google Scholar] [CrossRef] [PubMed]
- Kato, A.; Fukai, K.; Oiso, N.; Hosomi, N.; Murakami, T.; Ishii, M. Association of SPINK5 Gene Polymorphisms with Atopic Dermatitis in the Japanese Population. Br. J. Dermatol. 2003, 148, 665–669. [Google Scholar] [CrossRef] [PubMed]
- Bin Huraib, G.; Al Harthi, F.; Arfin, M.; Al-Sugheyr, M.; Rizvi, S.; Al-Asmari, A. Cytokine Gene Polymorphisms in Saudi Patients with Atopic Dermatitis: A Case-Control Study. Biomark Insights 2018, 13, 1177271918777760. [Google Scholar] [CrossRef] [PubMed]
- Behniafard, N.; Amirzargar, A.A.; Gharagozlou, M.; Delavari, F.; Hosseinverdi, S.; Sotoudeh, S.; Farhadi, E.; Mahmoudi, M.; Khaledi, M.; Moghaddam, Z.G.; et al. Single Nucleotide Polymorphisms of the Genes Encoding IL-10 and TGF-Β1 in Iranian Children with Atopic Dermatitis. Allergol. Immunopathol. 2018, 46, 155–159. [Google Scholar] [CrossRef] [PubMed]
- Sohn, M.H.; Song, J.S.; Kim, K.-W.; Kim, E.-S.; Kim, K.-E.; Lee, J.M. Association of Interleukin-10 Gene Promoter Polymorphism in Children with Atopic Dermatitis. J. Pediatr. 2007, 150, 106–108. [Google Scholar] [CrossRef]
- Miyake, Y.; Tanaka, K.; Arakawa, M. Case-Control Study of Eczema in Relation to IL4Rα Genetic Polymorphisms in Japanese Women: The Kyushu Okinawa Maternal and Child Health Study. Scand. J. Immunol. 2013, 77, 413–418. [Google Scholar] [CrossRef]
- Miyake, Y.; Tanaka, K.; Arakawa, M. IL13 Genetic Polymorphisms, Smoking, and Eczema in Women: A Case-Control Study in Japan. BMC Med. Genet. 2011, 12, 142. [Google Scholar] [CrossRef] [Green Version]
- Miyake, Y.; Kiyohara, C.; Koyanagi, M.; Fujimoto, T.; Shirasawa, S.; Tanaka, K.; Sasaki, S.; Hirota, Y. Case-Control Study of Eczema Associated with IL13 Genetic Polymorphisms in Japanese Children. Int. Arch. Allergy Immunol. 2011, 154, 328–335. [Google Scholar] [CrossRef]
- Takahashi, N.; Akahoshi, M.; Matsuda, A.; Ebe, K.; Inomata, N.; Obara, K.; Hirota, T.; Nakashima, K.; Shimizu, M.; Tamari, M.; et al. Association of the IL12RB1 Promoter Polymorphisms with Increased Risk of Atopic Dermatitis and Other Allergic Phenotypes. Hum. Mol. Genet. 2005, 14, 3149–3159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Namkung, J.-H.; Lee, J.-E.; Kim, E.; Park, G.T.; Yang, H.S.; Jang, H.Y.; Shin, E.-S.; Cho, E.-Y.; Yang, J.-M. An Association between IL-9 and IL-9 Receptor Gene Polymorphisms and Atopic Dermatitis in a Korean Population. J. Dermatol. Sci. 2011, 62, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Choi, J.; Hahn, H.-J.; Lee, Y.-B.; Yu, D.-S.; Kim, J.-W. Association of Macrophage Migration Inhibitory Factor Polymorphisms with Total Plasma IgE Levels in Patients with Atopic Dermatitis in Korea. PLoS ONE 2016, 11, e0162477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, L.; Xue, H.-B.; Guan, X.-H.; Qi, R.-Q.; Liu, Y.-B. Macrophage Migration Inhibitory Factor Promoter 173G/C Polymorphism Is Associated with Atopic Dermatitis Risk. Int. J. Dermatol. 2014, 53, e75–e77. [Google Scholar] [CrossRef]
- Miyake, Y.; Tanaka, K.; Arakawa, M. IL5RA Polymorphisms, Smoking and Eczema in Japanese Women: The Kyushu Okinawa Maternal and Child Health Study. Int. J. Immunogenet. 2015, 42, 52–57. [Google Scholar] [CrossRef] [PubMed]
- Kato, T.; Tsunemi, Y.; Saeki, H.; Shibata, S.; Sekiya, T.; Nakamura, K.; Kakinuma, T.; Kagami, S.; Fujita, H.; Tada, Y.; et al. Interferon-18 Gene Polymorphism -137 G/C Is Associated with Susceptibility to Psoriasis Vulgaris but Not with Atopic Dermatitis in Japanese Patients. J. Dermatol. Sci. 2009, 53, 162–163. [Google Scholar] [CrossRef] [PubMed]
- Osawa, K.; Etoh, T.; Ariyoshi, N.; Ishii, I.; Ohtani, M.; Kariya, S.; Uchino, K.; Kitada, M. Relationship between Kaposi’s Varicelliform Eruption in Japanese Patients with Atopic Dermatitis Treated with Tacrolimus Ointment and Genetic Polymorphisms in the IL-18 Gene Promoter Region. J. Dermatol. 2007, 34, 531–536. [Google Scholar] [CrossRef]
- Behniafard, N.; Gharagozlou, M.; Farhadi, E.; Khaledi, M.; Sotoudeh, S.; Darabi, B.; Fathi, S.M.; Gholizadeh Moghaddam, Z.; Mahmoudi, M.; Aghamohammadi, A.; et al. TNF-alpha Single Nucleotide Polymorphisms in Atopic Dermatitis. Eur. Cytokine Netw. 2012, 23, 163–165. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Zhou, Y.; Lin, L.; Wang, J.; Peng, X.; Li, J.; Li, L. Association of Polymorphisms in the Promoter Region of FCER1A Gene with Atopic Dermatitis, Chronic Uticaria, Asthma, and Serum Immunoglobulin E Levels in a Han Chinese Population. Hum. Immunol. 2012, 73, 301–305. [Google Scholar] [CrossRef]
- Park, K.Y.; Park, M.K.; Kim, E.J.; Lee, M.-K.; Seo, S.J. FCεRI Gene Promoter Polymorphisms and Total IgE Levels in Susceptibility to Atopic Dermatitis in Korea. J. Korean Med. Sci. 2011, 26, 870–874. [Google Scholar] [CrossRef]
- Tsunemi, Y.; Komine, M.; Sekiya, T.; Saeki, H.; Nakamura, K.; Hirai, K.; Kakinuma, T.; Kagami, S.; Fujita, H.; Asano, N.; et al. The -431C>T Polymorphism of Thymus and Activation-Regulated Chemokine Increases the Promoter Activity but Is Not Associated with Susceptibility to Atopic Dermatitis in Japanese Patients. Exp. Dermatol. 2004, 13, 715–719. [Google Scholar] [CrossRef] [PubMed]
- Sekiya, T.; Tsunemi, Y.; Miyamasu, M.; Ohta, K.; Morita, A.; Saeki, H.; Matsushima, K.; Yoshie, O.; Tsuchiya, N.; Yamaguchi, M.; et al. Variations in the Human Th2-Specific Chemokine TARC Gene. Immunogenetics 2003, 54, 742–745. [Google Scholar] [CrossRef] [PubMed]
- Grafanaki, K.; Antonatos, C.; Maniatis, A.; Petropoulou, A.; Vryzaki, E.; Vasilopoulos, Y.; Georgiou, S.; Gregoriou, S. Intrinsic Effects of Exposome in Atopic Dermatitis: Genomics, Epigenomics and Regulatory Layers. J. Clin. Med. 2023, 12, 4000. [Google Scholar] [CrossRef] [PubMed]
- McAleer, M.A.; Irvine, A.D. The Multifunctional Role of Filaggrin in Allergic Skin Disease. J. Allergy Clin. Immunol. 2013, 131, 280–291. [Google Scholar] [CrossRef]
- Rodríguez, E.; Baurecht, H.; Herberich, E.; Wagenpfeil, S.; Brown, S.J.; Cordell, H.J.; Irvine, A.D.; Weidinger, S. Meta-Analysis of Filaggrin Polymorphisms in Eczema and Asthma: Robust Risk Factors in Atopic Disease. J. Allergy Clin. Immunol. 2009, 123, 1361–1370.e7. [Google Scholar] [CrossRef]
- Nedoszytko, B.; Olszewska, B.; Roszkiewicz, J.; Glen, J.; Zabłotna, M.; Ługowska-Umer, H.; Nowicki, R.; Sokołowska-Wojdyło, M. The Role of Polymorphism of Interleukin-2, -10, -13 and TNF-α Genes in Cutaneous T-Cell Lymphoma Pathogenesis. Postepy Dermatol. Alergol. 2016, 33, 429–434. [Google Scholar] [CrossRef]
- Park, A.; Wong, L.; Lang, A.; Kraus, C.; Anderson, N.; Elsensohn, A. Cutaneous T-Cell Lymphoma Following Dupilumab Use: A Systematic Review. Int. J. Dermatol. 2023, 62, 862–876. [Google Scholar] [CrossRef]
- Imboden, M.; Nieters, A.; Bircher, A.J.; Brutsche, M.; Becker, N.; Wjst, M.; Ackermann-Liebrich, U.; Berger, W.; Probst-Hensch, N.M. Cytokine Gene Polymorphisms and Atopic Disease in Two European Cohorts. (ECRHS-Basel and SAPALDIA). Clin. Mol. Allergy 2006, 4, 9. [Google Scholar] [CrossRef] [Green Version]
- Weissler, K.A.; Frischmeyer-Guerrerio, P.A. Genetic Evidence for the Role of Transforming Growth Factor-β in Atopic Phenotypes. Curr. Opin. Immunol. 2019, 60, 54–62. [Google Scholar] [CrossRef]
- Bozza, M.T.; Lintomen, L.; Kitoko, J.Z.; Paiva, C.N.; Olsen, P.C. The Role of MIF on Eosinophil Biology and Eosinophilic Inflammation. Clin. Rev. Allergy Immunol. 2020, 58, 15–24. [Google Scholar] [CrossRef]
- Egger, M.; Smith, G.D. Bias in Location and Selection of Studies. BMJ 1998, 316, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Lou, C.; Mitra, N.; Wubbenhorst, B.; D’Andrea, K.; Hoffstad, O.; Kim, B.S.; Yan, A.; Zaenglein, A.L.; Fuxench, Z.C.; Nathanson, K.L.; et al. Association between Fine Mapping Thymic Stromal Lymphopoietin and Atopic Dermatitis Onset and Persistence. Ann. Allergy Asthma Immunol. 2019, 123, 595–601.e1. [Google Scholar] [CrossRef] [PubMed]
- Gupta, J.; Margolis, D.J. Filaggrin Gene Mutations with Special Reference to Atopic Dermatitis. Curr. Treat. Options Allergy 2020, 7, 403–413. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Study, Year | Rs ID | Sample Size |
|---|---|---|
| Vardar Acar N et al., 2020 [14] | rs558269137, rs61816761, rs138726443, rs150597413 | 189 |
| Jurakic Toncic R et al., 2020 [15] | rs558269137, rs61816761, rs138726443 | 150 |
| González-Tarancón R et al., 2020 [16] | rs558269137, rs61816761, rs138726443, CG2 | 214 |
| Gimalova GF et al., 2016 [17] | rs558269137, rs61816761 | 564 |
| Woźniak M et al., 2016 [18] | rs558269137, rs61816761, CG1 | 121 |
| Trzeciak M et al., 2016 [19] | rs558269137, rs61816761, rs1800925, rs187238 | 275 |
| Trzeciak M et al., 2016 [20] | rs558269137 | 256 |
| Komova EG et al., 2014 [21] | rs558269137, rs61816761, rs138726443, rs150597413 | 230 |
| Ballardini N et al., 2013 [22] | rs558269137, rs61816761, rs138726443, CG2 | 1854 |
| Ercan H et al., 2013 [23] | rs61816761 | 99 |
| Mlitz V et al., 2012 [24] | rs558269137, rs61816761, rs138726443, CG2 | 196 |
| O’Regan GM et al., 2010 [25] | rs558269137, rs61816761, rs138726443, rs150597413, CG3, rs7927894, rs877776 | 1511 |
| Greisenegger E et al., 2010 [26] | rs558269137, rs61816761, rs138726443, rs150597413, CG3 | 864 |
| Gao PS et al., 2009 [27] | rs558269137, rs61816761, CG1 | 435 |
| Brown SJ et al., 2008 [28] | rs558269137, rs61816761, rs138726443, rs150597413, rs397507563 | 811 |
| Brown SJ et al., 2008 [29] | rs558269137, rs61816761, rs138726443, rs150597413, rs397507563 | 1221 |
| Giardina E et al., 2008 [30] | rs558269137, rs61816761 | 388 |
| Weidinger S et al., 2008 [31] | rs558269137, rs61816761, rs138726443, rs150597413, CG3 | 3099 |
| Rogers AJ et al., 2007 [32] | rs558269137, rs61816761, CG1 | 646 |
| Lerbaek A et al., 2007 [33] | rs558269137, rs61816761, CG1 | 215 |
| Sandilands A et al., 2007 [34] | rs558269137, rs61816761, rs138726443, rs150597413, rs397507563 | 924 |
| Weidinger S et al., 2007 [35] | rs558269137, rs61816761, CG1 | 526 |
| Marenholz I et al., 2006 [36] | rs558269137, rs61816761, CG1 | 507 |
| Stemmler S et al., 2007 [37] | rs558269137, rs61816761, CG1 | 1078 |
| Barker JN et al., 2007 [38] | rs558269137, rs61816761, CG1 | 1626 |
| Palmer CN et al., 2006 [39] | rs558269137, rs61816761, CG1 | 241 |
| Dêbiñska A et al., 2020 [40] | CG3, rs7927894 | 188 |
| Can C et al., 2017 [41] | rs5743708, rs4696480 | 139 |
| Salpietro C et al., 2011 [42] | rs5743708, rs4696480, rs4986790, rs4986791 | 337 |
| Galli E et al., 2010 [43] | rs5743708 | 249 |
| Oh DY et al., 2009 [44] | rs5743708, rs4696480, rs4986790, rs4986791 | 265 |
| Niebuhr M et al., 2008 [45] | rs5743708 | 19 |
| Levchenko L Yu et al., 2013 [46] | rs5743708 | 131 |
| Ahmad-Nejad et al., 2003 [47] | rs5743708 | 117 |
| Esposito S et al., 2015 [48] | rs1800896, rs1800872 | 223 |
| Lesiak A et al., 2014 [49] | rs1800896, rs1800925, rs2243250 | 136 |
| Kayserova J et al., 2012 [50] | rs1800896, rs1800871, rs1800872, rs2243250, rs2243248, rs1800795, rs1800797, rs1801275 | 197 |
| Stavric K et al., 2012 [51] | rs1800896, rs1800871, rs1800872, rs2243250, rs2243248, rs1800471, rs1800470, rs1800795, rs1800797, rs16944, rs1143634, rs1800629, rs361525, rs1801275 | 367 |
| Lesiak A et al., 2011 [52] | rs1800896, rs1800925 | 367 |
| Reich K et al., 2003 [53] | rs1800896, rs1800795, rs16944, rs1143634, rs1800629, rs361525 | 308 |
| Arkwright PD et al., 2001 [54] | rs1800896, rs1800471, rs1800470 | 118 |
| Ponińska JK et al., 2017 [55] | rs7927894 | 810 |
| Greisenegger EK et al., 2013 [56] | rs7927894, rs877776 | 518 |
| Marenholz I et al., 2011 [57] | rs7927894 | 2485 |
| Hummelshoj T et al., 2003 [58] | rs1800925 | 159 |
| Dêbiñska A et al., 2022 [59] | rs877776 | 188 |
| Trzeciak M et al., 2010 [60] | rs187238 | 113 |
| Kılıç S et al., 2016 [61] | rs2228570, rs7975232, rs1544410, rs731236 | 138 |
| Heine G et al., 2013 [62] | rs2228570, rs7975232, rs1544410, rs731236 | 530 |
| Dežman K et al., 2017 [63] | rs2303067 | 405 |
| Fölster-Holst R et al., 2005 [64] | rs2303067 | 569 |
| Vavilin VA et al., 2003 [65] | rs1695 | 325 |
| Safronova OG et al., 2003 [66] | rs1695 | 274 |
| Study, Year | Rs ID | Sample Size |
|---|---|---|
| Kim BJ et al., 2019 [67] | rs200519781, rs146466242, rs2303064, rs2303070, rs2303065, rs393548, rs436857, rs31563, rs334809 | 325 |
| Sasaki T et al., 2014 [68] | rs200519781, rs121909626, rs761212672, rs145738429, rs61816761, rs146466242, rs772851618 | 721 |
| Meng L et al., 2014 [69] | rs200519781 | 1988 |
| Lee DE et al., 2013 [70] | rs200519781 | 175 |
| Chen H et al., 2011 [71] | rs200519781, rs61816761 | 865 |
| Zhang H et al., 2011 [72] | rs200519781 | 353 |
| Osawa R et al., 2010 [73] | rs200519781, rs121909626, rs761212672, rs145738429, S2889X, rs61816761, rs146466242, rs772851618, CG | 306 |
| Nomura Y et al., 2010 [74] | rs200519781, rs121909626, rs761212672, rs145738429, S2889X, rs61816761, rs146466242, rs772851618, CG | 307 |
| Ma L et al., 2010 [75] | rs200519781 | 329 |
| Nemoto-Hasebe I et al., 2009 [76] | rs200519781, rs121909626, rs761212672, rs145738429, S2889X, rs61816761, rs146466242, rs772851618, CG | 271 |
| Nomura T et al., 2009 [77] | rs200519781, rs121909626, rs761212672, rs145738429, S2889X, rs61816761, rs772851618 | 252 |
| Nomura T et al., 2008 [78] | rs200519781, rs121909626, rs761212672, S2889X | 235 |
| Enomoto H et al., 2008 [79] | rs200519781, rs121909626 | 1299 |
| Nomura T et al., 2007 [80] | rs200519781, rs121909626 | 299 |
| Ching GK et al., 2009 [81] | rs121909626, S2889X, rs558269137, rs61816761 | 365 |
| Zhong WL et al., 2016 [82] | rs558269137 | 1017 |
| Kim SY et al., 2013 [83] | rs11584340 | 527 |
| Wang IJ et al., 2011 [84] | rs11584340 | 328 |
| Hua L et al., 2021 [85] | rs2243250, rs2070874, rs1801275, rs1805010, rs20541 | 597 |
| Shang H et al., 2016 [86] | rs2243250, rs2070874 | 182 |
| Hussein YM et al., 2016 [87] | rs2243250, rs2070874, rs1805010 | 100 |
| Gharagozlou M et al., 2015 [88] | rs2243250, rs2070874, rs1801275 | 228 |
| Hussein YM et al., 2014 [89] | rs2243250, rs2070874 | 206 |
| Tanaka K et al., 2001 [90] | rs2243250, rs2070874, rs1805010 | 424 |
| Kawashima T et al., 1998 [91] | rs2243250, rs2070874 | 425 |
| Morizane S et al., 2018 [92] | rs2303064, rs2303063, rs2303067, rs2303070 | 107 |
| Zhao LP et al., 2012 [93] | rs2303064, rs2303063, rs2303067, rs2303070 | 341 |
| Kato A et al., 2003 [94] | rs2303064, rs2303063, rs2303067, rs2303065 | 234 |
| Bin Huraib G et al., 2018 [95] | rs1800871, rs1800872, rs1800896, rs1800629 | 315 |
| Behniafard N et al., 2018 [96] | rs1800871, rs1800872, rs1800896 | 229 |
| Sohn MH et al., 2007 [97] | rs1800871, rs1800872, rs1800896 | 416 |
| Miyake Y et al., 2013 [98] | rs1801275 | 823 |
| Miyake Y et al., 2011 [99] | rs20541, rs1800925 | 1270 |
| Miyake Y et al., 2011 [100] | rs20541, rs1800925 | 533 |
| Takahashi N et al., 2005 [101] | rs393548, rs436857 | 1040 |
| Namkung JH et al., 2011 [102] | rs31563 | 1090 |
| Kim JS et al., 2016 [103] | rs755622 | 258 |
| Ma L et al., 2013 [104] | rs755622 | 391 |
| Miyake Y et al., 2015 [105] | rs334809 | 1318 |
| Kato T et al., 2009 [106] | rs187238 | 264 |
| Osawa K et al., 2007 [107] | rs187238 | 121 |
| Behniafard N et al., 2012 [108] | rs1800629 | 226 |
| Zhou J et al., 2012 [109] | rs2427837 | 380 |
| Park KY et al., 2011 [110] | rs2427837 | 231 |
| Tsunemi Y et al., 2004 [111] | 431C > T | 351 |
| Sekiya T et al., 2003 [112] | 431C > T | 306 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pontikas, A.; Antonatos, C.; Evangelou, E.; Vasilopoulos, Y. Candidate Gene Association Studies in Atopic Dermatitis in Participants of European and Asian Ancestry: A Systematic Review and Meta-Analysis. Genes 2023, 14, 1456. https://doi.org/10.3390/genes14071456
Pontikas A, Antonatos C, Evangelou E, Vasilopoulos Y. Candidate Gene Association Studies in Atopic Dermatitis in Participants of European and Asian Ancestry: A Systematic Review and Meta-Analysis. Genes. 2023; 14(7):1456. https://doi.org/10.3390/genes14071456
Chicago/Turabian StylePontikas, Alexandros, Charalabos Antonatos, Evangelos Evangelou, and Yiannis Vasilopoulos. 2023. "Candidate Gene Association Studies in Atopic Dermatitis in Participants of European and Asian Ancestry: A Systematic Review and Meta-Analysis" Genes 14, no. 7: 1456. https://doi.org/10.3390/genes14071456