
The Relationship between TLR3 rs3775291 Polymorphism and Infectious Diseases: A Meta-Analysis of Case-Control Studies

 ,
,  ,
,
Abstract
:1. Introduction
2. Material and Methods
2.1. Study Design
2.2. Search Strategy
2.3. Data Extraction
2.4. Statistical Analysis
3. Results
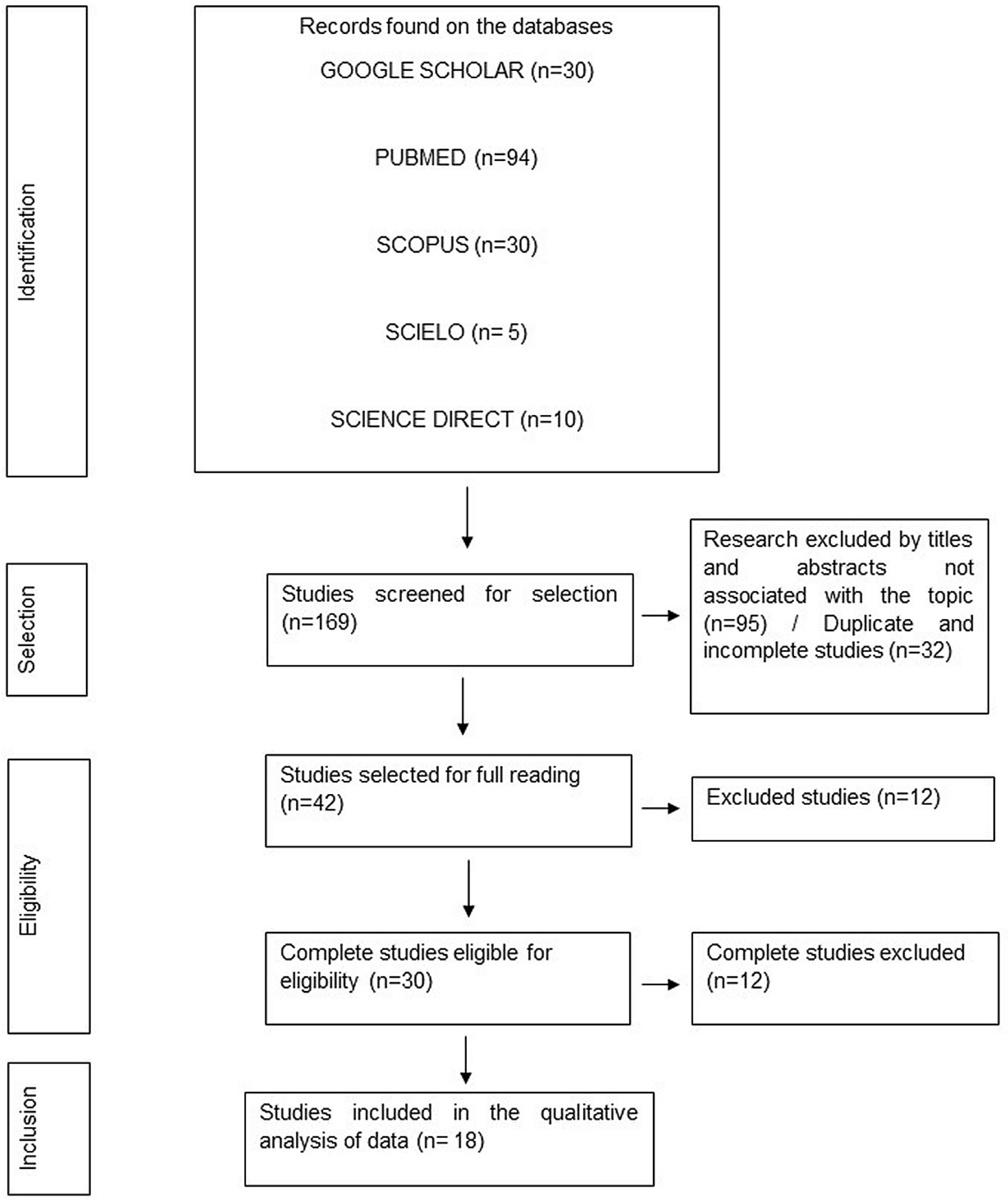
3.1. Literature Search
3.2. Characteristics of the Included Studies
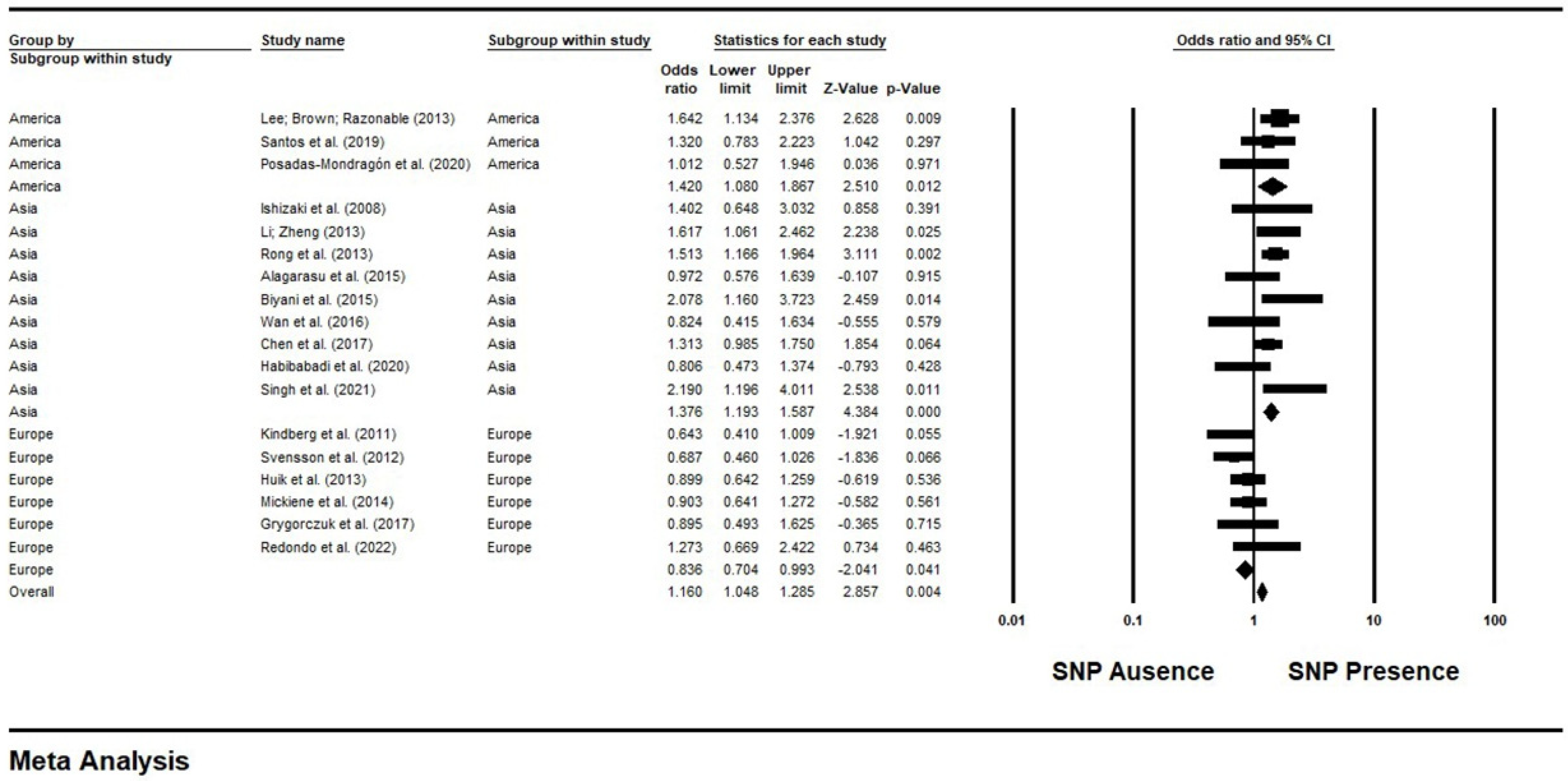
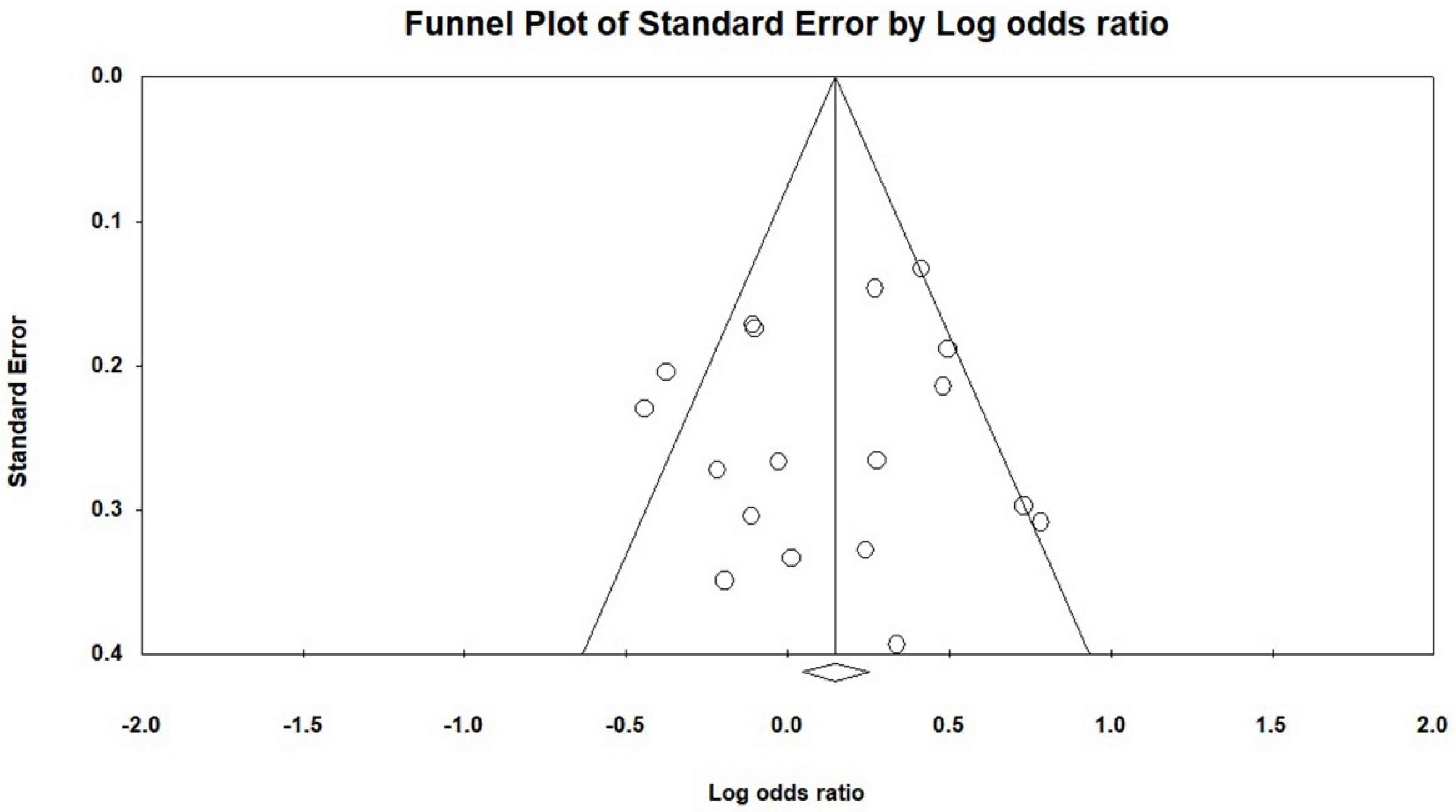
3.3. Results of a Meta-Analysis and Publication Bias between the TLR3 rs3775291 Polymorphism and the Risk of Infections in Subgroups of Continents
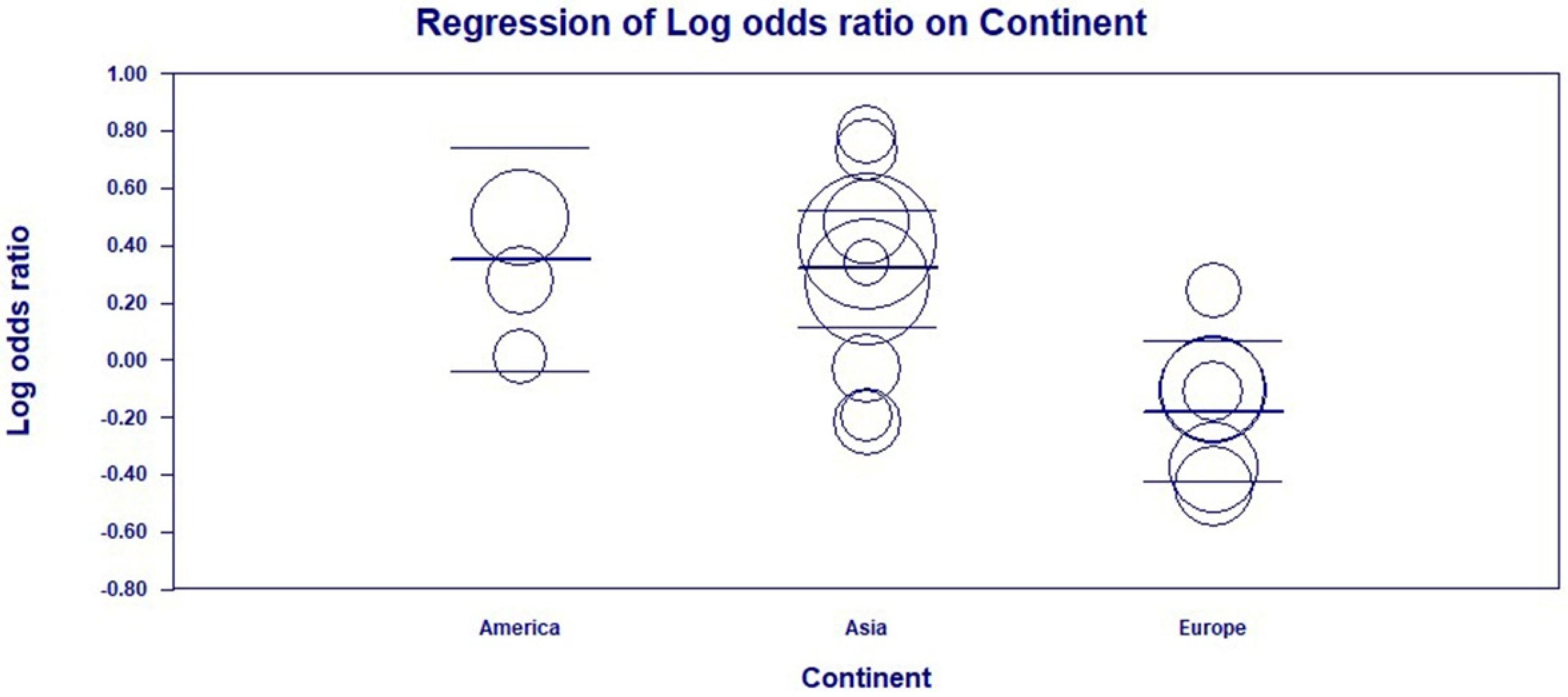
3.4. Subgroup and Univariate Meta-Regression Analyses
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Borges, G.M. Health transition in Brazil: Regional variations and divergence/convergence in mortality. Cad. Saúde Pública 2017, 33, e00080316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization (Ed.) The World Health Report 2000: Health Systems: Improving Performance; World Health Organization: Geneva, Switzerland, 2000; ISBN 92-4-156198-X. [Google Scholar]
- Gadelha, M.R.; Kasuki, L.; Korbonits, M. The genetic background of acromegaly. Pituitary 2017, 20, 10–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez, D.; Lefebvre, C.; Rioux, J.; García, L.F.; Barrera, L.F. Evaluation of Toll-like receptor and adaptor molecule polymorphisms for susceptibility to tuberculosis in a Colombian population. Int. J. Immunogenet. 2012, 39, 216–223. [Google Scholar] [CrossRef]
- Rai, R.C. Host inflammatory responses to intracellular invaders: Review study. Life Sci. 2020, 240, 117084. [Google Scholar] [CrossRef]
- Teräsjärvi, J.; Hakanen, A.; Korppi, M.; Nuolivirta, K.; Gröndahl-Yli-Hannuksela, K.; Mertsola, J.; Peltola, V.; He, Q. Rapid detection of functional gene polymorphisms of TLRs and IL-17 using high resolution melting analysis. Sci. Rep. 2017, 7, 41522. [Google Scholar] [CrossRef] [Green Version]
- Alseoudy, M.M.; Elgamal, M.; Abdelghany, D.A.; Borg, A.M.; El-Mesery, A.; Elzeiny, D.; Hammad, M.O. Prognostic impact of toll-like receptors gene polymorphism on outcome of COVID-19 pneumonia: A case-control study. Clin. Immunol. 2022, 235, 108929. [Google Scholar] [CrossRef] [PubMed]
- Moresco, E.M.Y.; LaVine, D.; Beutler, B. Toll-like receptors. Curr. Biol. 2011, 21, R488–R493. [Google Scholar] [CrossRef] [Green Version]
- Ribet, D.; Cossart, P. How bacterial pathogens colonize their hosts and invade deeper tissues. Microbes Infect. 2015, 17, 173–183. [Google Scholar] [CrossRef]
- Castro, F.A.; Försti, A.; Buch, S.; Kalthoff, H.; Krauss, C.; Bauer, M.; Egberts, J.; Schniewind, B.; Broering, D.C.; Schreiber, S.; et al. TLR-3 polymorphism is an independent prognostic marker for stage II colorectal cancer. Eur. J. Cancer 2011, 47, 1203–1210. [Google Scholar] [CrossRef]
- Chen, Y.; Lin, J.; Zhao, Y.; Ma, X.; Yi, H. Toll-like receptor 3 (TLR3) regulation mechanisms and roles in antiviral innate immune responses. J. Zhejiang Univ. Sci. B 2021, 22, 609–632. [Google Scholar] [CrossRef]
- Caetano, A.R. SNP markers: Basic concepts, applications in animal breeding and management and perspectives for the future. R. Bras. Zootec. 2009, 38, 64–71. [Google Scholar] [CrossRef] [Green Version]
- Habibabadi, H.M.; Parsania, M.; Pourfathollah, A.A.; Haghighat, S.; Sharifi, Z. Association of TLR3 single nucleotide polymorphisms with susceptibility to HTLV-1 infection in Iranian asymptomatic blood donors. Rev. Soc. Bras. Med. Trop. 2020, 53, e20200026. [Google Scholar] [CrossRef]
- Redondo, N.; Rodríguez-Goncer, I.; Parra, P.; Ruiz-Merlo, T.; López-Medrano, F.; González, E.; Polanco, N.; Trujillo, H.; Hernández, A.; San Juan, R.; et al. Influence of single-nucleotide polymorphisms in TLR3 (rs3775291) and TLR9 (rs352139) on the risk of CMV infection in kidney transplant recipients. Front. Immunol. 2022, 13, 929995. [Google Scholar] [CrossRef] [PubMed]
- Stroup, D.F. Meta-analysis of Observational Studies in EpidemiologyA Proposal for Reporting. JAMA 2000, 283, 2008. [Google Scholar] [CrossRef] [PubMed]
- Tugwell, P.; Tovey, D. PRISMA 2020. J. Clin. Epidemiol. 2021, 134, A5–A6. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. BMJ 2021, n71. [Google Scholar] [CrossRef] [PubMed]
- Tarsilla, M. Cochrane handbook for systematic reviews of interventions. J. Multidiscip. Eval. 2010, 6, 142–148. [Google Scholar] [CrossRef]
- Wang, B.-G.; Yi, D.-H.; Liu, Y.-F. TLR3 gene polymorphisms in cancer: A systematic review and meta-analysis. Chin. J. Cancer 2015, 34, 19. [Google Scholar] [CrossRef] [Green Version]
- Tang, J.-L.; Liu, J.L. Misleading funnel plot for detection of bias in meta-analysis. J. Clin. Epidemiol. 2000, 53, 477–484. [Google Scholar] [CrossRef]
- Khanmohammadi, S.; Rezaei, N. Role of Toll-like receptors in the pathogenesis of COVID-19. J. Med. Virol. 2021, 93, 2735–2739. [Google Scholar] [CrossRef]
- Kawai, T.; Akira, S. The role of pattern-recognition receptors in innate immunity: Update on Toll-like receptors. Nat. Immunol. 2010, 11, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Lester, S.N.; Li, K. Toll-Like Receptors in Antiviral Innate Immunity. J. Mol. Biol. 2014, 426, 1246–1264. [Google Scholar] [CrossRef] [PubMed]
- Ishizaki, Y.; Takemoto, M.; Kira, R.; Kusuhara, K.; Torisu, H.; Sakai, Y.; Sanefuji, M.; Yukaya, N.; Hara, T. Association of toll-like receptor 3 gene polymorphism with subacute sclerosing panencephalitis. J. Neurovirol. 2008, 14, 486–491. [Google Scholar] [CrossRef] [PubMed]
- Ranjith-Kumar, C.T.; Miller, W.; Sun, J.; Xiong, J.; Santos, J.; Yarbrough, I.; Lamb, R.J.; Mills, J.; Duffy, K.E.; Hoose, S.; et al. Effects of single nucleotide polymorphisms on Toll-like receptor 3 activity and expression in cultured cells. J. Biol. Chem. 2007, 282, 17696–17705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, D.; Wang, X.D.; Jin, G.Y. Research progress of Toll-like receptor and ligand complex structure. China J. Cell. Mol. Immunol. 2015, 31, 553–556. [Google Scholar]
- Qi, R.; Hoose, S.; Schreiter, J.; Sawant, K.V.; Lamb, R.; Ranjith-Kumar, C.T.; Mills, J.; San Mateo, L.; Jordan, J.L.; Kao, C.C. Secretion of the human Toll-like receptor 3 ectodomain is affected by single nucleotide polymorphisms and regulated by Unc93b1. J. Biol. Chem. 2010, 285, 36635–36644. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.; Shu, H.-B. Deciphering the pathways to antiviral innate immunity and inflammation. Adv. Immunol. 2020, 145, 1–36. [Google Scholar]
- Matsumoto, M.; Oshiumi, H.; Seya, T. Antiviral responses induced by the TLR3 pathway. Rev. Med. Virol. 2011, 21, 67–77. [Google Scholar] [CrossRef]
- Redondo, N.; Rodríguez-Goncer, I.; Parra, P.; López-Medrano, F.; González, E.; Hernández, A.; Trujillo, H.; Ruiz-Merlo, T.; San Juan, R.; Folgueira, M.D.; et al. Genetic polymorphisms in TLR3, IL10 and CD209 influence the risk of BK polyomavirus infection after kidney transplantation. Sci. Rep. 2022, 12, 11338. [Google Scholar] [CrossRef]
- Lee, S.-O.; Brown, R.A.; Razonable, R.R. Association between a functional polymorphism in Toll-like receptor 3 and chronic hepatitis C in liver transplant recipients. Transpl. Infect. Dis. 2013, 15, 111–119. [Google Scholar] [CrossRef]
- Santos, C.N.O.; Ribeiro, D.R.; Cardoso Alves, J.; Cazzaniga, R.A.; Magalhães, L.S.; de Souza, M.S.F.; Fonseca, A.B.L.; Bispo, A.J.B.; Porto, R.L.S.; dos Santos, C.A.; et al. Association Between Zika Virus Microcephaly in Newborns With the rs3775291 Variant in Toll-Like Receptor 3 and rs1799964 Variant at Tumor Necrosis Factor-α Gene. J. Infect. Dis. 2019, 220, 1797–1801. [Google Scholar] [CrossRef] [PubMed]
- Posadas-Mondragón, A.; Aguilar-Faisal, J.L.; Zuñiga, G.; Magaña, J.J.; Santiago-Cruz, J.A.; Guillén-Salomón, E.; Alcántara-Farfán, V.; Arellano-Flores, M.L.; Salas-Benito, J.S.; Neri-Bazán, R.M.; et al. Association of Genetic Polymorphisms in TLR3, TLR4, TLR7, and TLR8 with the Clinical Forms of Dengue in Patients from Veracruz, Mexico. Viruses 2020, 12, 1230. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Zheng, Z. Toll-like receptor 3 genetic variants and susceptibility to hepatocellular carcinoma and HBV-related hepatocellular carcinoma. Tumour Biol. 2013, 34, 1589–1594. [Google Scholar] [CrossRef] [PubMed]
- Rong, Y.; Song, H.; You, S.; Zhu, B.; Zang, H.; Zhao, Y.; Li, Y.; Wan, Z.; Liu, H.; Zhang, A.; et al. Association of Toll-like receptor 3 polymorphisms with chronic hepatitis B and hepatitis B-related acute-on-chronic liver failure. Inflammation 2013, 36, 413–418. [Google Scholar] [CrossRef] [PubMed]
- Alagarasu, K.; Bachal, R.V.; Memane, R.S.; Shah, P.S.; Cecilia, D. Polymorphisms in RNA sensing toll like receptor genes and its association with clinical outcomes of dengue virus infection. Immunobiology 2015, 220, 164–168. [Google Scholar] [CrossRef]
- Biyani, S.; Garg, R.K.; Jain, A.; Malhotra, H.S.; Kumar, R.; Prakash, S.; Verma, R.; Sharma, P.K. Toll-like receptor-3 gene polymorphism in patients with Japanese encephalitis. J. Neuroimmunol. 2015, 286, 71–76. [Google Scholar] [CrossRef]
- Wan, Z.; Lin, X.; Li, T.; Zhou, A.; Yang, M.; Hu, D.; Feng, L.; Peng, S.; Fan, L.; Tu, S.; et al. Genetic variant in CXCL13 gene is associated with susceptibility to intrauterine infection of hepatitis B virus. Sci. Rep. 2016, 6, 26465. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Xie, W.; Lu, Y.; Su, S.; Nong, L.; Jia, Y.; Liu, Y.; Zhou, W.; Wang, H.; Tan, A. Gene polymorphisms of TLR2 and TLR3 in HBV clearance and HBV-related hepatocellular carcinoma in a Chinese male population. Int. J. Biol. Mrk. 2017, 32, e195–e201. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.K.; Prakash, S.; Garg, R.K.; Jain, P.; Kumar, R.; Jain, A. Study of Single Nucleotide Polymorphisms in Endosomal Toll-Like Receptors-3, 7, and 9 Genes in Patients With Dengue: A Case-Control Study. Cureus 2021, 13, e14883. [Google Scholar] [CrossRef]
- Préhaud, C.; Mégret, F.; Lafage, M.; Lafon, M. Virus Infection Switches TLR-3-Positive Human Neurons To Become Strong Producers of Beta Interferon. J. Virol. 2005, 79, 12893–12904. [Google Scholar] [CrossRef] [Green Version]
- Huik, K.; Avi, R.; Pauskar, M.; Kallas, E.; Jõgeda, E.-L.; Karki, T.; Marsh, K.; Des Jarlais, D.; Uusküla, A.; Lutsar, I. Association between TLR3 rs3775291 and resistance to HIV among highly exposed Caucasian intravenous drug users. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2013, 20, 78–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mickienė, A.; Pakalnienė, J.; Nordgren, J.; Carlsson, B.; Hagbom, M.; Svensson, L.; Lindquist, L. Polymorphisms in chemokine receptor 5 and Toll-like receptor 3 genes are risk factors for clinical tick-borne encephalitis in the Lithuanian population. PLoS ONE 2014, 9, e106798. [Google Scholar] [CrossRef]
- Svensson, A.; Tunbäck, P.; Nordström, I.; Padyukov, L.; Eriksson, K. Polymorphisms in Toll-like receptor 3 confer natural resistance to human herpes simplex virus type 2 infection. J. Gen. Virol. 2012, 93, 1717–1724. [Google Scholar] [CrossRef] [PubMed]
- Reinert, L.S.; Harder, L.; Holm, C.K.; Iversen, M.B.; Horan, K.A.; Dagnæs-Hansen, F.; Ulhøi, B.P.; Holm, T.H.; Mogensen, T.H.; Owens, T.; et al. TLR3 deficiency renders astrocytes permissive to herpes simplex virus infection and facilitates establishment of CNS infection in mice. J. Clin. Investig. 2012, 122, 1368–1376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.; Audry, M.; Ciancanelli, M.; Alsina, L.; Azevedo, J.; Herman, M.; Anguiano, E.; Sancho-Shimizu, V.; Lorenzo, L.; Pauwels, E.; et al. Herpes simplex virus encephalitis in a patient with complete TLR3 deficiency: TLR3 is otherwise redundant in protective immunity. J. Exp. Med. 2011, 208, 2083–2098. [Google Scholar] [CrossRef]
- Wu, J.; Lu, M.; Meng, Z.; Trippler, M.; Broering, R.; Szczeponek, A.; Krux, F.; Dittmer, U.; Roggendorf, M.; Gerken, G.; et al. Toll-like receptor-mediated control of HBV replication by nonparenchymal liver cells in mice. Hepatology 2007, 46, 1769–1778. [Google Scholar] [CrossRef]
- Horscroft, N.J.; Pryde, D.C.; Bright, H. Antiviral applications of Toll-like receptor agonists. J. Antimicrob. Chemother. 2012, 67, 789–801. [Google Scholar] [CrossRef] [Green Version]
- Levi, J.E. Emerging Infectious Agents and Blood Safety in Latin America. Front. Med. 2018, 5, 71. [Google Scholar] [CrossRef] [Green Version]
- Tanabe, M.; Kurita-Taniguchi, M.; Takeuchi, K.; Takeda, M.; Ayata, M.; Ogura, H.; Matsumoto, M.; Seya, T. Mechanism of up-regulation of human Toll-like receptor 3 secondary to infection of measles virus-attenuated strains. Biochem. Biophys. Res. Commun. 2003, 311, 39–48. [Google Scholar] [CrossRef]
- Dhiman, N.; Ovsyannikova, I.G.; Vierkant, R.A.; Ryan, J.E.; Pankratz, V.S.; Jacobson, R.M.; Poland, G.A. Associations Between SNPs in Toll-like Receptors and Related Intracellular Signaling Molecules and Immune Responses to Measles Vaccine: Preliminary Results. Vaccine 2008, 26, 1731–1736. [Google Scholar] [CrossRef] [Green Version]
- Fenizia, C.; Fiocchi, M.; Jones, K.; Parks, R.W.; Ceribelli, M.; Chevalier, S.A.; Edwards, D.; Ruscetti, F.; Pise-Masison, C.A.; Franchini, G. Human T-Cell Leukemia/Lymphoma Virus Type 1 p30, but Not p12/p8, Counteracts Toll-Like Receptor 3 (TLR3) and TLR4 Signaling in Human Monocytes and Dendritic Cells. J. Virol. 2014, 88, 393–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doetschman, T. Influence of Genetic Background on Genetically Engineered Mouse Phenotypes. Methods Mol. Biol. 2009, 530, 423–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhangadamajhi, G.; Rout, R. Association of TLR3 functional variant (rs3775291) with COVID-19 susceptibility and death: A population-scale study. Hum. Cell 2021, 34, 1025–1027. [Google Scholar] [CrossRef] [PubMed]
- Pati, A.; Padhi, S.; Chaudhury, S.; Panda, A.K. TLR3 (rs3775291) variant is not associated with SARS-CoV-2 infection and related mortality: A population-based correlation analysis. Hum. Cell 2021, 34, 1274–1277. [Google Scholar] [CrossRef]
- Pessoa, N.L.; Bentes, A.A.; de Carvalho, A.L.; de Souza Silva, T.B.; Alves, P.A.; de Sousa Reis, E.V.; Rodrigues, T.A.; Kroon, E.G.; Campos, M.A. Case report: Hepatitis in a child infected with SARS-CoV-2 presenting toll-like receptor 7 Gln11Leu single nucleotide polymorphism. Virol. J. 2021, 18, 1–5. [Google Scholar] [CrossRef]
- Shi, H.; He, H.; Sun, C.; Fu, J.; Ghosh, D.; Deng, C.; Sheng, Y. Association of toll-like receptor polymorphisms with acquisition of HIV infection and clinical findings. Medicine 2020, 99, e23663. [Google Scholar] [CrossRef]
- Ye, S.; Zhang, X.; Zhang, Y.; Tian, X.; Liu, A.; Cui, C.; Shi, L.; Xia, D. Association of TLR3 (rs3775291) and IL-10 (rs1800871) gene polymorphisms with susceptibility to Hepatitis B infection: A meta-analysis. Epidemiol. Infect. 2020, 148, e228. [Google Scholar] [CrossRef]
- Geng, P.-L.; Song, L.-X.; An, H.; Huang, J.-Y.; Li, S.; Zeng, X.-T. Toll-Like Receptor 3 is Associated With the Risk of HCV Infection and HBV-Related Diseases. Medicine 2016, 95, e2302. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reference/Database | Type of Infectious Agent | Methodology/Genotyping Method/Population Size | Country/Continent/Ethnic Group | Gender Ratio (Male/Female)/Average Age of Participants | Absolute Count of Alleles and Genotypes (Cases/Controls) | HWE | p-Value | Results |
|---|---|---|---|---|---|---|---|---|
| Ishizaki et al. (2008) [20]/Science Direct | Virus/ssRNA- | Case-control/TaqMan essays/124 subjects (40 patients and 84 controls) | Japan/Asia/Not reported. | Male: 27/Female: 13 and 84 children (with a no reported gender); Age: 12.4 years | T alleles (33/46) Dominant Model Genotypes (17/29) | p > 0.05 | p = 0.03 | The mutant T allele of the SNP has been associated with the risk of subacute sclerosing panencephalitis (SSPE) related to measles virus persistence. |
| Kindberg et al. (2011) [21]/PUBMED | Virus/ssRNA+ | Case-control/PCR Genotyping/340 subjects (128 patients and 212 controls) | Lithuania/Europe/Not reported. | Gender ratio and average age not reported. | T alleles (59/141) Dominant Model Genotypes (50/101) | p > 0.05 | p < 0.05 | The wild-type C allele has been associated with the risk of contracting tick-borne encephalitis virus (TBEV). |
| Svensson et al. (2012) [22]/PUBMED | Virus/dsDNA | Case-control/TaqMan Assay/401 individuals (239 patients and 162 controls) | Sweden/Europe/Not reported. | Cases (Male: 136; Female: 103)/Controls (Male: 88; Female: 74); Age: 38 years. | T alleles (120/109) Dominant Model Genotypes (106/87) | p > 0.05 | p = 0.0272 | This SNP conferred protection against herpes simplex virus type 2 (HSV-2). |
| Lee; Brown; Razonable (2013) [23]/PUBMED | Virus/ssRNA+ | Case-control/PCR Genotyping/611 subjects (153 patients and 458 controls) | USA/North America/Not reported. | Male: 395/Female: 216; Age: 52 years | T alleles (102/237) Dominant Model Genotypes (88/207) | p > 0.05 | p = 0.03 | The presence of the SNP promoted susceptibility to the Hepatitis C virus (HCV). |
| Li; Zheng (2013) [24]/PUBMED | Virus/dsDNA | Case-control/PCR Genotyping/948 subjects (466 patients and 482 controls) | China/Asia/Not reported. | Cases (Male: 383/Female: 83); Controls (Male: 386/Female: 96); Age: 53.8 years | T alleles (326/249) Dominant Model Genotypes (274/226) | p > 0.05 | p = 0.004 | It is a risk factor for developing HBV infection. |
| Rong et al. (2013) [25]/PUBMED | Virus/dsDNA | Case-control/PCR Genotyping/914 subjects (452 patients and 462 controls) | China/Asia/Not reported. | Cases (Male: 340/Female: 112)/Controls (Male: 344/Female: 118); Age: 38.88 years | T alleles (296/235) Model Genotypes (254/212) | p > 0.05 | p = 0.002 | The SNP increased the risk of HBV infection. |
| Huik et al. (2013) [26]/PUBMED | Virus/ssRNA+ | Case-control/TaqMan Assay/842 subjects (172 HIV-positive patients and 670 controls) | Estonia/Europe/White. | Cases (Male: 133/Female: 69)/Controls (Male:Not reported/Female: Not reported); Age: 30 years. | T alleles (108/455) Dominant Model Genotypes (92/376) | p > 0.05 | p = 0.03 | The SNP promoted HIV-1 protection. |
| Mickienė et al. (2014) [27]/PUBMED | Virus/ssRNA+ | Case-control/PCR Genotyping/560 subjects (348 patients and 212 controls) | Lithuania/Europe/Not reported. | Cases (Male: 195/Female: 154); Controls (Male: Not reported/Female: Not reported). Age—Children TBE (Cases: 12.07/Controls: 11.43 years) Age—Adult severe TBE (Cases: 51.93 ± 15.419/Controls: 57.27 ± 15.108) Age—Adult TBE (Cases: 43.56/Controls: 46.97 years) | T alleles (195/141) Dominant Model Genotypes (157/101) | p > 0.05 | p = 0.02 | The SNP confers less risk of getting TBEV infection. |
| Alagarasu et al. (2015) [28]/PUBMED | Virus/ssRNA+ | Case-control/PCR-RFLP/229 subjects (120 patients and 109 controls) | India/Asia/Not reported. | Cases (Male:73/Female: 47)/Controls (Male: 67/Female: 42); Age: 31.3 years | T alleles (59/63) Dominant Model Genotypes (52/48) | p > 0.05 | p = 0.04 | The SNP confers a greater risk of acquiring the dengue virus. |
| BiyaniByyani et al. (2015) [29]/PUBMED and Science Direct | Virus/ssRNA+ | Case-control/PCR Genotyping/206 individuals (103 patients and 103 controls) | India/Asia/Not informed. | Gender ratio not reported; Age: 18.04. | T alleles (60/33) Dominant Model Genotypes (45/28) | p > 0.05 | p = 0.013 | The SNP has been associated with the risk of Japanese encephalitis virus (JEV) infection. |
| Wan et al. (2016) [30]/PUBMED | Virus/dsDNA | Case-control/TaqMan Assay/563 subjects (35 patients and 528 controls) | China/Asia/Not reported. | Cases (Male: 18/Female: 26)/Controls (Male: 358/Female: 295); Age: 26.43 years. | T alleles (17/329) Dominant Model Genotypes (17/282) | p > 0.05 | p = 0.736 | No significant association for SNP and HBV infection. |
| Chen et al. (2017) [31]/PUBMED | Virus/dsDNA | Case-control/PCR Genotyping/978 subjects (292 patients and 686 controls) | China/Asia/Not reported. | Male: 686/Female: 0; Age: 37 years. | T alleles (240/235) Dominant Model Genotypes (195/415) | p > 0.05 | p = 0.0001 | The SNP is a protective factor for HBV infection. |
| Grygorczuk et al. (2017) [32]/PUBMED | Virus/ssRNA+ | Case-control/TaqMan Assay/180 subjects (108 patients and 72 controls) | Poland/Europe/Not informed. | Gender: Not reported; Age: 42.44 years. | T alleles (59/41) Dominant Model Genotypes (51/36) | p > 0.05 | p < 0.05 | The higher frequency of wild-type C allele in patients was associated with TBEV infection. |
| Santos et al. (2019) [33]/PUBMED | Virus/ssRNA+ | Case-control/Quantitative Real Time-PCR/255 subjects (168 patients and 87 controls) | Brazil/South America/Not reported. | Not reported. | T alleles (94/43) Dominant Model Genotypes (83/37) | p > 0.05 | p = 0.042 | The SNP increased risk of developing Zika virus infection. |
| Posadas-Mondragón et al. (2020) [34]/PUBMED | Virus/ssRNA+ | Case-control/Real Time PCR/211 participants (165 patients and 46 controls) | Mexico/North America/Not reported. | DEN: 88/77; GP: Not reported; Age—Cases: 39.85, Controls: 49.52. | T alleles (96/56) Dominant Model Genotypes (83/23) | p > 0.05 | p = 0.31 | The mutant T allele of this SNP has been associated with protection from the dengue virus. |
| Habibabadi et al. (2020) [13]/PUBMED | Virus/ssRNA+ | Case-control/PCR-RFLP/218 subjects (100 patients and 118 controls) | Iran/Asia/Not reported. | Male: 179/Female: 39; Age—Case Group: 38.55, Control Group: 36.72 years. | T alleles (53/70) Dominant Model Genotypes (48/63) | p > 0.05 | p = 0.46 | No association between this SNP and human T-cell lymphotropic virus type 1 (HTLV-1). |
| Singh et al. (2021) [35]/PUBMED | Virus/ssRNA+ | Case-control/PCR Genotyping/196 individuals (98 patients and 98 controls) | India/Asia/Not informed. | Cases (Male: 36/Female: 62)/Controls (Male: 39; Female: 59); Age: 37.7 years | T alleles (52/28) Dominant Model Genotypes (42/25) | p > 0.05 | p = 0.10 | The presence of the mutant T allele of this SNP was associated with dengue susceptibility. |
| Redondo et al. (2022) [36]/PUBMED | Virus/dsDNA | Case-control/TaqMan Assay/204 individuals (50 cases and 154 controls) | Spain/Europe/Not reported. | Male: 146/Female: 58; Age: 54.6 years | T alleles (39/90) Dominant Model Genotypes (28/75) | p > 0.05 | p = 0.029 | Homozygous carriers of the T minor allele (TT genotype) of the highlighted SNP had a twofold increased risk of BK polyomavirus viremia (BKPyV) after kidney transplantation. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, M.J.A.; Silva, C.S.; da Silva Vieira, M.C.; dos Santos, P.A.S.; Frota, C.C.; Lima, K.V.B.; Lima, L.N.G.C. The Relationship between TLR3 rs3775291 Polymorphism and Infectious Diseases: A Meta-Analysis of Case-Control Studies. Genes 2023, 14, 1311. https://doi.org/10.3390/genes14071311
Silva MJA, Silva CS, da Silva Vieira MC, dos Santos PAS, Frota CC, Lima KVB, Lima LNGC. The Relationship between TLR3 rs3775291 Polymorphism and Infectious Diseases: A Meta-Analysis of Case-Control Studies. Genes. 2023; 14(7):1311. https://doi.org/10.3390/genes14071311
Chicago/Turabian StyleSilva, Marcos Jessé Abrahão, Caroliny Soares Silva, Marcelo Cleyton da Silva Vieira, Pabllo Antonny Silva dos Santos, Cristiane Cunha Frota, Karla Valéria Batista Lima, and Luana Nepomuceno Gondim Costa Lima. 2023. "The Relationship between TLR3 rs3775291 Polymorphism and Infectious Diseases: A Meta-Analysis of Case-Control Studies" Genes 14, no. 7: 1311. https://doi.org/10.3390/genes14071311