
The Unique Homothallic Mating-Type Loci of the Fungal Tree Pathogens Chrysoporthe syzygiicola and Chrysoporthe zambiensis from Africa

Abstract
:1. Introduction
2. Materials and Methods
2.1. Genome Sequencing, Assembly, and Analysis
2.2. Structure of the Mating-Type Loci of C. zambiensis and C. syzygiicola
2.3. Phylogenetic Analysis of Mating-Type Genes
3. Results
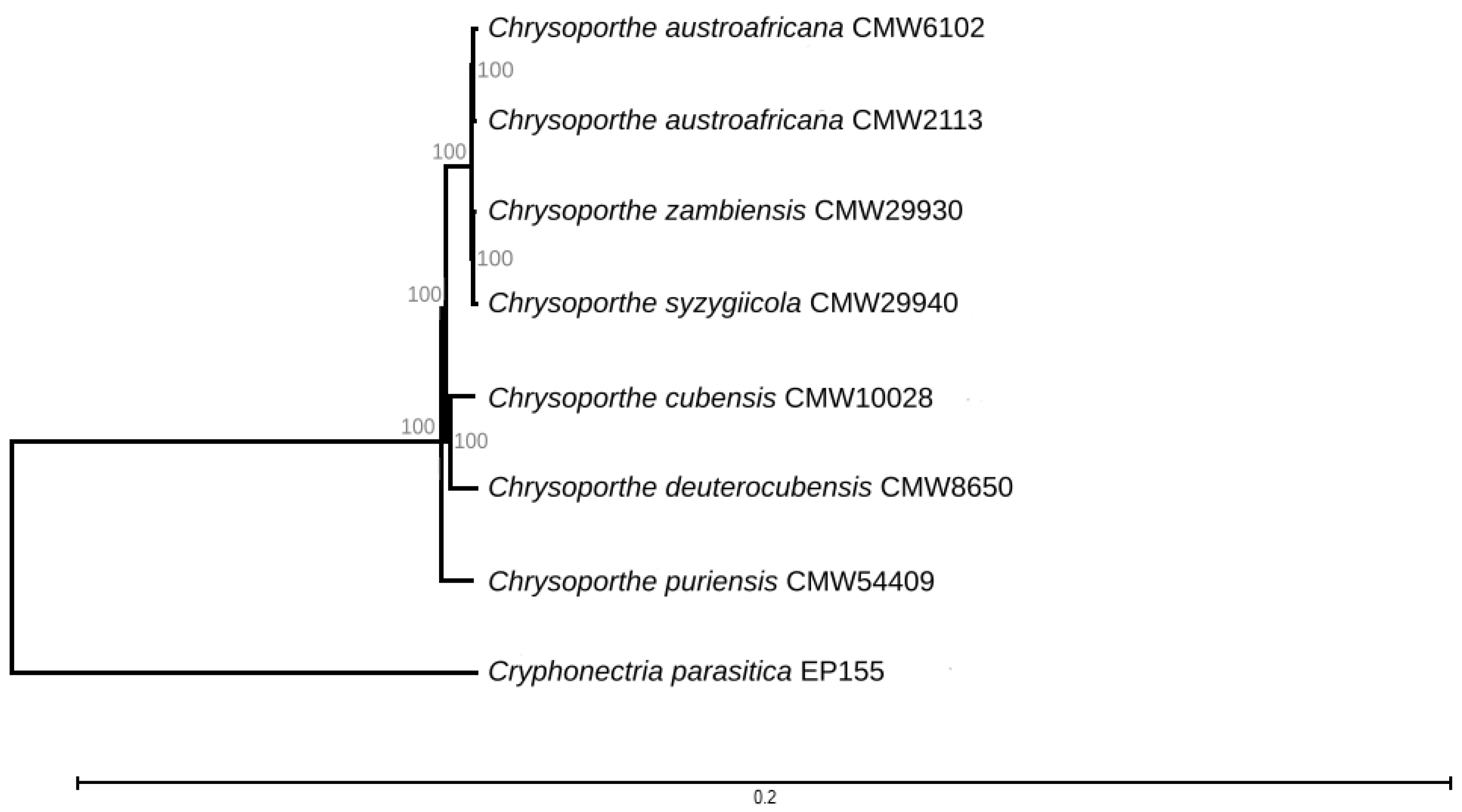
3.1. Genome Sequencing and Taxonomic Confirmation
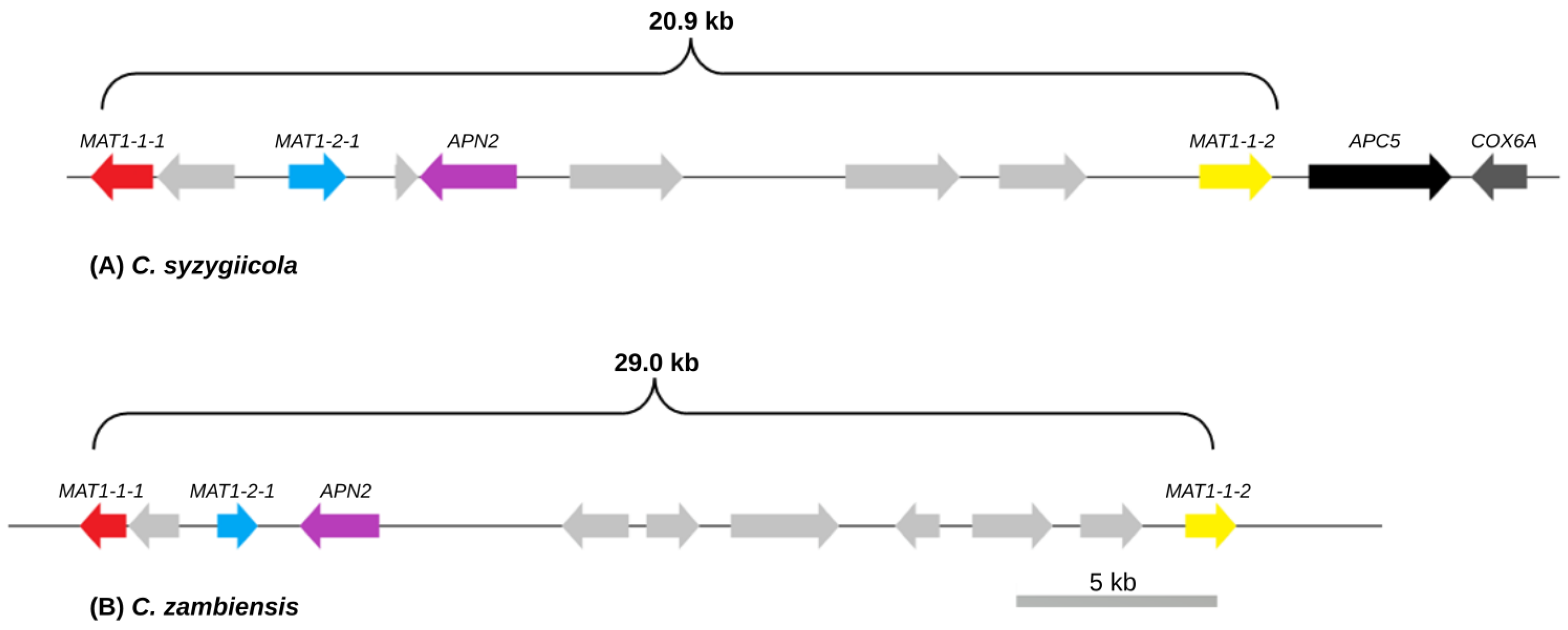
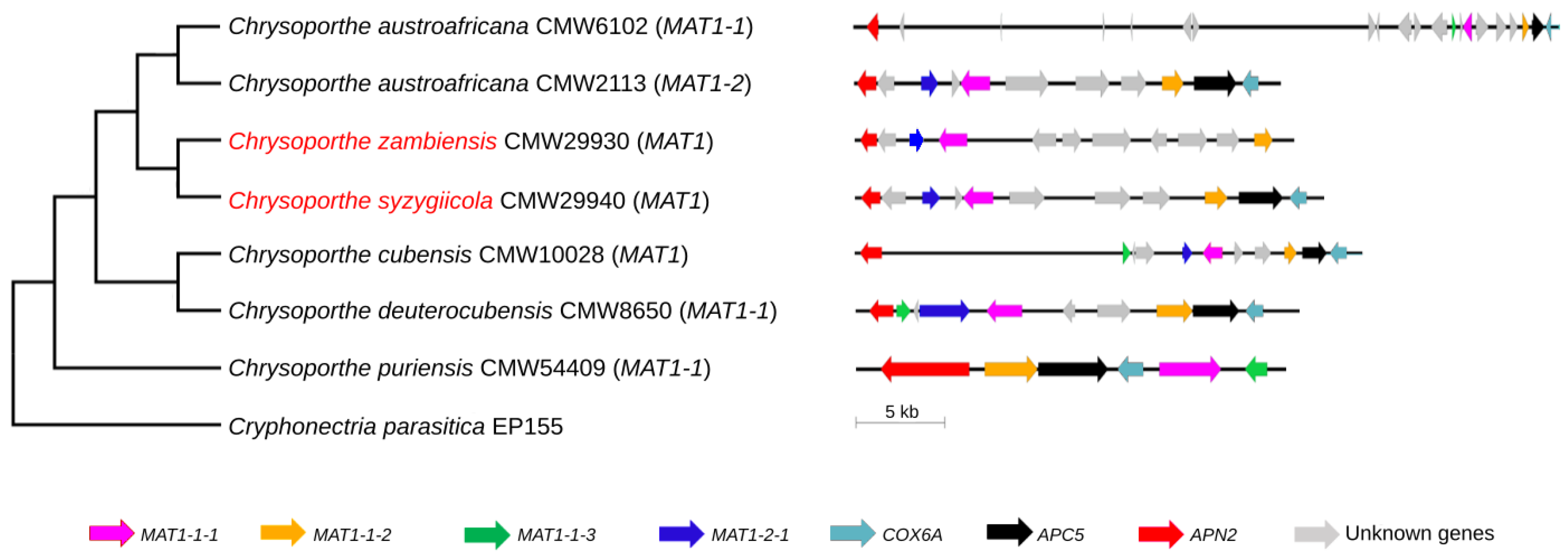
3.2. Mating-Type Genes and Structure of the Mating-Type Loci of C. syzygiicola and C. zambiensis
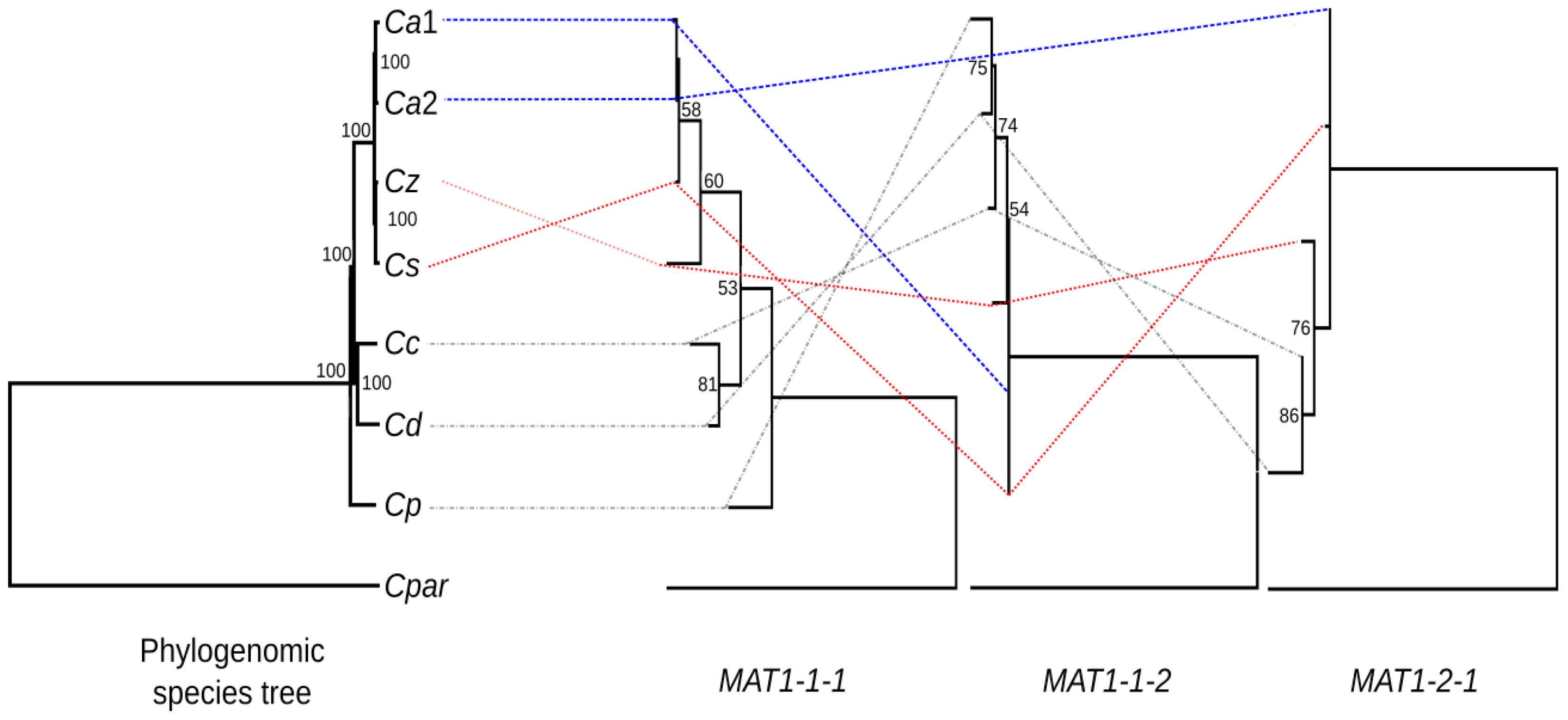
3.3. Phylogenetic Analysis of the Mating-Type Genes
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Debuchy, R.; Turgeon, B.G. Mating-type structure, evolution, and function in Euascomycetes. In The Mycota; Esser, K., Ed.; Springer: Berlin/Heidelberg, Germany, 2006; Volume 1, pp. 293–323. [Google Scholar] [CrossRef]
- Dyer, P.S.; Inderbitzin, P.; Debuchy, R. Mating-type structure, function, regulation and evolution in the Pezizomycotina. In The Mycota; Esser, K., Ed.; Springer: Berlin/Heidelberg, Germany, 2006; Volume 1, pp. 351–358. [Google Scholar] [CrossRef]
- Coppin, E.; Debuchy, R.; Arnaise, S.; Picard, M. Mating types and sexual development in filamentous ascomycetes. Micobiol. Mol. Biol. Rev. 1997, 61, 411–428. [Google Scholar] [CrossRef]
- Wilson, A.M.; Wilken, P.M.; van der Nest, M.A.; Steenkamp, E.T.; Wingfield, M.J.; Wingfield, B.D. Homothallism: An umbrella term for describing diverse sexual behaviours. IMA Fungus 2015, 6, 207–214. [Google Scholar] [CrossRef]
- Attanayake, R.N.; Tennekoon, V.; Johnson, D.A.; Porter, L.D.; del Río-Mendoza, L.; Jiang, D.; Chen, W. Inferring outcrossing in the homothallic fungus Sclerotinia sclerotiorum using linkage disequilibrium decay. Heredity 2014, 113, 353–363. [Google Scholar] [CrossRef] [PubMed]
- Billiard, S.; López-Villavicencio, M.; Hood, M.E.; Giraud, T. Sex, outcrossing and mating types: Unsolved questions in fungi and beyond. J. Evol. Biol. 2012, 25, 1020–1038. [Google Scholar] [CrossRef] [PubMed]
- Wilson, A.M.; Wilken, P.M.; Wingfield, M.J.; Wingfield, B.D. Genetic networks that govern sexual reproduction in the Pezizomycotina. Microbiol. Mol. Biol. Rev. 2021, 85, e00020-21. [Google Scholar] [CrossRef]
- Wilson, A.M.; Wilken, P.M.; van der Nest, M.A.; Wingfield, M.J.; Wingfield, B.D. It’s all in the genes: The regulatory pathways of sexual reproduction in filamentous ascomycetes. Genes 2019, 10, 330. [Google Scholar] [CrossRef]
- Casselton, L.A. Mate recognition in fungi. Heredity 2002, 88, 142–147. [Google Scholar] [CrossRef]
- Turgeon, B.G.; Yoder, O.C. Proposed nomenclature for mating type genes of filamentous ascomycetes. Fungal Genet. Biol. 2000, 31, 1–5. [Google Scholar] [CrossRef]
- Wilken, P.M.; Steenkamp, E.T.; Wingfield, M.J.; de Beer, Z.W.; Wingfield, B.D. Which MAT gene? Pezizomycotina (Ascomycota) mating-type gene nomenclature reconsidered. Fungal Biol. Rev. 2017, 31, 199–211. [Google Scholar] [CrossRef]
- Myburg, H.; Gryzenhout, M.; Heath, R.; Roux, J.; Wingfield, B.D.; Wingfield, M.J. Cryphonectria canker on Tibouchina in South Africa. Mycol. Res. 2002, 106, 1299–1306. [Google Scholar] [CrossRef]
- Nakabonge, G.; Roux, J.; Gryzenhout, M.; Wingfield, M.J. Distribution of Chrysoporthe canker pathogens on Eucalyptus and Syzygium spp. in Eastern and Southern Africa. Plant Dis. 2006, 90, 734–740. [Google Scholar] [CrossRef] [PubMed]
- Heath, R.N.; Gryzenhout, M.; Roux, J.; Wingfield, M.J. Discovery of the canker pathogen Chrysoporthe austroafricana on native Syzygium spp. in South Africa. Plant Dis. 2006, 90, 433–438. [Google Scholar] [CrossRef]
- Chen, S.F.; Gryzenhout, M.; Roux, J.; Xie, Y.J.; Wingfield, M.J.; Zhou, X.D. Identification and pathogenicity of Chrysoporthe cubensis on Eucalyptus and Syzygium spp. in South China. Plant Dis. 2010, 94, 1143–1150. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, M.E.S.; van der Merwe, N.A.; Wingfield, M.J.; Wingfield, B.D.; Soares, T.P.F.; Kanzi, A.M.; Ferreira, M.A. Chrysoporthe puriensis sp. nov. from Tibouchina spp. in Brazil: An emerging threat to Eucalyptus. Australas. Plant Pathol. 2021, 50, 29–40. [Google Scholar] [CrossRef]
- Van Heerden, S.W.; Wingfield, M.J. Genetic diversity of Cryphonectria cubensis isolates in South Africa. Mycol. Res. 2001, 105, 94–99. [Google Scholar] [CrossRef]
- Chungu, D.; Gryzenhout, M.; Muimba-Kankolongo, A.; Wingfield, M.J.; Roux, J. Taxonomy and pathogenicity of two novel Chrysoporthe species from Eucalyptus grandis and Syzygium guineense in Zambia. Mycol. Prog. 2010, 9, 379–393. [Google Scholar] [CrossRef]
- Gryzenhout, M.; Myburg, H.; van der Merwe, N.A.; Wingfield, B.D.; Wingfield, M.J. Chrysoporthe, a new genus to accommodate Cryphonectria cubensis. Stud. Mycol. 2004, 50, 119–142. [Google Scholar]
- Kanzi, A.M.; Steenkamp, E.T.; van der Merwe, N.A.; Wingfield, B.D. The mating system of the Eucalyptus canker pathogen Chrysoporthe austroafricana and closely related species. Fungal Genet. Biol. 2019, 123, 41–52. [Google Scholar] [CrossRef]
- Pöggeler, S.; Kück, U. Comparative analysis of the mating-type loci from Neurospora crassa and Sordaria macrospora: Identifcation of novel transcribed ORFs. Mol. Gen. Genet. 2000, 263, 292–301. [Google Scholar] [CrossRef]
- McGuire, I.C.; Marra, R.E.; Turgeon, B.G.; Milgroom, M.G. Analysis of mating-type genes in the chestnut blight fungus, Cryphonectria parasitica. Fungal Genet. Biol. 2001, 34, 131–144. [Google Scholar] [CrossRef]
- Duong, T.A.; de Beer, Z.W.; Wingfield, B.D.; Wingfield, M.J. Characterization of the mating-type genes in Leptographium procerum and Leptographium profanum. Fungal Biol. 2013, 117, 411–421. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, M.E.S.; Kanzi, A.M.; van der Merwe, N.A.; Wingfield, M.J.; Wingfield, B.D.; Silva, G.A.; Ferreira, M.A. Genetic variability in populations of Chrysoporthe cubensis and Chr. puriensis in Brazil. Australas. Plant Pathol. 2022, 51, 175–191. [Google Scholar] [CrossRef]
- Steenkamp, E.T.; Wingfield, B.D.; Coutinho, T.A.; Wingfield, M.J.; Marasas, W.F.O. Differentiation of Fusarium subglutinans f. sp. pini by histone gene sequence data. Appl. Environ. Microbiol. 1999, 65, 3401–3406. [Google Scholar]
- Jalili, V.; Afgan, E.; Gu, Q.; Clements, D.; Blankenberg, D.; Goecks, J.; Taylor, J.; Nekrutenko, A. The Galaxy platform for accessible, reproducible and collaborative biomedical analyses: 2020 update. Nucleic Acids Res. 2020, 48, W395–W402. [Google Scholar] [CrossRef]
- Koren, S.; Walenz, B.P.; Berlin, K.; Miller, J.R.; Bergman, N.H.; Phillippy, A.M. Canu: Scalable and accurate long-read assembly via adaptive k-mer weighting and repeat separation. Genome Res. 2017, 27, 22–736. [Google Scholar] [CrossRef] [PubMed]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality assessment tool for genome assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef]
- Simão, F.A.; Waterhouse, R.M.; Ioannidis, P.; Kriventseva, E.V.; Zdobnov, E.M. BUSCO: Assessing genome assembly and annotation completeness with single-copy orthologs. Bioinformatics 2015, 31, 3210–3212. [Google Scholar] [CrossRef] [PubMed]
- Stanke, M.; Morgenstern, B. AUGUSTUS: A web server for gene prediction in eukaryotes that allows user-defined constraints. Nucleic Acids Res. 2005, 33, W465–W467. [Google Scholar] [CrossRef]
- Stanke, M.; Tzvetkova, A.; Morgenstern, B. AUGUSTUS at EGASP: Using EST, protein and genomic alignments for improved gene prediction in the human genome. Genome Biol. 2006, 7, S11. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: A multiple sequence alignment method with reduced time and space complexity. Bmc Bioinform. 2004, 5, 113. [Google Scholar] [CrossRef] [PubMed]
- McGowan, J.; O’Hanlon, R.; Owens, R.A.; Fitzpatrick, D.A. Comparative denomic and proteomic analyses of three widespread Phytophthora species: Phytophthora chlamydospora, Phytophthora gonapodyides and Phytophthora pseudosyringae. Microorganisms 2020, 8, 653. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Quang Minh, B. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2014, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Gertz, E.M.; Yu, Y.K.; Agarwala, R.; Schäffer, A.A.; Altschul, S.F. Composition-based statistics and translated nucleotide searches: Improving the TBLASTN module of BLAST. BMC Biol. 2006, 4, 41. [Google Scholar] [CrossRef] [PubMed]
- Yin, Z.; Ke, X.; Li, Z.; Chen, J.; Gao, X.; Huang, L. Unconventional recombination in the mating type locus of heterothallic apple canker pathogen Valsa mali. Genes Genomes Genet. 2017, 7, 1259–1265. [Google Scholar] [CrossRef]
- Jones, P.; Binns, D.; Chang, H.Y.; Fraser, M.; Li, W.; McAnulla, C.; McWilliam, H.; Maslen, J.; Mitchell, A.; Nuka, G.; et al. InterProScan 5: Genome-scale protein function classification. Bioinformatics 2014, 30, 1236–1240. [Google Scholar] [CrossRef]
- Finn, R.D.; Attwood, T.K.; Babbitt, P.C.; Bateman, A.; Bork, P.; Bridge, A.J.; Chang, H.Y.; Dosztányi, Z.; El-Gebali, S.; Fraser, M.; et al. InterPro in 2017—Beyond protein family and domain annotations. Nucleic Acids Res. 2017, 45, D190–D199. [Google Scholar] [CrossRef]
- El-Gebali, S.; Mistry, J.; Bateman, A.; Eddy, S.R.; Luciani, A.; Potter, S.C.; Qureshi, M.; Richardson, L.J.; Salazar, G.A.; Smart, A.; et al. The Pfam protein families database in 2019. Nucleic Acids Res. 2019, 47, D427–D432. [Google Scholar] [CrossRef]
- Van der Nest, M.A.; Chávez, R.; De Vos, L.; Duong, T.A.; Gil-Durán, C.; Ferreira, M.A.; Lane, F.A.; Levicán, G.; Santana, Q.C.; Steenkamp, E.T.; et al. IMA genome—F14: Draft genome sequences of Penicillium roqueforti, Fusarium sororula, Chrysoporthe puriensis, and Chalaropsis populi. IMA Fungus 2021, 12, 5. [Google Scholar] [CrossRef]
- Wingfield, B.D.; Ades, P.K.; Al-Naemi, F.A.; Beirn, L.A.; Bihon, W.; Crouch, J.A.; de Beer, Z.W.; De Vos, L.; Duong, T.A.; Fields, C.J.; et al. IMA Genome—F4: Draft genome sequences of Chrysoporthe austroafricana, Diplodia scrobiculata, Fusarium nygamai, Leptographium lundbergii, Limonomyces culmigenus, Stagonosporopsis tanaceti, and Thielaviopsis punctulata. IMA Fungus 2015, 6, 233–248. [Google Scholar] [CrossRef]
- Wingfield, B.D.; Barnes, I.; de Beer, Z.W.; De Vos, L.; Duong, T.A.; Kanzi, A.M.; Naidoo, K.; Nguyen, H.D.T.; Santana, Q.C.; Sayari, M.; et al. IMA Genome—F5: Draft genome sequences of Ceratocystis eucalypticola, Chrysoporthe cubensis, C. deuterocubensis, Davidsoniella virescens, Fusarium temperatum, Graphilbum fragrans, Penicillium nordicum, and Thielaviopsis musarum. IMA Fungus 2015, 6, 493–506. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Sullivan, T.D.; Walton, E.; Floyd Averette, A.; Sakthikumar, S.; Cuomo, C.A.; Klein, B.S.; Heitman, J. Identification of the mating-type (MAT) locus that controls sexual reproduction of Blastomyces dermatitidis. Eukaryot. Cell 2013, 12, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Bihon, W.; Wingfield, M.J.; Slippers, B.; Duong, T.A.; Wingfield, B.D. MAT gene idiomorphs suggest a heterothallic sexual cycle in a predominantly asexual and important pine pathogen. Fungal Genet. Biol. 2014, 62, 55–61. [Google Scholar] [CrossRef]
- Nagel, J.H.; Wingfield, M.J.; Slippers, B. Evolution of the mating types and mating strategies in prominent genera in the Botryosphaeriaceae. Fungal Genet. Biol. 2018, 114, 24–33. [Google Scholar] [CrossRef]
- Kück, U.; Böhm, J. Mating type genes and cryptic sexuality as tools for genetically manipulating industrial molds. Appl. Microbiol. Biotechnol. 2013, 97, 9609–9620. [Google Scholar] [CrossRef]
- Kück, U.; Pöggeler, S.; Nowrousian, M.; Nolting, N.; Engh, I. Sordaria macrospora, a model system for fungal development. In Mycota XV; Anke, T., Weber, D., Eds.; Physiology and Genetics; Springer: Berlin/Heidelberg, Germany, 2009; Chapter 2; pp. 17–39. [Google Scholar]
- Kanematsu, S.; Adachi, Y.; Ito, T. Mating-type loci of heterothallic Diaporthe spp.: Homologous genes are present in opposite mating-types. Curr. Genet. 2007, 52, 11–22. [Google Scholar] [CrossRef]
- Martin, S.H.; Wingfield, B.D.; Wingfield, M.J.; Steenkamp, E.T. Structure and evolution of the Fusarium mating type locus: New insights from the Gibberella fujikuroi complex. Fungal Genet. Biol. 2011, 48, 731–740. [Google Scholar] [CrossRef]
- Yokoyama, E.; Yamagishi, K.; Hara, A. Structures of the mating-type loci of Cordyceps takaomontana. Appl. Environ. Microbiol. 2003, 69, 5019–5022. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, E.; Yamagishi, K.; Hara, A. Heterothallism in Cordyceps takaomontana. Fems Microbiol. Lett. 2005, 250, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Wilken, P.M.; Steenkamp, E.T.; Wingfield, M.J.; de Beer, Z.W.; Wingfield, B.D. DNA loss at the Ceratocystis fimbriata mating locus results in self-sterility. PLoS ONE 2014, 9, e92180. [Google Scholar] [CrossRef] [PubMed]
- Li, J.Q.; Wingfield, B.D.; Wingfield, M.J.; Barnes, I.; Fourie, A.; Crous, P.W.; Chen, S.F. Mating genes in Calonectria and evidence for a heterothallic ancestral state. Persoonia 2020, 45, 163–176. [Google Scholar] [CrossRef] [PubMed]
- Yong, M.; Yu, J.; Pan, X.; Yu, M.; Cao, H.; Qi, Z.; Du, Y.; Zhang, R.; Song, T.; Yin, X.; et al. MAT1-1-3, a mating type gene in the Villosiclava virens, is required for fruiting bodies and sclerotia formation, asexual development and pathogenicity. Front. Microbiol. 2020, 11, 1137. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, F.E.; Duhamel, M.; Carpentier, F.; Hood, M.E.; Foulongne-Oriol, M.; Silar, P.; Malagnac, F.; Grognet, P.; Giraud, T. Recombination suppression and evolutionary strata around mating-type loci in fungi: Documenting patterns and understanding evolutionary and mechanistic causes. New Phytol. 2020, 229, 2470–2491. [Google Scholar] [CrossRef]
- Fraser, J.A.; Stajich, J.E.; Tarcha, E.J.; Cole, G.T.; Inglis, D.O.; Sil, A.; Heitman, J. Evolution of the mating type locus: Insights gained from the dimorphic primary fungal pathogens Histoplasma capsulatum, Coccidioides immitis, and Coccidioides posadasii. Eukaryot. Cell 2007, 6, 622–629. [Google Scholar] [CrossRef]
- Roux, J.; Nkuekam, G.K.; Marincowitz, S.; van der Merwe, N.A.; Uchida, J.; Wingfield, M.J.; Chen, S.F. Cryphonectriaceae associated with rust-infected Syzygium jambos in Hawaii. MycoKeys 2020, 76, 49–79. [Google Scholar] [CrossRef]
- Kanzi, A.M.; Trollip, C.; Wingfield, M.J.; Barnes, I.; van der Nest, M.A.; Wingfield, B.D. Phylogenomic incongruence in Ceratocystis: A clue to speciation? BMC Genom. 2020, 21, 362. [Google Scholar] [CrossRef]
- Kanzi, A.M.; Wingfield, B.D.; Steenkamp, E.T.; Naidoo, S.; van der Merwe, N.A. Intron derived size polymorphism in the mitochondrial genomes of closely related Chrysoporthe species. PLoS ONE 2016, 11, e0156104. [Google Scholar] [CrossRef]
- Heitman, J.; Carter, D.A.; Dyer, P.S.; Soll, D.R. Sexual reproduction of human fungal pathogens. Cold Spring Harb. Perspect. Med. 2014, 4, 1–19. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Assembly Metric | C. zambiensis | C. syzygiicola |
|---|---|---|
| Genome size (bp) | 48,317,394 | 42,500,337 |
| Number of contigs | 211 | 233 |
| GC content (%) | 56.57 | 55.43 |
| N50 (bp) | 691,378 | 617,420 |
| L50 | 19 | 21 |
| BUSCO Statistic | C. zambiensis | C. syzygiicola |
|---|---|---|
| Overall completeness (%) | 96.2 | 95 |
| Complete BUSCO genes (C) | 3672 | 3631 |
| Single copy orthologs (S) | 3664 | 3625 |
| Duplicated orthologs (D) | 8 | 6 |
| Fragmented orthologs (F) | 34 | 39 |
| Missing orthologs (M) | 111 | 147 |
| NCBI | C. syzygiicola | C. zambiensis | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Species | Isolate | Assembly | MAT1-1-1 | MAT1-1-2 | MAT1-2-1 | AVG | MAT1-1-1 | MAT1-1-2 | MAT1-2-1 | AVG |
| C. syzygiicola | CMW29940 | PRJNA971112 | — | — | — | — | 91.29 | 98.11 | 97.12 | 95.50 |
| C. zambiensis | CMW29930 | PRJNA971112 | 91.29 | 98.11 | 97.12 | 95.50 | — | — | — | — |
| C. austroafricana | CMW6102 (MAT1-1) | ASM105115 | 97.51 | 100 | — | 98.76 | 92.57 | 98.11 | — | 95.34 |
| C. austroafricana | CMW2113 (MAT1-2) | ASM1607180 | 98.76 | 100 | 99.64 | 99.47 | 84.57 | 98.11 | 97.48 | 93.39 |
| C. cubensis | CMW10028 | ASM128231 | 84.65 | 97.57 | 97.12 | 93.11 | 87.55 | 96.22 | 97.48 | 93.75 |
| C. deuterocubensis | CMW8650 | ASM151382 | 87.55 | 96.76 | 94.24 | 92.85 | 83.55 | 95.41 | 94.60 | 91.19 |
| C. puriensis | CMW54409 (MAT1-1) | ASM1567895 | 84.23 | 95.41 | — | 89.82 | 83.82 | 94.05 | — | 88.94 |
| Cry. parasitica | EP155 | Crypa2 | 46.47 | 52.15 | 62.23 | 53.62 | 45.23 | 51.61 | 61.87 | 52.90 |
| Per gene average similarity | 84.35 | 91.43 | 90.07 | 81.23 | 90.23 | 89.71 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
van der Merwe, N.A.; Phakalatsane, T.; Wilken, P.M. The Unique Homothallic Mating-Type Loci of the Fungal Tree Pathogens Chrysoporthe syzygiicola and Chrysoporthe zambiensis from Africa. Genes 2023, 14, 1158. https://doi.org/10.3390/genes14061158
van der Merwe NA, Phakalatsane T, Wilken PM. The Unique Homothallic Mating-Type Loci of the Fungal Tree Pathogens Chrysoporthe syzygiicola and Chrysoporthe zambiensis from Africa. Genes. 2023; 14(6):1158. https://doi.org/10.3390/genes14061158
Chicago/Turabian Stylevan der Merwe, Nicolaas A., Tshiamo Phakalatsane, and P. Markus Wilken. 2023. "The Unique Homothallic Mating-Type Loci of the Fungal Tree Pathogens Chrysoporthe syzygiicola and Chrysoporthe zambiensis from Africa" Genes 14, no. 6: 1158. https://doi.org/10.3390/genes14061158