
An Attempt to Identify the Medaka Receptor for Somatolactin Alpha Using a Reverse Genetics Approach


Department of Chemical and Biological Sciences, Japan Women’s University, Tokyo 112-8681, Japan
*
Author to whom correspondence should be addressed.
Genes 2023, 14(4), 796; https://doi.org/10.3390/genes14040796
Submission received: 27 February 2023
/
Revised: 19 March 2023
/
Accepted: 24 March 2023
/
Published: 26 March 2023
(This article belongs to the Special Issue Genetic Studies of Fish)
Abstract
:Somatolactin alpha (SLα) is a fish-specific hormone involved in body color regulation. The growth hormone (GH) is another hormone that is expressed in all vertebrates and promotes growth. These peptide hormones act by binding to receptors (SLα receptor (SLR) and GH receptor (GHR)); however, the relationships between these ligands and their receptors vary among species. Here, we first performed phylogenetic tree reconstruction by collecting the amino-acid sequences classified as SLR, GHR, or GHR-like from bony fish. Second, we impaired SLR or GHR functions in medaka (Oryzias sakaizumii) using CRISPR/Cas9. Lastly, we analyzed SLR and GHR mutants for phenotypes to deduce their functions. Phylogenetic tree reconstruction was performed using a total of 222 amino-acid sequences from 136 species, which revealed that many GHRa and GHRb are vaguely termed as GHR or GHR-like, while showing no orthologous/paralogous relationships. SLR and GHR mutants were successfully established for phenotyping. SLR mutants exhibited premature lethality after hatching, indicating an essential role for SLR in normal growth. GHR mutations did not affect viability, body length, or body color. These results provide no evidence that either SLR or GHR functions as a receptor for SLα; rather, phylogenetically and functionally, they seem to be receptors for GH, although their (subfunctionalized) roles warrant further investigation.
1. Introduction
Somatolactin alpha (SLα) is a peptide hormone specific to fish that is secreted from the pars intermedia of the pituitary gland and regulates body color in medaka [1,2,3,4]. The body color of fish is adjusted according to the density and physiological condition of pigment cells, termed chromatophores, located on the body surface [5,6]. Chromatophores are classified into six types, according to the color of the pigment granules they contain: melanophores, erythrophores, xanthophores, leucophores, iridophores, and cyanophores. Medaka have four types of chromatophores and a spontaneous mutant, called color interfere (ci), which has a brighter gray color on the body surface than its wild-type (WT) counterpart because of an increase in leucophores and iridophores and the decrease in melanophores and xanthophores [2]. Conversely, the excessive expression of SLα in ci causes a darker yellow color on the body surface compared to ci with reduced expression because of the increase in melanophores and xanthophores and the decrease in leucophores and iridophores [3].
The growth hormone (GH) is another peptide hormone that is expressed in all vertebrates. This hormone is secreted from the anterior pituitary gland and is involved in growth promotion (e.g., somatic growth, metabolism of lipids and carbohydrates, and cell differentiation), reproduction, osmotic regulation, and immunogenicity [7]. The overexpression of GH in medaka (O. sakaizumii) triggers an increase in body size and severe infertility in most female fish [8].
SLα and GH are the closest relatives in the GH/prolactin (PRL) family and function by binding to the extracellular domain of their receptors (the SLα receptor (SLR) and GH receptor (GHR)) [9,10]. The amino-acid identity between SLα (231 residues) and GH (210 residues) in medaka is 23% [8]. Hence, we assumed previously that SLR and GHR would also be the closest relatives, which could be a clue for identifying an SLR in fish that is absent in land vertebrates.
Fukada et al. revealed the differences in the GH-binding ability of the two GH receptors present in masu salmon (Oncorhynchus masou) using an in vitro approach: one binds to both SLα and GH with low binding specificity, and more strongly to SLα than to GH; the other binds to GH alone [9,10]. Therefore, these receptors were hypothesized to be SLR and GHR, respectively. Conversely, both GHR1 and GHR2 bind to GH, but not to SLα, in zebrafish (Danio rerio) [11]. One of the two GHRs (GHR1) also binds specifically to GH in Japanese eels (Anguilla japonica) [12]. Therefore, the binding ability strangely varies according to the fish species. In addition, the SLR/GHR nomenclature is currently quite confusing; thus, it is necessary to clarify the evolutionary and functional relationships between these receptors and ligands (see Ocampo Daza et al. [13,14] for details).
Phylogenetic tree construction is a method that has been used to improve the understanding of genetic relationships because it can visualize the clade position of the gene sequences systematically and help elucidate the relationships among genes under an evolutional process. Reverse genetics is another useful technique for understanding gene function by genetically engineering specific nucleic acid sequences to assess phenotypes. To ensure the impairment of receptor function, the Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR)/CRISPR-associated protein 9 (CRISPR/Cas9) system [15,16,17] is a good candidate for generating frameshift mutations on the SLR and GHR genes and for assessing phenotypes.
Therefore, in this study, first, we performed an in silico phylogenetic tree reconstruction using amino-acid sequences from bony fish classified as SLR, GHR, or GHR-like using the RefSeq database at the National Center for Biotechnology Information (NCBI). Based on the position of the gene sequences, we propose the GHRa/GHRb nomenclature instead of the SLR/GHR one. Second, we impaired the receptor functions of SLR and GHR in medaka, using the CRISPR/Cas9 system, to produce mutants with frameshift mutations and establish various lineages. Lastly, we analyzed the phenotype of the slr and ghr mutants and estimated the function of these receptors.
2. Materials and Methods
2.1. Animals
Japanese rice fish of the HNI strain (WT) were used to establish new strains with specific mutations in the SLR or GHR gene induced by the CRISPR/Cas9 system. All fish were hatched and bred in our laboratory. This study was reviewed and approved by the Animal Experiment Committee of Japan Women’s University. Fertilized medaka eggs were reared in a Petri dish. After hatching, they were reared in a static water tank. A few weeks after hatching, the grown fish were transferred to a circulating tank with filtered water that was maintained at a temperature between 25 °C and 28 °C with light provided by white LED bulbs and white fluorescent lamps under a 14 h/10 h light/dark cycle. Larval fish were fed paramecium or powder food, and adult fish were provided with brine shrimp and commercial flake food (TetraMin) five times a day between 10h00 and 18h00.
2.2. Phylogenetic Tree Reconstruction
The amino-acid sequences from bony fish that were classified as SLR, GHR, or GHR-like were collected from the RefSeq database at the NCBI. GHRs from several land vertebrates were also collected for use as an outgroup. The listed sequences were aligned using ClustalW, and many redundant (e.g., isoforms X1, X2, or X3) or apparently odd (e.g., exceptionally short/long) sequences were manually eliminated. For some sequences, we trimmed the N-terminal region in accordance with other sequences. A maximum-likelihood tree was drawn using the RAxML (version 8.10.12)-installed Genetyx-Mac software (version 21.2.0), and some sequences with phylogenetic positions that apparently contradicted the traditional tree (likely because they were not GHR orthologs) were excluded from the list (we suspect that these were genuine SLRs). This final list (Supplementary Table S1) was used for phylogenetic reconstruction using the RAxML-installed Genetyx-Mac software with the amino-acid substitution model of PROTGAMMA, the substitution matrix of BLOSUM62a, and a bootstrapping value of 100.
2.3. Preparation and Microinjection of the gRNA and Cas9 mRNA
The target sequences for CRISPR/Cas9 were TCCTGCGCCCAAAATCAAAGG for SLR (Figure 1a) and CAGGAGCCTTGTGTTTGGTGG for GHR (Figure 1b). Properly designed oligonucleotide DNAs were purchased from Thermo Fisher Scientific (Waltham, MA, USA). A circular pDR274 plasmid (Addgene plasmid 42,250) was linearized with BsaI and confirmed by electrophoresis on a 1% agarose gel with ethidium bromide staining. The linear plasmid was excised from the gel and purified with the Wizard® SV Gel and PCR Clean-UP System (Promega, Madison, WI, USA). The annealed double-stranded oligonucleotides were subcloned into linearized pDR274 using the Ligation high Ver.2 reagent (Toyobo, Osaka, Japan), and proper insertion was confirmed by colony polymerase chain reaction (PCR) using M13 primers and direct sequencing. The double-stranded oligonucleotide-inserted pDR274 plasmid and the hCas9 plasmid were digested with DraI and NotI, respectively. The gRNA and capped Cas9 mRNA were synthesized using the AmpliScribe T7-flash Transcription Kit (Illumina, San Diego, CA, USA) and the mMESSENGE mMACHINE SP6 kit (Thermo Fisher Scientific), respectively. After DNase treatment, the transcripts were purified using the Rneasy Mini Kit (Qiagen, Venlo, Netherlands) and confirmed by electrophoresis on a 1% agarose gel. The concentration of the gRNA was measured using a NanoDrop Lite instrument (Thermo Fisher Scientific). All procedures were in accordance with the manufacturers’ protocols.
A mixed solution of gRNA and Cas9 mRNA (final concentrations of 25 ng/µL and 100 ng/µL, respectively) was placed in an injection needle and set in an IM-12 microinjector (Narishige, Tokyo, Japan). The RNA mixture (one type of gRNA and the Cas9 mRNA when checking mutagenesis efficiency, or two types of gRNA (SLR8 and GHR6) and the Cas9 mRNA for actual mutagenesis) was injected into the cytoplasm of 1-cell-stage medaka embryos of the HNI strain (WT) under an SMZ18 stereoscopic microscope (Nikon, Tokyo, Japan). The microinjected embryos were examined, dead ones were removed, and the remaining embryos were used for analysis and breeding.
2.4. Identification of ins/del and Frameshift Mutations in SLR and GHR
To confirm the insertion/deletion (ins/del) and frameshift mutations generated in SLR and GHR by the CRISPR/Cas9 system, a heteroduplex mobility assay (HMA) was conducted as described elsewhere [16,18,19]. Briefly, genomic DNA was extracted from 4-day-old embryos or the caudal fins of adult fish, and amplified by PCR (initiated at 98 °C for 30 s, followed by 30 cycles of 98 °C for 20 s, 57 °C for 1 min, 68 °C for 30 s, and 72 °C for 10 min) using the KAPA Taq polymerase (Roche, Basel, Switzerland) and the following appropriate primers (at a final concentration of 0.5 µM each): F, 5′-GATAAGCTTGTAAGGTAAATATTGAGG-3′ and R, 5′-TCTCATTGCTCTCAAACAAATC-3′ for SLR; and F, 5′-GTTCAGTTTCCTTGTGTCTTATATTTTCTGTAAAGGTTAAG-3′ and R, 5′-GGAACGCTTTAAAAATAGATCACATGACCGTAG-3′ for GHR. The amplification products were then electrophorized on a 12% polyacrylamide gel.
G0 adults that were confirmed to carry ins/del mutations in the caudal fin were mated with WT medaka (HNI) to obtain heterozygous F1 fish with the ins/del mutations. To confirm the transmission of the frameshift mutations to the F1 generation, HMA and sequencing were performed using a part of the caudal fin from F1 adult fish, as described above. After males and females with identical heterozygous frameshift mutations were obtained, the F1 fish were mated to generate the F2 generation. If males and females with identical frameshift mutations were not obtained, the F1 fish were backcrossed with WT fish.
2.5. Analysis of Viability
The heterozygous F1 males and females carrying identical frameshift mutations were mated with each other and the fertilized F2 eggs were collected. The littermates with unknown genotypes were bred in the same tanks en masse (one or more tanks per batch, depending on the number of individuals) and were genotyped using whole larvae or the caudal fin of adults, as described above. The expected ratio of WT, heterozygote, and homozygote fish in the F2 generation was 1:2:1 at any stage if the viability did not depend on the genotype.
2.6. Analysis of Body Color and Body Length
The body color and body size in the F2 generation were examined among genotypes by counting the density of chromatophores and measuring the distance between the snout and the tip of the caudal fin, respectively. On the day of chromatophore counting, adult medaka (over 3 months old (F2)) were placed in a white tank for at least 30 min to induce melanophore aggregation. Subsequently, they were anesthetized on ice for 1–2 min, placed on a Petri dish, and the epidermis on the dorsal side of the trunk (near the base of the dorsal fin) was photographed under a stereoscopic microscope. The number of melanophores, xanthophores, and leucophores was counted manually in an area of 1 mm × 3 mm, using Image J®, to calculate the cell density (cells/mm2). Because xanthophores were difficult to distinguish in raw micrographs, the contrast was increased by adjusting the balance of the blue color. The color and size measurements of the F2 littermates were performed before the identification of the genotypes of the subjects via HMA or sequencing, as described above.
2.7. Reverse Transcription Polymerase Chain Reaction
To confirm the nonsense-mediated mRNA decay (NMD) caused by the mutations in GHR, a reverse transcription PCR (RT-PCR) was performed. Total RNA was extracted from the livers of three adult fish, each from the WT, homozygous for ghr+1, and homozygous for ghr−1 groups, using ISOGENII (Nippon Gene). After treatment with deoxyribonuclease (RT Grade) for Heat Stop (Nippon Gene), the mRNA was reverse transcribed using a polyT primer (5′-ATTCTAGAGGCCGAGGCGGCCGACATGTTTTTTTTTTTTTTTTTVN-3′) and ReverTra Ace (Toyobo) to synthesize cDNA. All procedures were performed according to the manufacturers’ protocols. PCR was carried out as described above using the cDNA as a template and the following primers: F, 5′-GGTCTTCTGCTCATGCTCATGATGTC-3′ and R, 5′-CATTTCATGGTAGGAGGTTTCCCAGC-3′ for SLR; F, 5′-ATGGCGGCTGCGCTCAC-3′ and R, 5′-ATGCTGCTCAAAAGGTCAGGAATCAG-3′ for GHR; and F, 5′-ATGGATGATGACATTGCCGCACTG-3′ and R, 5′-TTAGAAGCATTTGCGGTGGACGATG-3′ for beta-actin. The expression of the transcripts was confirmed by electrophoresis on agarose gels with ethidium bromide staining. The number of PCR cycles was adjusted for each gene so that the WT band could be detected before reaching a plateau.
2.8. Statistics
The viability (ratio) of F2 offspring among genotypes (see Section 2.5) was analyzed using a chi-squared test with a null hypothesis of WT:hetero:homo = 1:2:1. The body length and chromatophore density between two or among three genotypes (see Section 2.6) were analyzed using Student’s t-test or one-way ANOVA, respectively. When there was a significant effect in the one-way ANOVA, the Tukey–Kramer test was additionally carried out to identify statistically significant differences among the levels (genotypes) of a parameter (body length or chromatophore density). Significant differences in the Tukey–Kramer test are indicated by p < 0.05. We did not correct the p value to reduce the type 1 error rate. The results are presented as the mean ± standard error (SE).
3. Results
3.1. Phylogenetic Reassessment of GHRa and GHRb in Teleosts
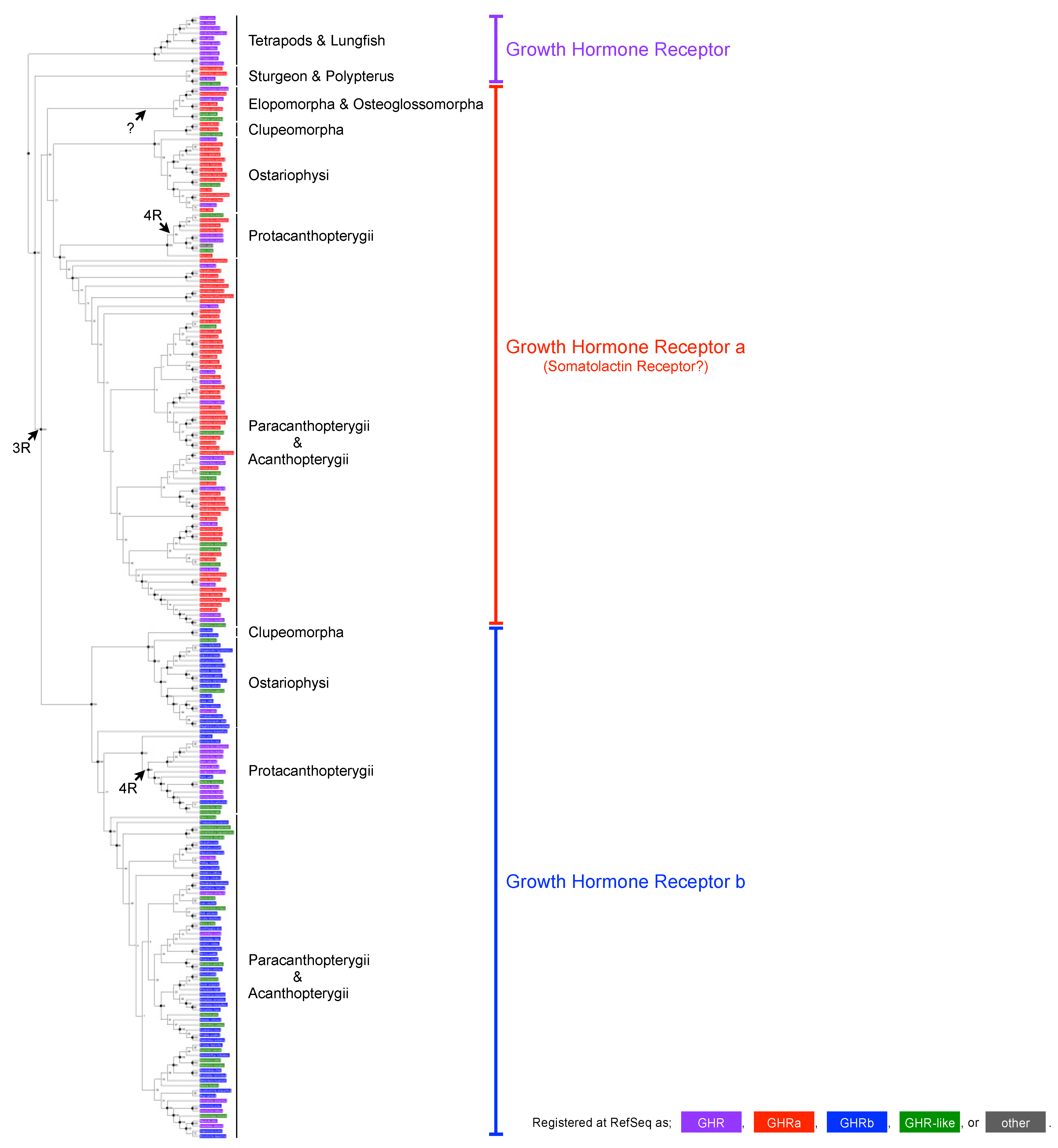
The common ancestor of teleosts underwent a third-round (3R) genome duplication, and many teleosts have two copies of GHR, which are currently termed GHRa and GHRb, whereas other vertebrates (e.g., tetrapods, lungfish, sturgeon, and polypterus) have only one copy. An additional genome duplication (4R) occurred at the common ancestor of Salmoninae, resulting in four copies of GHR. This evolutionary scenario was largely (with the exception of Elopomorpha) supported by the phylogenetic reconstruction performed here, using a total of 222 sequences from 136 species (Figure 2). However, this analysis revealed serious problems in the current nomenclature, as exemplified below.
First, the GHRs of sturgeons and polypterus, which did not experience the 3R genome duplication, should simply be termed as “GHR” however, two of the four sequences were classified as “GHRa”, and one was defined as “GHR-like”. Second, two copies of GHR were found in Elopomorpha (but not in Osteoglossomorpha), both of which belonged to the GHRa clade (because of a lineage-specific GHRa duplication?); however, one was designated as “GHR-like” despite the fact that it must be a genuine GHR (GHRa). Third, there were many other “GHR-like” entries in the GHRa and GHRb clades, which confusingly and wrongly indicated that these sequences are similar to, but different from, GHR. Fourth, several other “GHR-like” entries in the database could not be included in the present phylogenetic reconstruction. Their phylogenetic positions clearly contradicted the traditional ones, indicating that they are not orthologous to GHR; rather, they seem to be genuine “GHR-like” receptors. Fifth, there were many “GHR” entries in the GHRa and GHRb clades, which obscured the paralogous relationships. Sixth, GHRa and GHRb (i.e., phylogenetically distinct receptors) were indistinguishably designated as “GHR” or “GHR-like” for many species (particularly Salmoninae, having four copies of GHR).
The sequences of medaka (O. sakaizumii or O. latipes) were also indistinguishably designated as “GHR” in the database, whereas the phylogenetic reconstruction clearly demonstrated that XP_011477148 (and NP_001098560) and NP_001116377 (and XP_023816119, XP_023816118, and XP_023816117) are GHRa and GHRb, respectively. The former and latter were originally designated as SLR (DQ002886) and GHR (DQ010539), respectively. Although we do agree with the GHRa/GHRb nomenclature, we followed the original SLR/GHR nomenclature in the present study.
3.2. Mutagenesis Efficiency
Before we finally decided on the target sequences for CRISPR/Cas9 (see Methods), a total of eight sequences were tested. Among the eight gRNAs (SLR5/6/7/8 and GHR5/6/7/8), we were not able to transcribe SLR6 and SLR7 sufficiently for unknown reasons; therefore, the mutagenesis efficacy was examined using the remaining six gRNAs (Table 1). Co-injection of each gRNA with the Cas9 mRNA into 22–143 fertilized eggs yielded the 1-day viability, which ranged from 24.5% to 68.2% (Table 1). Hatched larvae injected with SLR8 or GHR6 were confirmed to have ins/del mutations in 35.7% or 25.0% of cases, respectively (Table 1), whereas SLR5, GHR5, GHR7, and GHR8 injections did not trigger any mutagenesis activity. Therefore, SLR8 and GHR6 were used thereafter as gRNAs for the SLR and GHR genes, respectively.
Using the genomic sequence of the HNI strain (version2.2.4; http://viewer.shigen.info/medaka/download.php, accessed on 25 March 2023), we manually verified that no genomic sequence other than the actual targets on chromosome 9 or 12 was identical to the 18 nucleotides adjacent to the PAM of SLR8 or GHR6 (i.e., TCCTGCGCCCAAAATCAA or CAGGAGCCTTGTGTTTGG). We also surveyed a genomic sequence using the Pattern Match program at the NBRP medaka (http://viewer.shigen.info/medakavw/crisprtool/, accessed on 25 March 2023); however, this tool is currently available only for the Hd-rR strain, O. latipes. When up to 2 mismatches were allowed in the 21-base target sequences, the 0 or 3 sequences could be potential off-targets for SLR8 or GHR6, respectively (Table 2). One of them was located about 320 kb downstream of the GHR locus, which is part of an intron of the PDZ and pleckstrin homology domains 1 gene.
Identical/similar sequences could exist in the HNI genome, and these observations cannot perfectly exclude a possible off-target effect; however, the risk of the simultaneous induction of frameshift mutations at multiple loci (see Table 3) and their co-inheritance by F1 and F2 siblings should not be high (with the exception of the tightly linked off-target). We also assumed that an intronic ins/del mutation would hardly affect the phenotype. Hence, we decided to use the SLR8 or GHR6 gRNAs for mutagenesis, rather than designing/testing additional gRNAs.
The mixture of two gRNAs and the Cas9 mRNA was microinjected into a total of 679 fertilized WT eggs. The mortality of embryos, the incidence of deformed embryos, and the incidence of normal development were 52.6%, 12.8%, and 34.6%, respectively. The overall efficiency of mutagenesis in the G0 larvae was predicted to be obtained with the following probabilities: fish with a single ins/del mutation on SLR (26.8%); fish with a single ins/del mutation on GHR (16.1%); fish with double ins/del mutations on both SLR and GHR (8.9%) (see Table 1). Of the 679 microinjected G0 eggs, 41 finally matured; therefore, the predicted values became 11.0 for a single mutation on SLR, 6.6 for a single mutation on GHR, 3.7 for double mutations on both SLR and GHR, and 19.8 for no mutations. The actual values as a result of HMAs were 8, 0, 7, and 26, respectively, which were significantly different from the predicted ones (p = 0.006, chi-squared test), albeit for an unknown reason.
A total of 9 out of the 15 G0 fish (numbered #1 to #15) that were confirmed to have ins/del mutations in the caudal fin were mated with WT fish or with each other, and 16 F1 larvae were collected from each cross. HMAs of the larvae revealed that 13 mutation types had been passed from 6 (or 7) G0 fish (#1, #2, #4 and/or #7, #5, #6, and #8) to the F1 generation, many of which were shown to be frameshift mutations by sequencing (Table 3).
Among them, the #1, #6, and #8 males with good growth conditions were preferentially selected, and 99, 128, and 176 F1 adult fish were obtained, respectively. A total of 12 and 4 mutation types were found, including 8 and 3 frameshift mutations in the SLR and GHR genes, respectively (Table 3). F1 fish with identical frameshift mutations in SLR were mated with each other to obtain the F2 generation. Regarding GHR, because the number of F1 adult fish with identical frameshift mutations was not sufficient for mating, they were backcrossed with WT fish to increase the number of heterozygotes.
3.3. Confirmation of Frameshift Mutations in slr and ghr Mutants
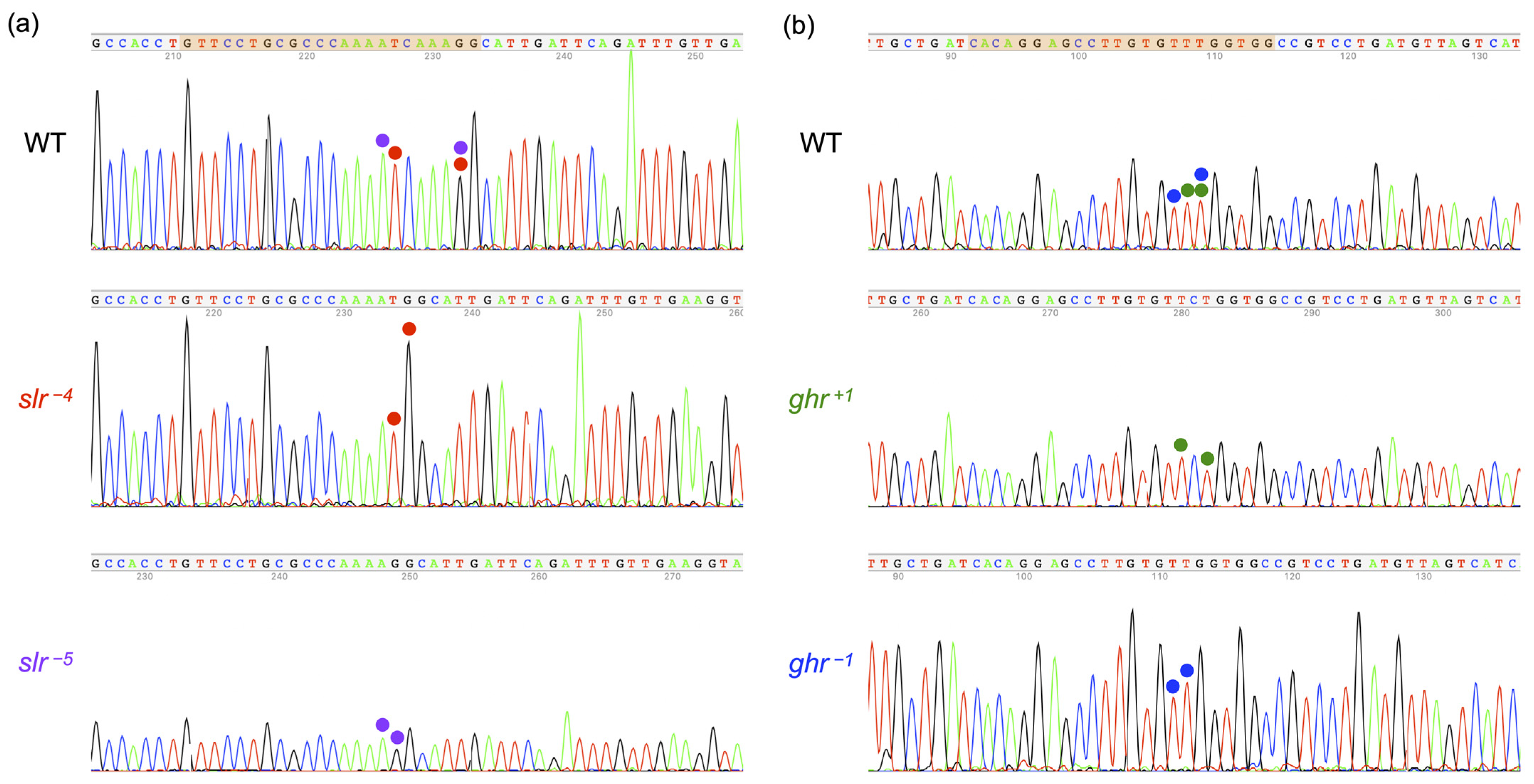
The successful introduction of frameshift mutations into slr and ghr mutants was confirmed by electropherograms of the target sequences: a 4- or 5-base deletion (CAAA or TCAAA) in slr−4 or slr−5 homozygotes, respectively (Figure 3a), and a 1-base insertion or deletion of C or T in ghr+1 or ghr−1 homozygotes, respectively (Figure 3b).
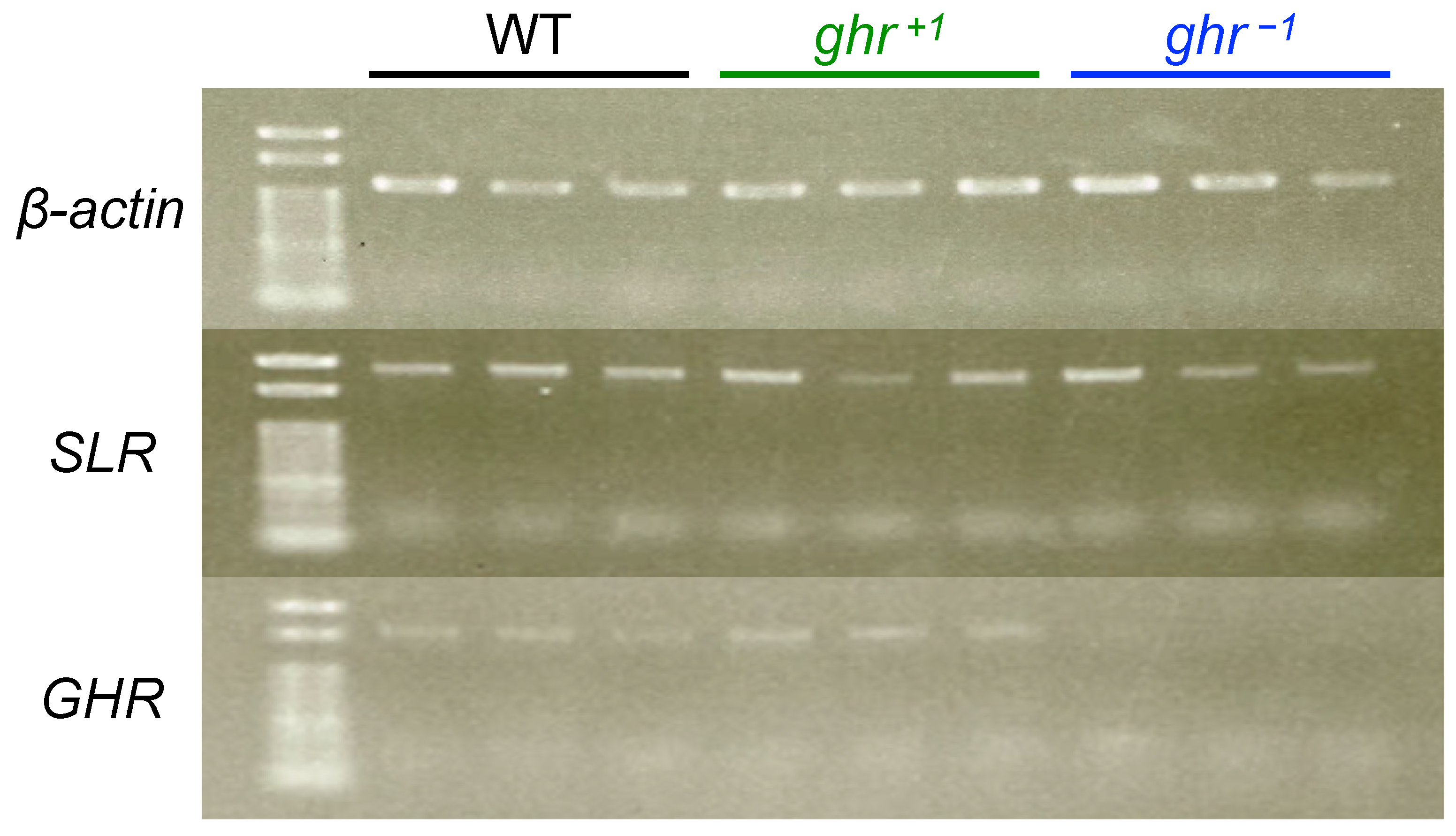
The frameshift mutations (premature stop codons) in GHR were also checked by NMD analysis. RT-PCR was performed using the total RNA from three adult fish livers, each among WT fish, ghr+1 homozygotes, and ghr−1 homozygotes; the products were electrophoresed on an agarose gel. The expression of beta-actin and SLR was detected in each individual from all strains (Figure 4). The expression of GHR was detected in WT and ghr+1 individuals, whereas the apparent NMD could be detected in three ghr−1 individuals, suggesting that the frameshift mutation successfully induced a premature stop codon in GHR, at least in ghr−1 fish.
It was not possible to confirm NMD in slr−4 or slr−5 homozygotes because of death before maturity (see Section 3.4). The only adults obtained were four slr−5 homozygotes, which were used in the body-color analysis (see Section 3.5) and were not obtained again in later crossings. Experiments using whole larvae might have solved this problem; however, the frameshift mutations do not always induce NMD (e.g., ghr+1) and, thus, we did not perform this experiment.
3.4. Viability and Body Size of Full Siblings Obtained by Crossing Heterozygous Mutants
The genotype ratio for WT, heterozygous, and homozygous fish among the siblings obtained between heterozygotes is expected to be 1:2:1. For slr−4 siblings, the ratio was 75:114:0 in the adult stage (a sum of six independent crosses), and no homozygotes were detected, which was significantly different from the expected value of 47.25:94.5:47.25 (p < 0.001, chi-squared test; Table 4). This result clearly demonstrated that the slr−4 mutation recessively suppresses the adult stage in medaka. For slr−5 siblings, the ratio (a sum of three independent crosses) was 18:33:4, which was also different from the expected ratio of 13.75:27.5:13.75 (p = 0.009). We could not obtain fertilized eggs from the slr−5 homozygotes or even slr−5 heterozygotes, and this mutation was unfortunately lost. The frozen sperm of slr−4 heterozygotes is available at the NBRP medaka (MT1356), together with those of missense mutants of slrC100F (MT1081) and slrF55I (MT1082).
For ghr+1 and ghr−1 siblings, the genotype ratios of 12:24:9 and 32:68:28 observed in adult fish (the sum of two or seven independent crosses, respectively) were not different from the expected values of 11.25:22.5:11.25 and 32:64:32, respectively (p = 0.741 or 0.687, respectively, chi-squared test; Table 4). The frozen sperm of ghr−1 homozygotes is available at the NBRP medaka (MT1354), together with those of missense mutants of ghrP161S (MT1078), ghrV166L (MT1079), and ghrW103R (MT1080).
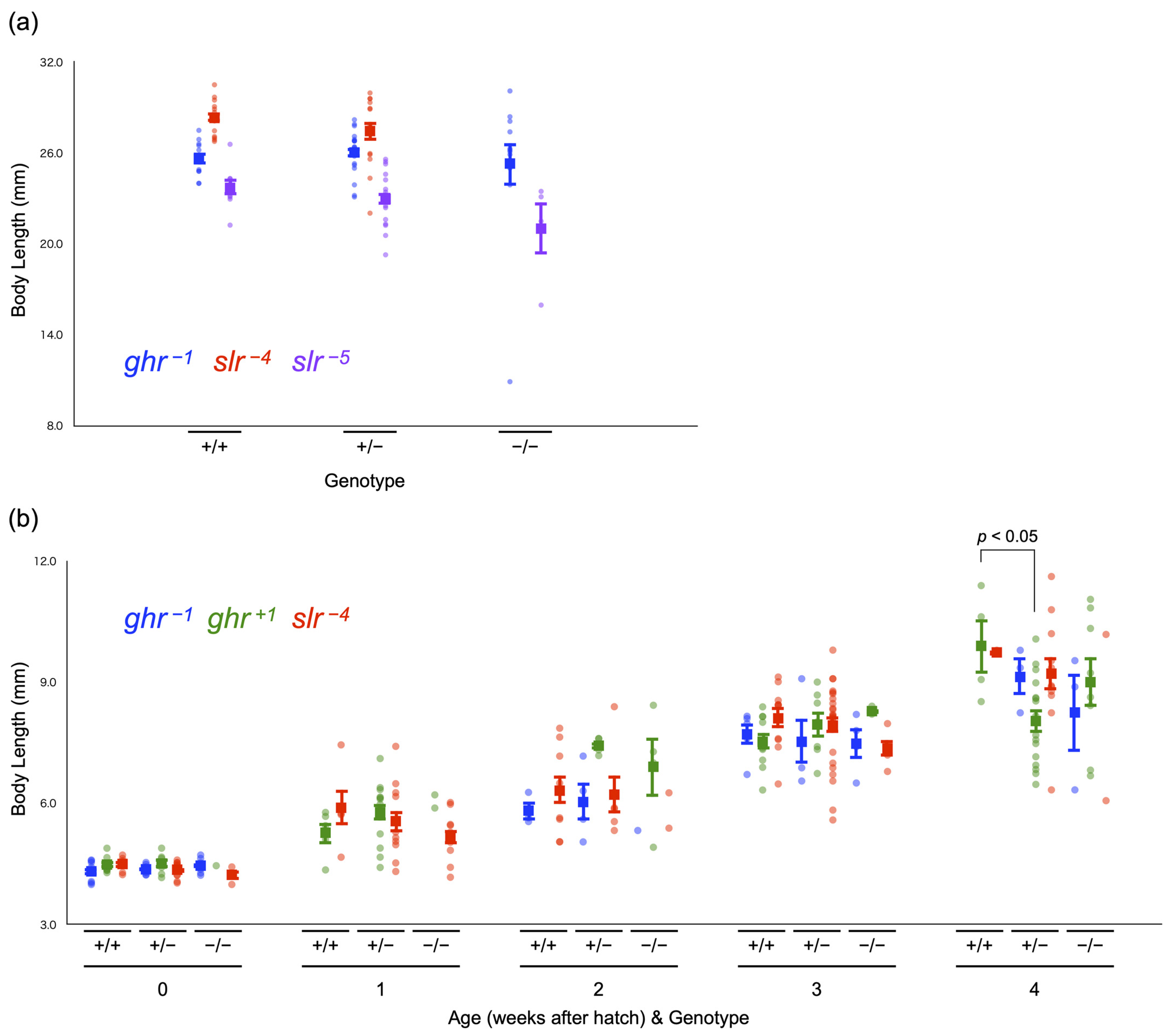
We measured the body length of some of these adults (Figure 5a). Among ghr−1 fish, the average body length of 5-month-old adult fish was similar among genotypes, as follows: WT: 25.6 ± 0.4 cm (n = 11), heterozygotes: 26.0 ± 0.3 cm (n = 23), and homozygotes: 25.3 ± 1.4 cm (n = 12) (mean ± SE; p = 0.736, one-way ANOVA). Although there were no homozygotes for slr−4, no difference in body length was observed between 4–11-month-old WT fish (28.3 ± 0.3 cm, n = 13) and heterozygotes (27.5 ± 0.6 cm, n = 14) (p = 0.249, Student’s t-test). For slr−5 (7-month-old fish), no significant difference was detected among genotypes, as follows: WT: 23.7 ± 0.5 cm (n = 8), heterozygotes: 23.0 ± 0.4 cm (n = 18), and homozygotes: 21.0 ± 1.7 cm (n = 4) (p = 0.097, one-way ANOVA); however, there seemed to be a tendency for homozygotes to be smaller than WT and heterozygote fish.
We also examined the genotype ratio at the larval stage (0–4 weeks post-hatching (wph)) among ghr−1, ghr+1, and slr−4 individuals. The ratios (a sum of 1–4 batches) fluctuated because of the small number of larvae included in the analysis (6–47 individuals per week per strain), which might accidentally have caused a significantly biased ratio at 3 wph in ghr+1 fish (p = 0.035, chi-squared test; Table 5). We noted that the percentages of slr−4 homozygotes at 2, 3, and 4 wph were consistently lower (by ~10%) than the expected value (25%), with one of these values (that obtained at 2 wph) being statistically significant (p = 0.027). This result indicates that slr−4 homozygous larvae are less viable than the full siblings of other genotypes, and this resulted in the absence of homozygotes in the adult stage.
We measured the body length of all larvae before the genotyping analysis (Figure 5b). For ghr+1 siblings, heterozygotes (8.0 ± 0.3 cm, n = 16) had a significantly shorter body length than WT fish (9.9 ± 0.7 cm, n = 4) at 4 wph (p = 0.036, one-way ANOVA and Tukey–Kramer test), whereas there was no difference among genotypes at other weeks of age (all p > 0.05, Student’s t-test or one-way ANOVA). Because we did not correct the p values to reduce the type 1 error rate, the above difference that was surprisingly detected between WT fish and heterozygotes could be an accidental event. For ghr−1 and slr−4, no difference in body length was observed at any wph (all p > 0.05, one-way ANOVA). These results do not statistically support the contention that the death of slr−4 homozygous larvae is caused by growth retardation, although there might be a trend toward this effect (e.g., at 1 and 3 wph).
3.5. Body Color of Full Siblings Obtained by Crossing Heterozygous Mutant Fish
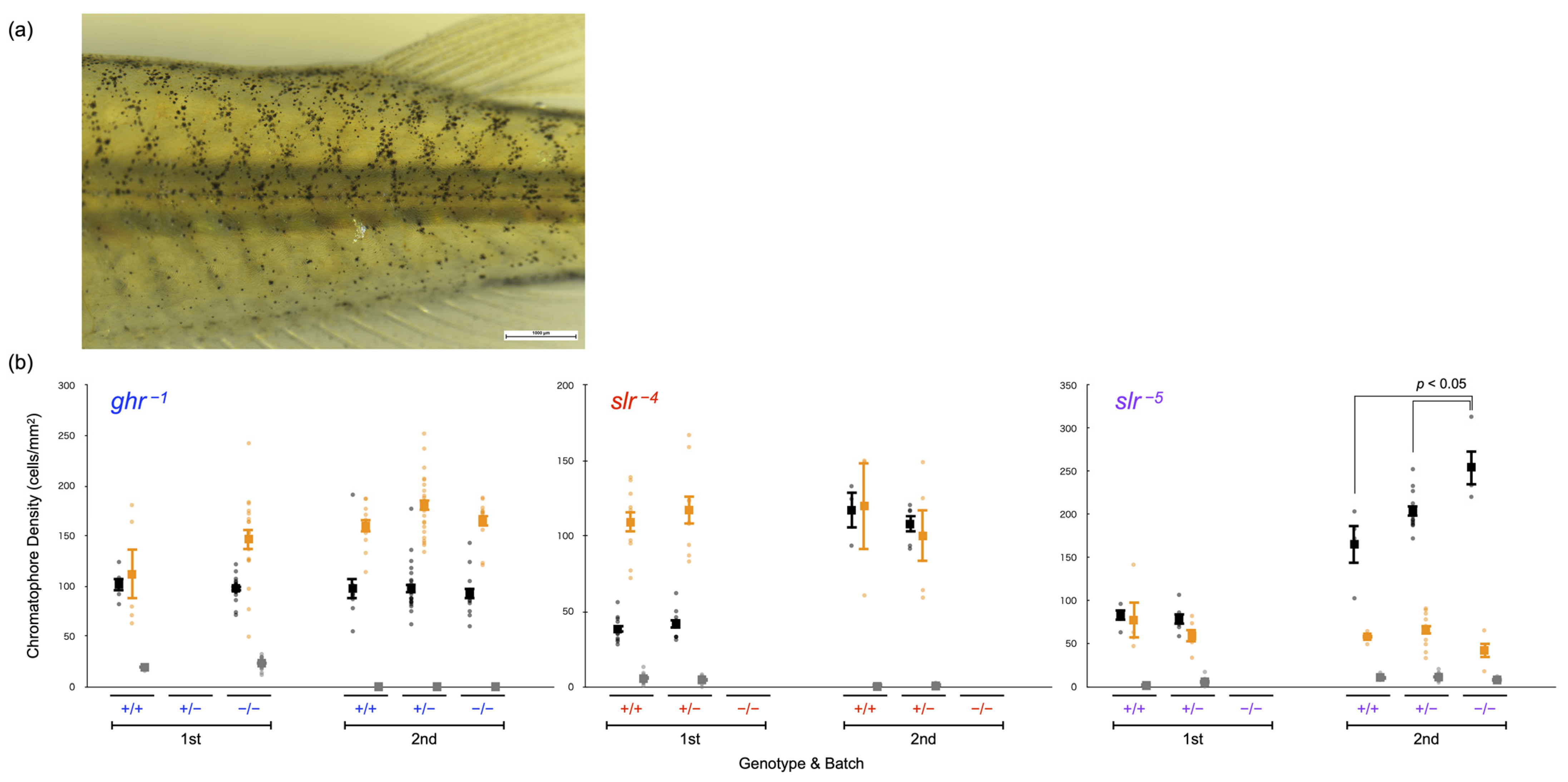
Lastly, we evaluated the body color (Figure 6), supposing that the density of leucophores and xanthophores in the slr mutants should, respectively, be increased and decreased if SLR functions as the receptor for SLα. The density of chromatophores in adult fish was examined twice for each mutation (ghr−1, slr−4, and slr−5) using independent batches. The results (body color) were not necessarily identical between the batches because the color of breeding tanks depends on accidental factors (e.g., algal growth) and medaka adapt their body color to the tank color. Therefore, the comparison among genotypes was performed only within (and not between) each batch.
Regarding the slr−4 mutation, for which no homozygotes were obtained, there was no significant difference between WT fish (n = 10 or 3) and heterozygotes (n = 9 or 5) for any of the chromatophores (p > 0.05, Student’s t-test). Furthermore, no significant difference was detected between WT fish (n = 8) and slr−5 heterozygotes (n = 18) in the first batch (p > 0.05, Student’s t-test). In the second batch, the density of melanophores in slr−5 homozygotes (n = 4, 254.9 ± 20.6 cells/mm2) was greater than that observed in WT fish (n = 12, 165.3 ± 21.9 cells/mm2) and heterozygotes (n = 4, 203.2 ± 6.7 cells/mm2) (p = 0.003, one-way ANOVA and Tukey–Kramer test); in turn, the differences in xanthophores or leucophores were not significant (p > 0.05).
We also examined the body color of the ghr−1 mutant and found no differences in any of the chromatophores among the genotypes (first batch: n = 5 and 17 for WT and homozygotes, respectively; second batch: n = 11, 23, and 12 for WT, heterozygotes, and homozygotes, respectively) (p > 0.05, Student’s t-test or one-way ANOVA).
4. Discussion
4.1. Confirmation of Frameshift Mutations in ghr/slr Mutants
We impaired the receptor function of SLR and GHR in medaka fish using the CRISPR/Cas9 system and used sequencing to confirm the successful generation of mutants carrying frameshift mutations (Figure 3). The introduction of the frameshift mutations into ghr−1 mutants was also double-checked using NMD (Figure 4). NMD is one of the eukaryotic mRNA quality control mechanisms that selectively degrade abnormal mRNA before translation when an immature stop codon occurs in the translation region [20]. Bands from transcripts obtained by RT-PCR, that cannot be confirmed or can be judged to be fainter than that obtained for the WT, imply that the mRNA is degraded after transcription, and it can be judged that NMD has occurred. Because the mRNA was degraded in ghr−1 fish because of NMD, an adequate amount of cDNA was not synthesized by reverse transcription, resulting in the lack of confirmation of GHR transcripts by RT-PCR.
Conversely, NMD does not always occur for mRNAs with immature stop codons [20]. The confirmation of the bands from transcripts implies that the transcribed RNA is retained without being degraded, and it can be judged that NMD has not occurred. The transcript products of GHR detected in all three ghr+1 homozygotes seemed to be equivalent in amount to those detected in WT fish, indicating that NMD had not occurred in ghr+1 homozygotes, although a functional GHR should not be transcribed from the mutated mRNA.
Regarding the slr mutations, we could not assess NMD because slr−4 or slr−5 homozygotes could never or seldom be obtained at the adult stage, respectively (Table 4). This severe mortality commonly shown by the slr mutants should support the successful introduction of frameshift mutations in the same gene, SLR. For instance, the establishment and analysis of multiple mutants could reduce the risk of misunderstanding the genotype–phenotype correlations caused by off-target effects.
4.2. Estimation of Medaka SLR Functions
SLR is supposed to be a receptor for SLα. Biallelic mutations in SLR in medaka decrease the melanophores in larval fish [21]; however, medaka homozygous for slr−5 had more melanophores than WT fish. These results indicate that SLR participates in body color regulation to a certain extent (e.g., it darkens larvae but brightens adults). If SLR functions as a necessary and sufficient receptor for SLα, the SLR (receptor) mutants should have exhibited the same body color as the SLα (ligand) mutant, ci. However, the dramatic increase and decrease in leucophores and xanthophores, respectively, that have repeatedly been reported for the ligand mutant [2,3,8], were not at all reproduced in the receptor mutant (Figure 6).
In addition, the receptor mutations had a serious effect on vital activities before sexual maturity. Despite repeated attempts to preserve them, all individuals that were homozygous for slr−4 died before maturity and we obtained only four slr−5 homozygous adults that were unable to reproduce (Table 4). The results obtained for the receptor mutants were also incompatible with those of the ligand mutant, ci, which exhibited ordinary growth and viability [8].
The present results of a dissimilar body coloration in slr−5 individuals compared with that of ci and the difficulty in growth (survival) commonly observed in the slr−5 and slr−4 mutants suggest one of the two following possibilities: (1) SLR functions as a receptor for SLα, but there is another factor (e.g., receptor) that compensates for the leucophore/xanthophore deficiency; or (2) SLR is a receptor for GH, rather than SLα. The phylogenetic reconstruction (Figure 2) showed that the medaka SLR was surely orthologous to the GHR of tetrapods. Moreover, we found several “GHR-like” sequences that did not belong to either the GHRa (SLR) or GHRb (GHR) clades. Based on the present results, we favor the second possibility described above, i.e., SLR is a receptor that is essential for normal growth and possibly binds to GH. The slr mutants might be unable to receive sufficient GH signals, resulting in low viability, although inhibition of larval growth could not clearly be demonstrated by measuring the body length (Figure 5b).
4.3. Estimation of Medaka GHR Functions
GHR mutations in humans, pigs, and mice have been shown to inhibit growth [22,23,24,25]; however, none of the medaka ghr mutants (but not the slr mutants) showed any specific phenotype regarding viability, body length, or body color, suggesting that GHR is unlikely to be essential for GH-mediated growth promotion or SLα-mediated body color regulation in medaka. When the effect of gene knockout is not reflected in the phenotype, the role of the gene cannot be elucidated; therefore, the reverse genetics method used in this study could not clarify the function of medaka GHR. The phylogeny (Figure 2) at least supported the contention that medaka GHR (and SLR) is orthologous to tetrapod GHR. Even if both medaka orthologs could function as receptors for GH, the present results indicate that the role of GHR (but not SLR) is dispensable for growth. We were interested in the phenotypes of SLR/GHR double mutants and intercrossed the double heterozygotes. However, the high mortality of the slr mutations did not allow the establishment of the double-mutant lines (similar to the single mutants; Table 4).
We hope that the function of GHR will be elucidated by methods other than reverse genetics or by reverse genetics in other fish species. In addition, it is possible that SLR and GHR complement each other’s functions. If so, SLR/GHR double-knockout lineages need to be established and their phenotypes must be examined in the future.
5. Conclusions
A phylogenetic tree was systematically reconstructed, and a GHRa/GHRb nomenclature, instead of the SLR/GHR nomenclature, was proposed based on the positions of the gene sequences. SLR and GHR mutants were successfully established in medaka, and slr−4, slr−5, ghr+1, and ghr−1 were selected for phenotyping. The slr mutants exhibited premature lethality, indicating the inactivation of the GH signal at the receptor. In turn, the ghr mutations did not affect viability, body length, or body color. These results indicate that the medaka GHR paralogs would phylogenetically and functionally be GH receptors. The receptor for SLα remains an open question.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/genes14040796/s1, Table S1: A list of sequences used for the phylogenetic reconstruction.
Author Contributions
Conceptualization, S.F.; methodology, S.F.; validation, Y.M. and S.F.; formal analysis, Y.M. and S.F.; investigation, M.K., Y.O., E.N. and H.O.; resources, S.F.; data curation, Y.M. and S.F.; writing—original draft preparation, Y.M.; writing—review and editing, S.F.; visualization, Y.M. and S.F.; supervision, S.F.; project administration, S.F.; funding acquisition, S.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by research funds from J.W.U. to S.F.
Institutional Review Board Statement
This study was reviewed and approved by the Animal Experiment Committee of Japan Women’s University (20210406).
Informed Consent Statement
Not applicable.
Data Availability Statement
Data will be made available on request.
Acknowledgments
We are grateful to Moeka Takizawa and Ayane Hasegawa in our laboratory for data collection.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ono, M.; Takayama, Y.; Rand-Weaver, M.; Sakata, S.; Yasunaga, T.; Noso, T.; Kawauchi, H. cDNA cloning of somatolactin, a pituitary protein related to growth hormone and prolactin. Proc. Natl. Acad. Sci. USA 1990, 87, 4330–4334. [Google Scholar] [CrossRef] [Green Version]
- Fukamachi, S.; Sugimoto, M.; Mitani, H.; Shima, A. Somatolactin selectively regulates proliferation and morphogenesis of neural-crest derived pigment cells in medaka. Proc. Natl. Acad. Sci. USA 2004, 101, 10661–10666. [Google Scholar] [CrossRef] [Green Version]
- Fukamachi, S.; Yada, T.; Meyer, A.; Kinoshita, M. Effects of constitutive expression of somatolactin alpha on skin pigmentation in medaka. Gene 2009, 442, 81–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasano, Y.; Yoshimura, A.; Fukamachi, S. Reassessment of the function of somatolactin alpha in lipid metabolism using medaka mutant and transgenic strains. BMC Genet. 2012, 13, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugimoto, M.; Uchida, N.; Hatayama, M. Apoptosis in skin pigment cells of the medaka, Oryzias latipes (Teleostei), during long-term chromatic adaptation: The role of sympathetic innervation. Cell Tissue Res. 2000, 301, 205–216. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, M. Morphological color changes in fish: Regulation of pigment cell density and morphology. Microsc. Res. Tech. 2002, 58, 496–503. [Google Scholar] [CrossRef]
- Reinecke, M.; Björnsson, B.T.; Dickhoff, W.W.; McCormick, S.D.; Navarro, I.; Power, D.M.; Gutiérrez, J. Growth hormone and insulin-like growth factors in fish: Where we are and where to go. Gen. Comp. Endocrinol. 2005, 142, 20–24. [Google Scholar] [CrossRef]
- Komine, R.; Nishimaki, T.; Kimura, T.; Oota, H.; Naruse, K.; Homma, N.; Fukamachi, S. Transgenic medaka that overexpress growth hormone have a skin color that does not indicate the activation or inhibition of somatolactin-a signal. Gene 2016, 584, 38–46. [Google Scholar] [CrossRef]
- Fukada, H.; Ozaki, Y.; Pierce, A.L.; Adachi, S.; Yamauchi, K.; Hara, A.; Swanson, P.; Dickhoff, W.W. Salmon growth hormone receptor: Molecular cloning, ligand specificity, and response to fasting. Gen. Comp. Endocrinol. 2004, 139, 61–71. [Google Scholar] [CrossRef]
- Fukada, H.; Ozaki, Y.; Pierce, A.L.; Adachi, S.; Yamauchi, K.; Hara, A.; Swanson, P.; Dickhoff, W.W. Identification of the salmon somatolactin receptor, a new member of the cytokine receptor family. Endocrinology 2005, 146, 2354–2361. [Google Scholar] [CrossRef]
- Chen, M.; Huang, X.; Yuen, D.S.; Cheng, C.H. A study on the functional interaction between the GH/PRL family of polypeptides with their receptors in zebrafish: Evidence against GHRI being the receptor for somatolactin. Mol. Cell. Endocrinol. 2011, 337, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Ozaki, Y.; Fukada, H.; Kazero, Y.; Adachi, S.; Hara, A.; Yamauchi, K. Molecular cloning and characterization of growth hormone receptor and its homologue in the Japanese eel (Anguilla japonica). Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2006, 143, 422–431. [Google Scholar] [CrossRef] [PubMed]
- Ocampo Daza, D.; Larhammar, D. Evolution of the growth hormone, prolactin, prolactin 2 and somatolactin family. Gen. Comp. Endocrinol. 2018, 264, 94–112. [Google Scholar] [CrossRef]
- Ocampo Daza, D.; Larhammar, D. Evolution of the receptors for growth hormone, prolactin, erythropoietin and thrombopoietin in relation to the vertebrate tetraploidizations. Gen. Comp. Endocrinol. 2018, 257, 143–160. [Google Scholar] [CrossRef] [PubMed]
- Hwang, W.Y.; Fu, Y.; Reyon, D.; Maeder, M.L.; Tsai, S.Q.; Sander, J.D.; Peterson, R.T.; Yeh, J.R.; Joung, J.K. Efficient genome editing in zebrafish using a CRISPR-Cas system. Nat. Biotechnol. 2013, 3, 227–229. [Google Scholar] [CrossRef] [Green Version]
- Ansai, S.; Kinoshita, M. Targeted mutagenesis using CRISPR/Cas system in medaka. Biol. Open. 2014, 3, 362–371. [Google Scholar] [CrossRef] [Green Version]
- Lander, E.S. The heroes of CRISPR. Cell 2016, 164, 18–28. [Google Scholar] [CrossRef] [Green Version]
- Homma, N.; Harada, Y.; Uchikawa, T.; Kamei, Y.; Fukamachi, S. Protanopia (red color-blindness) in medaka: A simple system for producing color-blind fish and testing their spectral sensitivity. BMC Genet. 2017, 18, 10. [Google Scholar] [CrossRef] [Green Version]
- Kanazawa, N.; Goto, M.; Harada, Y.; Takimoto, C.; Sasaki, Y.; Uchikawa, T.; Kamei, Y.; Matsuo, M.; Fukamachi, S. Changes in a cone opsin repertoire affect color-dependent social behavior in medaka but not behavioral photosensitivity. Front. Genet. 2020, 11, 801. [Google Scholar] [CrossRef]
- Supek, F.; Lehner, B.; Lindeboom, R.G.H. To NMD or not to NMD: Nonsense-mediated mRNA decay in cancer and other genetic diseases. Trends Genet. 2021, 37, 657–668. [Google Scholar] [CrossRef]
- Delgadin, T.H.; Castañeda-Cortés, D.C.; Sacks, C.; Breccia, A.; Fernandino, J.I.; Vissio, P.G. Morphological colour adaptation during development in fish: Involvement of growth hormone receptor 1. J. Exp. Biol. 2020, 223, jeb230375. [Google Scholar] [CrossRef] [PubMed]
- Godowski, P.J.; Leung, D.W.; Meacham, L.R.; Galgani, J.P.; Hellmiss, R.; Keret, R.; Rotwein, P.S.; Parks, J.S.; Laron, Z.; Wood, W.I. Characterization of the human growth hormone receptor gene and demonstration of a partial gene deletion in two patients with Laron-type dwarfism. Proc. Natl. Acad. Sci. USA 1989, 20, 8083–8087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laron, Z. Laron syndrome (primary growth hormone resistance or insensitivity): The personal experience 1958–2003. J. Clin. Endocrinol. Metab. 2004, 89, 1031–1044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, D.; Li, F.; Li, Q.; Li, J.; Zhao, Y.; Hu, X.; Zhang, R.; Li, N. Generation of a miniature pig disease model for human Laron syndrome. Sci. Rep. 2015, 5, 15603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Xu, B.C.; Maheshwari, H.G.; He, L.; Reed, M.; Lozykowski, M.; Okada, S.; Cataldo, L.; Coschigamo, K.; Wagner, T.E.; et al. A mammalian model for Laron syndrome produced by targeted disruption of the mouse growth hormone receptor/binding protein gene (the Laron mouse). Proc. Natl. Acad. Sci. USA 1997, 94, 13215–13220. [Google Scholar] [CrossRef] [Green Version]
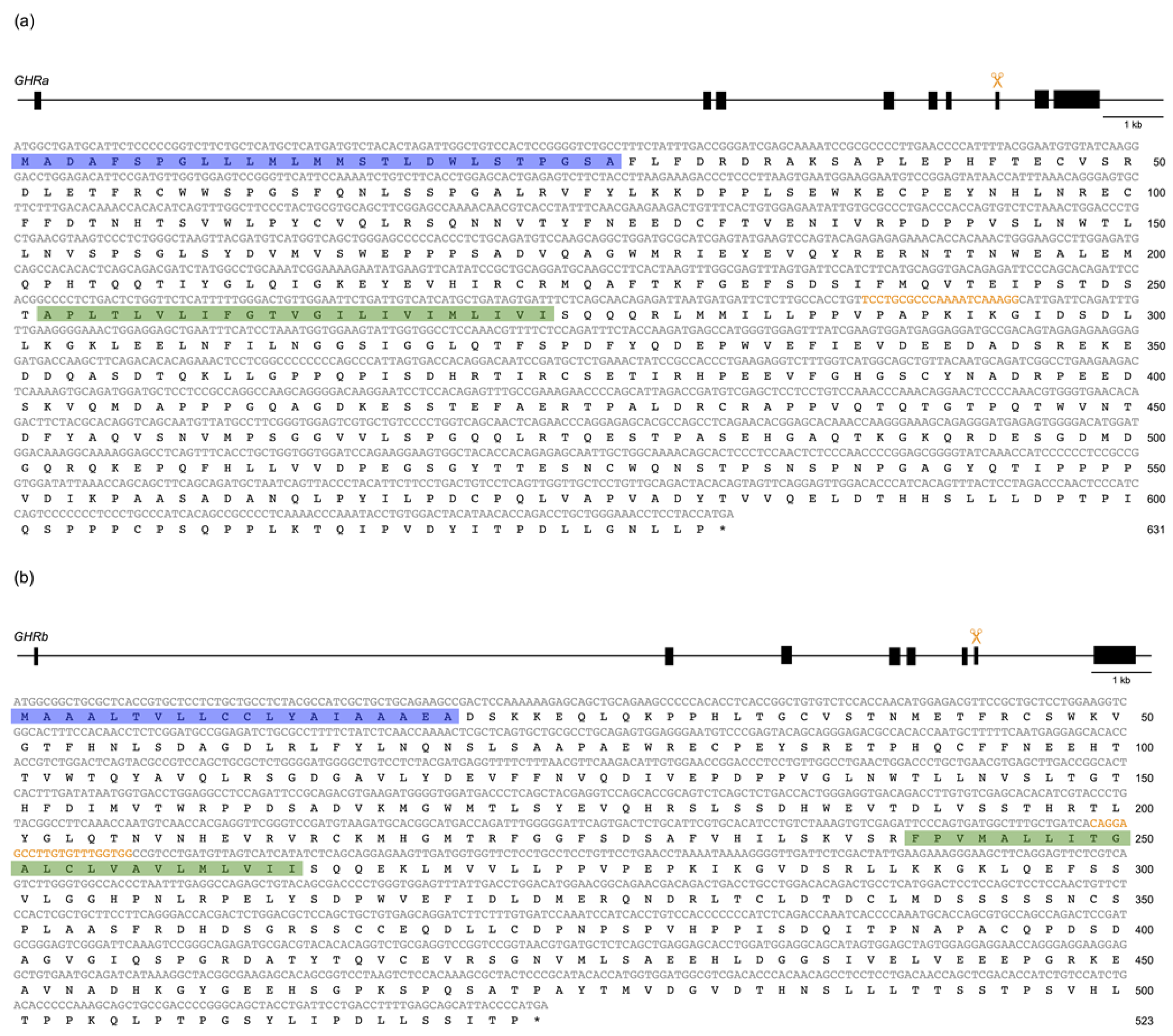
Figure 1.
Genomic structure of the somatolactin alpha receptor (SLR: GHRa) (a) and growth hormone receptor (GHR: GHRb) (b) genes (GenBank accession numbers: DQ002886 and DQ010539, respectively). The black boxes represent translated regions (the 5′/3′ UTRs are not shown). The orange scissors indicate the approximate positions of the target sequences in CRISPR/Cas9. The cDNA and amino-acid sequences are shown in gray and black, respectively. The target sequences are shown in orange. The signal peptides predicted by SignalIP 5.0 and the transmembrane regions predicted by TMHMM 2.0 are highlighted in blue and green, respectively.
Figure 1.
Genomic structure of the somatolactin alpha receptor (SLR: GHRa) (a) and growth hormone receptor (GHR: GHRb) (b) genes (GenBank accession numbers: DQ002886 and DQ010539, respectively). The black boxes represent translated regions (the 5′/3′ UTRs are not shown). The orange scissors indicate the approximate positions of the target sequences in CRISPR/Cas9. The cDNA and amino-acid sequences are shown in gray and black, respectively. The target sequences are shown in orange. The signal peptides predicted by SignalIP 5.0 and the transmembrane regions predicted by TMHMM 2.0 are highlighted in blue and green, respectively.

Figure 2.
Cladogram of the GHR sequences registered at the RefSeq protein database. A total of 222 sequences (136 species) were used for phylogenetic tree reconstruction using the maximum-likelihood method. Sequences classified in the database as GHR, GHRa, GHRb, GHR-like, or other are colored in purple, red, blue, green, or gray, respectively. Bootstrap values are shown at each node, and nodes with a bootstrap value of 90% or more are labeled by black dots. Three groups (GHR of tetrapods, lungfish, sturgeon, and polypterus [purple]; GHRa [red] of teleosts; and GHRb [blue] of teleosts) are supported by bootstrap values of 100%. This figure revealed many problems in the nomenclature of GHRs.
Figure 2.
Cladogram of the GHR sequences registered at the RefSeq protein database. A total of 222 sequences (136 species) were used for phylogenetic tree reconstruction using the maximum-likelihood method. Sequences classified in the database as GHR, GHRa, GHRb, GHR-like, or other are colored in purple, red, blue, green, or gray, respectively. Bootstrap values are shown at each node, and nodes with a bootstrap value of 90% or more are labeled by black dots. Three groups (GHR of tetrapods, lungfish, sturgeon, and polypterus [purple]; GHRa [red] of teleosts; and GHRb [blue] of teleosts) are supported by bootstrap values of 100%. This figure revealed many problems in the nomenclature of GHRs.

Figure 3.
Electropherograms of target sequences and frameshift mutations in SLR and GHR. The target sequences (highlighted in orange) for the wild-type (WT) fish are shown at the top for SLR (a) and GHR (b). Corresponding nucleotides in the WT and mutant fish are marked by colored circles. Four bases (CAAA) or five bases (TCAAA) are deleted in slr−4 and slr−5, respectively. One base (C or T) is inserted or deleted in ghr+1 or ghr−1, respectively.
Figure 3.
Electropherograms of target sequences and frameshift mutations in SLR and GHR. The target sequences (highlighted in orange) for the wild-type (WT) fish are shown at the top for SLR (a) and GHR (b). Corresponding nucleotides in the WT and mutant fish are marked by colored circles. Four bases (CAAA) or five bases (TCAAA) are deleted in slr−4 and slr−5, respectively. One base (C or T) is inserted or deleted in ghr+1 or ghr−1, respectively.

Figure 4.
PT-PCR for three adults, each among WT, ghr+1, and ghr−1 fish. The upper lanes, middle lanes, and bottom lanes show the expression of beta-actin (22 cycles), SLR (28 cycles), and GHR (28 cycles), respectively. The transcripts of GHR were not detected in all three ghr−1 individuals, whereas these bands were observed in all WT and ghr+1 individuals. Bands from RT-PCR products for beta-actin and SLR were detected in all individuals from all strains.
Figure 4.
PT-PCR for three adults, each among WT, ghr+1, and ghr−1 fish. The upper lanes, middle lanes, and bottom lanes show the expression of beta-actin (22 cycles), SLR (28 cycles), and GHR (28 cycles), respectively. The transcripts of GHR were not detected in all three ghr−1 individuals, whereas these bands were observed in all WT and ghr+1 individuals. Bands from RT-PCR products for beta-actin and SLR were detected in all individuals from all strains.

Figure 5.
Body length at the adult and larval stages. (a) Body length of adult fish among genotypes in ghr−1 (blue dots), slr−4 (red dots), and slr−5 (purple dots) individuals. (b) Transition of body length among genotypes during the larval stage in ghr−1 (blue dots), ghr+1 (green dots), and slr−4 (red dots) individuals. Genotypes are indicated as +/+ (WT fish), +/− (heterozygotes), and −/− (homozygotes). Each dot represents the body length of each individual, and a closed box with bars indicates the mean and standard error.
Figure 5.
Body length at the adult and larval stages. (a) Body length of adult fish among genotypes in ghr−1 (blue dots), slr−4 (red dots), and slr−5 (purple dots) individuals. (b) Transition of body length among genotypes during the larval stage in ghr−1 (blue dots), ghr+1 (green dots), and slr−4 (red dots) individuals. Genotypes are indicated as +/+ (WT fish), +/− (heterozygotes), and −/− (homozygotes). Each dot represents the body length of each individual, and a closed box with bars indicates the mean and standard error.

Figure 6.
Comparison of body color. (a) Example of the images used for counting chromatophores. (b) Chromatophore densities in adult medaka. The numbers of melanophores (black dots), xanthophores (orange dots), and leucophores (gray dots) in ghr−1, slr−4, and slr−5 adult fish examined twice in two different batches are shown. Genotypes are indicated as +/+ (WT fish), +/− (heterozygotes), and −/− (homozygotes). A dot represents the chromatophore density of each individual and a closed box with bars indicates the mean and standard error.
Figure 6.
Comparison of body color. (a) Example of the images used for counting chromatophores. (b) Chromatophore densities in adult medaka. The numbers of melanophores (black dots), xanthophores (orange dots), and leucophores (gray dots) in ghr−1, slr−4, and slr−5 adult fish examined twice in two different batches are shown. Genotypes are indicated as +/+ (WT fish), +/− (heterozygotes), and −/− (homozygotes). A dot represents the chromatophore density of each individual and a closed box with bars indicates the mean and standard error.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
gRNA target sequences and mutagenesis efficiency for the SLR and GHR genes in G0 fish.
| gRNA | Target Sequence (5′→3′) | Eggs Injected (N) | Survivors on the Day Following Injection (N) | Survivors on the Day Following Injection (%) | Normal Development (N) | Larvae Subjected to HMA (N) | Larvae Confirmed to Have a Band Shift (N) | Mutagenesis Efficiency (%) |
|---|---|---|---|---|---|---|---|---|
| SLR5 | CCAGAGTCAGAGGGGCCGTGG | 40 | 16 | 40.0 | 15 | 15 | 0 | 0 |
| SLR6 | TCTCATTTTTGGGACTGTTGG | ― | ― | ― | ― | ― | ― | ― |
| SLR7 | TTTGGGCGCAGGAACAGGTGG | ― | ― | ― | ― | ― | ― | ― |
| SLR8 | TCCTGCGCCCAAAATCAAAGG | 29 | 15 | 51.7 | 14 | 14 | 5 | 35.7 |
| GHR5 | GATCAGCAAAGCCATCACTGG | 143 | 35 | 24.5 | 11 | 11 | 0 | 0 |
| GHR6 | CAGGAGCCTTGTGTTTGGTGG | 127 | 48 | 37.8 | 17 | 16 | 4 | 25.0 |
| GHR7 | GGTTCAGGAACAGGAGGCAGG | 35 | 20 | 57.1 | 19 | 13 | 0 | 0 |
| GHR8 | TCCTGAACCTAAAATAAAAGG | 22 | 15 | 68.2 | 15 | 12 | 0 | 0 |
SLR8 and GHR6, which were finally selected as gRNAs for the SLR and GHR genes, respectively, are highlighted in bold.
Table 2.
Potential off-targets of the GHR6 gRNA in the Hd-rR genome.
| gRNA | Potential Off-Target Sequence (5′→3′) | Chromosome |
|---|---|---|
| GHR6 | CAGGAGCCTTGTGTTTGGTGG | 12 (target) |
| CAGGAGACTTGTGTTTGTGGG | 1 | |
| CAGGAGCCATGTTTTTGGGGG | 12 | |
| GAGCAGCCTTGTGTTTGAGGG | 19 |
Table 3.
Ins/del mutations in SLR and GHR inherited by the F1 generation from the G0 fish.
| Target Gene | G0 Individual Number | Mutation Type | Mutation | Sequence (5′→3′) | Number of F1 Larvae with Mutations (%) | Number of F1 Adults with Mutations (%) |
|---|---|---|---|---|---|---|
| SLR | - | WT | - | GCCCAAAATCAAAGGCATTGATTCAGATTTGTTGAAG | - | - |
| 1 | six-deletion | SLR-6 (1) | GCCCAAAATCA––––––TTGATTCAGATTTGTTGAAG | 3 (18.8) | 13 (13.1) | |
| 2 | one-deletion | SLR+3-4 | GCCCAAAA–CAGAGGCATTGATTCAGATTTGTTGAAG | 1 (6.3) | 0 (0) | |
| 4 × 7 | nine-insertion | SLR+9 | GCCCAAAAATCAATGAATCAAAGGCATTGATTCAGATTTGTTGAAG | 2 (12.5) | 0 (0) | |
| 4 × 7, 6 | five-deletion | SLR-5 | GCCCAAAA–––––GGCATTGATTCAGATTTGTTGAAG | 4 (18.8, 6.3) | 5 (3.9) | |
| 4 × 7, 8 | six-deletion | SLR-6 (2) | GCCCAAA––––––GGCATTGATTCAGATTTGTTGAAG | 2 (12.5) | 3 (1.7) | |
| 5, 6 | two-insertion | SLR+2 (1) | GCCCAAACAATCAAAGGCATTGATTCAGATTTGTTGAAG | 1 (6.3) | 1 (0.8) | |
| 6 | sixteen-insertion | SLR+17-1 | GCCCAAAAGAGCCTGAAACGATGTCCAAAGGCATTGATTCAGATTTGTTGAAG | 0 (0) | 3 (2.4) | |
| 6 | four-deletion | SLR-4 | GCCCAAAAT––––GGCATTGATTCAGATTTGTTGAAG | 1 (6.3) | 9 (7) | |
| 6 | three-deletion | SLR-3 | GCCCAAAA–––AAGGCATTGATTCAGATTTGTTGAAG | 0 (0) | 3 (2.3) | |
| 6 | two-insertion | SLR+2 (2) | GCCCAAAATGCCAAAGGCATTGATTCAGATTTGTTGAAG | 1 (6.3) | 9 (7) | |
| 6 | two-deletion | SLR-2 | GCCCAAAA––AAAGGCATTGATTCAGATTTGTTGAAG | 0 (0) | 2 (1.6) | |
| 8 | seven-deletion | SLR+6-13 | GCCCAAAGGTCTT–––––––ATTCAGATTTGTTGAAG | 0 (0) | 1 (0.6) | |
| 8 | six-insertion | SLR+7-1 | GCCCAAAAATGGGCCCAAAGGCATTGATTCAGATTTGTTGAAG | 2 (12.5) | 4 (2.3) | |
| 8 | one-insertion | SLR+1 | GCCCAAAATTCAAAGGCATTGATTCAGATTTGTTGAAG | 2 (12.5) | 29 (16.5) | |
| Total | 19 | 82 | ||||
| GHR | - | WT | - | GCCTTGTGTTTGGTGGCCATCCTGATGTTAGTCATCA | - | - |
| 1 | one-insertion | GHR+1 | GCCTTGTGTTCTGGTGGCCATCCTGATGTTAGTCATCA | 1 (6.3) | 4 (4) | |
| 4 × 7, 8 | one-deletion | GHR-1 | GCCTTGTGTT–GGTGGCCATCCTGATGTTAGTCATCA | 3 (12.5, 6.3) | 5 (2.8) | |
| 6 | six-insertion | GHR+6 | GCCTTGTGTGGTGCCTTGGTGGCCATCCTGATGTTAGTCATCA | 1 (6.3) | 7 (5.5) | |
| 8 | two-deletion | GHR-2 | GCCTTGTGT––GGTGGCCATCCTGATGTTAGTCATCA | 0 (0) | 2 (1.1) | |
| Total | 5 | 18 |
The wild-type (WT) sequence is shown at the top for each gene. Three G0 individuals, #1, #6, and #8, who were subjected to pass the mutations to F1 adult fish are highlighted in bold. Inserted or deleted nucleotides are shown as red uppercase letters or red hyphens, respectively. The mutants were named based on the number of ins/del mutations in SLR and GHR (e.g., slr−5 means that five nucleotides were deleted from WT SLR). Four mutations, slr−5, slr−4, ghr+1, and ghr−1, were subjected to phenotyping and are highlighted in bold.
Table 4.
Viability of slr−4, slr−5, ghr+1, and ghr−1 among the genotypes of F2 adult fish.
| Strain | Genotypes | ||
|---|---|---|---|
| +/+ | +/− | −/− | |
| slr–4 | 75 | 114 | 0 |
| 0.000 | |||
| slr–5 | 18 | 33 | 4 |
| 0.009 | |||
| ghr+1 | 12 | 24 | 9 |
| 0.741 | |||
| ghr–1 | 32 | 68 | 28 |
| 0.687 | |||
Genotypes are indicated as +/+ (WT fish), +/− (heterozygotes), and −/− (homozygotes).
Table 5.
Viability of ghr+1, ghr−1, and slr−4 F2 larval fish according to genotype.
| Strain | Age (weeks) and Genotype | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 1 | 2 | 3 | 4 | |||||||||||
| +/+ | +/− | −/− | +/+ | +/− | −/− | +/+ | +/− | −/− | +/+ | +/− | −/− | +/+ | +/− | −/− | |
| ghr+1 | 7 | 7 | 1 | 5 | 12 | 2 | 0 | 4 | 4 | 10 | 7 | 3 | 4 | 16 | 8 |
| 0.088 | 0.323 | 0.135 | 0.035 | 0.424 | |||||||||||
| ghr–1 | 7 | 7 | 7 | 0 | 0 | 0 | 3 | 4 | 1 | 5 | 4 | 4 | 0 | 3 | 3 |
| 0.311 | ― | 0.607 | 0.354 | 0.223 | |||||||||||
| slr–4 | 10 | 18 | 9 | 8 | 18 | 15 | 9 | 6 | 2 | 15 | 27 | 5 | 4 | 11 | 2 |
| 0.960 | 0.223 | 0.027 | 0.071 | 0.379 | |||||||||||
Upper line: number of surviving individuals. Lower line: p value according to the chi-squared test. Genotypes are indicated as +/+ (WT fish), +/− (heterozygotes), and −/− (homozygotes).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Moroki, Y.; Komori, M.; Ogawa, Y.; Nagumo, E.; Ohno, H.; Fukamachi, S. An Attempt to Identify the Medaka Receptor for Somatolactin Alpha Using a Reverse Genetics Approach. Genes 2023, 14, 796. https://doi.org/10.3390/genes14040796
AMA Style
Moroki Y, Komori M, Ogawa Y, Nagumo E, Ohno H, Fukamachi S. An Attempt to Identify the Medaka Receptor for Somatolactin Alpha Using a Reverse Genetics Approach. Genes. 2023; 14(4):796. https://doi.org/10.3390/genes14040796
Chicago/Turabian StyleMoroki, Yuko, Mamiko Komori, Yuko Ogawa, Erina Nagumo, Haruna Ohno, and Shoji Fukamachi. 2023. "An Attempt to Identify the Medaka Receptor for Somatolactin Alpha Using a Reverse Genetics Approach" Genes 14, no. 4: 796. https://doi.org/10.3390/genes14040796
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.