
Social Diversification Driven by Mobile Genetic Elements

Department of Molecular Biology, University of Wyoming, Laramie, WY 82071, USA
*
Author to whom correspondence should be addressed.
Genes 2023, 14(3), 648; https://doi.org/10.3390/genes14030648
Submission received: 9 January 2023
/
Revised: 17 February 2023
/
Accepted: 28 February 2023
/
Published: 4 March 2023
(This article belongs to the Special Issue The Genetic Basis of Microbial Sociality)
Abstract
:Social diversification in microbes is an evolutionary process where lineages bifurcate into distinct populations that cooperate with themselves but not with other groups. In bacteria, this is frequently driven by horizontal transfer of mobile genetic elements (MGEs). Here, the resulting acquisition of new genes changes the recipient’s social traits and consequently how they interact with kin. These changes include discriminating behaviors mediated by newly acquired effectors. Since the producing cell is protected by cognate immunity factors, these selfish elements benefit from selective discrimination against recent ancestors, thus facilitating their proliferation and benefiting the host. Whether social diversification benefits the population at large is less obvious. The widespread use of next-generation sequencing has recently provided new insights into population dynamics in natural habitats and the roles MGEs play. MGEs belong to accessory genomes, which often constitute the majority of the pangenome of a taxon, and contain most of the kin-discriminating loci that fuel rapid social diversification. We further discuss mechanisms of diversification and its consequences to populations and conclude with a case study involving myxobacteria.
1. Introduction
Evolution is a story of diversification. It essentially involves changes in lineage genomes that may eventually result in new taxa. Different selective forces drive diversification and impactful changes, made by mutations, recombination or horizontal gene transfer (HGT), are maintained provided there is a fitness benefit [1]. Environmental factors, such as spatial separation, distinct ecological niches, competition and other forces, including genetic drift, promote diversification. Central to this is the formation of new social groups, where individuals within a group cooperate, but not with those that belong to other groups. Social diversification in turn can lead to recombination barriers that ultimately facilitate the evolution of distinct taxa.
Social groups consist of cooperative interactions between related individuals that lead to fitness gains, which are often not attainable by individuals [2,3]. However, when cooperation is blocked, and/or antagonism is initiated, diversification ensues. In microbes, social diversification is directly linked to gene content changes. In bacteria, this is frequently caused by HGT. This review focuses on the role of HGT in social diversification and, in particular, how mobile genetic elements (MGEs) serve as powerful engines of diversification. Here, the acquisition of new genes, or their loss, results in cells that no longer productively cooperative with their recent siblings. Instead, such diversification typically leads to antagonism between related individuals, herein broadly defined as kin discrimination (KD). In turn, social conflict itself can lead to accelerated evolution manifested as arms races or Red Queen evolution [4]. These arms races likely explain why many bacterial genomes contain a large number of KD genes [5,6,7,8,9,10,11]. Tied to diversification, we also discuss the dynamic nature of bacterial genomes and mechanisms that govern KD.
1.1. Intraspecific Genome Diversity and Fluidity
The dynamic nature of prokaryotic genomes typically results in an exceptionally high degree of intraspecific diversity. This diversity is readily apparent when comparing genomes within a species and assessing pangenome content, which represents all genes found within a taxon [12]. In most cases, pangenomes mostly consist of accessory genes that are present in only a subset of lineages. For some species, their core genomes, genes found in all isolates, only represent <20% of their pangenome [12,13,14]. Many of these accessory genes are derived from MGEs, which themselves can represent >25% of a genome [15]. As highlighted here, MGEs play key roles in social diversification, in part because they are the most dynamic class of genes in prokaryotes [16].
Although bacterial genomes are fluid, where new genes are added by HGT, the sizes of genomes within a species typically remain relatively constant [17]. This is partly explained by gene addition being countered by gene loss. More broadly, in the absence of HGT and selective pressures, the prevailing mode of change is by genome contraction [18], where the compact nature of bacterial genomes is caused by a mutational bias toward deletions [14,19]. In fact, the rate of deletion to insertion is favored about 10:1 [20], and thus DNA that does not benefit the host is lost over time, as dramatically revealed by the small genomes of many obligate intracellular parasites [19]. Additionally, microbes can adapt to environmental changes by gene loss, leading to the ‘less-is-more hypothesis’ [21,22]. In support of this, a meta-analysis of studies on E. coli MG1655 grown in 144 different environmental conditions determined that adaptive null mutations had occurred in 139 of the 144 conditions [23]. Strikingly, under nutrient deprivation, adaptive null mutations were found that nearly doubled the growth rate [22,23].
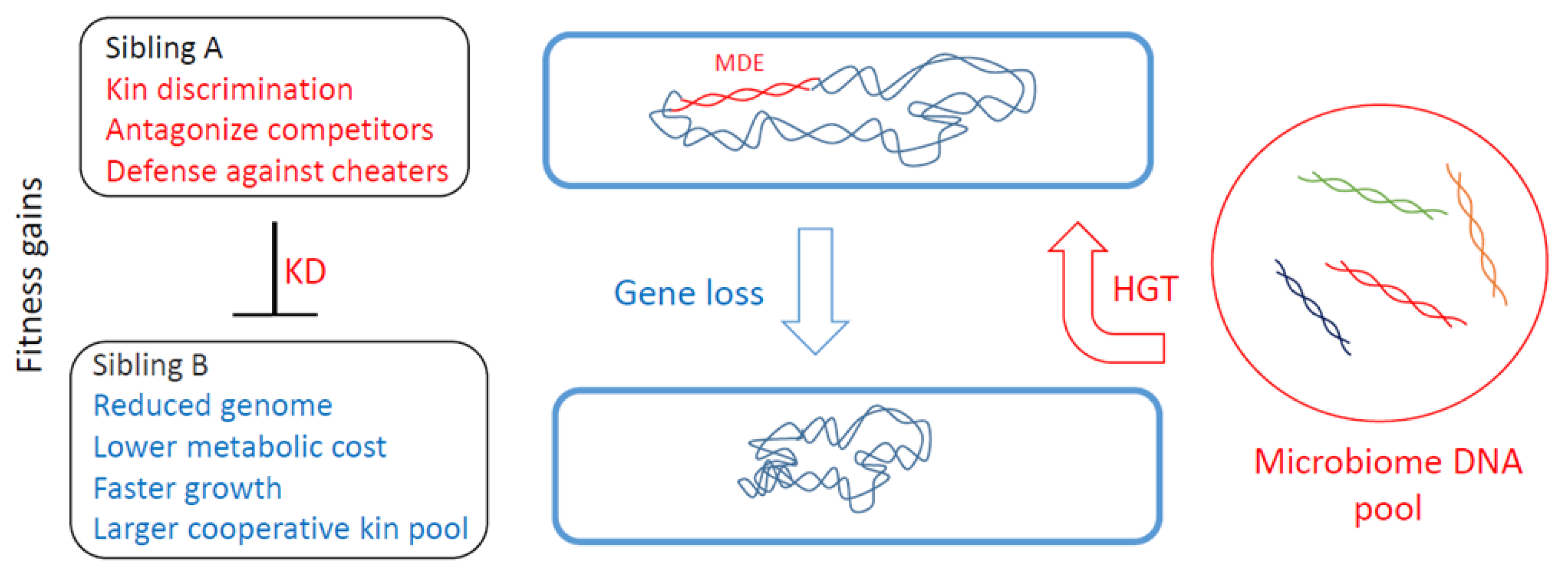
Although MGEs are selfish entities that replicate for their own gain, they also carry traits that impact the host. Many of these traits influence how the host interacts with others, including means for KD, which we describe as mobile discriminating elements (MDEs). Genes that govern interactions with kin are by definition social genes, and the constellation of social genes determines the degree of relatedness between individuals [3], as expressed by Hamilton’s rule (see below) [24]. Thus, the acquisition or loss of MDEs changes an individual’s social relationships and overall fitness (Figure 1). Moreover, the acquisition of a new MDE can confer an adaptive benefit whereby the individual discriminates against siblings and thus creates a new social group (Figure 2). In these scenarios, the new discriminatory behavior might be driven by a single locus, e.g., a toxin-immunity cassette. In such cases, this behavior can be called kind or greenbeard discrimination to highlight its genetic simplicity [2]. However, in many cases, other loci also contribute towards KD, such as genes required for toxin delivery (Figure 3).
1.2. Benefits of Kin Discrimination
Determining genetic relatedness between individuals allows microbes to form sustainable social interactions. That is, individuals need to discern close genetic relatives or siblings that cooperate with themselves from distant microbes with similar metabolic needs that are competitors or social cheaters. Although microorganisms are often thought of as autonomous unicellular organisms, cooperative and even altruistic behaviors are abundant in microbial populations. These behaviors range from secreting a shared public good to self-sacrifice, e.g., autolysis. Evolutionary theory and experimental work have addressed how social behaviors evolve, given a seemingly contradictory Darwinian framework that individuals only behave in their own self-interests. Hamilton [24] proposed that cooperative and altruistic behaviors evolve when they are directed toward close kin that share social genes. Hamilton’s rule states that rb > c, where r is the genetic relatedness between the actor and the recipient, b is the benefit to the actor, and c is the cost to the actor. Thus, an altruistic behavior that has a fitness cost to the actor can evolve if it provides a fitness benefit to a sufficient number of close relatives that share social alleles. This rule was summarized by evolutionary biologist JBS Haldane’s comment that he would not lay down his life for a brother, but would for two brothers or eight cousins [25]. In support of Hamilton’s rule, a meta-analysis of twelve studies found that altruism is often under positive selection [26]. Since altruistic behavior must be directed towards close kin to evolve, organisms need mechanisms to determine their genetic relatedness to other individuals.
In many cooperative systems, social cheating arises. A cheater is an individual that reaps the benefit of a cooperative action but incurs reduced or no cost to itself. If cheaters expand in a population, it leads to the loss of the cooperative trait and collapse of the social system [27,28,29]. Similarly, within multicellular organisms, such as animals, cancer cells cheat by exploiting their cooperative cellular environments, which often leads to the demise of the individual. Examples of cheaters in biofilms are individuals that that use public secreted goods produced by others, but they do not contribute themselves. Cheaters also arise in aggregative multicellular organisms, such as myxobacteria and social slime molds. In myxobacteria, >60% of cells lyse during fruiting-body development, while less than 20% form viable spores [30]. The lysed cells apparently represent an altruistic behavior that provides nutrients to the starving population and consequently supports the metabolically demanding process of spore formation, which occurs over days [31]. However, cheaters exploit this cooperative environment and become disproportionately represented in spores and thus do not undergo their fair share of lysis [31,32,33,34,35,36]. Similarly, in Dictyostelium discoideum fruiting bodies, cheaters arise in which they are over-represented among spores, and under-represented in terminally differentiated stock cells [37,38,39]. Because there are inherit selective advantages for cheater cells to arise, species need mechanisms to protect their cooperative traits from exploitation. To do so, organisms use self-recognition and/or KD systems [40,41].
As mentioned, secreted goods are particularly vulnerable to cheating, because once they leave the producer, any neighboring cell can exploit them. An alternative strategy is for cells to share goods privately. An example of shared private goods comes from outer membrane exchange (OME) in myxobacteria [42]. In this system, individuals recognize one another through homotypic binding between compatible (e.g., identical) TraA receptors, which are highly polymorphic and hence provide specificity for kin recognition [43,44]. After receptor binding, cells exchange portions of their outer membranes and proteins by an apparent transient membrane fusion event(s) [45]. This behavior is hypothesized to evolve as a mechanism to protect cooperative interactions.
1.3. KD Mechanisms Are Plentiful and Diverse
Given the importance of KD, it is not surprising that these systems are widespread and diverse throughout the bacterial kingdom [2,3,7,46,47,48]. Here we give a brief overview of antagonism- and non-antagonism-based KD systems. Although many of the examples given are associated with MGEs, there are also many cases where KD loci are separate from MGEs.
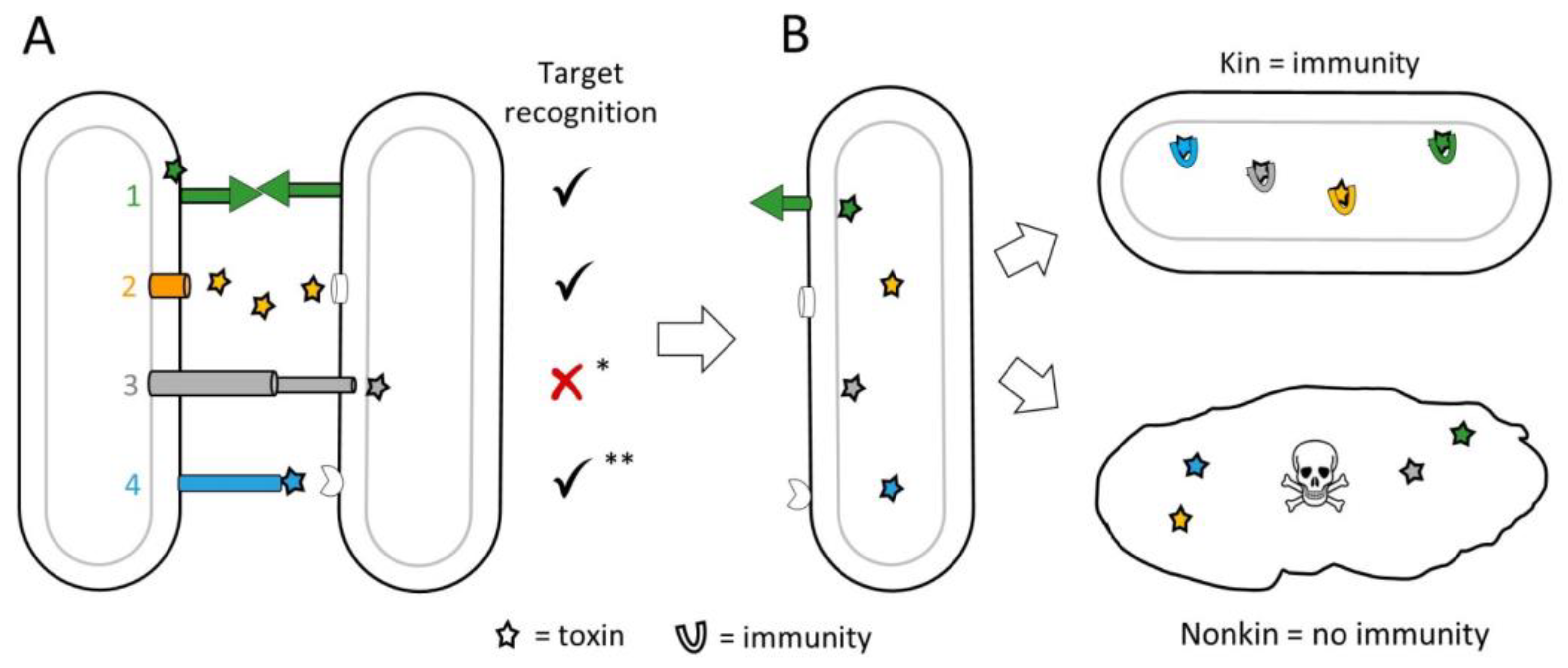
KD systems frequently contain toxin-immunity gene cassettes where an individual delivers effectors to neighboring cells, which only have immunity if they are related or clonal cells (Figure 3). In the case of myxobacteria, which display extensive multicellular and cooperative interactions [49], they use multi-layered KD systems. As described, the first level of kin recognition is mediated by binding of compatible TraA receptors that trigger OME. During OME, subsets of the exchanged cargo are lipoprotein effectors known as SitA toxins. These toxins function in KD, because only related individuals with compatible TraA receptors receive them [50]. After delivery to the outer membrane, SitA toxins employ their ‘escort domain’ to hijack inner membrane proteins to enter the cytoplasm where they act as nucleases [51]. sitAI toxin-immunity gene cassettes are widespread, where some myxobacteria contain >80 loci. In turn, this allows each strain to have its own exquisite ‘self-identity barcode’ consisting of a suite of unique toxin-immunity loci [52]. The exchange of SitA toxins thus serves as a second layer or a verification step for KD [53].
Since OME only functions in KD between strains with compatible TraA receptors [44,50,54], other systems are required. Indeed, when testing environmental Myxococcus xanthus isolates [5], the elimination of OME by inactivating traA did not prevent antagonism. Instead, to eliminate antagonism, double mutants that knocked out OME and the type VI secretion systems (T6SS) were required, and for two strains, a third system (Rhs) had to be knocked out. This revealed that M. xanthus uses parallel KD systems to deliver discriminating effectors. As found with sitAI, comparative genomic analysis found divergent T6SS and Rhs effector loci in an MGE called Mx-α, a polymorphic lineage of prophages [50,54,55]. Additionally, Gong et al. [56,57] identified T6SS effectors involved in colony merger incompatibility (KD) between isogenic M. xanthus strains. Thus, in one context, T6SS in M. xanthus allows KD against myxobacteria of variable degrees of relatedness, while OME allows KD against closely related individuals that share compatible TraA receptors.
Interestingly, the T6SS also discriminates against siblings that are physiologically less fit [58]. For example, in M. xanthus, when a histidine auxotroph is mixed with its isogenic parent, the parent kills the auxotroph in the absence of histidine, but not in the presence of histidine. Here, a phylogenetically conserved T6SS effector mediates sibling antagonism. Mechanistically, antagonism occurs because the level of the immunity protein decreases during histidine starvation, and thus the indiscriminate injection of this T6SS effector discriminates against less fit siblings.
Similarly, Bacillus subtilis uses a combination of KD mechanisms. Here, Lyons et al. [6] proposed that surface receptors, contact-dependent inhibition (CDI) effectors, secreted compounds and mobile elements all function in KD. Kraigher et al. [59] further investigated KD of environmental isolates at a centimeter scale, and found that when closely related strains were mixed, they swarmed together to colonize a surface. However, when genetically divergent strains were mixed, one strain outcompeted the other and colonized the surface alone. In a separate study, Lyons & Kolter [60] investigated Bacillus interactions from a broader relatedness scale, i.e., between Bacillus species. They found Bacillus cells discerned the degree of relatedness, where strong levels of antagonism occurred between closely related species, and low levels of antagonism occurred between distant isolates. Nevertheless, antagonism occurred against distantly related species that produced the same surfactant molecule, an exploitable common good.
In contrast to cell-contact-dependent discrimination, many organisms deploy diffusible discrimination factors. For example, Vacheron et al. [61] found that Pseudomonas protegens produce R-tailocins, which, following lysis of the producing cell, are dispersed and specifically target competing cells by puncturing their cell membranes. Other diffusible KD factors commonly found on MGEs include bacteriocins, colicins and antibiotics [2,3,62,63,64].
KD also occurs in the absence of lethal toxin delivery. It has long been recognized that when swarms of divergent strains of Proteus mirabilis meet, a demarcation zone appears [65]. Genetic analysis determined that the six-gene ids locus is responsible for this inhibition [66]. Subsequent studies determined that two cognate proteins, IdsD and IdsE, must interact for kin recognition. IdsD is delivered by T6SS, and if cells are kin, IdsD interacts with IdsE in the recipient, and the effector is neutralized. If cells are nonkin, the cognate IdsE is absent, allowing unbound IdsD to increase levels of the secondary signaling molecule ppGpp, which results in transcriptional changes and decreased swarming. Thus, nonkin cells drop out of a swarm, ensuring that a swarm is composed only of cooperative kin cells [67,68]. However, in other P. mirabilis strains, T6SS serves in antagonism delivering lethal toxins [69]. In another example of non-lethal KD, environmental isolates of Vibrio cholerae harboring differing variants of the PilA pilin lacked the ability to aggregate, and thus formed distinct social groups [70].
1.4. MGEs Harbor Diverse Types of KD Systems
Many KD loci are encoded on MGEs (Table 1). Perhaps the best-known examples are colicin toxins encoded on Escherichia coli plasmids (reviewed in [71]), which are secreted into the milieu and kill E. coli cells lacking that MGE. Additionally, toxin-delivery systems are also found on plasmids. For instance, Morgado and Vicente [72] identified 330 plasmids, primarily in the Proteobacteria, that encode T6SS genes. T6SS genes are also encoded on integrative conjugative elements (ICEs) and genomic islands, suggesting HGT [72,73]. Recently, a T6SS operon in Vibrio fischeri was found that functions in KD [74] and contains a putative lipoprotein that specifies target cells [75].
Lysogenic phage are another type of vehicle that disseminates KD loci and thus serve as catalysts for social diversification. For instance, MuF toxins are associated with phages prevalent in the human gut microbiome [76]. In E. coli, the formation of colony demarcations, i.e., KD, was found when one strain contains a prophage and the other does not [81]. Here, a sub-population of cells lyse and release phages, resulting in KD against siblings. In M. xanthus, sitAI and T6SS effector loci are frequently located on prophages and thus discriminate against related strains that lack immunity [5,54]. Additionally, HGT by prophages was proposed to disseminate KD genes in B. subtilis [6].
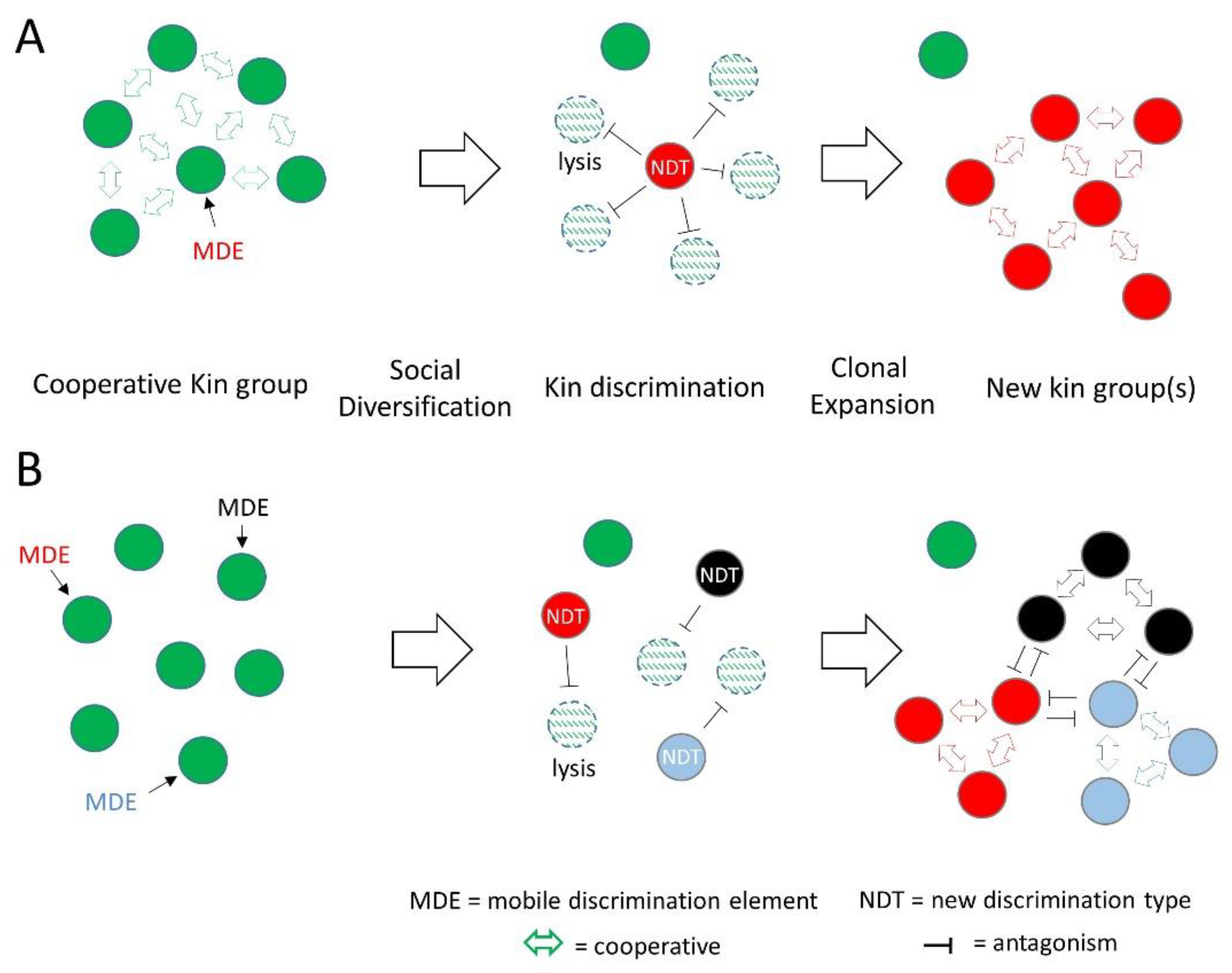
CDI systems are widely distributed in several classes of Proteobacteria where they antagonize close kin. In some cases, CDI genes are located in or near transposable element genes, suggesting horizontal transfer [77,80], and are also stably maintained in plasmids that become fixed in populations [82]. In Bacteroides and Parabacteroides residing in the human gut, ICE families were found, which contained T6SS operons and, strikingly, were horizontally transferred within a person and between related species [78,79]. Moreover, the horizontal transfer of these elements led to fixation of particular strains in at least eight cases in two human gut ecosystems [79]. These findings illustrate how the transfer of MDEs can rapidly change kin groups in an ecosystem, as shown in Figure 2.
1.5. Do MGEs Promote Cooperation?
An intriguing and debated idea is whether MGEs also promote social cooperation [83]. Some argue that cooperation, i.e., public-good sharing, is promoted by HGT, which can also act as a mechanism to guard against cheating. For example, when a secreted good is encoded on an MGE, a cheater that does not secrete the compound, but benefits and co-exists in the population, becomes a likely target for infection by that MGE, thus preventing a cheater from overtaking a population [83,84]. Furthermore, if KD loci are carried on that MGE, it acts as a policing mechanism to punish non-cooperators [83]. These ideas have sparked debates, which are primarily centered on secreted virulence factors encoded on plasmids [83,85,86,87,88].
Nogueira et al. [85] used modeling to show how horizontal transfer promotes cooperation. In support of their model, they analyzed 20 E. coli genomes and their plasmids and found that proteins that are secreted or localized in the outer membrane are more likely to be encoded on mobile elements and transfer hotspots. Likewise, other studies concluded that genes encoding secreted goods are overrepresented on plasmids [86]. In contrast, Dewar et al. [88] analyzed 1,632 genomes from 51 species and did not find support of this idea. Instead, they concluded that transfer of MGEs may promote cooperation in the initial invasion of a population, but not for their maintenance.
If HGT is not sufficient to maintain cooperation, then how do these elements persist? One solution comes from the Black Queen Hypothesis [89]. This hypothesis predicts genes that produce costly beneficial public goods will be lost by most individuals, but are retained in just enough individuals to support the population, resulting in dependency between individuals. Put another way, the Black Queen Hypothesis can be viewed as a ‘division of labor’ [83].
Besides plasmids, other MGE types contribute to cooperation, though less is known [83]. In one case, the spread of beneficial genomic islands (GIs) through an aquaculture community was found [90]. By using pangenome sequencing of Vibrio parahaemolyticus, GIs encoding virulence genes and antibiotic resistance genes were discovered. Some of these GIs were also found in other Vibrio taxa that interact positively with V. parahaemolyticus, suggesting a cooperative function mediated by HGT.
1.6. Does Social Diversification Benefit Bacterial Populations at Large?
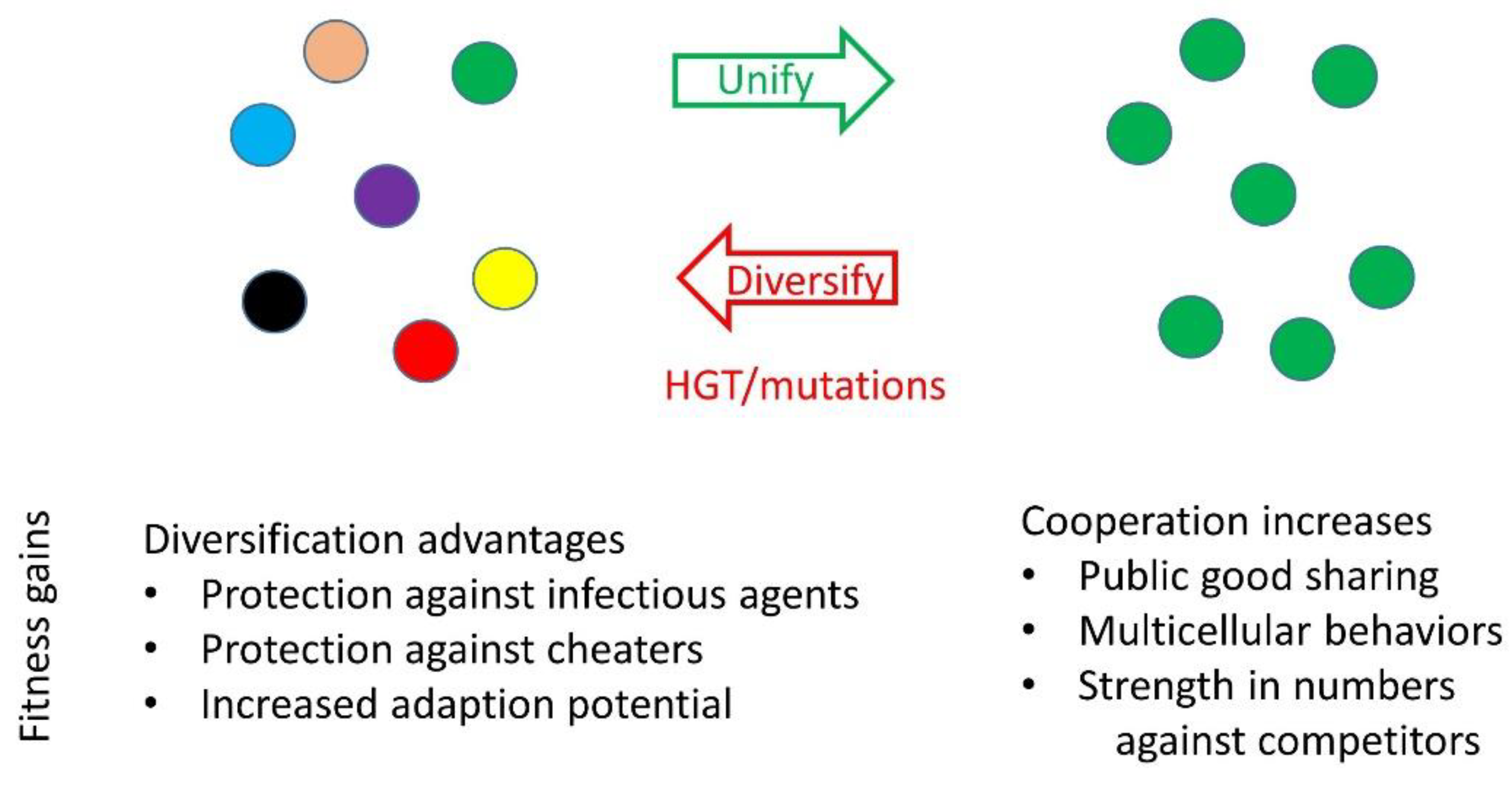
Successful MGEs provide a fitness benefit to the host, which in turn allows the element to propagate. For MDEs, both the element and the host benefit, but they do so at the expense of the host’s siblings (Figure 2). This leads to an underlying question: Does such social diversification benefit populations at large, or do they simply tolerate the stress? On the one hand, KD results in elevated infighting among kin and hence reduces a population’s size and fitness, making it vulnerable to larger and more cooperative competitor populations (Figure 4). Furthermore, considering competitive exclusion theory, one predicts that following diversification, the new lineages should not coexist indefinitely since they occupy the same niche [91]. In some environments, this may occur [79], while in other environments this may not happen because of the dynamics and diversity of MDE transfer [5].
On the other hand, diversification offers benefits (Figure 4). Empirically, when examined, new KD loci are often found in populations [5,6], thus highlighting the success of the elements and apparent fitness gains by hosts. For populations at large, diversification by KD functions as a barrier, or social moat, against cheaters and closely related competitors. Such division can also restrict the spread of deleterious infectious agents, e.g., phage and conjugating elements, by restricting spatial overlaps between groups. That is, although one group is compromised by an infection, it is less likely all groups are compromised, and thus the overall population is more resilient. Moreover, in the hypothetical absence of new KD loci, individuals in populations would continue to cooperate, even as they diversify by random mutations. Eventually, Hamilton’s rule (rb > c) would exceed a threshold, and genetic relatedness of recipients would no longer outweigh the cost to the actor. Therefore, MDEs provide social fluidity and selective forces that limit the level and size of cooperating groups. For a given situation, the optimal sizes of cooperating groups, as well their degree of relatedness, depends on the nature of their interactions, the properties of a taxa and local selective forces. Importantly, a specific benefit of fluid KD systems is that they target related individuals, which limits the size of cooperative groups and thus helps eliminate new competitors or cheaters that arise.
1.7. A Case Study: Rapid Social Diversification in a Local Natural Population by MDEs
To understand social diversification in nature, one needs natural isolates from a taxon that co-evolved in a particular niche. Such isolates are then tested for social compatibilities, and any incompatibilities can then be mapped to specific social genes. Unfortunately, there are few reports investigating social diversification in such a manner, nevertheless here we do outline one that involves a series of studies on M. xanthus.
Over 15 years ago, the Velicer group undertook a systematic approach to isolate M. xanthus strains from a small patch of forest soil [92]. From 100 adjacent soil plugs, they specifically sought M. xanthus isolates, where only one isolate was retained from each plug. In total, 78 attempts were successful. In a relatively simple social compatibility assay, they tested whether colony swarms would merge in harmony or whether they discriminated against one another, as indicated by demarcations. By analyzing a subset of strains, they found at least 45 distinct compatibility groups [9], though the total number is likely larger. These striking results revealed that M. xanthus isolates sharing the same habitat had diverged into many distinct social groups that actually kill their kin [5]. In separate work, draft genomes from the two largest clades were obtained [55]. These clades contained 22 of the 78 isolates and represented 11 distinct compatibility types. Their comparative genome analysis found numerous sequence differences between compatibility types that suggested a genetic basis for inter-strain incompatibilities, but the underlying mechanisms remained unknown [55].
From our work, we identified hundreds of unique KD sitAI loci, which in various theoretical combinations represented an astronomical number of ‘self-identity barcodes’ or kin groups [42,52]. With this information, we used comparative genomics to predict social group identities among the 22 strains with draft genomes. From this analysis of sitAI loci, we found a perfect correlation between unique sitAI loci and strains that belong to distinct compatibility types [5]. Of course, inherent in this analysis is that predictions are only possible between strains that have compatible TraA receptors that allow OME [44]. Therefore, since the clades represented two incompatible TraA receptors, there had to be a second KD mechanism to explain incompatibilities between isolates with divergent traA alleles. Based on prior reports [57,58], the T6SS was a plausible reservoir of KD effectors. By again using comparative genomics, we also found a perfect correlation between unique T6SS effectors and strain incompatibilities [5]. Next, we tested our predictions by constructing a series of mutants. Here, single mutations blocking OME or T6SS function did not relieve KD, but double mutants did for most strains. However, for two strains, a third KD system (Rhs), with unique effectors and apparent means of delivery, was identified [5,55], and when all three systems were inactivated, antagonism was relieved. Therefore, among strains within a clade (identical traA alleles), OME, T6SS and in some cases Rhs, governed KD. Finally, it is interesting to note that an HGT event occurred between these clades, presumably by transduction, whereby the traAB alleles were swapped. Therefore, these sub-clade isolates could no longer conduct OME with fellow clade members, but could from members of the other clade, thus dramatically changing their social interactions.
Strikingly, genome sequences among members within a clade showed they were highly related (e.g., DNA identities >99.99%) [55], but contained islands of unique sequences. We analyzed these islands and found they contained MGEs belonging to four prophage families and one insertion element family [5]. Importantly, all of the unique OME, T6SS and Rhs effector loci mapped to these MGEs (25 unique elements in 11 genomes). Therefore, in recent evolutionary time, these MDEs invaded ancestors of these clades and triggered their rapid diversification into 11 distinct social groups as depicted in Figure 2B. However, in this small patch of forest soil, these clades likely contained many additional social groups, because only a small number of strains were practically isolated. Moreover, these combined results suggest this particular microbiome contained a large pool of MDEs (Figure 1), the origin of which is unknown. In summary, these series of studies revealed how MGEs drove the rapid social diversification of M. xanthus populations in their natural habitat. This resulted in diverse social groups that appear to be resilient to changing and challenging environmental conditions.
2. Conclusions
Social diversification plays an important role in evolution because it facilitates genetic isolation, which, as described here, often coincides with antagonism. In turn, conflict and genetic isolation can accelerate evolution and lineage divergence by promoting arms races between groups [4]. Next-generation sequencing and metagenomics have provided powerful new approaches and insights into the dynamic nature of HGT of MGEs. This, combined with isolation methods and microbial assays, allows for exciting new discoveries into the mechanisms of diversification and its consequences in natural habitats. With these and other strategies, future work will unlock the fluid nature of microbial social interactions, how they change, and the impacts they have on a population’s fitness.
Author Contributions
M.L.W. and D.W. conceived, wrote, reviewed, edited and designed figures for the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Institutes of Health grant R35GM140886 to D.W.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lenski, R.E.; Travisano, M. Dynamics of adaptation and diversification: A 10,000-generation experiment with bacterial populations. Proc. Natl. Acad. Sci. USA 1994, 91, 6808–6814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strassmann, J.E.; Gilbert, O.M.; Queller, D.C. Kin discrimination and cooperation in microbes. Annu. Rev. Microbiol. 2011, 65, 349–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wall, D. Kin Recognition in bacteria. Annu. Rev. Microbiol. 2016, 70, 143–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Queller, D.C.; Strassmann, J.E. Evolutionary conflict. Annu. Rev. Ecol. Evol. Syst. 2018, 49, 73–93. [Google Scholar] [CrossRef]
- Vassallo, C.N.; Troselj, V.; Weltzer, M.L.; Wall, D. Rapid diversification of wild social groups driven by toxin-immunity loci on mobile genetic elements. ISME J. 2020, 14, 2474–2487. [Google Scholar] [CrossRef]
- Lyons, N.A.; Kraigher, B.; Stefanic, P.; Mandic-Mulec, I.; Kolter, R. A combinatorial kin discrimination system in Bacillus subtilis. Curr. Biol. 2016, 26, 733–742. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; de Souza, R.F.; Anantharaman, V.; Iyer, L.M.; Aravind, L. Polymorphic toxin systems: Comprehensive characterization of trafficking modes, processing, mechanisms of action, immunity and ecology using comparative genomics. Biol. Direct 2012, 7, 18. [Google Scholar] [CrossRef] [Green Version]
- Stefanic, P.; Kraigher, B.; Lyons, N.A.; Kolter, R.; Mandic-Mulec, I. Kin discrimination between sympatric Bacillus subtilis isolates. Proc. Natl. Acad. Sci. USA 2015, 112, 14042–14047. [Google Scholar] [CrossRef] [Green Version]
- Vos, M.; Velicer, G.J. Social Conflict in centimeter-and Ggobal-scale populations of the bacterium Myxococcus xanthus. Curr. Biol. 2009, 19, 1763–1767. [Google Scholar] [CrossRef] [Green Version]
- Gibbs, K.A.; Greenberg, E.P. Territoriality in Proteus: Advertisement and Aggression. Chem. Rev. 2010, 111, 188–194. [Google Scholar] [CrossRef] [Green Version]
- Rendueles, O.; Zee, P.C.; Dinkelacker, I.; Amherd, M.; Wielgoss, S.; Velicer, G.J. Rapid and widespread de novo evolution of kin discrimination. Proc. Natl. Acad. Sci. USA 2015, 112, 9076–9081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Z.; Yu, K.; Fu, S.; Xiao, Y.; Wei, Q.; Wang, D. Genomic analysis reveals high intra-species diversity of Shewanella algae. Microb. Genom. 2022, 8, 000786. [Google Scholar] [CrossRef]
- Innamorati, K.A.; Earl, J.P.; Aggarwal, S.D.; Ehrlich, G.D.; Hiller, N.L. The bacterial guide to designing a diversified gene portfolio. In The Pangenome: Diversity, Dynamics and Evolution of Genomes; Tettelin, H., Medini, D., Eds.; Springer: Cham, Switzerland, 2020; pp. 51–87. [Google Scholar]
- Douglas, G.M.; Shapiro, B.J. Genic selection within prokaryotic pangenomes. Genome Biol. Evol. 2021, 13, evab234. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.N.; Sheriff, E.K.; Duerkop, B.A.; Chatterjee, A. Let me upgrade you: Impact of mobile genetic elements on Enterococcal adaptation and evolution. J. Bacteriol. 2021, 203, e0017721. [Google Scholar] [CrossRef]
- Makarova, K.S.; Wolf, Y.I. Evolutionary genomics of defense systems in archaea and bacteria. Annu. Rev. Microbiol. 2017, 71, 233–261. [Google Scholar] [CrossRef]
- Tatusova, T.; Ciufo, S.; Fedorov, B.; O’Neill, K.; Tolstoy, I. RefSeq microbial genomes database: New representation and annotation strategy. Nucleic Acids Res. 2013, 42, D553–D559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puigbò, P.; Lobkovsky, A.E.; Kristensen, D.M.; Wolf, Y.I.; Koonin, E.V. Genomes in turmoil: Quantification of genome dynamics in prokaryote supergenomes. BMC Biol. 2014, 12, 66. [Google Scholar] [CrossRef]
- Mira, A.; Ochman, H.; Moran, N.A. Deletional bias and the evolution of bacterial genomes. Trends Genet. 2001, 17, 589–596. [Google Scholar] [CrossRef] [PubMed]
- Kuo, C.-H.; Ochman, H. Deletional bias across the three domains of life. Genome Biol. Evol. 2009, 1, 145–152. [Google Scholar] [CrossRef] [Green Version]
- Albalat, R.; Cañestro, C. Evolution by gene loss. Nat. Rev. Genet. 2016, 17, 379–391. [Google Scholar] [CrossRef]
- Olson, M.V. When Less Is More: Gene loss as an engine of evolutionary change. Am. J. Hum. Genet. 1999, 64, 18–23. [Google Scholar] [CrossRef] [Green Version]
- Hottes, A.K.; Freddolino, P.L.; Khare, A.; Donnell, Z.N.; Liu, J.; Tavazoie, S. Bacterial adaptation through loss of function. PLoS Genet. 2013, 9, e1003617. [Google Scholar] [CrossRef]
- Hamilton, W.D. The genetical theory of social behaviour. II. J. Thour. Biol. 1964, 7, 1–52. [Google Scholar] [CrossRef]
- Fleischman, D.S. Pathological altruism isn’t pathological or altruistic. Cogn. Neuropsychiatry 2013, 18, 631–636. [Google Scholar] [CrossRef]
- Bourke, A.F. Hamilton’s rule and the causes of social evolution. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014, 369, 20130362. [Google Scholar] [CrossRef] [Green Version]
- Özkaya, O.; Xavier, K.B.; Dionisio, F.; Balbontín, R. Maintenance of microbial cooperation mediated by public goods in single- and multiple-trait scenarios. J. Bacteriol. 2017, 199, e00297-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavaliere, M.; Yang, G.; Danos, V.; Dakos, V. Detecting the collapse of cooperation in evolving networks. Sci. Rep. 2016, 6, 30845. [Google Scholar] [CrossRef] [PubMed]
- Fiegna, F.; Velicer, G.J. Competitive fates of bacterial social parasites: Persistence and self-induced extinction of Myxococcus xanthus cheaters. Proc. Biol. Sci. 2003, 270, 1527–1534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, B.; Holkenbrink, C.; Treuner-Lange, A.; Higgs, P.I. Myxococcus xanthus developmental cell fate production: Heterogeneous accumulation of developmental regulatory proteins and reexamination of the role of MazF in developmental lysis. J. Bacteriol. 2012, 194, 3058–3068. [Google Scholar] [CrossRef] [Green Version]
- Fiegna, F.; Yu, Y.-T.N.; Kadam, S.V.; Velicer, G.J. Evolution of an obligate social cheater to a superior cooperator. Nature 2006, 441, 310–314. [Google Scholar] [CrossRef]
- Velicer, G.J.; Kroos, L.; Lenski, R.E. Developmental cheating in the social bacterium Myxococcus xanthus. Nature 2000, 404, 598–601. [Google Scholar] [CrossRef] [PubMed]
- Velicer, G.J.; Yu, Y.-T.N. Evolution of novel cooperative swarming in the bacterium Myxococcus xanthus. Nature 2003, 425, 75–78. [Google Scholar] [CrossRef]
- Travisano, M.; Velicer, G.J. Strategies of microbial cheater control. Trends Microbiol. 2004, 12, 72–78. [Google Scholar] [CrossRef]
- Smith, J.; Van Dyken, J.D.; Velicer, G.J. Nonadaptive processes can create the appearance of facultative cheating in microbes. Evolution 2013, 68, 816–826. [Google Scholar] [CrossRef]
- Pande, S.; Escriva, P.P.; Yu, Y.-T.N.; Sauer, U.; Velicer, G.J. Cooperation and cheating among germinating spores. Curr. Biol. 2020, 30, 4745–4752.e4. [Google Scholar] [CrossRef] [PubMed]
- Buss, L.W. Somatic cell parasitism and the evolution of somatic tissue compatibility. Proc. Natl. Acad. Sci. USA 1982, 79, 5337–5341. [Google Scholar] [CrossRef] [Green Version]
- Strassmann, J.E.; Zhu, Y.; Queller, D.C. Altruism and social cheating in the social amoeba Dictyostelium discoideum. Nature 2000, 408, 965–967. [Google Scholar] [CrossRef]
- Ennis, H.L.; Dao, D.N.; Pukatzki, S.U.; Kessin, R.H. Dictyostelium amoebae lacking an F-box protein form spores rather than stalk in chimeras with wild type. Proc. Natl. Acad. Sci. USA 2000, 97, 3292–3297. [Google Scholar] [CrossRef]
- Ho, H.-I.; Hirose, S.; Kuspa, A.; Shaulsky, G. Kin recognition protects cooperators against cheaters. Curr. Biol. 2013, 23, 1590–1595. [Google Scholar] [CrossRef] [Green Version]
- Diggle, S.P.; Griffin, A.S.; Campbell, G.S.; West, S.A. Cooperation and conflict in quorum-sensing bacterial populations. Nature 2007, 450, 411–414. [Google Scholar] [CrossRef] [PubMed]
- Sah, G.P.; Wall, D. Kin recognition and outer membrane exchange (OME) in myxobacteria. Curr. Opin. Microbiol. 2020, 56, 81–88. [Google Scholar] [CrossRef]
- Cao, P.; Wei, X.; Awal, R.P.; Müller, R.; Wall, D. A highly polymorphic receptor governs many distinct self-recognition types within the Myxococcales Order. mBio 2019, 10, e02751-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pathak, D.T.; Wei, X.; Dey, A.; Wall, D. Molecular recognition by a polymorphic cell surface receptor governs cooperative behaviors in bacteria. PLoS Genet. 2013, 9, e1003891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pathak, D.T.; Wei, X.; Bucuvalas, A.; Haft, D.H.; Gerloff, D.L.; Wall, D. Cell contact–dependent outer membrane exchange in myxobacteria: Genetic Determinants and Mechanism. PLoS Genet. 2012, 8, e1002626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stubbendieck, R.M.; Straight, P.D. Multifaceted interfaces of bacterial competition. J. Bacteriol. 2016, 198, 2145–2155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Bayona, L.; Comstock, L.E. Bacterial antagonism in host-associated microbial communities. Science 2018, 361, eaat2456. [Google Scholar] [CrossRef] [Green Version]
- Ruhe, Z.C.; Low, D.A.; Hayes, C.S. Polymorphic toxins and their immunity proteins: Diversity, evolution, and mechanisms of delivery. Annu. Rev. Microbiol. 2020, 74, 497–520. [Google Scholar] [CrossRef]
- Cao, P.; Dey, A.; Vassallo, C.N.; Wall, D. How myxobacteria cooperate. J. Mol. Biol. 2015, 427, 3709–3721. [Google Scholar] [CrossRef] [Green Version]
- Vassallo, C.N.; Cao, P.; Conklin, A.; Finkelstein, H.; Hayes, C.S.; Wall, D. Infectious polymorphic toxins delivered by outer membrane exchange discriminate kin in myxobacteria. eLife 2017, 6, e29397. [Google Scholar] [CrossRef]
- Vassallo, C.N.; Sah, G.P.; Weltzer, M.L.; Wall, D. Modular lipoprotein toxins transferred by outer membrane exchange target discrete cell entry pathways. mBio 2021, 12, e02388-21. [Google Scholar] [CrossRef]
- Vassallo, C.N.; Wall, D. Self-identity barcodes encoded by six expansive polymorphic toxin families discriminate kin in myxobacteria. Proc. Natl. Acad. Sci. USA 2019, 116, 24808–24818. [Google Scholar] [CrossRef]
- Patra, P.; Vassallo, C.N.; Wall, D.; Igoshin, O.A. Mechanism of kin-discriminatory demarcation line formation between colonies of swarming bacteria. Biophys. J. 2017, 113, 2477–2486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dey, A.; Vassallo, C.N.; Conklin, A.C.; Pathak, D.T.; Troselj, V.; Wall, D. Sibling rivalry in Myxococcus xanthus is mediated by kin recognition and a polyploid prophage. J. Bacteriol. 2016, 198, 994–1004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wielgoss, S.; Didelot, X.; Chaudhuri, R.; Liu, X.; Weedall, G.; Velicer, G.J.; Vos, M. A barrier to homologous recombination between sympatric strains of the cooperative soil bacterium Myxococcus xanthus. ISME J. 2016, 10, 2468–2477. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Wang, J.; Zhang, Z.; Wang, F.; Gong, Y.; Sheng, D.-H.; Li, Y.-Z. Two PAAR proteins with different C-terminal extended domains have distinct ecological functions in Myxococcus xanthus. Appl. Environ. Microbiol. 2021, 87, e00080-21. [Google Scholar] [CrossRef]
- Gong, Y.; Zhang, Z.; Liu, Y.; Zhou, X.; Anwar, M.N.; Li, Z.; Hu, W.; Li, Y. A nuclease-toxin and immunity system for kin discrimination inMyxococcus xanthus. Environ. Microbiol. 2018, 20, 2552–2567. [Google Scholar] [CrossRef] [PubMed]
- Troselj, V.; Treuner-Lange, A.; Søgaard-Andersen, L.; Wall, D. Physiological heterogeneity triggers sibling conflict mediated by the type VI secretion system in an aggregative multicellular bacterium. mBio 2018, 9, e01645-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kraigher, B.; Butolen, M.; Stefanic, P.; Mulec, I.M. Kin discrimination drives territorial exclusion during Bacillus subtilis swarming and restrains exploitation of surfactin. ISME J. 2021, 16, 833–841. [Google Scholar] [CrossRef] [PubMed]
- Lyons, N.A.; Kolter, R. Bacillus subtilis protects public goods by extending kin discrimination to closely related species. mBio 2017, 8, e00723-17. [Google Scholar] [CrossRef] [Green Version]
- Vacheron, J.; Heiman, C.M.; Keel, C. Live cell dynamics of production, exPLoSive release and killing activity of phage tail-like weapons for Pseudomonas kin exclusion. Commun. Biol. 2021, 4, 87. [Google Scholar] [CrossRef]
- Riley, M.A.; Wertz, J.E. Bacteriocins: Evolution, ecology, and application. Annu. Rev. Microbiol. 2002, 56, 117–137. [Google Scholar] [CrossRef] [Green Version]
- Koonin, E.V.; Makarova, K.S.; Wolf, Y.I.; Krupovic, M. Evolutionary entanglement of mobile genetic elements and host defense systems: Guns for hire. Nat. Rev. Genet. 2019, 21, 119–131. [Google Scholar] [CrossRef] [PubMed]
- Dragoš, A.; Andersen, A.J.; Lozano-Andrade, C.N.; Kempen, P.J.; Kovács, Á.T.; Strube, M.L. Phages carry interbacterial weapons encoded by biosynthetic gene clusters. Curr. Biol. 2021, 31, 3479–3489.e5. [Google Scholar] [CrossRef]
- Dienes, L. Reproductive processes in Proteus cultures. Exp. Biol. Med. 1946, 63, 265–270. [Google Scholar] [CrossRef]
- Gibbs, K.A.; Urbanowski, M.L.; Greenberg, E.P. Genetic determinants of self identity and social recognition in bacteria. Science 2008, 321, 256–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saak, C.C.; Gibbs, K.A. The self-identity protein IdsD is communicated between cells in swarming Proteus mirabilis colonies. J. Bacteriol. 2016, 198, 3278–3286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tipping, M.J.; Gibbs, K.A. Peer pressure from a Proteus mirabilis self-recognition system controls participation in cooperative swarm motility. PLoS Pathog. 2019, 15, e1007885. [Google Scholar] [CrossRef] [Green Version]
- Alteri, C.; Himpsl, S.D.; Pickens, S.R.; Lindner, J.R.; Zora, J.S.; Miller, J.; Arno, P.D.; Straight, S.W.; Mobley, H.L.T. Multicellular bacteria deploy the type VI secretion system to preemptively strike neighboring cells. PLoS Pathog. 2013, 9, e1003608. [Google Scholar] [CrossRef]
- Adams, D.W.; Stutzmann, S.; Stoudmann, C.; Blokesch, M. DNA-uptake pili of Vibrio cholerae are required for chitin colonization and capable of kin recognition via sequence-specific self-interaction. Nat. Microbiol. 2019, 4, 1545–1557. [Google Scholar] [CrossRef]
- Cascales, E.; Buchanan, S.K.; Duché, D.; Kleanthous, C.; Lloubes, R.; Postle, K.; Riley, M.; Slatin, S.; Cavard, D. Colicin biology. Microbiol. Mol. Biol. Rev. 2007, 71, 158–229. [Google Scholar] [CrossRef] [Green Version]
- Morgado, S.; Vicente, A.C. Diversity and distribution of type VI secretion system gene clusters in bacterial plasmids. Sci. Rep. 2022, 12, 8249. [Google Scholar] [CrossRef] [PubMed]
- Hussain, N.A.S.; Kirchberger, P.C.; Case, R.J.; Boucher, Y.F. Modular molecular weaponry plays a key role in competition within an environmental Vibrio cholerae population. Front. Microbiol. 2021, 12, 671092. [Google Scholar] [CrossRef] [PubMed]
- Speare, L.; Cecere, A.G.; Guckes, K.R.; Smith, S.; Wollenberg, M.S.; Mandel, M.J.; Miyashiro, T.; Septer, A.N. Bacterial symbionts use a type VI secretion system to eliminate competitors in their natural host. Proc. Natl. Acad. Sci. USA 2018, 115, E8528–E8537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Speare, L.; Woo, M.; Dunn, A.K.; Septer, A.N. A putative lipoprotein mediates cell-cell contact for type VI secretion system-dependent killing of specific competitors. mBio 2022, 13, e03085-21. [Google Scholar] [CrossRef] [PubMed]
- Jamet, A.; Touchon, M.; Ribeiro-Gonçalves, B.; Carrico, J.; Charbit, A.; Nassif, X.; Ramirez, M.; Rocha, E. A widespread family of polymorphic toxins encoded by temperate phages. BMC Biol. 2017, 15, 75. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Guo, Y.; Kim, J.-S.; Wang, X.; Wood, T.K. Phages mediate bacterial self-recognition. Cell Rep. 2019, 27, 737–749.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poole, S.J.; Diner, E.J.; Aoki, S.K.; Braaten, B.A.; De Roodenbeke, C.T.; Low, D.A.; Hayes, C.S. Identification of functional toxin/Immunity genes linked to contact-dependent growth inhibition (CDI) and rearrangement hotspot (Rhs) systems. PLoS Genet. 2011, 7, e1002217. [Google Scholar] [CrossRef]
- Ocasio, A.B.; Cotter, P.A. CDI/CDS system-encoding genes of Burkholderia thailandensis are located in a mobile genetic element that defines a new class of transposon. PLoS Genet. 2019, 15, e1007883. [Google Scholar] [CrossRef] [Green Version]
- Ruhe, Z.C.; Nguyen, J.Y.; Chen, A.J.; Leung, N.Y.; Hayes, C.S.; Low, D.A. CDI systems are stably maintained by a cell-contact mediated surveillance mechanism. PLoS Genet. 2016, 12, e1006145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coyne, M.J.; Zitomersky, N.L.; McGuire, A.M.; Earl, A.M.; Comstock, L.E. Evidence of extensive DNA transfer between Bacteroidales species within the human gut. mBio 2014, 5, e01305-14. [Google Scholar] [CrossRef] [Green Version]
- García-Bayona, L.; Coyne, M.J.; Comstock, L.E. Mobile Type VI secretion system loci of the gut Bacteroidales display extensive intra-ecosystem transfer, multi-species spread and geographical clustering. PLoS Genet. 2021, 17, e1009541. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.P.A.; Eldakar, O.T.; Gogarten, J.P.; Andam, C.P. Bacterial cooperation through horizontal gene transfer. Trends Ecol. Evol. 2021, 37, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Smith, J. The social evolution of bacterial pathogenesis. Proc. R. Soc. B Boil. Sci. 2001, 268, 61–69. [Google Scholar] [CrossRef] [Green Version]
- Nogueira, T.; Rankin, D.J.; Touchon, M.; Taddei, F.; Brown, S.P.; Rocha, E.P. Horizontal gene transfer of the secretome drives the evolution of bacterial cooperation and virulence. Curr. Biol. 2009, 19, 1683–1691. [Google Scholar] [CrossRef] [Green Version]
- Rankin, D.J.; Rocha, E.P.C.; Brown, S.P. What traits are carried on mobile genetic elements, and why? Heredity 2011, 106, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Giraud, T.; A Shykoff, J. Bacterial cooperation controlled by mobile elements: Kin selection versus infectivity. Heredity 2011, 107, 277–278, author reply 279–281. [Google Scholar] [CrossRef]
- Dewar, A.E.; Thomas, J.L.; Scott, T.W.; Wild, G.; Griffin, A.S.; West, S.A.; Ghoul, M. Plasmids do not consistently stabilize cooperation across bacteria but may promote broad pathogen host-range. Nat. Ecol. Evol. 2021, 5, 1624–1636. [Google Scholar] [CrossRef] [PubMed]
- Morris, J.J.; Lenski, R.E.; Zinser, E.R. The Black Queen Hypothesis: Evolution of dependencies through adaptive genelLoss. mBio 2012, 3, e00036-12. [Google Scholar] [CrossRef] [Green Version]
- Fu, S.; Wang, Q.; Wang, R.; Zhang, Y.; Lan, R.; He, F.; Yang, Q. Horizontal transfer of antibiotic resistance genes within the bacterial communities in aquacultural environment. Sci. Total. Environ. 2022, 820, 153286. [Google Scholar] [CrossRef]
- Hardin, G. The competitive exclusion principle. Science 1960, 131, 1292–1297. [Google Scholar] [CrossRef] [Green Version]
- Vos, M.; Velicer, G.J. Genetic population structure of the soil bacterium Myxococcus xanthus at the centimeter scale. Appl. Environ. Microbiol. 2006, 72, 3615–3625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Genome fluidity and social evolution. Top cell receives mobile discrimination element (MDE) from the environment, resulting in sibling A antagonizing sibling B. However, after many rounds of cell division, a daughter cell may lose the MDE, rendering it susceptible to its ancestral population, but nevertheless contributing toward a reduced genome and resulting in different fitness gains.
Figure 1.
Genome fluidity and social evolution. Top cell receives mobile discrimination element (MDE) from the environment, resulting in sibling A antagonizing sibling B. However, after many rounds of cell division, a daughter cell may lose the MDE, rendering it susceptible to its ancestral population, but nevertheless contributing toward a reduced genome and resulting in different fitness gains.

Figure 2.
Social diversification mediated by HGT of MDEs. (A) A cell in a clonal population is transformed with an MDE containing a novel toxin-immunity locus that creates a new discrimination type (NDT; different color), which antagonizes siblings and overtakes the population. (B) Multiple cells in a clonal population transformed by different MDEs. These NDTs antagonize their siblings, resulting in new social groups that likely antagonize each other and do not cooperate.
Figure 2.
Social diversification mediated by HGT of MDEs. (A) A cell in a clonal population is transformed with an MDE containing a novel toxin-immunity locus that creates a new discrimination type (NDT; different color), which antagonizes siblings and overtakes the population. (B) Multiple cells in a clonal population transformed by different MDEs. These NDTs antagonize their siblings, resulting in new social groups that likely antagonize each other and do not cooperate.

Figure 3.
Mechanisms of kin discrimination. (A) Toxin delivery by four general strategies: 1. Homotypic binding between compatible receptors. In myxobacteria, this occurs between TraAB receptors triggering OME and delivery of a suite of SitA lipoprotein toxins. 2. Cell releases diffusible bacteriocins/tailocins that target distant relatives. 3. Cell-contact-dependent secretion systems, e.g., T6SS, deliver toxins to target cell. 4. Toxin displayed on distal end of a protruding cell surface protein, e.g., CDI, targeting a receptor on kin. * Recognition not required but can occur; ** required for CDI, unknown for very large class of Rhs proteins found in myxobacteria. Check mark means yes; red x means no. (B) Delivered toxins either neutralized by cognate immunity factors in close kin, e.g., clonemates, or poison distant relatives lacking immunity.
Figure 3.
Mechanisms of kin discrimination. (A) Toxin delivery by four general strategies: 1. Homotypic binding between compatible receptors. In myxobacteria, this occurs between TraAB receptors triggering OME and delivery of a suite of SitA lipoprotein toxins. 2. Cell releases diffusible bacteriocins/tailocins that target distant relatives. 3. Cell-contact-dependent secretion systems, e.g., T6SS, deliver toxins to target cell. 4. Toxin displayed on distal end of a protruding cell surface protein, e.g., CDI, targeting a receptor on kin. * Recognition not required but can occur; ** required for CDI, unknown for very large class of Rhs proteins found in myxobacteria. Check mark means yes; red x means no. (B) Delivered toxins either neutralized by cognate immunity factors in close kin, e.g., clonemates, or poison distant relatives lacking immunity.

Figure 4.
Evolution of social diversification or unification. Selective forces drive populations to become more unified or diverse. Unification forces include a dominant genotype that outcompetes related genotypes or selective pressures favoring large cooperative groups. Rapid diversification is frequently driven by HGT (Figure 2).
Figure 4.
Evolution of social diversification or unification. Selective forces drive populations to become more unified or diverse. Unification forces include a dominant genotype that outcompetes related genotypes or selective pressures favoring large cooperative groups. Rapid diversification is frequently driven by HGT (Figure 2).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
KD systems and effectors found on MGEs.
| MGE Type | Type of Genes Encoded | Host Organism | References |
|---|---|---|---|
| Plasmid | Colicin, T6SS, SitA toxin | E. coli, Vibrio, Proteobacteria (e.g., Rhizobium, Ralstonia), Myxococcus | [52,71,72,74,75] |
| Prophage | Effectors T6SS, OME, Rhs; MuF toxins, bacteriocins | Myxococcus, E. coli, widespread | [5,64,76] |
| Transposon | CDI | E. coli, Dickeya, Burkholderia | [77,78,79,80] |
| ICE | T6SS, toxins, bacteriocins | Bacteroides, Parabacteroides | [78,79] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Weltzer, M.L.; Wall, D. Social Diversification Driven by Mobile Genetic Elements. Genes 2023, 14, 648. https://doi.org/10.3390/genes14030648
AMA Style
Weltzer ML, Wall D. Social Diversification Driven by Mobile Genetic Elements. Genes. 2023; 14(3):648. https://doi.org/10.3390/genes14030648
Chicago/Turabian StyleWeltzer, Michael L., and Daniel Wall. 2023. "Social Diversification Driven by Mobile Genetic Elements" Genes 14, no. 3: 648. https://doi.org/10.3390/genes14030648
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.