1. Introduction
China is the largest meat market in the world, approximately 18% of chicken production in the world is consumed in China. Likewise, today’s projection research has forecasted that the enlargement of the poultry market is expected to continue in the future. Furthermore, consumers prefer chicken meat, particularly breast meat, because of its delicious taste, healthy nutritional content, ease of preparation, and ability to meet modern consumer needs [
1,
2]. Broiler chickens have been selected over the last few decades to improve carcass traits and increase thigh and breast yields. As a result, meat quality traits have been negatively impacted, and some undesirable meat quality traits have emerged, such as a decrease in taste quality and sensory adequacy for consumers. Meat quality is a complete concept that includes tenderness, flavor, nutrition, appearance, and other factors. However, many factors influence meat quality, including breed, feeding, management system, sex, and age, with the breed being the most important [
3]. The CH Chicken breed, a native chicken breed in China, including the Yunnan chicken breed, has a unique meat quality compared to the DG broiler chicken and is extremely popular in Yunnan due to its meat characteristics. For this reason, the explanation of the molecular basis of meat quality in CH and DG chicken breeds will have an economic and biological effect. Furthermore, there are multiple genes that regulate meat quality traits in chicken, and some genes have been reported as associated with this trait, such as
TYRP1, TYR, DCT [
4],
AGA, COL28A1, COL1A2 [
5],
MB, HBAD, HBA1 [
6]
, ACACA, ACADL, ACSL1, ATP8A1, CAV1, FADS2, FASN [
7], DCN,
CHCHD10 [
8], and
MORN4 [
9].
In earlier decades, the main methods for identifying genes associated with meat quality and carcass trait performance in chickens were genome-wide association studies, quantitative trait locus mapping, and candidate gene analysis. As a result, many genomic regions and genetic associations have been identified. Similarly, genome-wide association studies can be used to identify the variants and genes that control economic traits [
10,
11] as they have been for meat quality [
5,
12] and carcass traits [
13,
14,
15]. Although these methods have contributed significantly to our improved understanding of the underlying mechanisms of genetic traits, some potential constraints are still present. The important restriction is a good mapping needed to identify relevant variations. Furthermore, some biological pathways or novel genes linked with the targeted trait can be excluded accidentally. Next-generation sequencing (NGS) technologies have been widely used in recent years to identify differential expression as well as potential novel transcripts [
16]. Based on the preceding discussion, RNA-Seq technology has been widely used to identify differentially expressed genes between two gene expression patterns, alternative splicing events, and causative variants.
Transcriptomic approach analysis is a foundation tool for gene function and structure studies [
17], and it has become a fairly standard methodology in genetic studies, with growing applications in animal breeding [
18]. General transcriptomes or complete transcripts include messenger RNA, ribosomal RNA, and transfer RNA [
19,
20]. RNA-sequencing analysis (transcriptional sequencing) has rapidly developed over the past few years, a technology of deep sequencing for transcriptome analysis [
21]. This technology can detect the whole transcriptome at a single nucleotide level. Transcript expression and structure can provide analysis by RNA-seq technology. Moreover, the approach to gene expression could contribute to the systematic studies undertaken by evaluating the suitability of pathways representing significantly responsible genes responsible for genetic characteristics [
22,
23].
Recently, many studies have reported using RNA-seq technology [
15,
24] or microarray technology [
25] on chicken transcriptomes. Besides that, chicken transcriptome analysis has already been performed for fatty acid metabolism [
8], ultimate pH (pHu) [
9], growth [
26,
27], feed efficiency [
28], meat quality and muscle growth [
29]. Wu et al. [
30] performed RNA-seq technology of leg muscle between fast and slow-growing chickens of Jinghai yellow chickens with different body weights at 300 days to study the molecular mechanism of chicken growth. Between fast and slow-growing chickens, 87 genes were found to be differentially expressed. Cui et al. [
31] used microarray technology to identify differentially expressed genes related to intramuscular fat (IMF) metabolism in Beijing-you chicken breast and thigh at 42 and 90 days of age, and 515 DEGs and 36 DEGs related to IMF metabolism were identified between the breast and thigh at 42 and 90 days. Furthermore, molecular pathways and genes related to regulating thigh meat quality and feed efficiency of Korat chickens were examined using microarray technology. The functional analysis emphasized numerous enriched function pathways, such as metabolic processes, immune responses, biological processes, and nucleotide metabolism [
25]. The study explored biological genes related to controlling melanogenesis pathways in the breast muscles of Wu Liangshan black-boned chickens and broiler chickens. The findings identified 25 differentially expressed genes related to the melanogenesis pathways [
4]. Zhang et al. [
27] showed that 4608 differentially expressed genes (DEGs) were obtained by comparison of the Jinghai yellow using RNA-seq to study the transcriptome of the breast muscle. Karimi et al. [
28] identified target genes that relate to feed efficiency by analyzing transcriptome profiles of liver tissue in native and commercial chicken breeds (Ross vs. native breeds) using RNA-Seq data. However, most of these studies have been focused on the broilers and layers of chicken breeds, still, the understanding of the mechanisms underlying meat quality and muscle development due to Chinese chicken breeds has remained limited. Chahua (CH2) is a new line of chickens. It was created through five generations of crossbreeding and selection from the CH Chicken breed and other breeds in Yunnan Province. It was established to develop the CH chicken breed to promote farming, ensure food safety in small villages, and maintain local chicken breeds. Slow-growing CH chickens are commonly slaughtered on the I2 to 16 weeks of age. Its meat is recognized as high quality [
32,
33]. However, the slow growth of CH chickens causes high production costs [
34].
On the other hand, Digao chicken is a commercial chicken breed (broiler). Yunling Guangda Breeding Poultry Feed Company imported this breed from Australia in 1997 (Yunling Guangda Breeding Poultry Feed Co., Ltd., Kunming, China). At present, DG chicken breeds are the only products in Yunnan province. This breed is characterized by a variety of production, strong adaptability, fast growth rate, and delicious meat, it is suitable for breeding and market needs at all levels. The parent breeders of this breed are red feathers, with a unique “recessive white feather” genetic trait, which can be used as the mother of local breed improvement. The body weight of a 56-day-old commercial chicken with rapid feather growth and easy identification of males and females (male for 3–3.2 kg, female 2.8–3 kg). For this reason, the improvement of CH chickens genetically to increase growth rate is required with maintaining high meat quality for this breed.
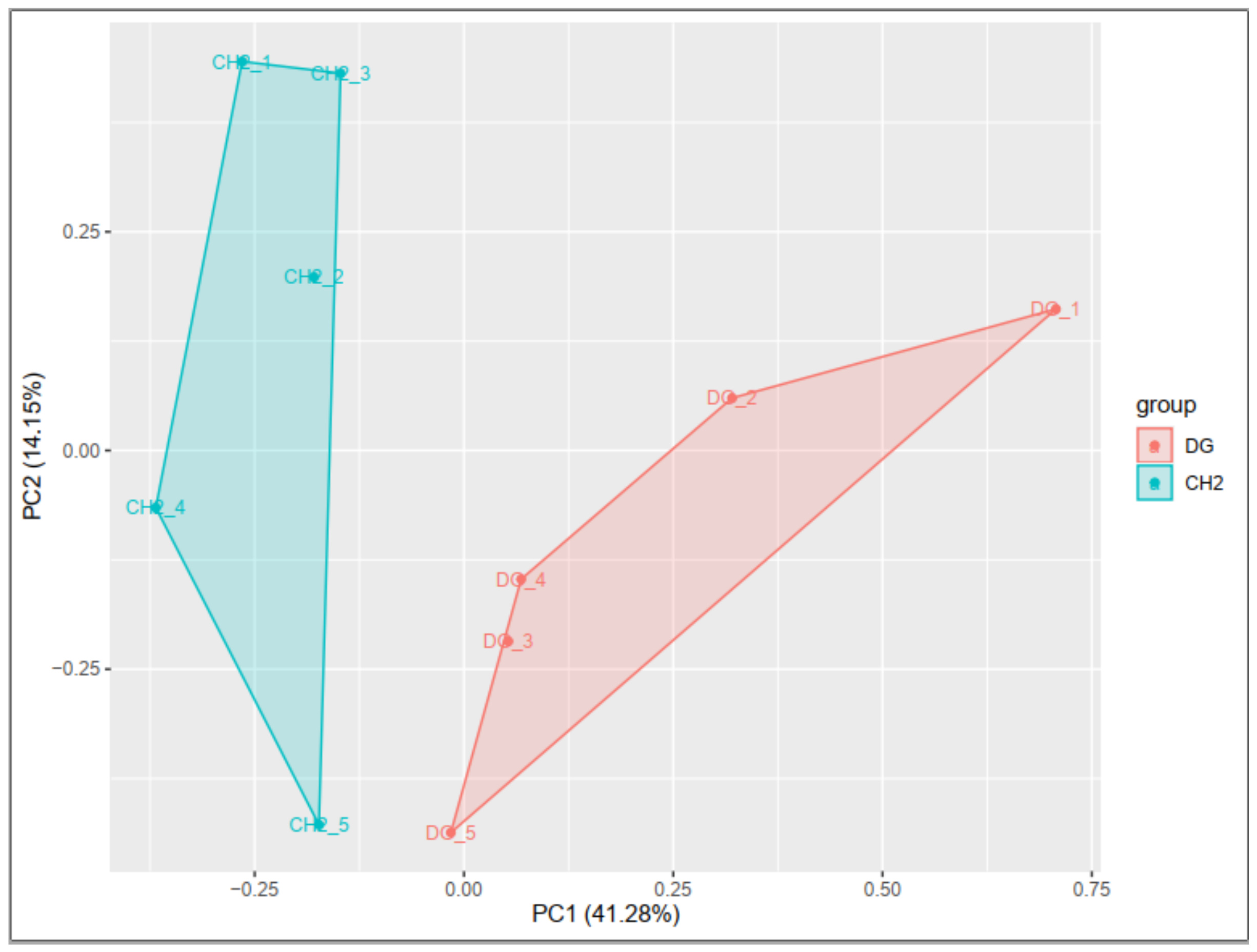
The perspectives for a genetic enhancement between local and commercial chicken breeds are completely different. Furthermore, local chickens are destined for high-quality chicken meat; for this reason, these breeds are characterized by high meat quality. As study subjects, we used 100-day-old DG and CH chicken breeds. The Chahua chicken breed and the Digao chicken breed are two distinct breeds, with the Chahua chicken having high meat quality and the Digao chicken having low meat quality when compared to the Chahua chicken. The aim of the research was to determine the pathways and genes that may be involved in the meat quality formation of breast muscle of these two breeds.
4. Discussion
Transcriptome sequencing via RNA resequencing or microarray technologies can be used to investigate the regulation of meat quality and carcass traits in chickens in a systematic manner, with some success in recent years.
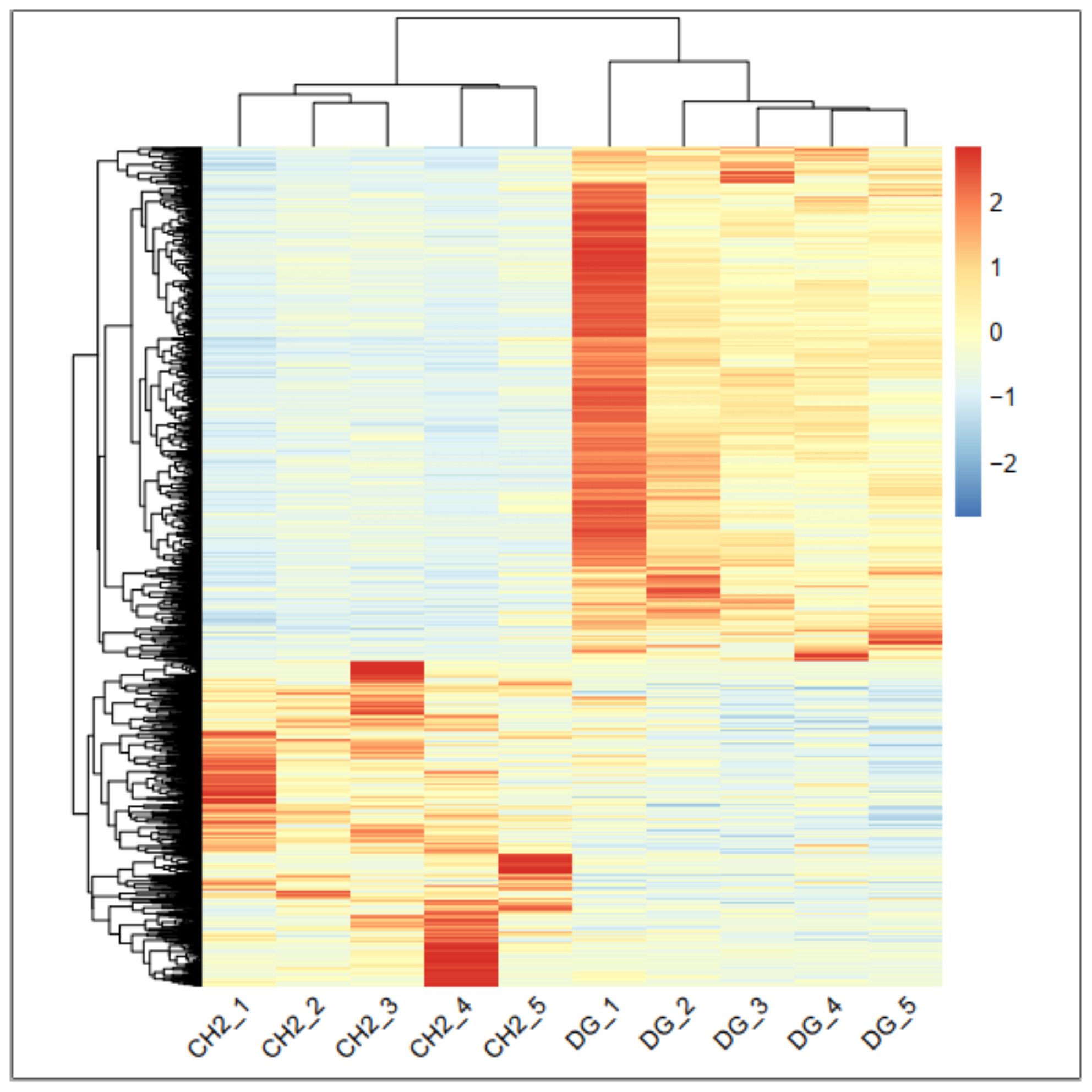
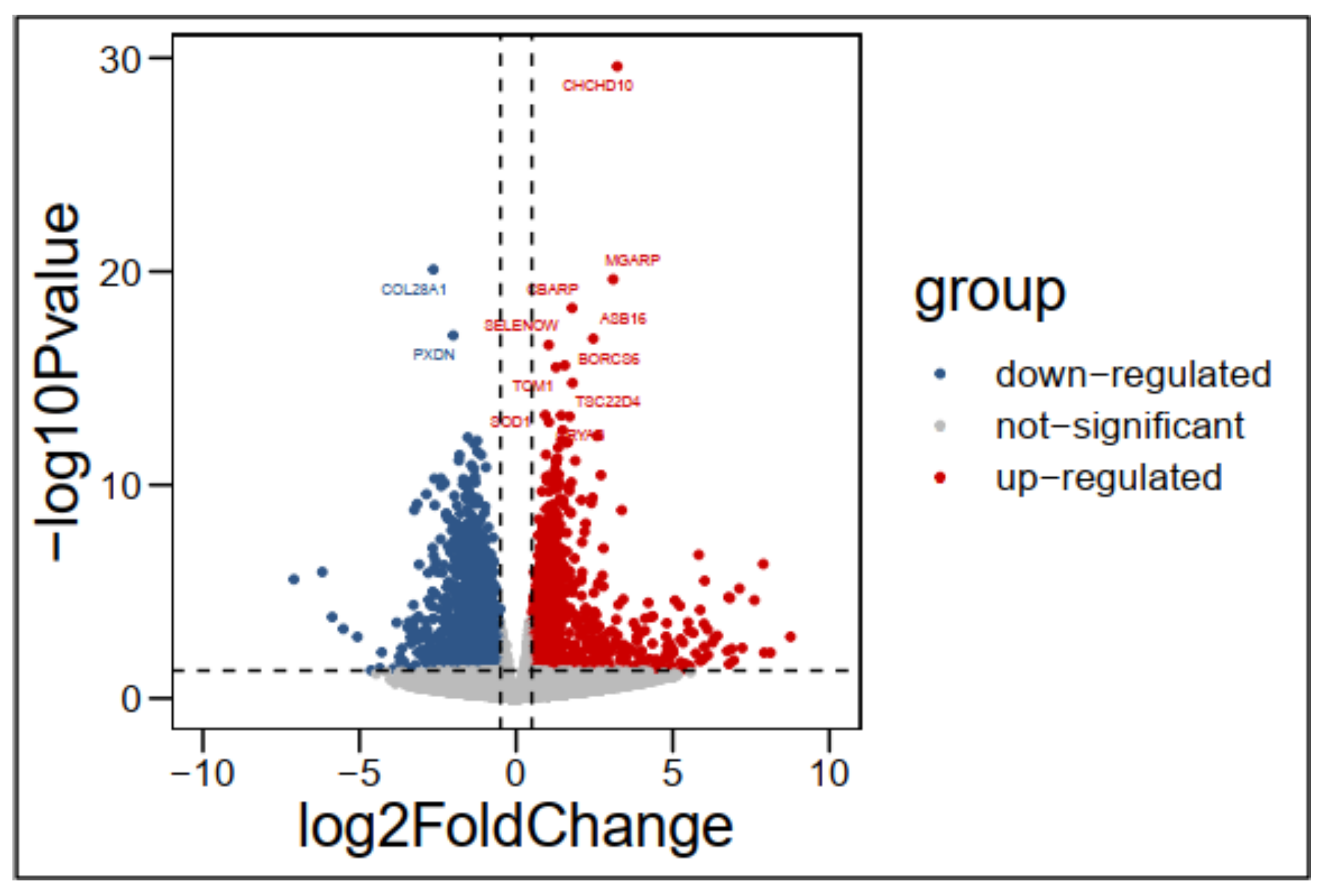
The results found 3525 differentially expressed genes DEGs of which 1750 were up-regulated and 1775 down regulated in CH compared to DG chicken. Using RNA-seq to study the transcriptome of the breast muscle, [
27] showed that 4608 differentially expressed genes (DEGs) were obtained by comparison of the Jinghai yellow. The CH chicken breed is a native breed distinguished by high-quality meat and slow growth. The DG chicken breed (broiler) is distinguished by low-quality meat and rapid growth. Therefore, evaluating the differences between these two breeds in regulatory pathways and genes is important to unravel the mechanisms of the DG breed’s rapid growth and the CH breed’s high-quality meat.
In this study, commercial chicken breed (DG) and native chicken (CH) breed have been used. These two breeds differ in meat quality traits and have a similar age. That’s why it is an ideal animal model to investigate the molecular mechanism of meat quality. In this study, RNA-Seq technique was used on breast muscle tissues from 100 days-old CH and DG broilers and identified a number of key DEGs that may affect meat quality.
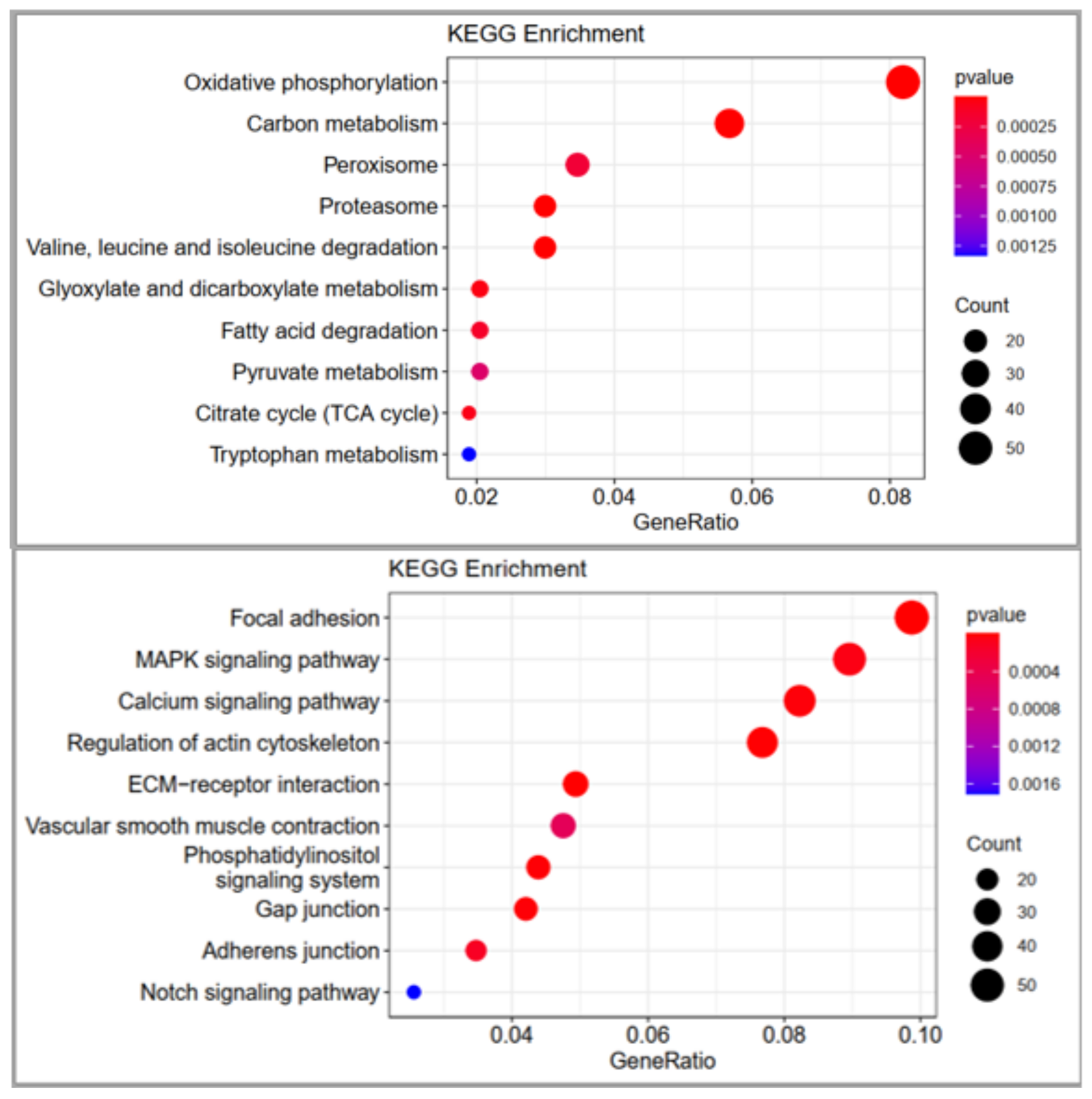
There are many pathways and genes that are related to meat quality traits reported in this study. Sun et al. [
5] revealed 14 genes as a candidate gene for meat quality, including
TYRO3 and
AGA genes for intramuscular fat content in breast muscle,
COL28A1 and
COL1A2 for meat color and RET gene for abdominal fat traits when conducting a genome-wide association study to identify genes associated with the meat quality of Beijing-You chickens and Cobb-Vantress. breeds Li et al. [
6] collected breast muscle samples of chicken and found several genes related to muscle color and amino acids metabolism, including
MB, HBAA, HBE1 HBM and
HBE, relating to the meat color and
PHGDH, DMGDH, AGXT and
FTCD are related to biosynthesis or metabolism of amino acids. Intriguingly, in our study, we also found many key genes that are linked to fatty acids synthesis, including DEGs (
ACADL; ACSL4; ECHS1; ECI1; ACAT2; HADH; HADHA; ECHS1; ACAA2; GCDH) and analysis of KEGG results showed these genes were enriched in the fatty acid degradation pathway. Wang et al. [
7] identified key genes that are associated with fatty acids synthesis, including
FASN, ACACA, HACD1, FADS2, ACSL1, HACD2, and
ACSBG1 using transcriptome (RNA-Seq) in broiler chicken and KEGG analysis showed that genes were enriched in the pathway of fatty acid biosynthesis and biosynthesis of unsaturated fatty acids. Yang et al. [
8] demonstrated promising candidate genes that regulate fatty acids composition by analyzing chicken transcriptomes from the thigh muscle tissue of Huangshan Black Chicken, including
DCN, FADS2, FRZB, OGN, CHCHD10, PRKAG3, ADGRD1 and
CYTL1, are the most promising candidate genes affecting fatty acids composition using RNA sequencing technology. Remarkably
ACACA can function as an important enzyme synthesized fatty acids.
ACACA is an enzyme that can catalyze the synthesis of malonyl-CoA from two acetyl-CoA molecules and produce fatty acids under the action of fatty acid synthase [
40]. Therefore, our study showed that the gene regulation associated with fatty acid metabolism was significantly greater in the CH chicken breed than the DG chicken breed, indicating the fat tissues in CH chicken have a stronger ability to synthesize the triglycerides cells. Thus, the expression of genes involved in fatty acid metabolism was significantly higher in CH chicken breeds compared to DG chicken breeds, and the content of fatty acids was also significantly higher in CH chicken breeds, implying that CH chicken breast muscles have a greater capacity for triglyceride synthesis. Moreover, our findings are consistent with the results of Zhang et al. [
29], who investigated molecular mechanisms underlying meat quality and muscle carcass traits in the pectoral tissues of Gushi chickens and AA broilers.
The ultimate pH (pHu) trait is more important and more closely identified with chicken meat processing ability and sensory quality, and it is also influenced by the glycogen content of chicken muscle after slaughter. Beauclercq et al. [
9] identified a biomarker that is related to meat pHu and glycogen metabolism by transcriptome analysis using microarray technology in the breast muscle of two chicken lines differentiated by pHu values, including
MORN4 and
MYLIP genes.
Meat color is an essential factor that may influence customers’ buying decisions [
41]. Many studies have reported that color of breast muscle was higher in several indigenous chicken breeds [
42,
43]. These are all changeable in parameters of meat color, maybe back to diluting of haem pigments. Our results are similar to Li et al. [
6] who studied the molecular mechanisms of chicken meat color and taste using RNA-seq-based quantitative transcriptome analysis. The top 10 enriched pathways in his study included glycolysis/gluconeogenesis, carbon metabolism, heme binding, hemoglobin complex and haemoglobin binding. Numbers of DEGs were also identified, such as hemoglobin subunit epsilon (
HBE), hemoglobin subunit epsilon 1 (
HBE1), and
MB, which were upregulated genes related to red meat color. Moreover, KEGG enrichment analysis identified DEGs related to amino acid metabolism all of which are connected with metabolism or amino acid biosynthesis, including
CHDH and
AGXT. The variable in meat color is believed to be caused by a complex interaction between the met-MB control enzyme system and oxidative processes [
44]. The color of meat is determined by the amount of oxy-MB and MB in the muscle, which determines the color, such as red, bright, and pink that consumers may desire. This relationship between oxy-MB and MB content has been investigated [
45,
46].
MB has a key role in color and promotes more than 80% of the pigment of meat, and the
MB gene expression is key to regulating meat color [
47,
48]. The current study suggested that the
MB gene was upregulated and was able to function in oxygen binding and heme-binding, and it may also be related to redness color value. An additional study reported that
MB gene expression was correlated to a∗ values in pork meat [
49]; hemoglobin is a major factor in the formation of red meats color [
50].
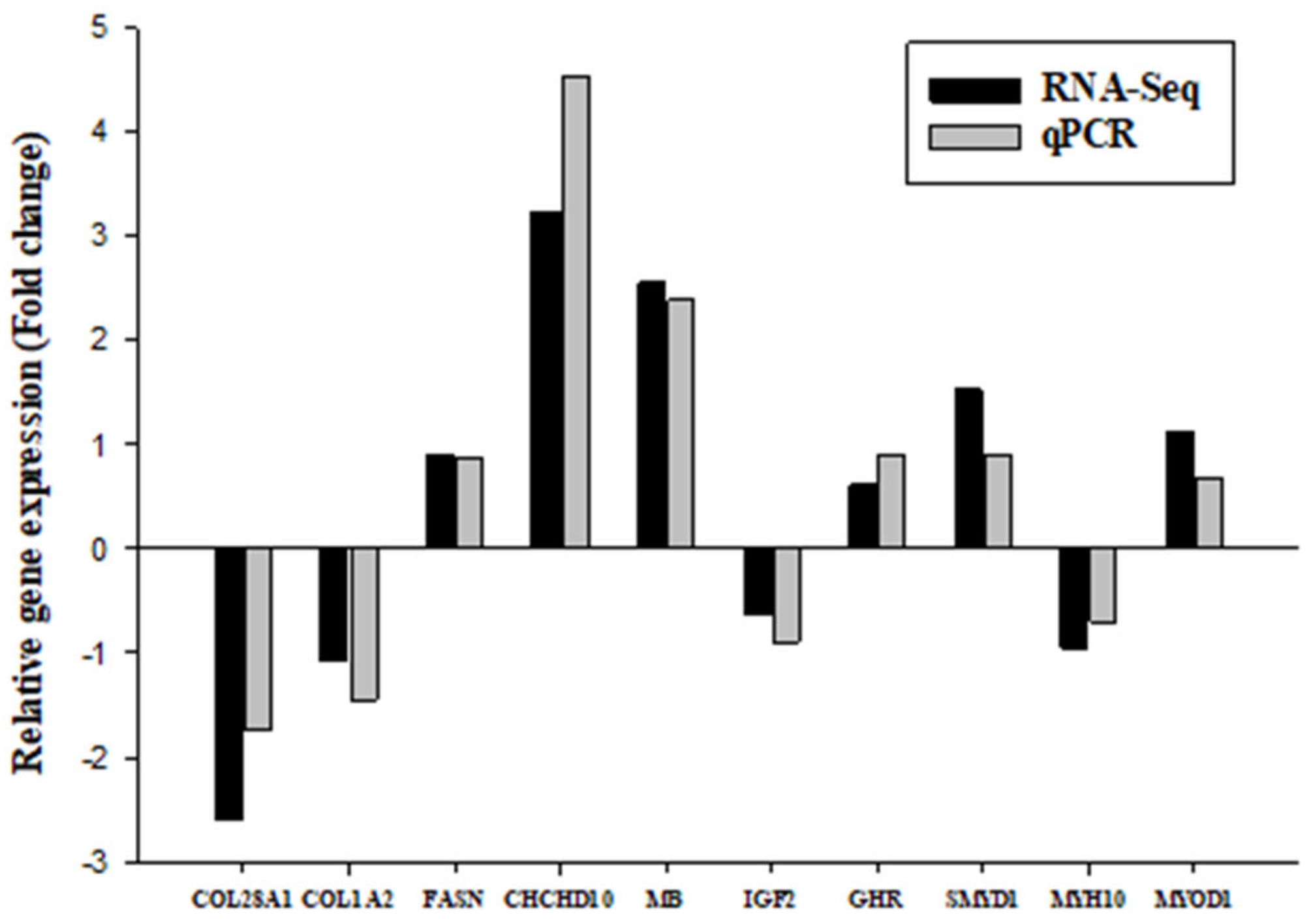
Therefore, MB, HBAD, and HBA1 genes were assumed as key candidate genes that can play an important role in promoting meat color in chickens; they encode subunits of hemoglobin and all of them were upregulated RT-qPCR and RNA-seq results.
5. Conclusions
In conclusion, this study has created the transcriptome profiles of the breast muscle from two chicken breeds (DG c and CH), which have different meat quality at the age of 100 days, using RNA sequencing technology. Successive bioinformatic analyses indicated that some DEGs, such as AGA, COL28A1, COL1A2, MB, HBAD, HBA1, ACACA, ACADL, ACSL1, ATP8A1, CAV1, FADS2, FASN, DCN, CHCHD10, AGXT2, ALDH3A2 and MORN4 and pathways, such as oxidative phosphorylation, carbon metabolism, valine, leucine and isoleucine degradation, fatty acid degradation, glyoxylate and dicarboxylate metabolism, pyruvate metabolism, tryptophan metabolism and glycolysis/gluconeogenesis, might be indispensable for the regulation of meat quality traits. This comparative transcriptome analysis of Digao and Chahua muscle has been reported for the first time. This study is useful in forecasting new gene functions and in the exploration of the mechanism of meat quality traits process in chickens.

 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}