
Genome-Wide Identification and Expression Profile of the HD-Zip Transcription Factor Family Associated with Seed Germination and Abiotic Stress Response in Miscanthus sinensis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification of HD-Zip Genes in M. Sinensis
2.2. Phylogenetic Analysis and Classification of HD-Zip Genes
2.3. Gene Structure and Motif Composition of HD-Zip Genes
2.4. Chromosomal Distribution, Gene Duplication, and Synteny Analysis of HD-Zip Genes
2.5. Plant Growth and Treatments
2.6. RNA Extraction, cDNA Synthesis, and qRT-PCR Gene Expression Analysis
3. Results
3.1. Identification of HD-Zip Genes in M. sinensis
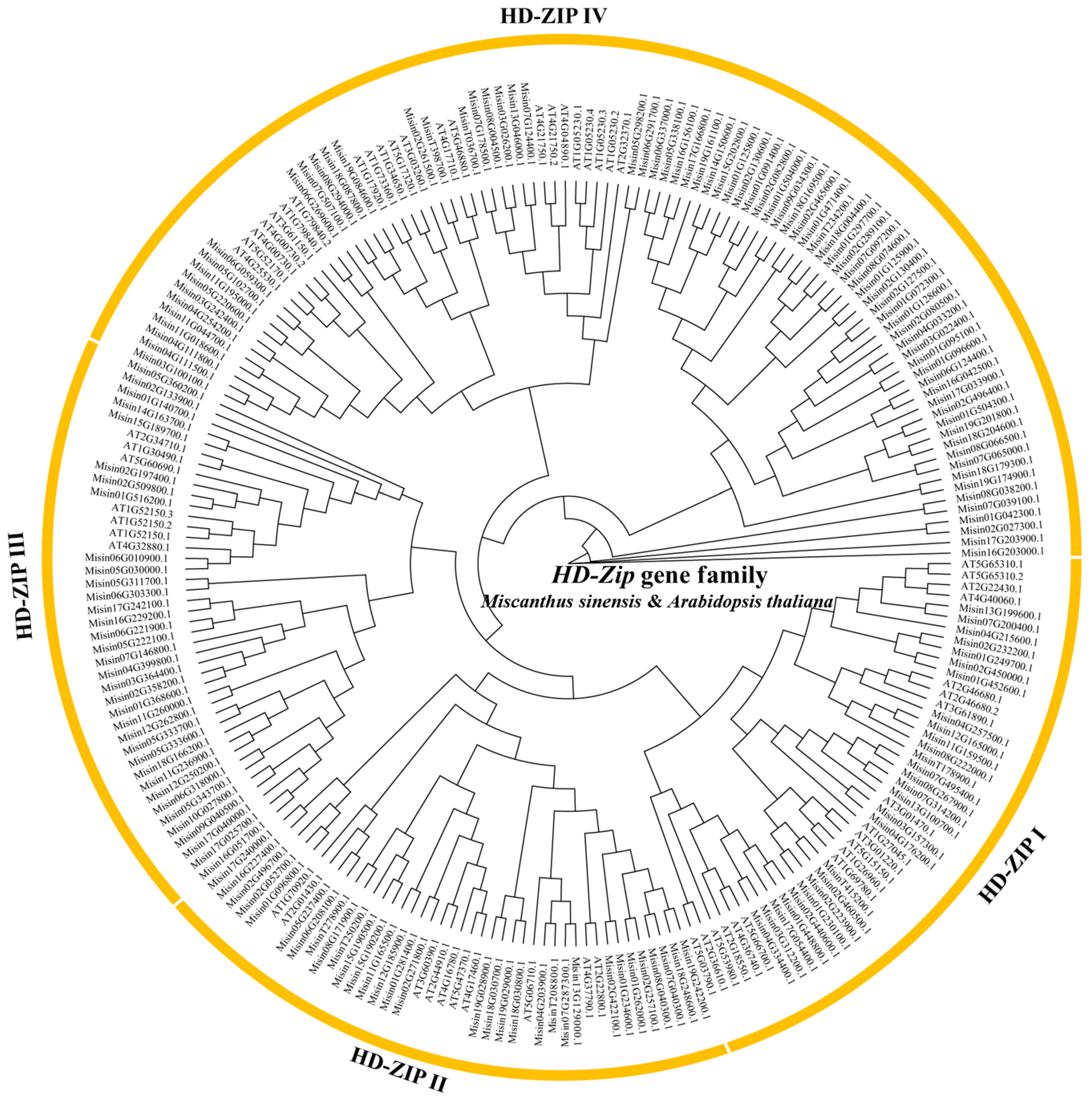
3.2. Phylogenetic Analysis and Classification of HD-Zip Genes
3.3. Gene Structure and Motif Composition of the HD-Zip Gene Family
3.4. Chromosomal Distribution and Gene Duplication of the HD-Zip Genes
3.5. Evolutionary Analysis of HD-Zip Genes in Several Different Species
3.6. Expression Analysis of HD-Zip Genes during Different Seed Germination Stages of M. sinensis
3.7. Expression Patterns of HD-Zip Genes in Response to Abiotic Stress
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Clifton-Brown, J.; Lewandowski, I. Screening Miscanthus genotypes in field trials to optimise biomass yield and quality in Southern Germany. Eur. J. Agron. 2002, 16, 97–110. [Google Scholar] [CrossRef]
- Nie, G.; Liao, Z.; Zhong, M.; Zhou, J.; Cai, J.; Liu, A.; Wang, X.; Zhang, X. MicroRNA-Mediated Responses to Chromium Stress Provide Insight Into Tolerance Characteristics of Miscanthus sinensis. Front. Plant Sci. 2021, 12, 666117. [Google Scholar] [CrossRef]
- Heaton, E.A.; Long, S.P.; Voigt, T.B.; Jones, M.B.; Clifton-Brown, J. Miscanthus for Renewable Energy Generation: European Union Experience and Projections for Illinois. Mitig. Adapt. Strat. Glob. Chang. 2004, 9, 433–451. [Google Scholar] [CrossRef]
- Nie, G.; Huang, L.; Ma, X.; Ji, Z.; Zhang, Y.; Tang, L.; Zhang, X. Enriching Genomic Resources and Transcriptional Profile Analysis of Miscanthus sinensis under Drought Stress Based on RNA Sequencing. Int. J. Genom. 2017, 2017, 9184731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nsanganwimana, F.; Waterlot, C.; Louvel, B.; Pourrut, B.; Douay, F. Metal, nutrient and biomass accumulation during the growing cycle of Miscanthus established on metal-contaminated soils. J. Soil Sci. Plant Nutr. 2016, 179, 257–269. [Google Scholar] [CrossRef]
- Ye, B.; Saito, A.; Minamisawa, K. Effect of Inoculation with Anaerobic Nitrogen-Fixing Consortium on Salt Tolerance of Miscanthus sinensis. Soil Sci. Plant Nutr. 2005, 51, 243–249. [Google Scholar] [CrossRef] [Green Version]
- Ezaki, B.; Nagao, E.; Yamamoto, Y.; Nakashima, S.; Enomoto, T. Wild plants, Andropogon virginicus L. and Miscanthus sinensis Anders, are tolerant to multiple stresses including aluminum, heavy metals and oxidative stresses. Plant Cell Rep. 2008, 27, 951–961. [Google Scholar] [CrossRef]
- Aso, T. Studies on the Germination of Seeds of Miscanthus sinensis ANDERSS. Science Reports of the Yokohama National University, Section II. Biol. Geology 1976, 23, 27–34. [Google Scholar]
- Bewley, J.D. Seed Germination and Dormancy. Plant Cell 1997, 9, 1055–1066. [Google Scholar] [CrossRef] [Green Version]
- Christian, E.J.; Goggi, A.S.; Moore, K.J. Temperature and Light Requirements for Miscanthus sinensis Laboratory Germination Test. Crop Sci. 2014, 54, 791–795. [Google Scholar] [CrossRef] [Green Version]
- Awty-Carroll, D.; Clifton-Brown, J.; Robson, P. Using k-NN to analyse images of diverse germination phenotypes and detect single seed germination in Miscanthus sinensis. Plant Methods 2018, 14, 5. [Google Scholar] [CrossRef] [PubMed]
- Nie, G.; Sun, M.; Huang, L.; Ma, X.; Zhang, X. Effect of moist pre-chill and dry pre-heat treatment on the germination of Miscanthus sinensis seed from southwest China. Grassl. Sci. 2017, 63, 93–100. [Google Scholar] [CrossRef]
- Clifton-Brown, J.; McCalmont, J.; Hastings, A. Development of Miscanthus as a bioenergy crop: Biofuels and bioenergy. In Biofuels and Bioenergy; Elsevier: Amsterdam, The Netherlands, 2017. [Google Scholar]
- Clifton-Brown, J.; Schwarz, K.-U.; Hastings, A. History of the development of Miscanthus as a bioenergy crop: From small beginnings to potential realisation. In Biology and Environment: Proceedings of the Royal Irish Academy; Royal Irish Academy: Dublin, Ireland, 2015; Volume 115B, p. 45. [Google Scholar] [CrossRef]
- Yang, Y.; Al-Baidhani, H.H.J.; Harris, J.; Riboni, M.; Li, Y.; Mazonka, I.; Bazanova, N.; Chirkova, L.; Hussain, S.S.; Hrmova, M.; et al. DREB/CBF expression in wheat and barley using the stress-inducible promoters of HD-Zip I genes: Impact on plant development, stress tolerance and yield. Plant Biotechnol. J. 2020, 18, 829–844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, J.C.; Hrmova, M.; Lopato, S.; Langridge, P. Modulation of plant growth by HD-Zip class I and II transcription factors in response to environmental stimuli. New Phytol. 2011, 190, 823–837. [Google Scholar] [CrossRef]
- Elhiti, M.; Stasolla, C. Structure and function of homodomain-leucine zipper (HD-Zip) proteins. Plant Signal. Behav. 2009, 4, 86–88. [Google Scholar] [CrossRef] [Green Version]
- Ariel, F.D.; Manavella, P.A.; Dezar, C.A.; Chan, R.L. The true story of the HD-Zip family. Trends Plant Sci. 2007, 12, 419–426. [Google Scholar] [CrossRef]
- Tognacca, R.S.; Carabelli, M.; Morelli, G.; Ruberti, I.; Botto, J.F. Author Correction: ATHB2 is a negative regulator of germination in Arabidopsis thaliana seeds. Sci. Rep. 2021, 11, 12879. [Google Scholar] [CrossRef]
- Seo, M.; Kanno, Y.; Frey, A.; North, H.M.; Marion-Poll, A. Dissection of Arabidopsis NCED9 promoter regulatory regions reveals a role for ABA synthesized in embryos in the regulation of GA-dependent seed germination. Plant Sci. 2016, 246, 91–97. [Google Scholar] [CrossRef]
- Ni, Y.; Wang, X.; Li, D.; Wu, Y.; Xu, W.; Li, X. Novel cotton homeobox gene and its expression profiling in root development and in response to stresses and phytohormones. Acta Biochim. Biophys. Sin. 2010, 40, 78–84. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.-K.; Son, O.; Kim, M.-R.; Nam, K.-H.; Kim, G.-T.; Lee, M.-S.; Choi, S.-Y.; Cheon, C.-I. ATHB23, an Arabidopsis class I homeodomain-leucine zipper gene, is expressed in the adaxial region of young leaves. Plant Cell Rep. 2007, 26, 1179–1185. [Google Scholar] [CrossRef]
- Depège-Fargeix, N.; Javelle, M.; Chambrier, P.; Frangne, N.; Gerentes, D.; Perez, P.; Rogowsky, P.M.; Vernoud, V. Functional characterization of the HD-ZIP IV transcription factor OCL1 from maize. J. Exp. Bot. 2011, 62, 293–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Zhang, S.; Dong, R.; Wang, L.; Yao, J.; van Nocker, S.; Wang, X. The grapevine homeobox gene VvHB58 influences seed and fruit development through multiple hormonal signaling pathways. BMC Plant Biol. 2019, 19, 523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohashi-Ito, K.; Kubo, M.; Demura, T.; Fukuda, H. Class III Homeodomain Leucine-Zipper Proteins Regulate Xylem Cell Differentiation. Plant Cell Physiol. 2005, 46, 1646–1656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Haider, I.; Kohlen, W.; Jiang, L.; Bouwmeester, H.; Meijer, A.H.; Schluepmann, H.; Liu, C.-M.; Ouwerkerk, P.B.F. Function of the HD-Zip I gene Oshox22 in ABA-mediated drought and salt tolerances in rice. Plant Mol. Biol. 2012, 80, 571–585. [Google Scholar] [CrossRef] [PubMed]
- Park, M.Y.; Kim, S.-A.; Lee, S.-J.; Kim, S.Y. ATHB17 is a positive regulator of abscisic acid response during early seedling growth. Mol. Cells 2013, 35, 125–133. [Google Scholar] [CrossRef] [Green Version]
- Zhou, C.-P.; Li, C.-P.; Liang, W.-W.; Guo, P.; Yang, L.-T.; Chen, L.-S. Identification of manganese-toxicity-responsive genes in roots of two citrus species differing in manganese tolerance using cDNA-AFLP. Trees 2017, 31, 813–831. [Google Scholar] [CrossRef]
- Hu, T.; Ye, J.; Tao, P.; Li, H.; Zhang, J.; Zhang, Y.; Ye, Z. The tomato HD-Zip I transcription factor SlHZ24 modulates ascorbate accumulation through positive regulation of thed-mannose/l-galactose pathway. Plant J. 2016, 85, 16–29. [Google Scholar] [CrossRef] [Green Version]
- Chen, E.; Zhang, X.; Yang, Z.; Wang, X.; Yang, Z.; Zhang, C.; Wu, Z.; Kong, D.; Liu, Z.; Zhao, G.; et al. Genome-wide analysis of the HD-ZIP IV transcription factor family in Gossypium arboreum and GaHDG11 involved in osmotic tolerance in transgenic Arabidopsis. Genet. Mol. Res. 2017, 292, 593–609. [Google Scholar] [CrossRef]
- Ding, Y.F.; Gong, S.H.; Wang, Y.; Wang, F.J.; Bao, H.X.G.D.L.; Sun, J.W.; Cai, C.; Yi, K.K.; Chen, Z.X.; Zhu, C. MicroRNA166 Modulates Cadmium Tolerance and Accumulation in Rice. Plant Physiol. 2018, 177, 1691–1703. [Google Scholar] [CrossRef] [Green Version]
- Mitros, T.; Session, A.M.; James, B.T.; Wu, G.A.; Belaffif, M.B.; Clark, L.V.; Shu, S.; Dong, H.; Barling, A.; Holmes, J.R.; et al. Genome biology of the paleotetraploid perennial biomass crop Miscanthus. Nat. Commun. 2020, 11, 5442. [Google Scholar] [CrossRef]
- Wilkins, M.R.; Gasteiger, E.; Bairoch, A.; Sanchez, J.C.; Williams, K.L.; Appel, R.D.; Hochstrasser, D.F. Protein Identification and Analysis Tools in the ExPASy Server. In 2-D Proteome Analysis Protocols. Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 1999; Volume 112, pp. 531–552. [Google Scholar] [CrossRef]
- Zhuo, X.; Zheng, T.; Zhang, Z.; Zhang, Y.; Jiang, L.; Ahmad, S.; Sun, L.; Wang, J.; Cheng, T.; Zhang, Q. Genome-Wide Analysis of the NAC Transcription Factor Gene Family Reveals Differential Expression Patterns and Cold-Stress Responses in the Woody Plant Prunus mume. Genes 2018, 9, 494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Rhee, S.Y.; Beavis, W.; Berardini, T.Z.; Chen, G.; Dixon, D.; Doyle, A.; Garcia-Hernandez, M.; Huala, E.; Lander, G.; Montoya, M.; et al. The Arabidopsis Information Resource (TAIR): A model organism database providing a centralized, curated gateway to Arabidopsis biology, research materials and community. Nucleic Acids Res. 2003, 31, 224–228. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.-T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2013, 14, 587–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoang, D.T.; Chernomor, O.; Haeseler, A.v.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the Ultrafast Bootstrap Approximation. Mol. Biol. Evol. 2017, 35, 518–522. [Google Scholar] [CrossRef]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME Suite. Nucleic Acids Res. 2015, 43, W39–W49. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Tang, H.; DeBarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.-H.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.J.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.H.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Zhong, M.; Yang, X.; Hu, Y.; Huang, L.; Peng, Y.; Li, Z.; Liu, Q.; Wang, X.; Zhang, X.; Nie, G. Identification of candidate reference genes for quantitative RT-PCR in Miscanthus sinensis subjected to various abiotic stresses. Mol. Biol. Rep. 2020, 47, 2913–2927. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Henriksson, E.; Olsson, A.S.; Johannesson, H.; Hanson, J.; Engström, P.; Söderman, E. Homeodomain Leucine Zipper Class I Genes in Arabidopsis. Expression Patterns and Phylogenetic Relationships. Plant Physiol. 2005, 139, 509–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, M.; Liu, A.; Zhang, Y.; Zhou, Y.; Li, D.; Dossa, K.; Zhou, R.; Zhang, X.; You, J. Genome-wide characterization and expression analysis of the HD-Zip gene family in response to drought and salinity stresses in sesame. BMC Genom. 2019, 20, 748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayer, K.F.X.; Waugh, R.; Langridge, P.; Close, T.J.; Wise, R.P.; Graner, A.; Matsumoto, T.; Sato, K.; Schulman, A.; Muehlbauer, G.J.; et al. A physical, genetic and functional sequence assembly of the barley genome. Nature 2012, 491, 711–716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Xiong, H.; Cuo, D.; Wu, X.; Duan, R. Genome-wide characterization and expression profiling of the relation of the HD-Zip gene family to abiotic stress in barley (Hordeum vulgare L.). Plant Physiol. Biochem. 2019, 141, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Chai, W.; Si, W.; Ji, W.; Qin, Q.; Zhao, M.; Jiang, H. Genome-Wide Investigation and Expression Profiling of HD-Zip Transcription Factors in Foxtail Millet (Setaria italica L.). BioMed Res. Int. 2018, 2018, 8457614. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Liu, X.; Quan, Z.; Cheng, S.; Xu, X.; Pan, S.; Xie, M.; Zeng, P.; Yue, Z.; Wang, W.; et al. Genome sequence of foxtail millet (Setaria italica) provides insights into grass evolution and biofuel potential. Nat. Biotechnol. 2012, 30, 549–554. [Google Scholar] [CrossRef] [Green Version]
- The Arabidopsis Genome Initiative. Analysis of the genome sequence of the flowering plant Arabidopsis thaliana. Nature 2000, 408, 796–815. [Google Scholar] [CrossRef] [Green Version]
- Cannon, S.B.; Mitra, A.; Baumgarten, A.; Young, N.D.; May, G. The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana. BMC Plant Biol. 2004, 4, 10. [Google Scholar] [CrossRef]
- Tian, R.; Yang, Y.; Chen, M. Genome-wide survey of the amino acid transporter gene family in wheat (Triticum aestivum L.): Identification, expression analysis and response to abiotic stress. Int. J. Biol. Macromol. 2020, 162, 1372–1387. [Google Scholar] [CrossRef]
- Liu, M.; Ma, Z.; Sun, W.; Huang, L.; Wu, Q.; Tang, Z.; Bu, T.; Li, C.; Chen, H. Genome-wide analysis of the NAC transcription factor family in Tartary buckwheat (Fagopyrum tataricum). BMC Genom. 2019, 20, 113. [Google Scholar] [CrossRef] [Green Version]
- Prigge, M.J.; Otsuga, D.; Alonso, J.M.; Ecker, J.R.; Drews, G.N.; Clark, S.E. Class III Homeodomain-Leucine Zipper Gene Family Members Have Overlapping, Antagonistic, and Distinct Roles in Arabidopsis Development. Plant Cell 2005, 17, 61–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rombolá-Caldentey, B.; Rueda-Romero, P.; Iglesias-Fernández, R.; Carbonero, P.; Oñate-Sánchez, L. Arabidopsis DELLA and Two HD-ZIP Transcription Factors Regulate GA Signaling in the Epidermis through the L1 Box cis-Element. Plant Cell 2014, 26, 2905–2919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi-Shinozaki, K.; Shinozaki, K. Transcriptional regulatory networks in cellular responses and tolerance to dehydration and cold stresses. Annu. Rev. Plant Biol. 2006, 57, 781–803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Zhu, R.; Ji, X.; Li, H.J.; Lv, H.; Zhang, H.Y. Genome-Wide Characterization and Expression Analysis of the HD-ZIP Gene Family in Response to Salt Stress in Pepper. Int. J. Genom. 2021, 2021, 8105124. [Google Scholar] [CrossRef]
- Zhao, S.; Gao, H.; Jia, X.; Wang, H.; Ke, M.; Ma, F. The HD-Zip I transcription factor MdHB-7 regulates drought tolerance in transgenic apple (Malus domestica). Environ. Exp. Bot. 2020, 180, 104246. [Google Scholar] [CrossRef]
- Hong, Y.; Liu, Y.; Zhang, Y.; Jia, L.; Yang, X.; Zhang, X.; Liu, J.; Luan, Y. Genome-wide characterization of homeobox-leucine zipper gene family in tomato (Solanum lycopersicum) and functional analysis of SlHDZ34 (III sub-family member) under salinity stress. Environ. Exp. Bot. 2021, 192, 104652. [Google Scholar] [CrossRef]
- Ebrahimian-Motlagh, S.; Ribone, P.A.; Thirumalaikumar, V.P.; Allu, A.D.; Chan, R.L.; Mueller-Roeber, B.; Balazadeh, S. JUNGBRUNNEN1 Confers Drought Tolerance Downstream of the HD-Zip I Transcription Factor AtHB13. Front. Plant Sci. 2017, 8, 2118. [Google Scholar] [CrossRef] [Green Version]
- Cabello, J.V.; Chan, R.L. The homologous homeodomain-leucine zipper transcription factors HaHB1 and AtHB13 confer tolerance to drought and salinity stresses via the induction of proteins that stabilize membranes. Plant Biotechnol. J. 2012, 10, 815–825. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, H.; Srivastava, A.K.; Pan, Y.; Bai, J.; Fang, J.; Shi, H.; Zhu, J.-K.; Zhang, J.; Zhang, H.; et al. Knockdown of Rice MicroRNA166 Confers Drought Resistance by Causing Leaf Rolling and Altering Stem Xylem Development. Plant Physiol. 2018, 176, 2082–2094. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Gao, Z.; Li, R.; Xu, Y.; Kong, Y.; Zhou, G.; Meng, C.; Hu, R. Genome-wide identification and expression profiling of HD-ZIP gene family in Medicago truncatula. Genomics 2020, 112, 3624–3635. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, J.; Ran, Q.; Yang, Z.; Zhou, Y.; Yuan, Z.; Lai, H.; Wang, J.; Nie, G.; Zhu, Y. Genome-Wide Identification and Expression Profile of the HD-Zip Transcription Factor Family Associated with Seed Germination and Abiotic Stress Response in Miscanthus sinensis. Genes 2022, 13, 2256. https://doi.org/10.3390/genes13122256
Chen J, Ran Q, Yang Z, Zhou Y, Yuan Z, Lai H, Wang J, Nie G, Zhu Y. Genome-Wide Identification and Expression Profile of the HD-Zip Transcription Factor Family Associated with Seed Germination and Abiotic Stress Response in Miscanthus sinensis. Genes. 2022; 13(12):2256. https://doi.org/10.3390/genes13122256
Chicago/Turabian StyleChen, Jiayi, Qifan Ran, Zhongfu Yang, Ying Zhou, Zhihang Yuan, Huiqi Lai, Jing Wang, Gang Nie, and Yongqun Zhu. 2022. "Genome-Wide Identification and Expression Profile of the HD-Zip Transcription Factor Family Associated with Seed Germination and Abiotic Stress Response in Miscanthus sinensis" Genes 13, no. 12: 2256. https://doi.org/10.3390/genes13122256