

Hyperpigmentation Inhibits Early Skeletal Muscle Development in Tengchong Snow Chicken Breed
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Experimentation Ethical Statement
2.2. Chicken, Diet and Housing
2.3. Slaughter Procedure and Sample Collecting
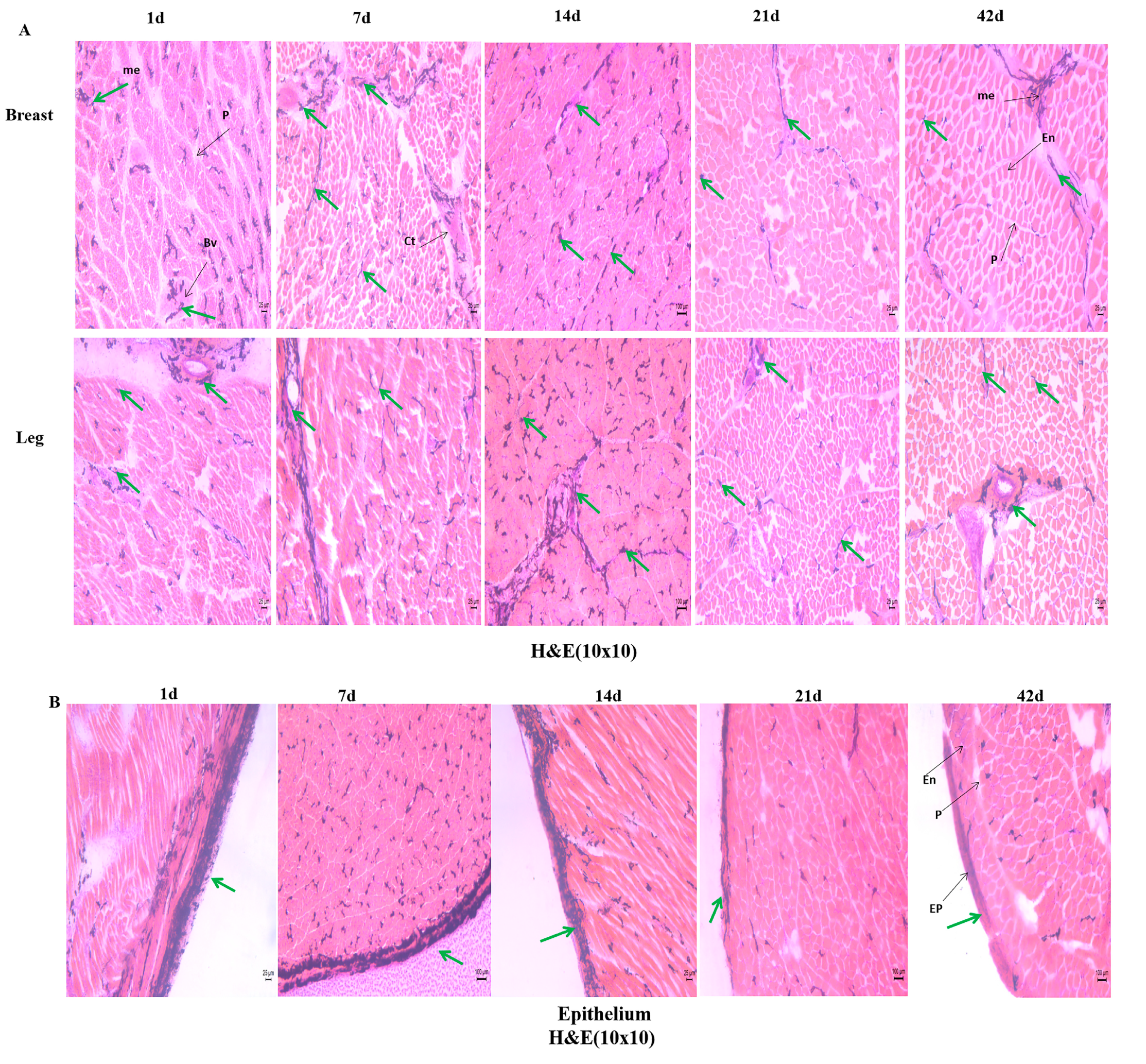
2.4. Histological Study
2.4.1. Cryosectioning and Hematoxylin and Eosin Staining
2.4.2. ATPase Staining
2.5. mRNA Expression Analysis
2.6. Data and Statistical Analysis
3. Results
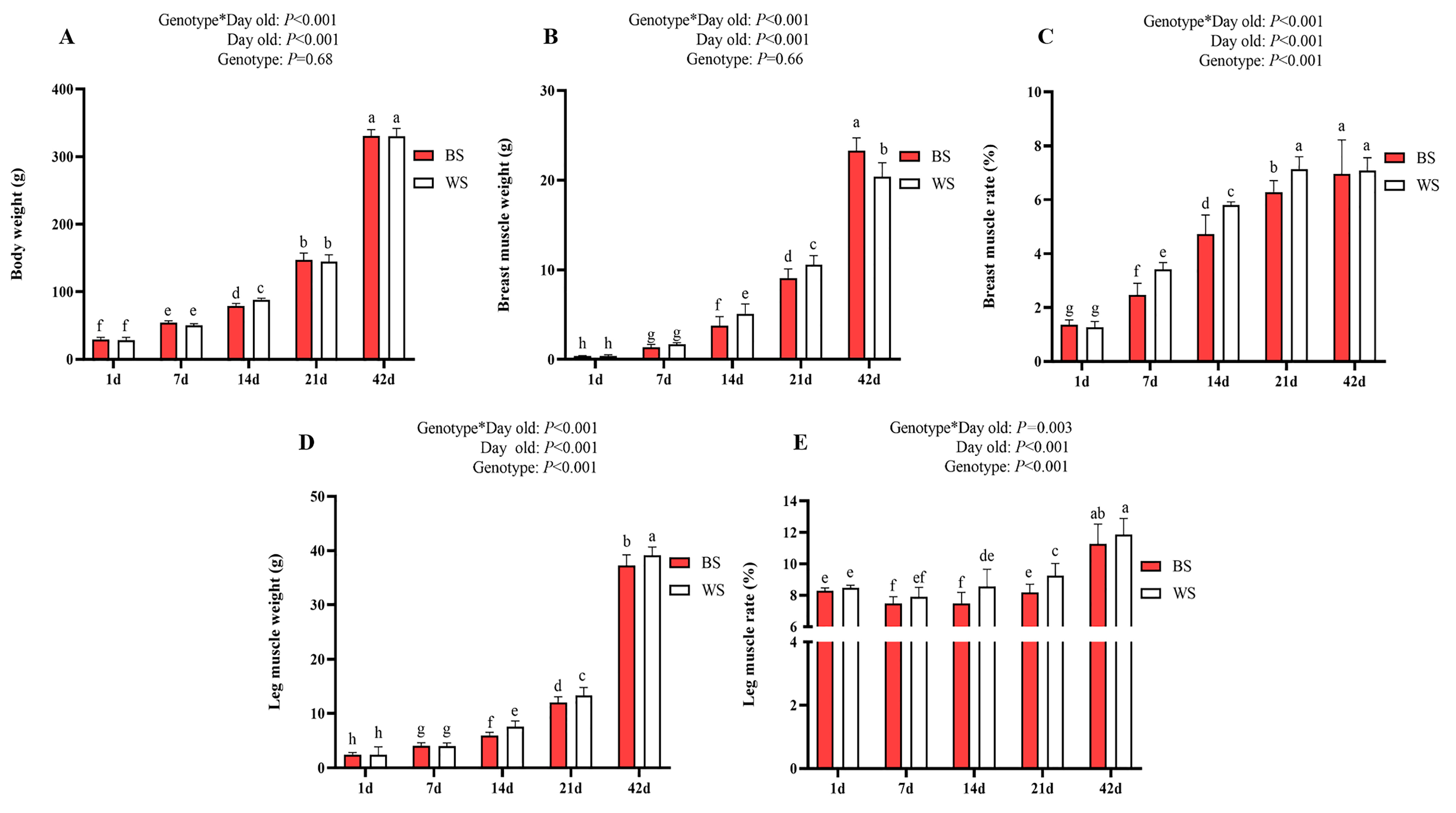
3.1. Comparative Analysis of Early Development in Skeletal Muscle
3.2. Comparative Analysis of Skeletal Muscle Fiber
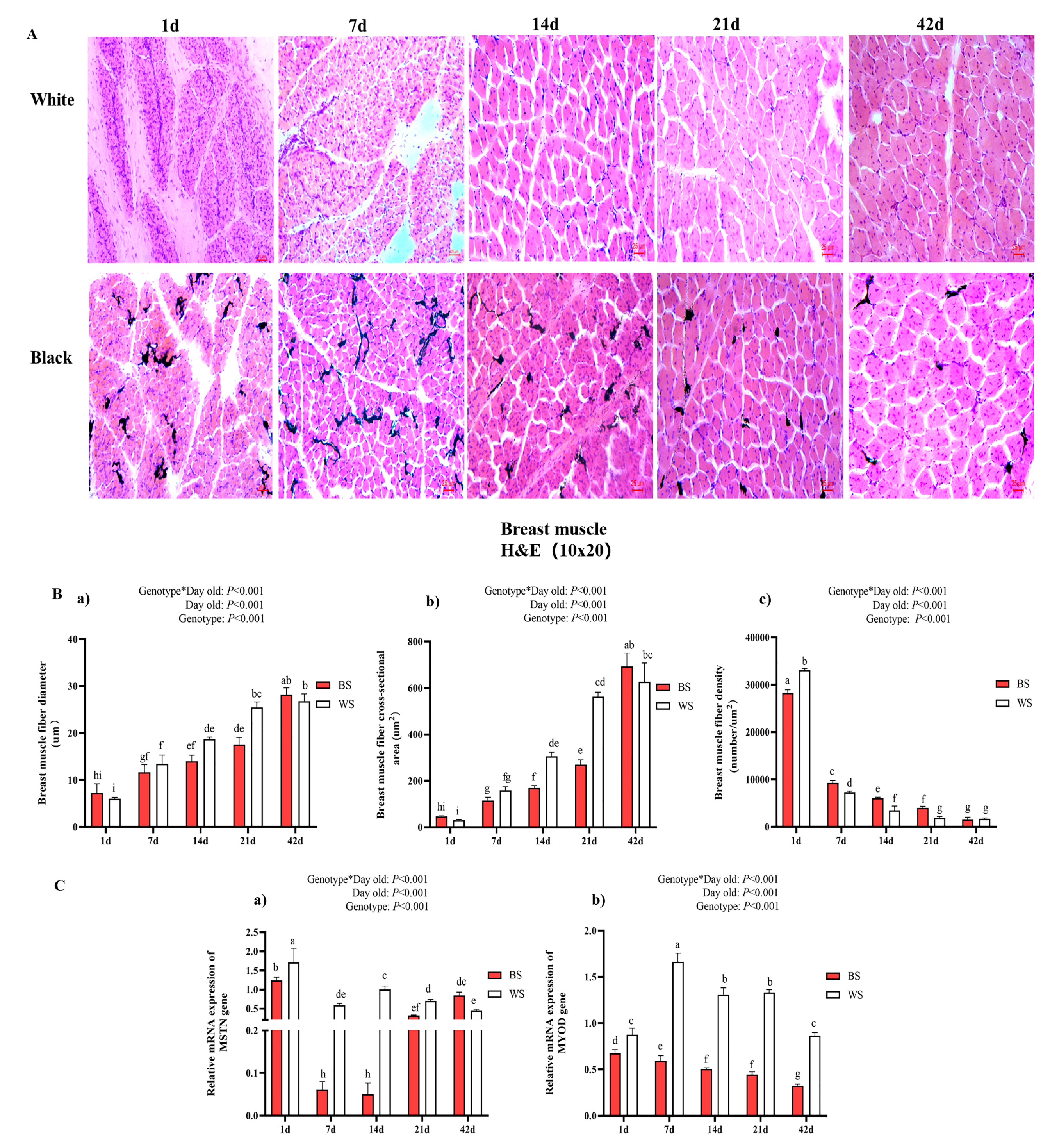
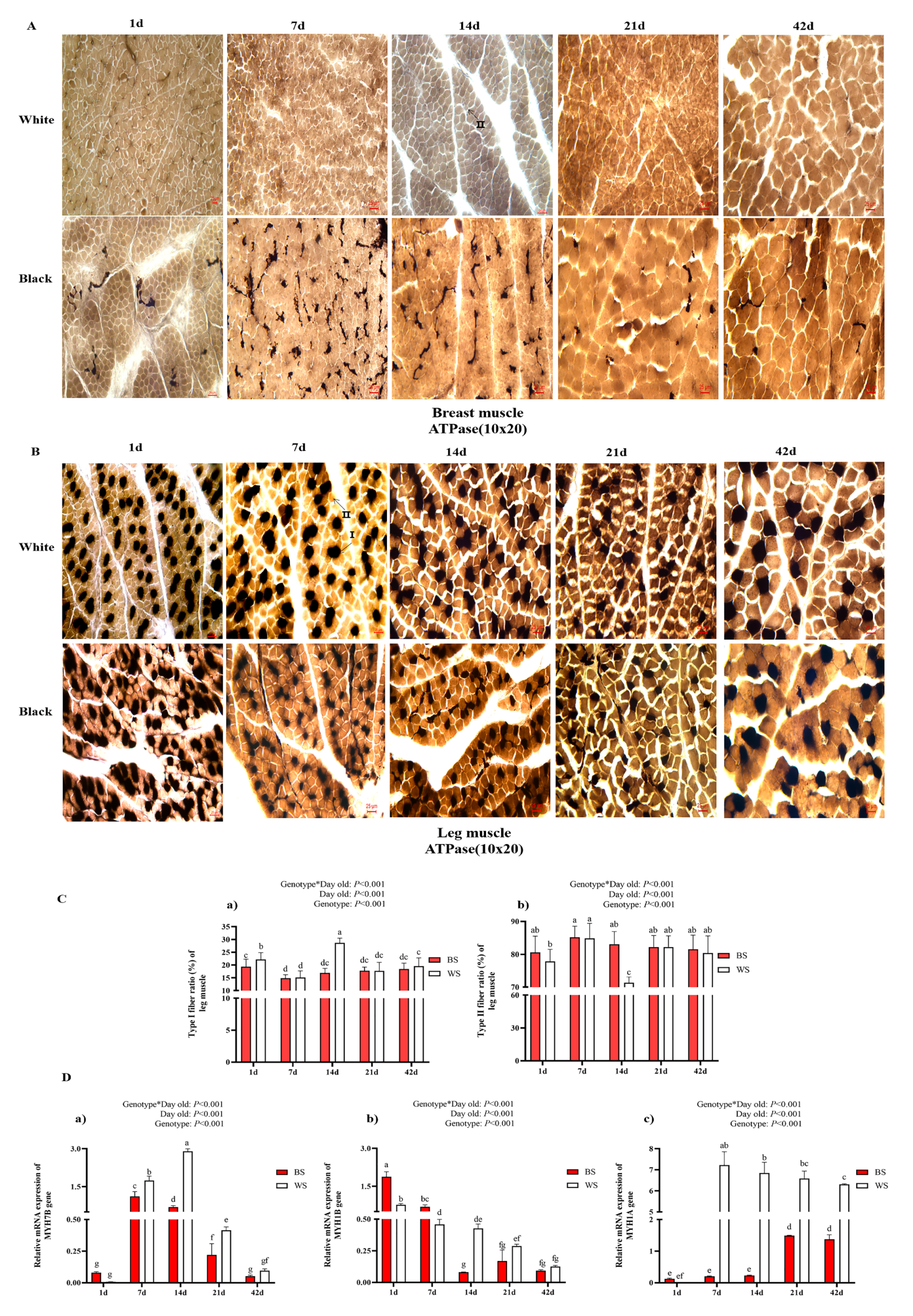
3.2.1. Comparative Analysis of Breast Muscle Fiber
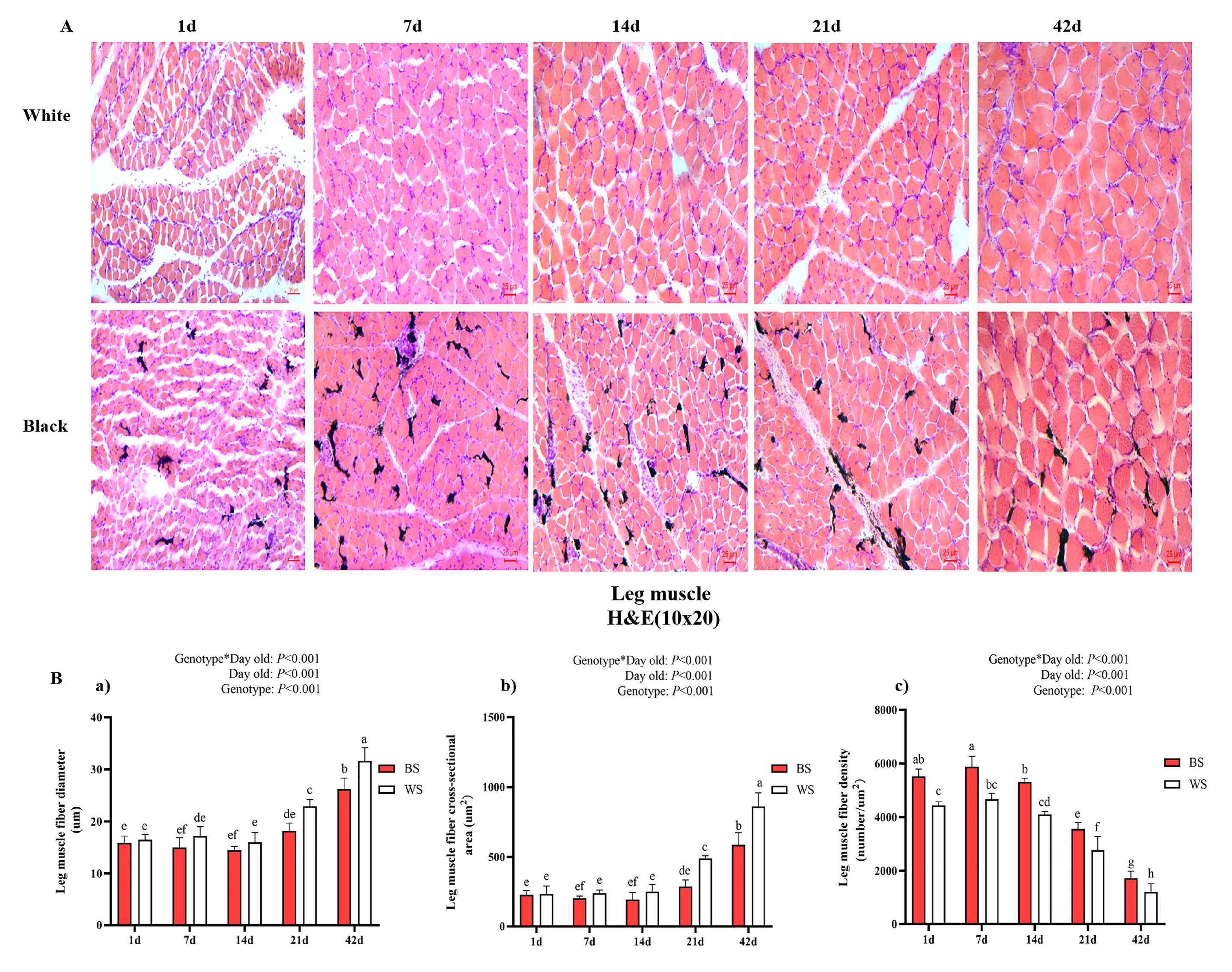
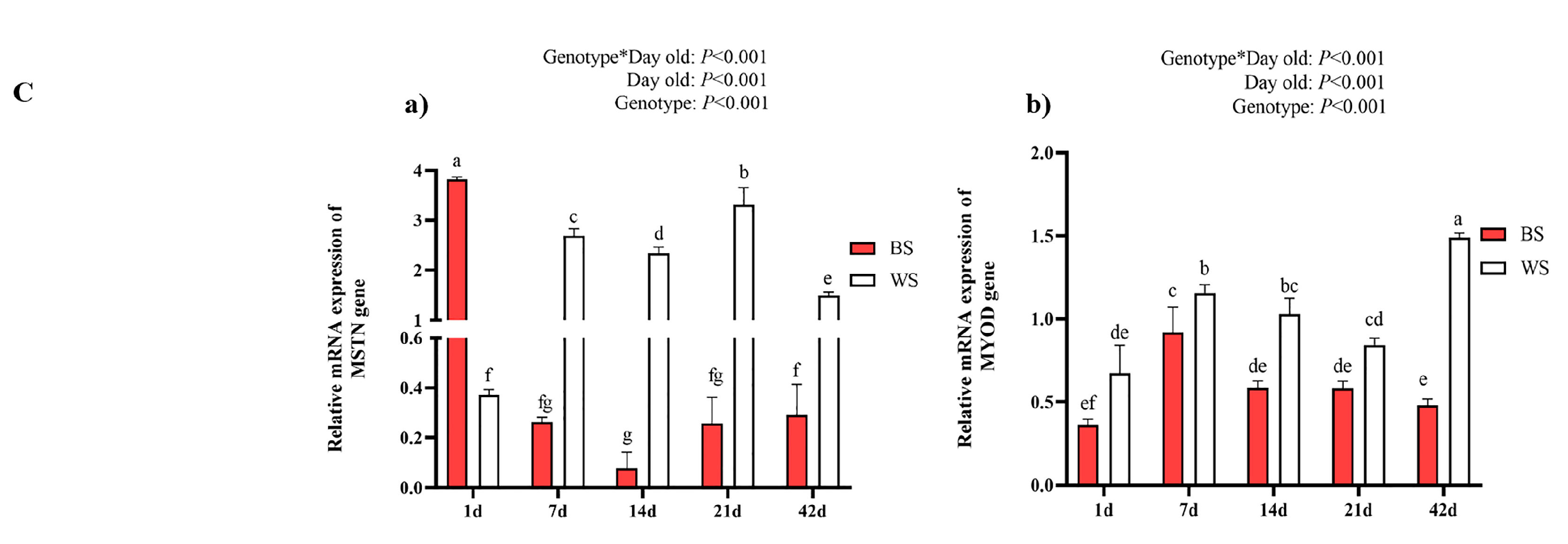
3.2.2. Comparative Analysis of Leg Muscle Fiber
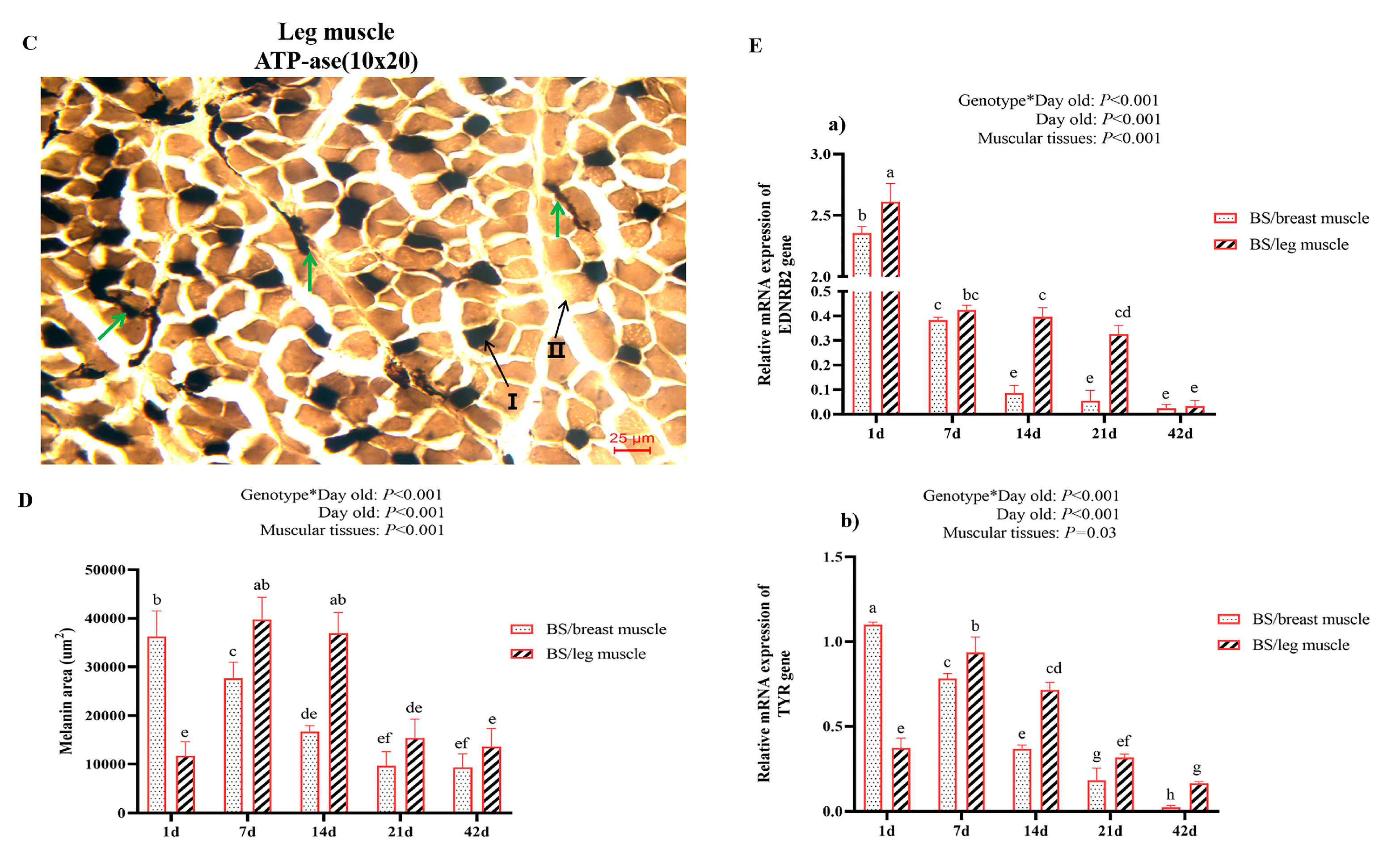
3.2.3. Comparative Analysis of the Leg Muscle Fiber Types
3.2.4. Melanin Distribution of the Breast and Leg Muscles in BS
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, D.; Sun, G.; Zhang, M.; Cao, Y.; Zhang, C.; Fu, Y.; Li, F.; Li, G.; Jiang, R.; Han, R.; et al. Breeding history and candidate genes responsible for black skin of Xichuan black-bone chicken. BMC Genom. 2020, 21, 511. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Wang, G.; Liao, J.; Tang, M. Transcriptome profile analysis identifies candidate genes for the melanin pigmentation of breast muscle in Muchuan black-boned chicken. Poult. Sci. 2018, 97, 3446–3455. [Google Scholar] [CrossRef] [PubMed]
- Jaturasitha, S.; Srikanchai, T.; Kreuzer, M.; Wicke, M. Differences in Carcass and Meat Characteristics Between Chicken Indigenous to Northern Thailand (Black-Boned and Thai Native) and Imported Extensive Breeds (Bresse and Rhode Island Red). Poult. Sci. 2008, 87, 160–169. [Google Scholar] [CrossRef]
- Tian, Y.; Sheng, Z.; Xie, M.; Wang, W.; Wu, H.; Gong, D. Composition of fatty acids in the muscle of black-bone silky chicken (Gallus gellus demesticus brissen) and its bioactivity in mice. Food Chem. 2011, 126, 479–483. [Google Scholar] [CrossRef]
- Han, D.; Wang, S.; Hu, Y.; Zhang, Y.; Dong, X.; Yang, Z.; Wang, J.; Li, J.; Deng, X. Hyperpigmentation Results in Aberrant Immune Development in Silky Fowl (Gallus gallus domesticus Brisson). PLoS ONE 2015, 10, e125686. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Zhou, H.; Han, C.; Xu, Z.; Ding, J.; Zhu, J.; Qin, C.; Luo, H.; Chen, K.; Jiang, S.; et al. Transcriptomic Analysis of MSTN Knockout in the Early Differentiation of Chicken Fetal Myoblasts. Genes 2021, 13, 58. [Google Scholar] [CrossRef] [PubMed]
- Muroya, S.; Tanabe, R.; Nakajima, I.; Chikuni, K. Molecular characteristics and site specific distribution of the pigment of the silky fowl. J. Vet. Med. Sci. 2000, 62, 391–395. [Google Scholar] [CrossRef] [Green Version]
- Ren, L.; Liu, A.; Wang, Q.; Wang, H.; Dong, D.; Liu, L. Transcriptome analysis of embryonic muscle development in Chengkou Mountain Chicken. BMC Genom. 2021, 22, 431. [Google Scholar] [CrossRef]
- Ding, S.; Nie, Y.; Zhang, X.; Liu, X.; Wang, C.; Yuan, R.; Chen, K.; Zhu, Q.; Cai, S.; Fang, Y.; et al. The SNPs in myoD gene from normal muscle developing individuals have no effect on muscle mass. BMC Genet. 2019, 20, 72. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.Q.; Qing, Y.; Zhu, Q.; Zhao, X.L.; Wang, Y.; Li, D.Y.; Liu, Y.P.; Yin, H.D. Genetic effects of polymorphisms in myogenic regulatory factors on chicken muscle fiber traits. Asian-Australas. J. Anim. Sci. 2015, 28, 782–787. [Google Scholar] [CrossRef]
- Yamamoto, M.; Legendre, N.P.; Biswas, A.A.; Lawton, A.; Yamamoto, S.; Tajbakhsh, S.; Kardon, G.; Goldhamer, D.J. Loss of MyoD and Myf5 in Skeletal Muscle Stem Cells Results in Altered Myogenic Programming and Failed Regeneration. Stem Cell Rep. 2018, 10, 956–969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, K.; Han, C.X.; Zhou, H.; Ding, J.M.; Xu, Z.; Yang, L.Y.; He, C.; Akinyemi, F.; Zheng, Y.M.; Qin, C.; et al. Effective MSTN Gene Knockout by AdV-Delivered CRISPR/Cas9 in Postnatal Chick Leg Muscle. Int. J. Mol. Sci. 2020, 21, 2584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.S.; Bobbili, N.K.; Paek, K.S.; Jin, H.J. Production of a monoclonal anti-myostatin antibody and the effects of in ovo administration of the antibody on posthatch broiler growth and muscle mass. Poult. Sci. 2006, 85, 1062–1071. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.D.; Lee, J.H.; Song, S.; Kim, S.W.; Han, J.S.; Shin, S.P.; Park, B.C.; Park, T.S. Generation of myostatin-knockout chickens mediated by D10A-Cas9 nickase. FASEB J. 2020, 34, 5688–5696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, Y.F.; Ding, Q.L.; Li, Y.M.; Fang, W.R. Identification of Differentially Expressed Genes and Pathways for Myofiber Characteristics in Soleus Muscles between Chicken Breeds Differing in Meat Quality. Anim. Biotechnol. 2017, 28, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Huo, W.; Weng, K.; Li, Y.; Zhang, Y.; Zhang, Y.; Xu, Q.; Chen, G. Comparison of muscle fiber characteristics and glycolytic potential between slow- and fast-growing broilers. Poult. Sci. 2022, 101, 101649. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.A.; Wang, Z.; Yang, X.; Ma, M.; Li, Z.; Nie, Q. LncRNA-FKBP1C regulates muscle fiber type switching by affecting the stability of MYH1B. Cell Death Discov. 2021, 7, 73. [Google Scholar] [CrossRef]
- Zeng, Y.T.; Wang, C.; Zhang, Y.; Xu, L.; Zhou, G.B.; Zeng, C.J.; Zuo, Z.C.; Song, T.Z.; Zhu, Q.; Yin, H.D.; et al. Improvac immunocastration affects the development of thigh muscles but not pectoral muscles in male chickens. Poult. Sci. 2020, 99, 5149–5157. [Google Scholar] [CrossRef]
- Zhao, T.J. A Preliminary Study on Breed Preservation and Development of Tengchong Snow Chicken. China Anim. Ind. 2018, 3, 54–56. (In Chinese) [Google Scholar]
- Ou, C.H.; Li, R.Q.; Ding, Z.B.; Ye, S.H.; Duan, G. Nutrient composition analysis of Tengchong snow meat at different ages and sexes. J. Southwest Minzu Univ. Nat. Sci. Ed. 1999, 77–81. (In Chinese) [Google Scholar]
- Wang, R.Q.; Zhang, Z.F.; Huang, J.; Qian, L.D.; Teng, X.H.; Miao, Y.W. Genetic Diversity of mtDNA D-loop Region in Tengchong White Chicken. J. Yunnan Agric. Univ. Nat. Sci. 2021, 36, 75–81. (In Chinese) [Google Scholar]
- Liao, G.Z.; Wang, G.Y.; Cheng, Z.B.; Jia, J.J.; Ge, C.R. Proteomic Analysis of Tengchong Snowcock Muscles in Yunnan. Meat Res. 2013, 27, 1–5. (In Chinese) [Google Scholar]
- Dong, S.Y.; Guo, L.; Gu, D.H.; Wang, X.R.; Kan, H. Study on decocting technology and nutritional component analysis of Tengchong snow cock muscle. Meat Ind. 2018, 9, 15–18. (In Chinese) [Google Scholar]
- Liu, S.Y.; Xiao, X.X.; Ying, H.; Miao, Y.W.; Ye, S.H. Analysis and fitting of the growth curve of Tengchong snowcock. China Poult. 2016, 38, 45–47. (In Chinese) [Google Scholar]
- Richard, V.N.D.; Susan, J.L.; Max, F.R.; Michael, E.P.; Chris, M.A.; Carl, J.S. Transcriptome analysis of post-hatch breast muscle in legacy and modern broiler chickens reveals enrichment of several regulators of myogenic growth. PLoS ONE 2017, 10, e0122525. [Google Scholar]
- Brooke, M.H.; Kaiser, K.K. Muscle fiber types: How many and what kind? Arch. Neurol. 1970, 23, 369–379. [Google Scholar] [CrossRef] [PubMed]
- Sen, U.; Sirin, E.; Ensoy, U.; Aksoy, Y.; Ulutas, Z.; Kuran, M. The effect of maternal nutrition level during mid-gestation on postnatal muscle fibre composition and meat quality in lambs. Anim. Prod. Sci. 2016, 56, 834–843. [Google Scholar] [CrossRef]
- Nganvongpanit, K.; Kaewkumpai, P.; Kochagul, V.; Pringproa, K.; Punyapornwithaya, V.; Mekchay, S. Distribution of Melanin Pigmentation in 33 Organs of Thai Black-Bone Chickens (Gallus gallus domesticus). Animals 2020, 10, 777. [Google Scholar] [CrossRef]
- Kriangwanich, W.; Piboon, P.; Sakorn, W.; Buddhachat, K.; Kochagul, V.; Pringproa, K.; Mekchay, S.; Nganvongpanit, K. Consistency of dark skeletal muscles in Thai native black-bone chickens (Gallus gallus domesticus). PeerJ 2021, 9, e10728. [Google Scholar] [CrossRef]
- Harris, M.L.; Hall, R.; Erickson, C.A. Directing pathfinding along the dorsolateral path—The role of EDNRB2 and EphB2 in overcoming inhibition. Development 2008, 135, 4113–4122. [Google Scholar] [CrossRef] [Green Version]
- Xi, Y.; Xu, Q.; Huang, Q.; Ma, S.; Wang, Y.; Han, C.; Zhang, R.; Wang, J.; Liu, H.; Li, L. Genome-wide association analysis reveals that EDNRB2 causes a dose-dependent loss of pigmentation in ducks. BMC Genom. 2021, 22, 381. [Google Scholar] [CrossRef] [PubMed]
- Tejeda, O.J.; Calderon, A.J.; Arana, J.A.; Meloche, K.J.; Starkey, J.D. Broiler chicken myofiber morphometrics and myogenic stem cell population heterogeneity1,2. Poult. Sci. 2019, 98, 4123–4130. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, T.K.; Shukla, R.; Chatterjee, R.N.; Bhanja, S.K. Comparative analysis of silencing expression of myostatin (MSTN) and its two receptors (ACVR2A and ACVR2B) genes affecting growth traits in knock down chicken. Sci. Rep. 2019, 9, 7789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tapscott, S.J.; Davis, R.L.; Thayer, M.J.; Cheng, P.F.; Weintraub, H.; Lassar, A.B. MyoD1: A nuclear phosphoprotein requiring a Myc homology region to convert fibroblasts to myoblasts. Science 1988, 242, 405–411. [Google Scholar] [CrossRef] [PubMed]
- Huo, W.; Weng, K.; Gu, T.; Zhang, Y.; Zhang, Y.; Chen, G.; Xu, Q. Effect of muscle fiber characteristics on meat quality in fast- and slow-growing ducks. Poult. Sci. 2021, 100, 101264. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items (Unit) | Period I | Period II |
|---|---|---|
| Corn (%) | 63.25 | 60.19 |
| Soybean meal (%) | 30.26 | 25.88 |
| Wheat bran (%) | 0.00 | 10.00 |
| Fish meal (%) | 2.50 | 0.00 |
| CaCO3 (%) | 1.11 | 1.06 |
| CaHPO4 (%) | 1.50 | 1.50 |
| Met (%) | 0.08 | 0.07 |
| NaCl (%) | 0.30 | 0.30 |
| Premix (%) | 1.00 | 1.00 |
| ME (MJ/kg−1) | 13.02 | 12.8 |
| CP (%) | 20.00 | 18.60 |
| Phenotypes | Measurement |
|---|---|
| Body weight | The weight of bird before slaughter |
| Breast muscle weight | The weight of the breast without skin and adherent fat |
| Breast muscle rate | Percentage of breast muscle weight in body weight |
| Leg muscle weight | The weight of the two legs without skin and adherent fat |
| Leg muscle rate | Percentage of leg muscle weight in body weight |
| Gene | GenBank Accession | Primer Sequence (5′-3′) | Length (bp) | Product Size (bp) | Annealing Temperature (°C) |
|---|---|---|---|---|---|
| ACTB | NM_205518.2 | F: TGAGCGCAAGTACTCTGTCTG | 21 | 154 | 61.7 |
| R: TGTGGGTGTTGGTAACAGTCC | 21 | ||||
| MSTN | NM_001001461.2 | F: AAACGGTCCCGCAGAGATTT | 20 | 195 | 61 |
| R: CAGGTGAGTGTGCGGGTATT | 20 | ||||
| MYOD | NM_204214.3 | F: ACTACAGCGGGGAGTCAGAT | 20 | 190 | 61 |
| R: ATGCTTGAGAGGCAGTCGAG | 20 | ||||
| MYH7B | NM_204587.4 | F: TGACAACGCCTACAACGACA | 20 | 154 | 61.7 |
| R: TACTTTCTTGCCCGGTGTGT | 20 | ||||
| MYH1B | NM_204228.4 | F: GGTCAACAAGCTCCGAGCAA | 20 | 200 | 61.7 |
| R:TGCTGTATATGCAGAGAATCTATG | 25 | ||||
| MYH1A | NM_001013396.1 | F: CTTCCAGTCAGCACAAGACCT | 21 | 168 | 61.7 |
| R: AAGGCTTATTCTGGGCCTCG | 20 | ||||
| EDNRB2 | NM_204120.1 | F:AGAGGGACAACACAAAAGTAAAC | 25 | 158 | 61 |
| R: GCCAGGTTCCTAGTGAATTAAACTT | 25 | ||||
| TYR | NM_204160.1 | F: TGCTCAGATGAACAACGGCT | 20 | 179 | 61.7 |
| R: GCAGAAAAGCACGATGCCAA | 20 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, H.; Fu, J.; He, Y.; Li, Z.; Kang, J.; Hu, C.; Zi, X.; Liu, Y.; Zhao, J.; Dou, T.; et al. Hyperpigmentation Inhibits Early Skeletal Muscle Development in Tengchong Snow Chicken Breed. Genes 2022, 13, 2253. https://doi.org/10.3390/genes13122253
Shi H, Fu J, He Y, Li Z, Kang J, Hu C, Zi X, Liu Y, Zhao J, Dou T, et al. Hyperpigmentation Inhibits Early Skeletal Muscle Development in Tengchong Snow Chicken Breed. Genes. 2022; 13(12):2253. https://doi.org/10.3390/genes13122253
Chicago/Turabian StyleShi, Hongmei, Jing Fu, Yang He, Zijian Li, Jiajia Kang, Changjie Hu, Xiannian Zi, Yong Liu, Jinbo Zhao, Tengfei Dou, and et al. 2022. "Hyperpigmentation Inhibits Early Skeletal Muscle Development in Tengchong Snow Chicken Breed" Genes 13, no. 12: 2253. https://doi.org/10.3390/genes13122253