
Salt and Metal Tolerance Involves Formation of Guttation Droplets in Species of the Aspergillus versicolor Complex
 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Characterization of Isolates
2.2. Microbiome Analyses
2.3. Physiological and Morphological Characterization
2.4. Scanning Electron Microscopy
2.5. Analysis of Guttation Droplets
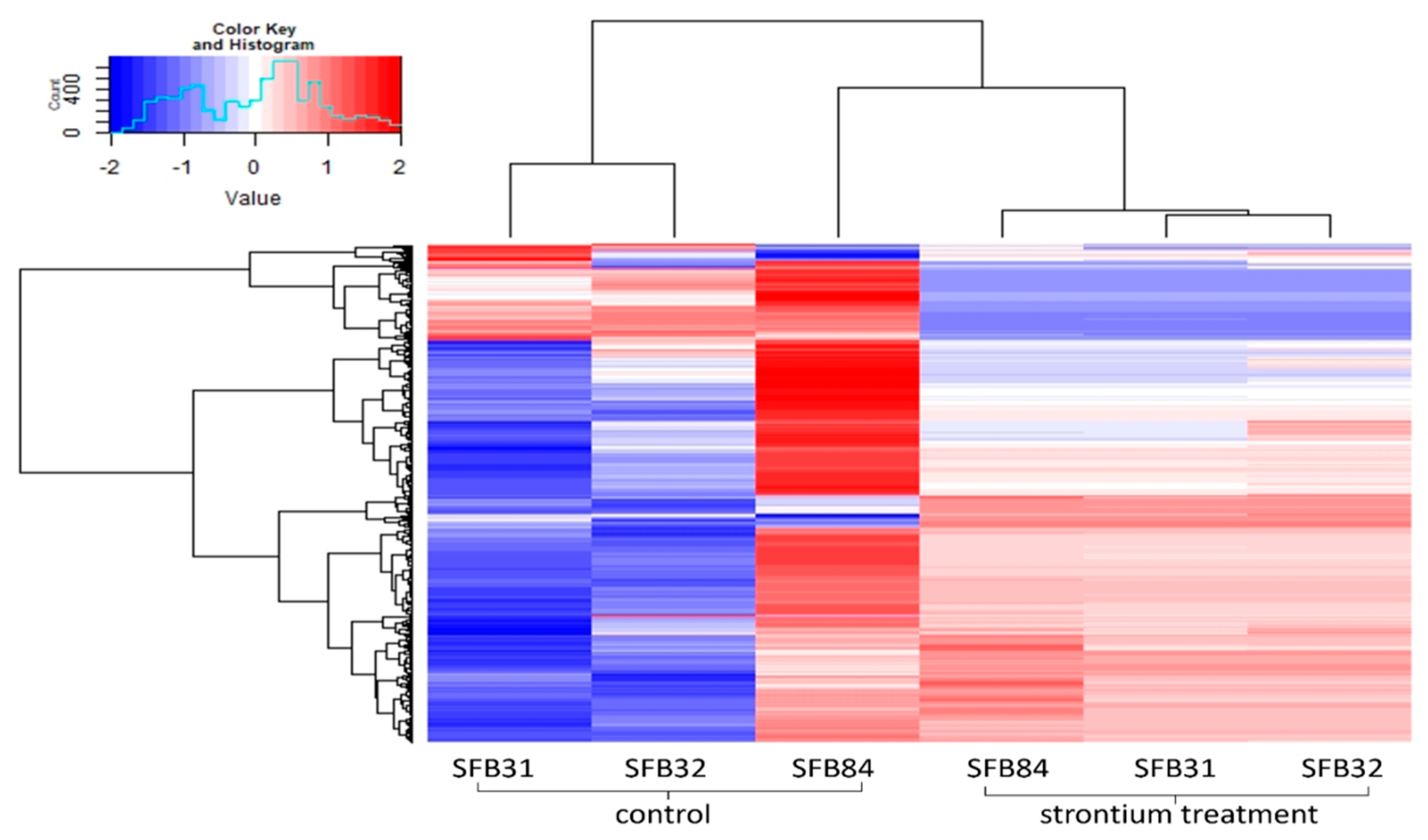
2.6. Proteomics Analysis
2.7. Metabolomics Analysis
3. Results
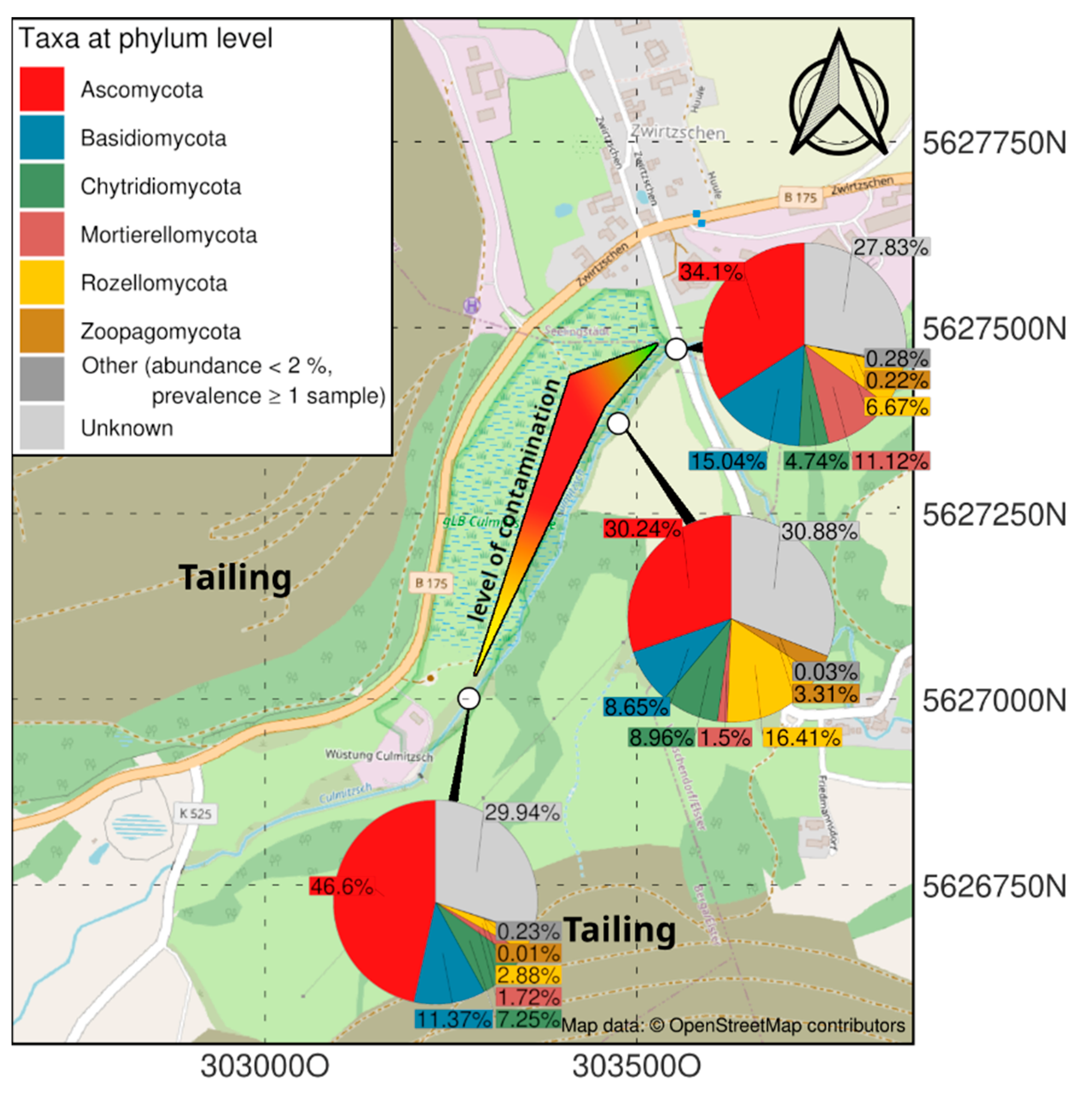
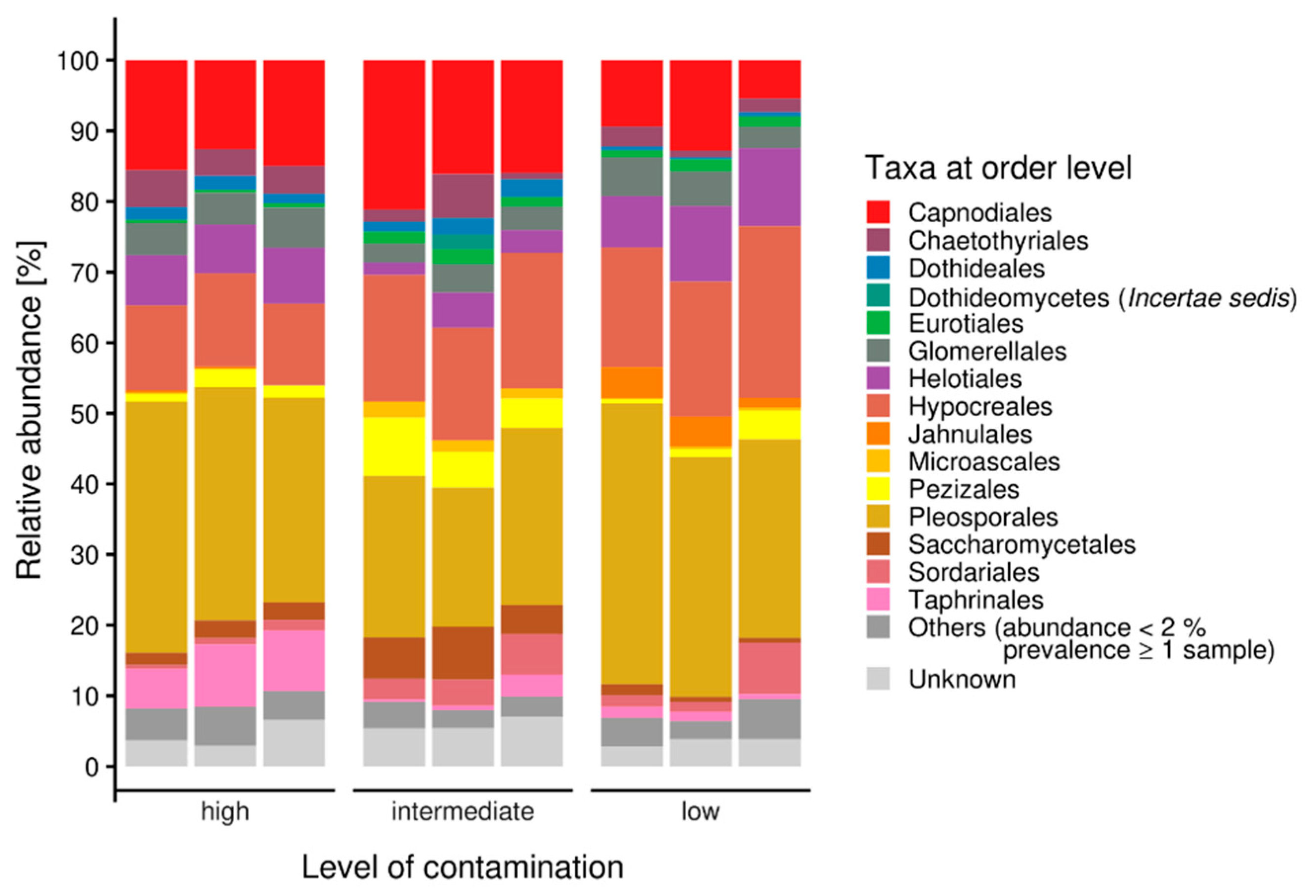
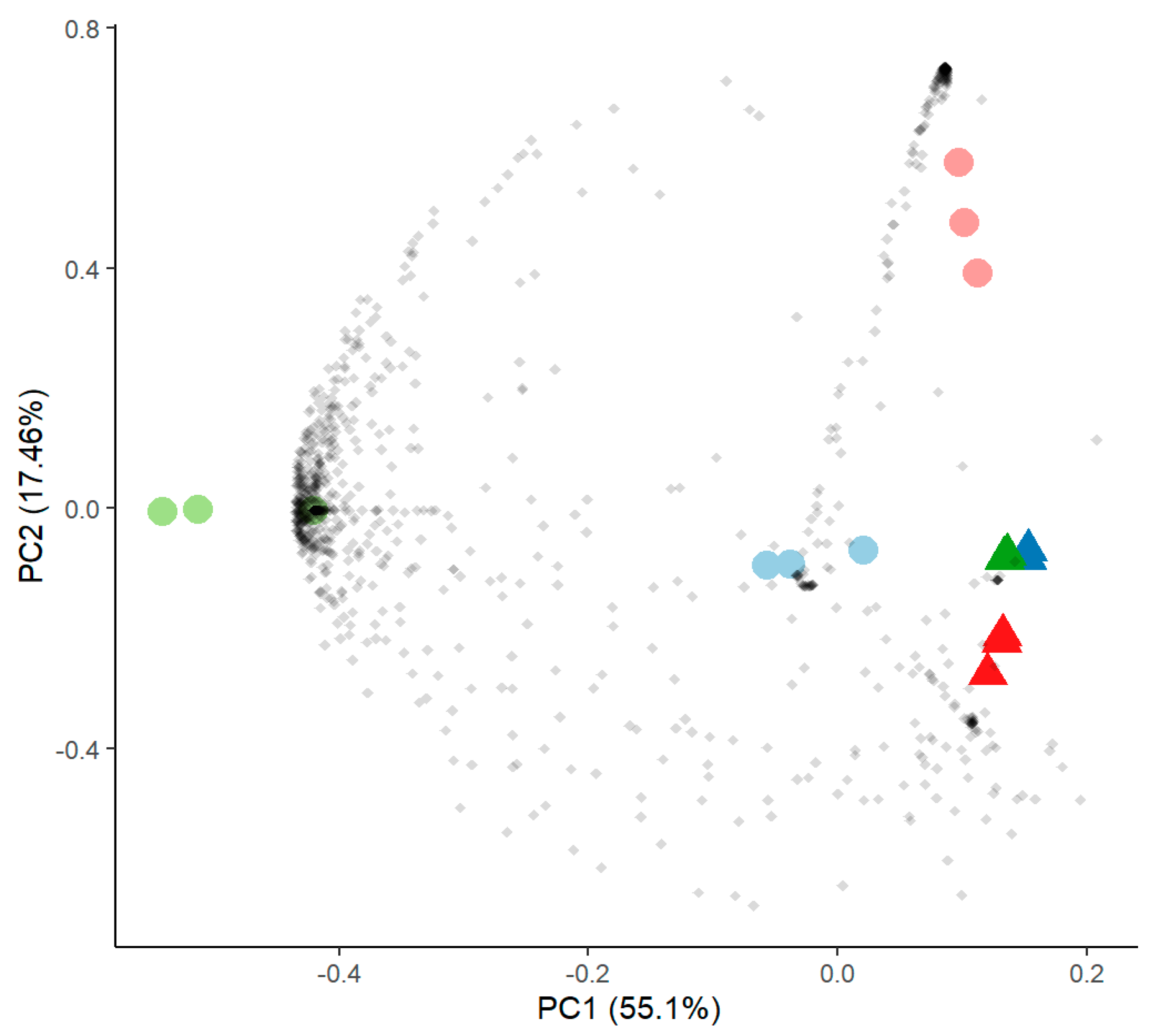
3.1. Microbiome Analysis
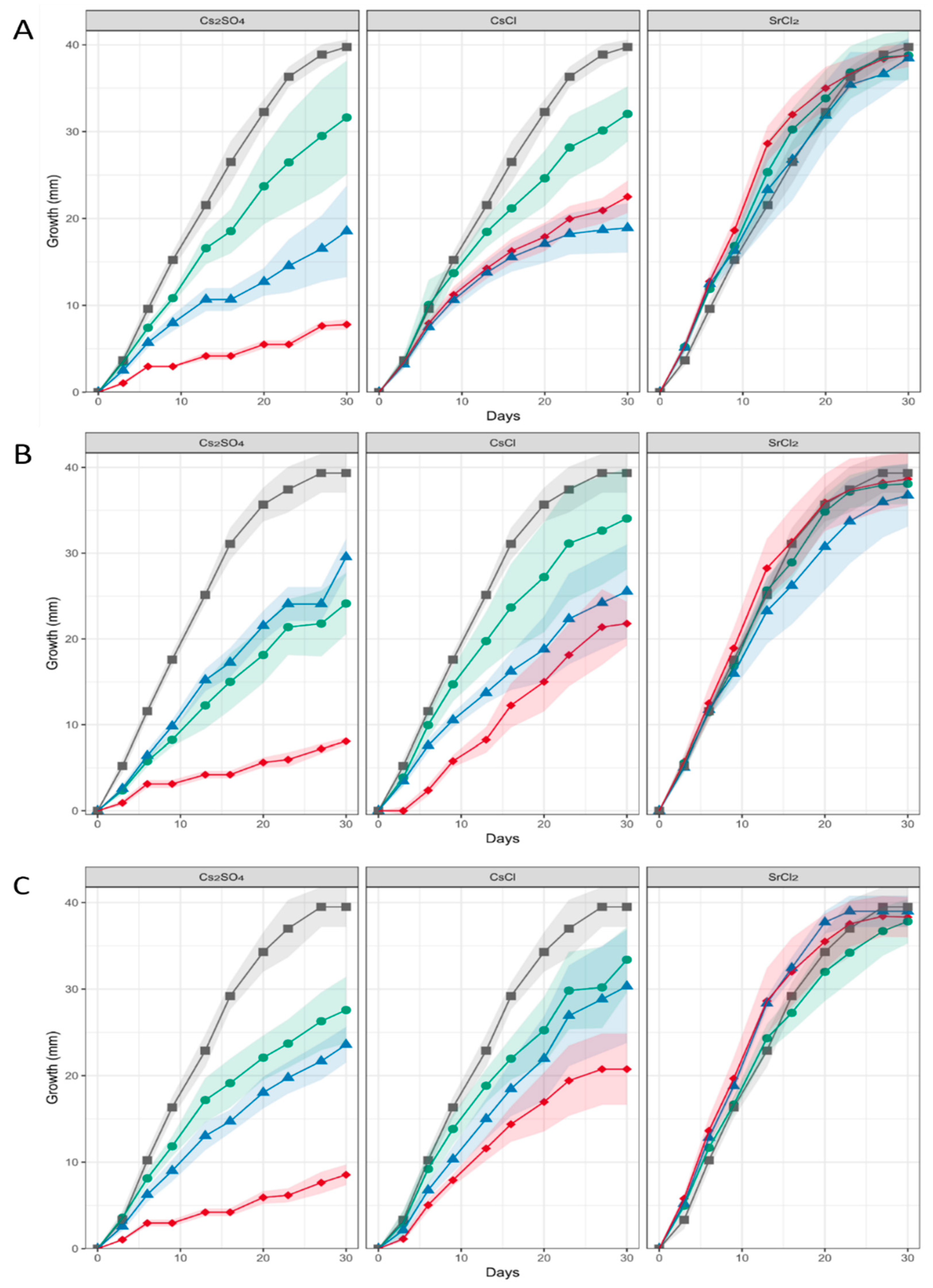
3.2. Three Aspergillus sp. Strains SFB31, SFB32 and SFB84 were Highly Metal and Salt Tolerant
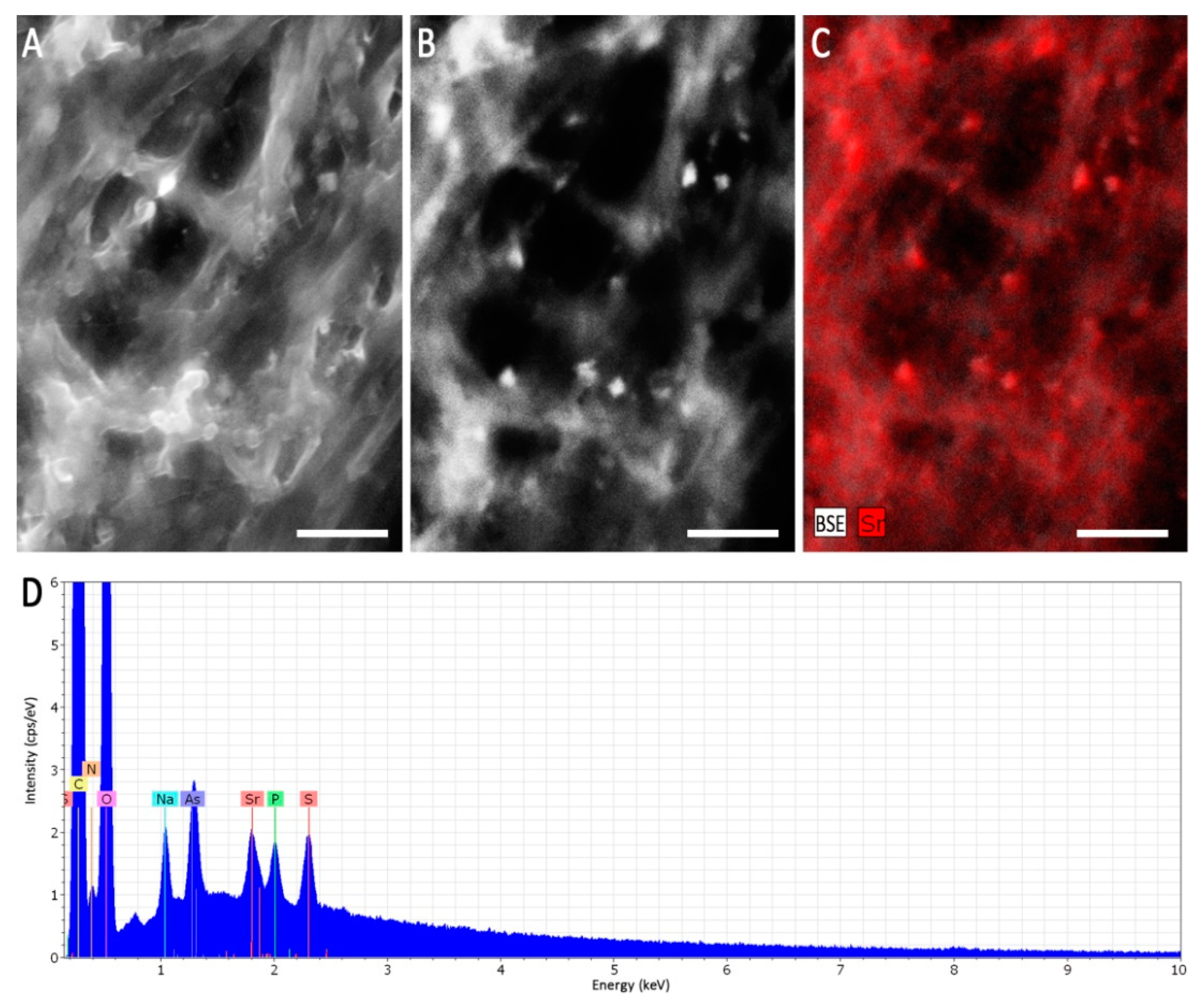
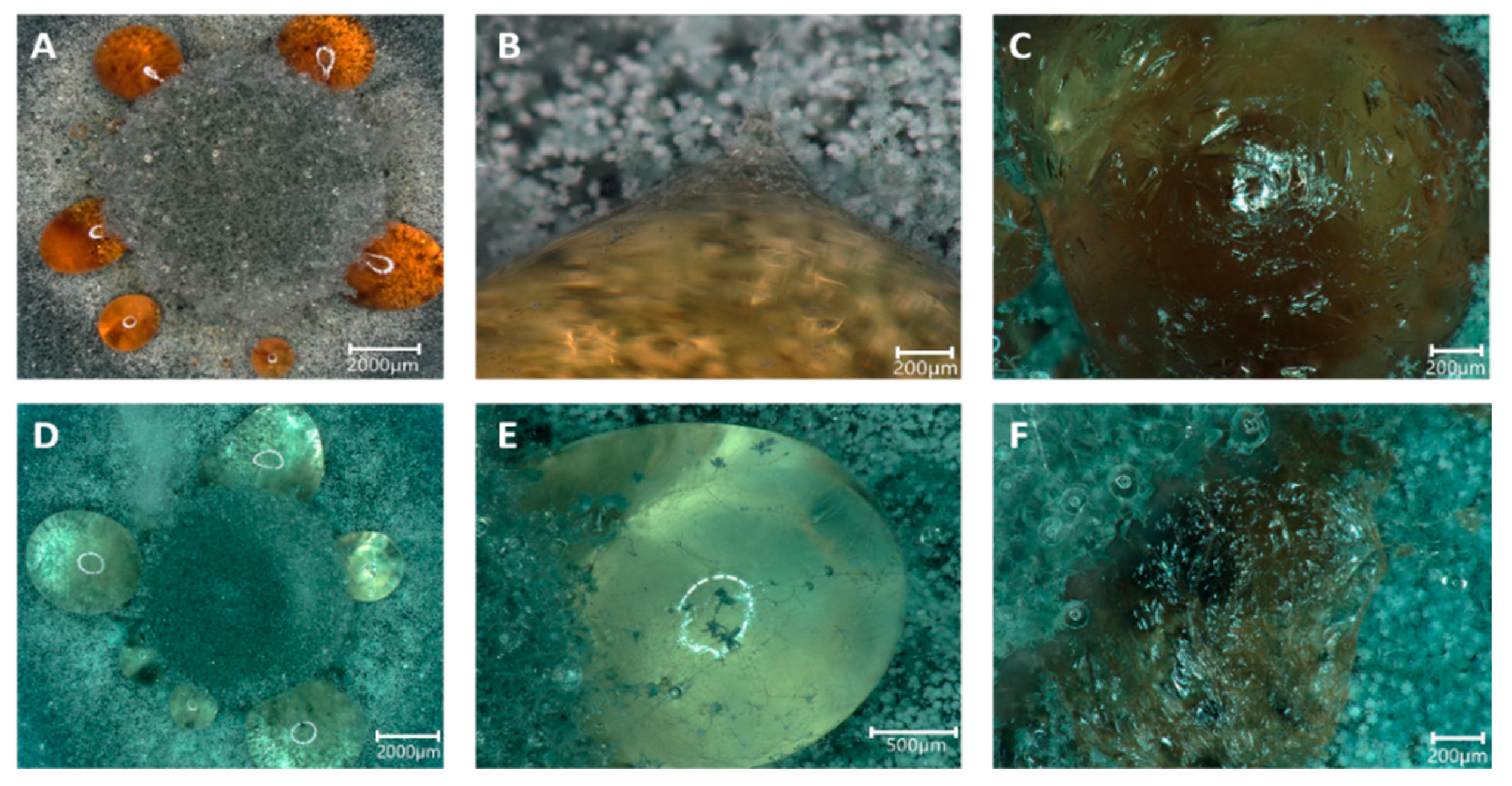
3.3. Formation of Guttation Droplets in Stress Response
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cowan, C.S.M.; Cryan, J.F. The microbiome-gut-brain axis in neurocognitive development and decline. Mod. Trends Psychiatry 2021, 32, 12–25. [Google Scholar] [CrossRef] [PubMed]
- Beyer, A.; Rzanny, M.; Weist, A.; Möller, S.; Burow, K.; Gutmann, F.; Neumann, S.; Lindner, J.; Müsse, S.; Brangsch, H.; et al. Aquifer community structure in dependence of lithostratigraphy in groundwater reservoirs. Environ. Sci. Pollut. Res. Int. 2015, 22, 19342–19351. [Google Scholar] [CrossRef] [PubMed]
- Mazière, C.; Agogué, H.; Cravo-Laureau, C.; Cagnon, C.; Lanneluc, I.; Sablé, S.; Fruitier-Arnaudin, I.; Dupuy, C.; Duran, R. New insights in bacterial and eukaryotic diversity of microbial mats inhabiting exploited and abandoned salterns at the Ré Island (France). Microbiol. Res. 2021, 252, 126854. [Google Scholar] [CrossRef] [PubMed]
- Harpke, M.; Pietschmann, S.; Costa, F.S.; Gansert, C.; Langenhorst, F.; Kothe, E. Biomineralization by extremely halophilic and metal-tolerant community members from a sulfate-dominated metal-rich environment. Microorganisms 2021, 10, 79. [Google Scholar] [CrossRef] [PubMed]
- Barnekow, U.; Metschies, T.; Paul, M. Remediation of Wismut’s Uranium Tailings Ponds at Seelingstädt, Germany. In Proceedings of the Uranium 2010—3rd International Conference on Uranium, Saskatoon, SK, Canada, 15–18 August 2010; Lam, E., Rowson, J., Özberk, E., Eds.; Volume II, pp. 507–516. [Google Scholar]
- Kothe, E.; Büchel, G. UMBRELLA: Using MicroBes for the REgulation of heavy metaL mobiLity at ecosystem and landscape scAle. Environ. Sci. Pollut. Res. Int. 2014, 21, 6761–6764. [Google Scholar] [CrossRef]
- Lersow, M.; Schmidt, P. The Wismut Remediation Project. In Proceedings of the Mine Closure 2006: First International Seminar on Mine Closure, Australian Centre for Geomechanics, Perth, Australia, 13–15 September 2006; Fourie, A.B., Tibbett, M., Eds.; pp. 181–190. [Google Scholar]
- Menéndez-Serra, M.; Triadó-Margarit, X.; Casamayor, E.O. Ecological and metabolic thresholds in the bacterial, protist, and fungal microbiome of Ephemeral Saline Lakes (Monegros Desert, Spain). Microb. Ecol. 2021, 82, 885–896. [Google Scholar] [CrossRef]
- Perez-Riverol, Y.; Bai, J.; Bandla, C.; García-Seisdedos, D.; Hewapathirana, S.; Kamatchinathan, S.; Kundu, D.J.; Prakash, A.; Frericks-Zipper, A.; Eisenacher, M.; et al. The PRIDE database resources in 2022: A hub for mass spectrometry-based proteomics evidences. Nucleic Acids Res. 2022, 50, D543–D552. [Google Scholar] [CrossRef]
- Chamekh, R.; Deniel, F.; Donot, C.; Jany, J.L.; Nodet, P.; Belabid, L. Isolation, identification and enzymatic activity of halotolerant and halophilic fungi from the Great Sebkha of Oran in Northwestern of Algeria. Mycobiology 2019, 47, 230–241. [Google Scholar] [CrossRef]
- Sepcic, K.; Zalar, P.; Gunde-Cimerman, N. Low water activity induces the production of bioactive metabolites in halophilic and halotolerant fungi. Mar. Drugs 2010, 9, 43–58. [Google Scholar] [CrossRef]
- Gunde-Cimerman, N.; Plemenitaš, A.; Oren, A. Strategies of adaptation of microorganisms of the three domains of life to high salt concentrations. FEMS Microbiol. Rev. 2018, 42, 353–375. [Google Scholar] [CrossRef] [Green Version]
- Hosseiniyan Khatibi, S.M.; Zununi Vahed, F.; Sharifi, S.; Ardalan, M.; Mohajel Shoja, M.; Zununi Vahed, S. Osmolytes resist against harsh osmolarity: Something old something new. Biochimie 2019, 158, 156–164. [Google Scholar] [CrossRef] [PubMed]
- Sleator, R.D.; Hill, C. Bacterial osmoadaptation: The role of osmolytes in bacterial stress and virulence. FEMS Microbiol. Rev. 2002, 26, 49–71. [Google Scholar] [CrossRef] [PubMed]
- Mevarech, M.; Frolow, F.; Gloss, L.M. Halophilic enzymes: Proteins with a grain of salt. Biophys. Chem. 2000, 86, 155–164. [Google Scholar] [CrossRef]
- Ventosa, A.; Márquez, M.C.; Garabito, M.J.; Arahal, D.R. Moderately halophilic gram-positive bacterial diversity in hypersaline environments. Extremophiles 1998, 2, 297–304. [Google Scholar] [CrossRef]
- Jiménez-Gómez, I.; Valdés-Muñoz, G.; Moreno-Perlin, T.; Mouriño-Pérez, R.R.; Sánchez-Carbente, M.D.R.; Folch-Mallol, J.L.; Pérez-Llano, Y.; Gunde-Cimerman, N.; Sánchez, N.D.C.; Batista-García, R.A. Haloadaptative responses of Aspergillus sydowii to extreme water deprivation: Morphology, compatible solutes, and oxidative stress at NaCl saturation. J. Fungi 2020, 6, 316. [Google Scholar] [CrossRef] [PubMed]
- Tafer, H.; Poyntner, C.; Lopandic, K.; Sterflinger, K.; Piñar, G. Back to the salt mines: Genome and transcriptome comparisons of the halophilic fungus Aspergillus salisburgensis and its halotolerant relative Aspergillus sclerotialis. Genes 2019, 10, 381. [Google Scholar] [CrossRef] [PubMed]
- Kis-Papo, T.; Weig, A.R.; Riley, R.; Peršoh, D.; Salamov, A.; Sun, H.; Lipzen, A.; Wasser, S.P.; Rambold, G.; Grigoriev, I.V.; et al. Genomic adaptations of the halophilic Dead Sea filamentous fungus Eurotium rubrum. Nat. Commun. 2014, 5, 3745. [Google Scholar] [CrossRef]
- Lotlikar, N.P.; Damare, S.R.; Meena, R.M.; Linsy, P.; Mascarenhas, B. Potential of marine-derived fungi to remove hexavalent chromium pollutant from culture broth. Indian J. Microbiol. 2018, 58, 182–192. [Google Scholar] [CrossRef]
- Rao, S.; Liang, J.; Song, W. Accumulation of 152+154Eu(III) by Aspergillus sydowii and Trichoderma harzianum. J. Environ. Radioact. 2018, 193, 75–81. [Google Scholar] [CrossRef]
- Zhang, J.; Miao, Y.; Rahimi, M.J.; Zhu, H.; Steindorff, A.; Schiessler, S.; Cai, F.; Pang, G.; Chenthamara, K.; Xu, Y.; et al. Guttation capsules containing hydrogen peroxide: An evolutionarily conserved NADPH oxidase gains a role in wars between related fungi. Environ. Microbiol. 2019, 21, 2644–2658. [Google Scholar] [CrossRef] [Green Version]
- Khani, M.H.; Pahlavanzadeh, H.; Alizadeh, K. Biosorption of strontium from aqueous solution by fungus Aspergillus terreus. Environ. Sci. Pollut. Res. Int. 2012, 19, 2408–2418. [Google Scholar] [CrossRef] [PubMed]
- Boczonádi, I.; Jakab, Á.; Baranyai, E.; Tóth, C.N.; Daróczi, L.; Csernoch, L.; Kis, G.; Antal, M.; Pusztahelyi, T.; Grawunder, A.; et al. Rare earth element sequestration by Aspergillus oryzae biomass. Environ. Technol. 2021, 42, 3725–3735. [Google Scholar] [CrossRef] [PubMed]
- Dong, Q.; Fei, L.; Wang, C.; Hu, S.; Wang, Z. Cadmium excretion via leaf hydathodes in tall fescue and its phytoremediation potential. Environ. Pollut. 2019, 252, 1406–1411. [Google Scholar] [CrossRef] [PubMed]
- Cerutti, A.; Jauneau, A.; Laufs, P.; Leonhardt, N.; Schattat, M.H.; Berthomé, R.; Routaboul, J.M.; Noël, L.D. Mangroves in the leaves: Anatomy, physiology, and immunity of epithemal hydathodes. Annu. Rev. Phytopathol. 2019, 57, 91–116. [Google Scholar] [CrossRef] [PubMed]
- Pereira, E.; Oliveira, I.; Baptista, P. Guttation droplets of the edible mushroom Suillus bovinus as a new source of natural antioxidants. Scientia Horticulturae 2021, 148, 89–92. [Google Scholar] [CrossRef]
- Andersson, M.A.; Salo, J.; Kedves, O.; Kredics, L.; Druzhinina, I.; Kurnitski, J.; Salonen, H. Bioreactivity, guttation and agents influencing surface tension of water emitted by actively growing indoor mould isolates. Microorganisms 2020, 8, 1940. [Google Scholar] [CrossRef]
- Muñoz, K.; Vega, M.; Rios, G.; Geisen, R.; Degen, G.H. Mycotoxin production by different ochratoxigenic Aspergillus and Penicillium species on coffee- and wheat-based media. Mycotoxin. Res. 2011, 27, 239–247. [Google Scholar] [CrossRef]
- Krain, A.; Siupka, P. Fungal guttation, a source of bioactive compounds, and its ecological role—A review. Biomolecules 2021, 11, 1270. [Google Scholar] [CrossRef]
- Karwehl, S.; Stadler, M. Exploitation of fungal biodiversity for discovery of novel antibiotics. Curr. Top. Microbiol. Immunol. 2016, 398, 303–338. [Google Scholar] [CrossRef]
- Hutwimmer, S.; Wang, H.; Strasser, H.; Burgstaller, W. Formation of exudate droplets by Metarhizium anisopliae and the presence of destruxins. Mycologia 2010, 102, 1–10. [Google Scholar] [CrossRef]
- Castagnoli, E.; Marik, T.; Mikkola, R.; Kredics, L.; Andersson, M.A.; Salonen, H.; Kurnitski, J. Indoor Trichoderma strains emitting peptaibols in guttation droplets. J. Appl. Microbiol. 2018, 125, 1408–1422. [Google Scholar] [CrossRef] [PubMed]
- Colotelo, N. Fungal exudates. Can. J. Microbiol. 1978, 24, 1173–1181. [Google Scholar] [CrossRef] [PubMed]
- Cenis, J.L. Rapid extraction of fungal DNA for PCR amplification. Nucleic Acids Res. 1992, 20, 2380. [Google Scholar] [CrossRef] [PubMed]
- Walters, W.; Hyde, E.R.; Berg-Lyons, D.; Ackermann, G.; Humphrey, G.; Parada, A. Improved bacterial 16S rRNA gene (V4 and V4-5) and fungal internal transcribed spacer marker gene primers for microbial community surveys. mSystems 2015, 1, e00009-15. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Jurjevic, Z.; Peterson, S.W.; Horn, B.W. Aspergillus section Versicolores: Nine new species and multilocus DNA sequence based phylogeny. IMA Fungus 2012, 3, 59–79. [Google Scholar] [CrossRef]
- Wessel, D.; Flügge, U.I. A method for the quantitative recovery of protein in dilute solution in the presence of detergents and lipids. Anal. Biochem. 1984, 138, 141–143. [Google Scholar] [CrossRef]
- Wang, Y.; Tan, Y.; Wang, Y.; Ge, Y.; Liu, Y.; Liu, H.; Shao, L.; Liu, Y.; Ren, X.; Liu, Z. Role of AcndtA in cleistothecium formation, osmotic stress response, pigmentation and carbon metabolism of Aspergillus cristatus. Fungal Biol. 2021, 125, 749–763. [Google Scholar] [CrossRef]
- Chen, G.; Yang, S.; Wang, C.; Shi, K.; Zhao, X.; Wu, Z. Investigation of the mycelial morphology of Monascus and the expression of pigment biosynthetic genes in high-salt-stress fermentation. Appl. Microbiol. Biotechnol. 2020, 104, 2469–2479. [Google Scholar] [CrossRef]
- Chang, P.K.; Cary, J.W.; Lebar, M.D. Biosynthesis of conidial and sclerotial pigments in Aspergillus species. Appl. Microbiol. Biotechnol. 2020, 104, 2277–2286. [Google Scholar] [CrossRef] [PubMed]
- Salo, M.J.; Marik, T.; Mikkola, R.; Andersson, M.A.; Kredics, L.; Salonen, H.; Kurnitski, J. Penicillium expansum strain isolated from indoor building material was able to grow on gypsum board and emitted guttation droplets containing chaetoglobosins and communesins A, B and D. J. Appl. Microbiol. 2019, 127, 1135–1147. [Google Scholar] [CrossRef] [PubMed]
- Koulman, A.; Lane, G.A.; Christensen, M.J.; Fraser, K.; Tapper, B.A. Peramine and other fungal alkaloids are exuded in the guttation fluid of endophyte-infected grasses. Phytochemistry 2007, 68, 355–360. [Google Scholar] [CrossRef] [PubMed]
- Chang, P.K.; Scharfenstein, L.L.; Mack, B.; Wie, Q.; Gilbert, M.; Lebar, M.; Cary, J.W. Identification of a copper-transporting ATPase involved in biosynthesis of A. flavus conidial pigment. Appl. Microbiol. Biotechnol. 2019, 103, 4889–4897. [Google Scholar] [CrossRef]
- Xie, N.; Ruprich-Robert, G.; Silar, P.; Herbert, E.; Ferrari, R.; Chapeland-Leclerc, F. Characterization of three multicopper oxidases in the filamentous fungus Podospora anserina: A new role of an ABR1-like protein in fungal development? Fungal Genet. Biol. 2018, 116, 1–13. [Google Scholar] [CrossRef]
- Sanyal, S.K.; Kanwar, P.; Yadav, A.K.; Sharma, C.; Kumar, A.; Pandey, G.K. Arabidopsis CBL interacting protein kinase 3 interacts with ABR1, an APETALA2 domain transcription factor, to regulate ABA responses. Plant Sci. 2017, 254, 48–59. [Google Scholar] [CrossRef]
- Aikawa, Y.; Martin, T.F. ARF6 regulates a plasma membrane pool of phosphatidylinositol(4,5)bisphosphate required for regulated exocytosis. J. Cell Biol. 2003, 162, 647–659. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harpke, M.; Pietschmann, S.; Ueberschaar, N.; Krüger, T.; Kniemeyer, O.; Brakhage, A.A.; Nietzsche, S.; Kothe, E. Salt and Metal Tolerance Involves Formation of Guttation Droplets in Species of the Aspergillus versicolor Complex. Genes 2022, 13, 1631. https://doi.org/10.3390/genes13091631
Harpke M, Pietschmann S, Ueberschaar N, Krüger T, Kniemeyer O, Brakhage AA, Nietzsche S, Kothe E. Salt and Metal Tolerance Involves Formation of Guttation Droplets in Species of the Aspergillus versicolor Complex. Genes. 2022; 13(9):1631. https://doi.org/10.3390/genes13091631
Chicago/Turabian StyleHarpke, Marie, Sebastian Pietschmann, Nico Ueberschaar, Thomas Krüger, Olaf Kniemeyer, Axel A. Brakhage, Sandor Nietzsche, and Erika Kothe. 2022. "Salt and Metal Tolerance Involves Formation of Guttation Droplets in Species of the Aspergillus versicolor Complex" Genes 13, no. 9: 1631. https://doi.org/10.3390/genes13091631