
Excavation of Genes Responsive to Brassinosteroids by Transcriptome Sequencing in Adiantum flabellulatum Gametophytes

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Materials and Drugs
2.2. Experimental Methods
2.2.1. Culture of Adiantum flabellulatum Gametophytes
2.2.2. RNA Extraction, Library Construction, Sequencing, and Data Analysis
2.2.3. Quality Evaluation of Transcriptome Sequencing
2.2.4. Screening of Differentially Expressed Genes, GO, and KEGG Analysis
2.2.5. Quantitative Real-Time Polymerase Chain Reaction
3. Results
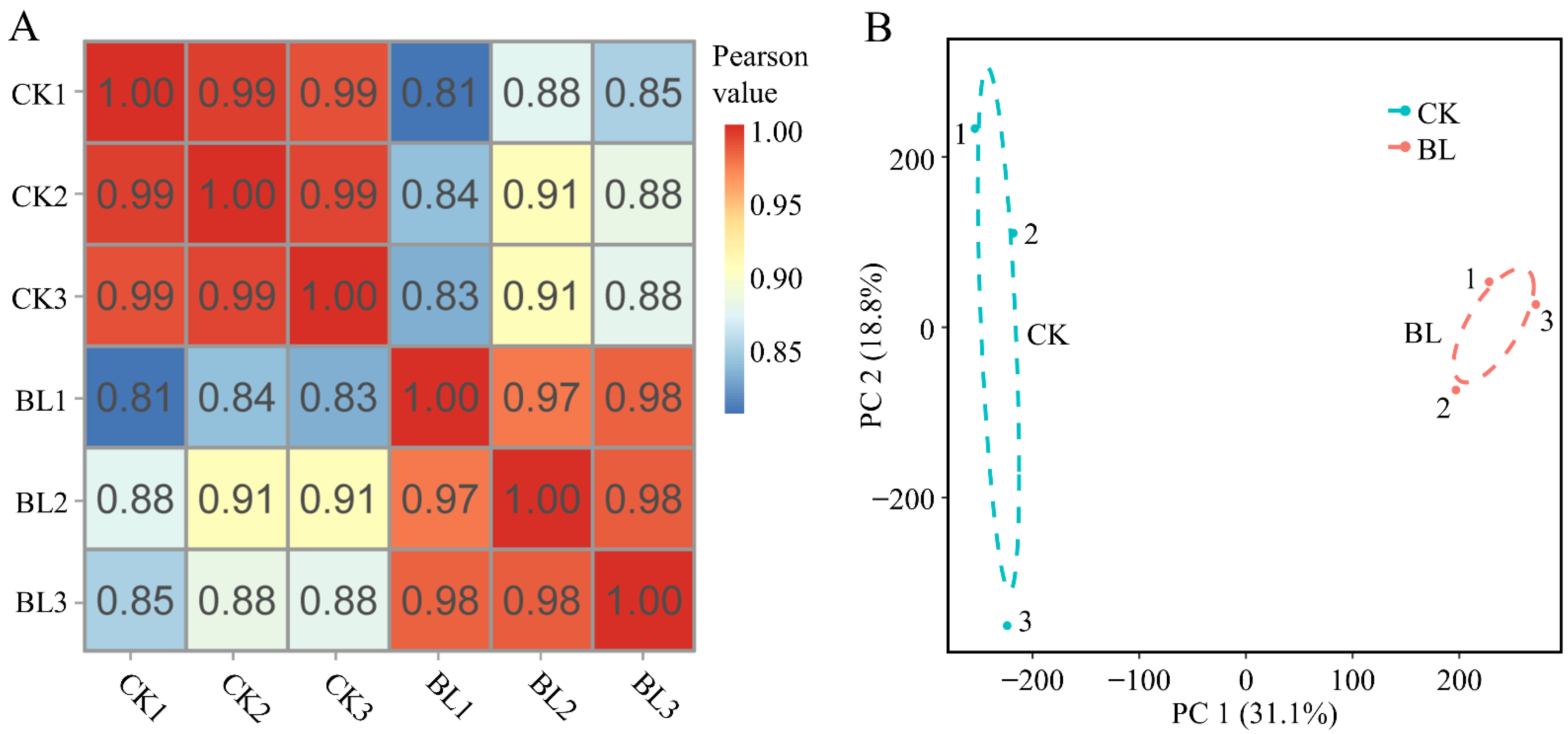
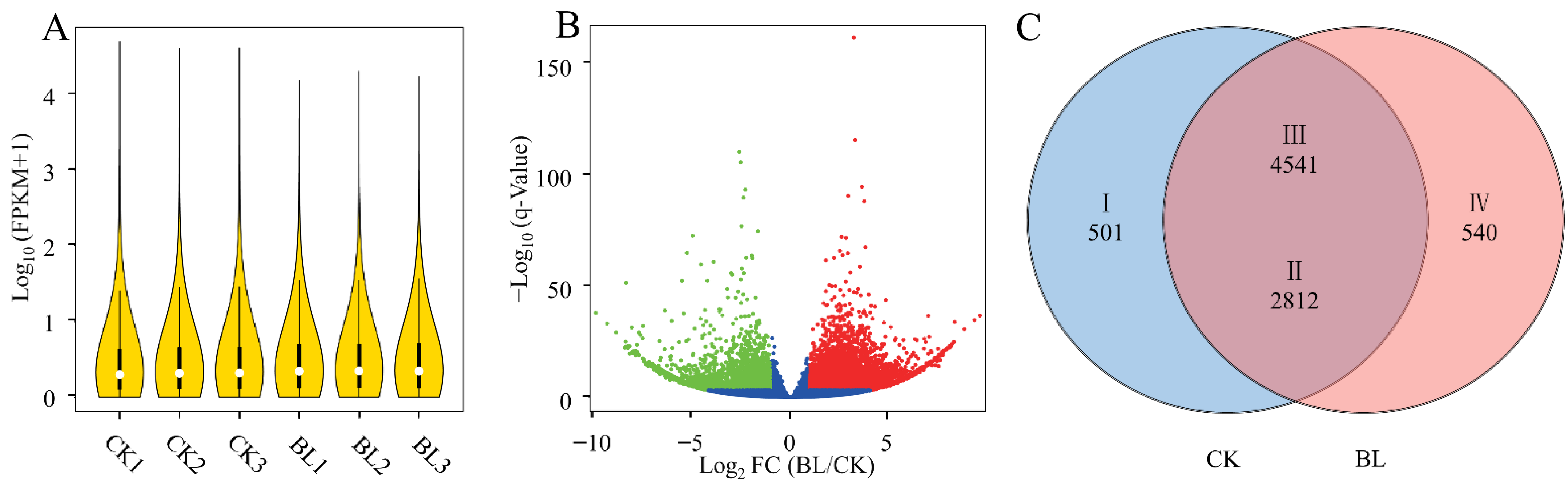
3.1. Quality Evaluation of Transcriptome Sequencing
3.2. Screening of DEGs
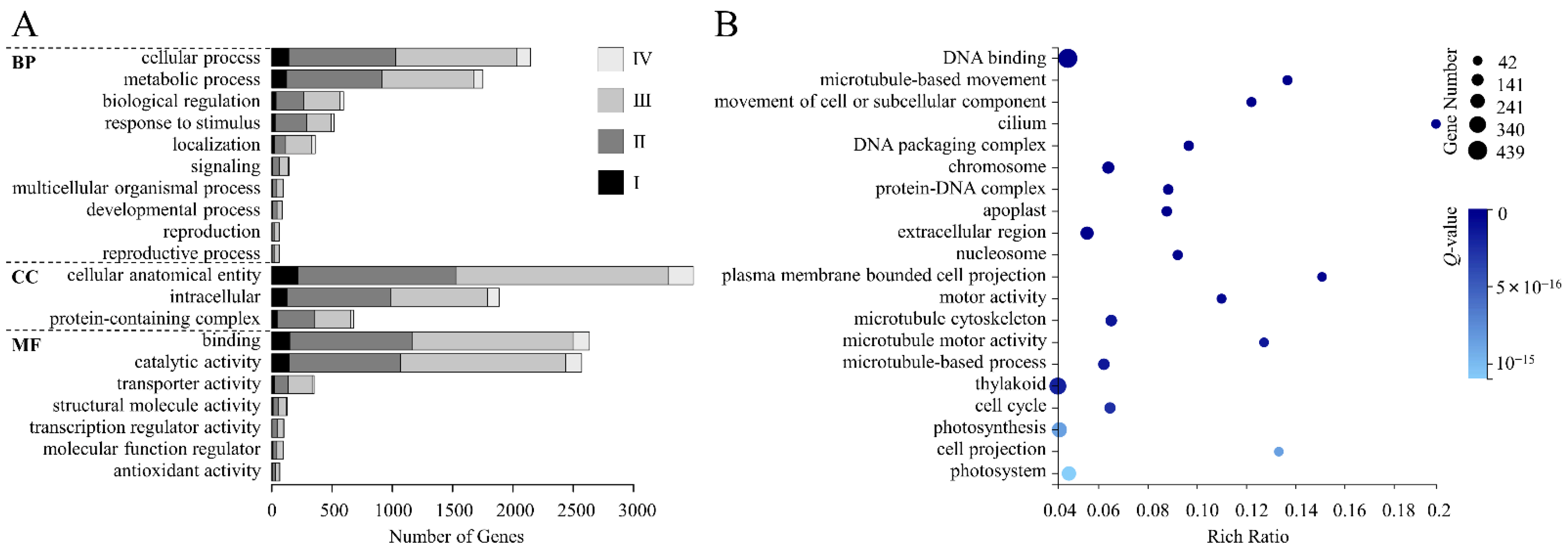
3.3. GO Analysis of DEGs
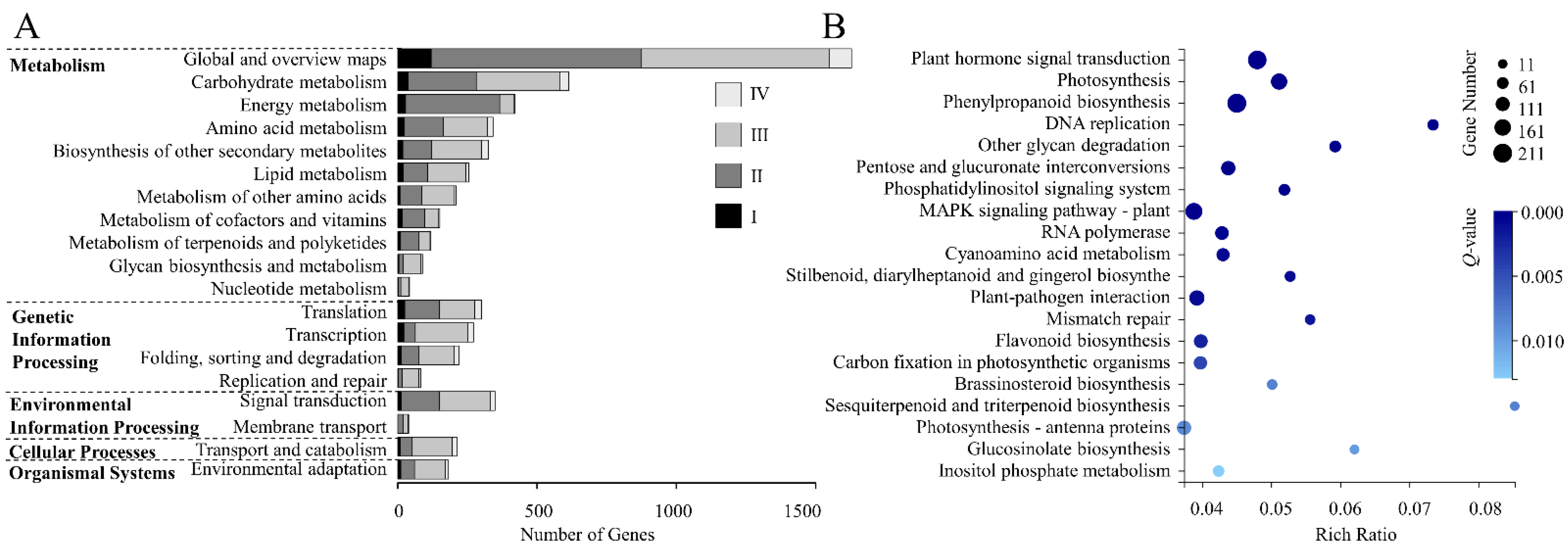
3.4. KEGG Analysis of DEGs
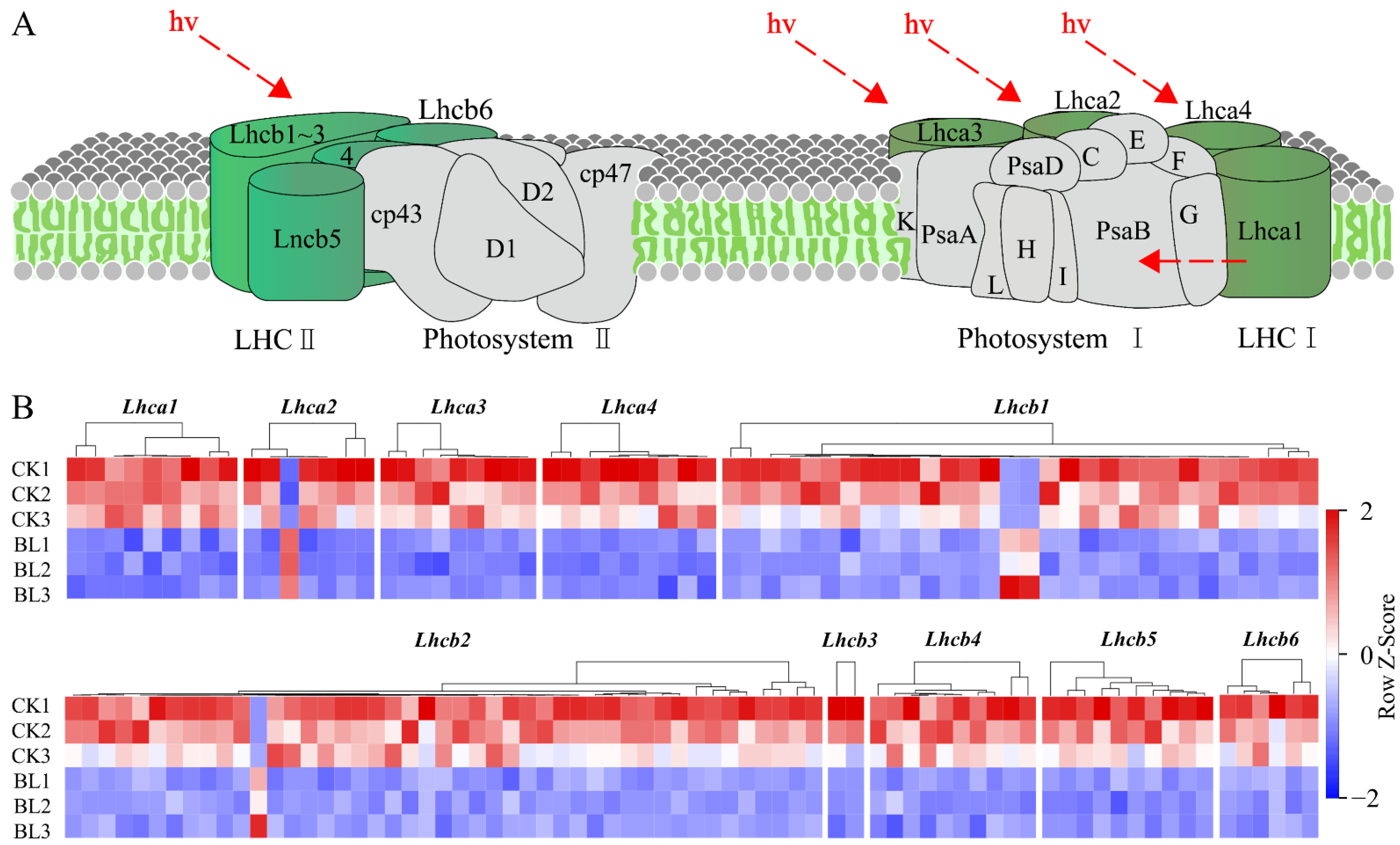
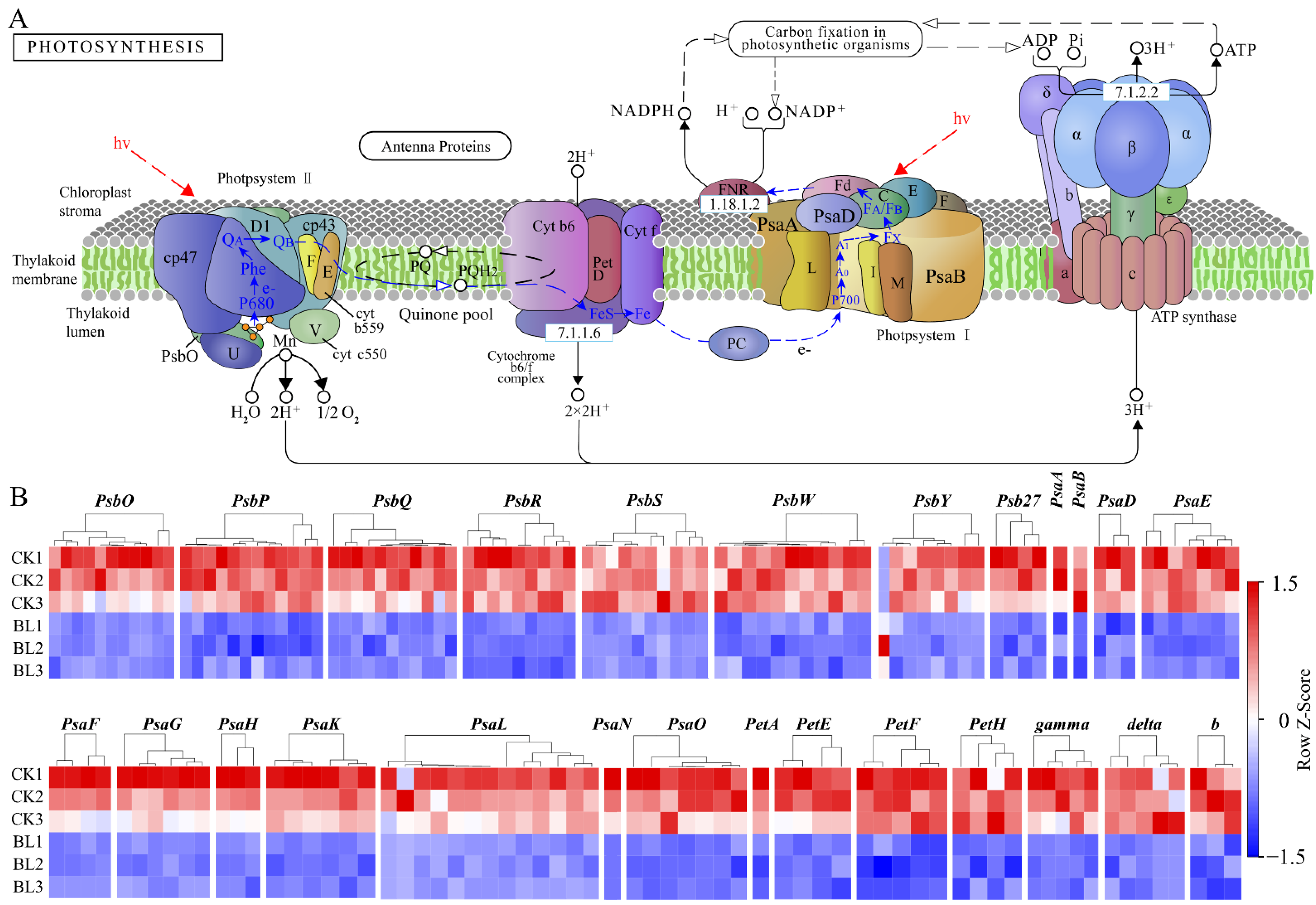
3.5. The Expressions of Photosynthesis-Related Genes Were Inhibited
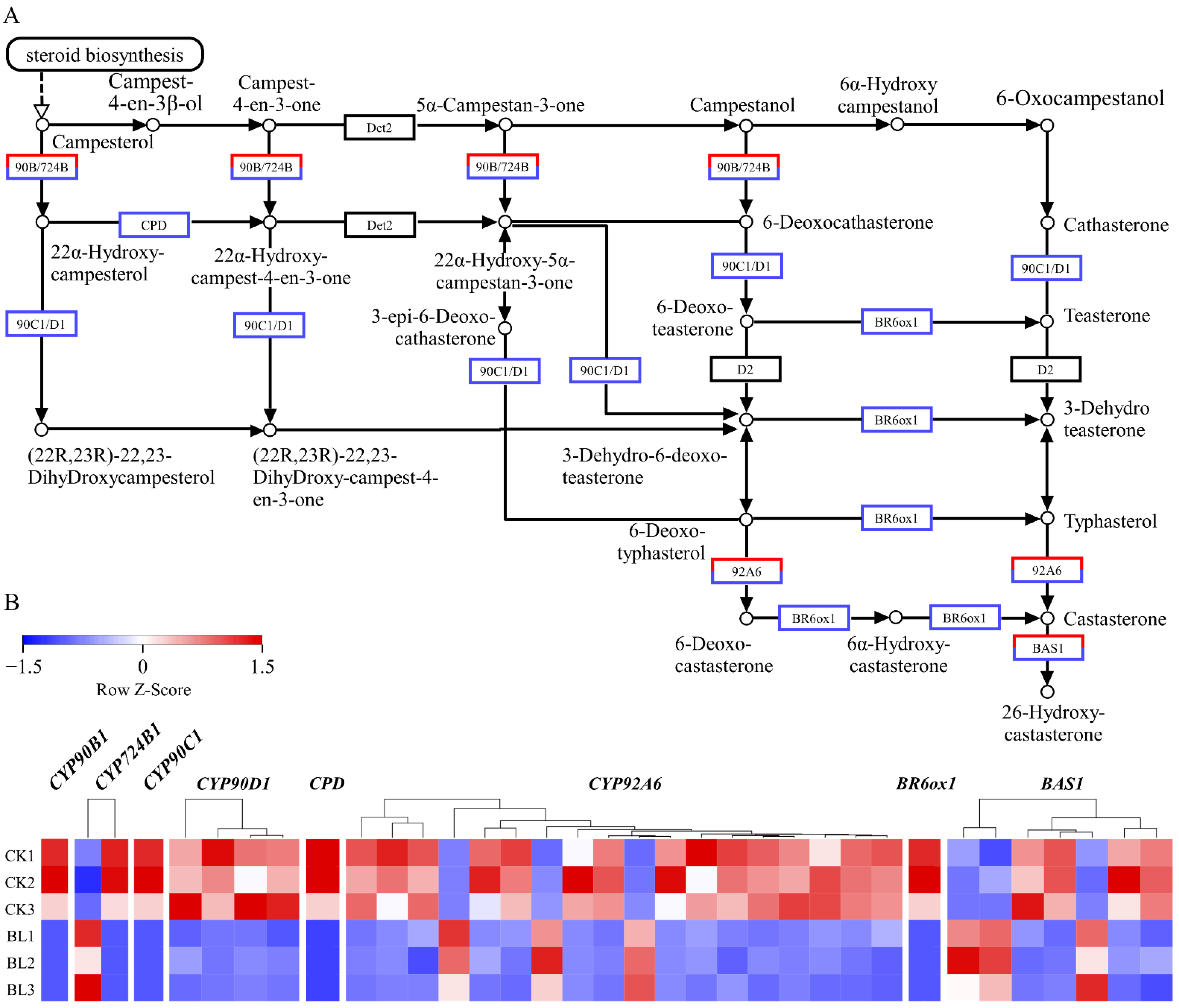
3.6. The Expressions of BR Synthase Genes Were Negatively Regulated by BL
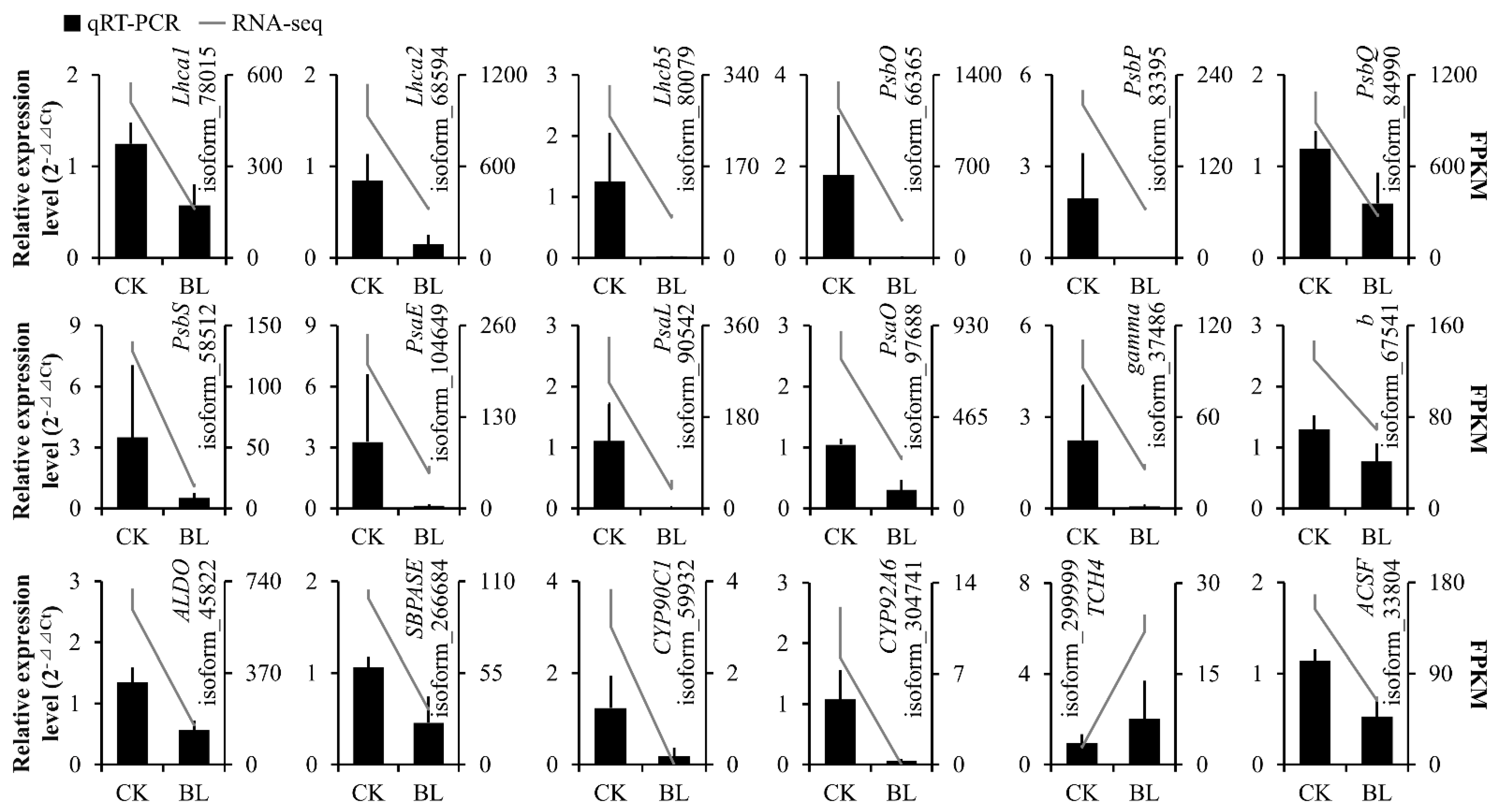
3.7. Quantitative Real-Time Polymerase Chain Reaction
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bajguz, A. Metabolism of brassinosteroids in plants. Plant Physiol. Biochem. 2007, 45, 95–107. [Google Scholar] [CrossRef] [PubMed]
- De Bruyne, L.; Höfte, M.; De Vleesschauwer, D. Connecting growth and defense: The emerging roles of brassinosteroids and gibberellins in plant innate immunity. Mol. Plant 2014, 7, 943–959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, M.A.; Fahad, S.; Sharif, R.; Jan, M.F.; Mujtaba, M.; Ali, Q.; Ahmad, A.; Ahmad, H.; Amin, N.; Ajayo, B.S.; et al. Multifunctional role of brassinosteroid and its analogues in plants. Plant Growth Regul. 2020, 92, 141–156. [Google Scholar] [CrossRef]
- Friedrichsen, D.; Chory, J. Steroid signaling in plants: From the cell surface to the nucleus. Bioessays 2001, 23, 1028–1036. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.J.; Zhang, C.; Lu, Y.N.; Jin, J.Q.; Wang, X.L. The mechanisms of brassinosteroids’ action: From signal transduction to plant development. Mol. Plant 2011, 4, 588–600. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Wen, J.; Lease, K.A.; Doke, J.T.; Tax, F.E.; Walker, J.C. BAK1, an Arabidopsis LRR receptor-like protein kinase, interacts with BRI1 and modulates brassinosteroid signaling. Cell 2002, 110, 213–222. [Google Scholar] [CrossRef] [Green Version]
- Nam, K.H.; Li, J. BRI1/BAK1, a receptor kinase pair mediating brassinosteroid signaling. Cell 2002, 110, 203–212. [Google Scholar] [CrossRef] [Green Version]
- Manghwar, H.; Hussain, A.; Ali, Q.; Liu, F. Brassinosteroids (BRs) Role in Plant Development and Coping with Different Stresses. Int. J. Mol. Sci. 2022, 23, 1012. [Google Scholar] [CrossRef]
- Gómez-Garay, A.; Gabriel y Galán, J.M.; Cabezuelo, A.; Pintos, B.; Prada, C.; Martín, L. Ecological Significance of Brassinosteroids in Three Temperate Ferns. In Current Advances in Fern Research; Fernández, H., Ed.; Springer International Publishing: Cham, Switzerland, 2018; pp. 453–466. [Google Scholar]
- Wang, J.M.; Yu, X.D.; Cai, Z.P.; Xu, S.T.; Luo, J.J.; Yang, Z.W. Effects of Brassinosteroids on gametophyte growth of Adiantum flabellulatum L. Chin. J. Trop. Crop. 2018, 39, 1739–1744. [Google Scholar] [CrossRef]
- Yokota, T.; Arima, M.; Takahashi, N. Castasterone, a new phytosterol with plant-hormone potency, from chestnut insect gall. Tetrahedron Lett. 1982, 23, 1275–1278. [Google Scholar] [CrossRef]
- Yokota, T.; Ohnishi, T.; Shibata, K.; Asahina, M.; Nomura, T.; Fujita, T.; Ishizaki, K.; Kohchi, T. Occurrence of brassinosteroids in non-flowering land plants, liverwort, moss, lycophyte and fern. Phytochemistry 2017, 136, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Fujioka, S.; Noguchi, T.; Yokota, T.; Takatsuto, S.; Yoshida, S. Brassinosteroids in Arabidopsis thaliana. Phytochemistry 1998, 48, 595–599. [Google Scholar] [CrossRef]
- Montoya, T.; Nomura, T.; Yokota, T.; Farrar, K.; Harrison, K.; Jones, J.D.; Kaneta, T.; Kamiya, Y.; Szekeres, M.; Bishop, G.J. Patterns of Dwarf expression and brassinosteroid accumulation in tomato reveal the importance of brassinosteroid synthesis during fruit development. Plant J. 2005, 42, 262–269. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.; Fujioka, S.; Kozuka, T.; Tax, F.; Takatsuto, S.; Yoshida, S.; Tsukaya, H. CYP90C1 and CYP90D1 are involved in different steps in the brassinosteroid biosynthesis pathway in Arabidopsis thaliana. Plant J. Cell Mol. Biol. 2005, 41, 710–721. [Google Scholar] [CrossRef] [PubMed]
- Kwon, M.; Choe, S. Brassinosteroid biosynthesis and dwarf mutants. J. Plant Biol. 2005, 48, 1. [Google Scholar] [CrossRef]
- Zhao, B.; Li, J. Regulation of brassinosteroid biosynthesis and inactivation. J. Integr. Plant Biol. 2012, 54, 746–759. [Google Scholar] [CrossRef]
- Je, B.I.; Han, C.D. Brassinosteroid homeostasis via coordinate regulation of signaling and synthetic pathways. Plant Signal. Behav. 2010, 5, 1440–1441. [Google Scholar] [CrossRef] [Green Version]
- Bancoş, S.; Nomura, T.; Sato, T.; Molnár, G.; Bishop, G.J.; Koncz, C.; Yokota, T.; Nagy, F.; Szekeres, M. Regulation of transcript levels of the Arabidopsis cytochrome p450 genes involved in brassinosteroid biosynthesis. Plant Physiol. 2002, 130, 504–513. [Google Scholar] [CrossRef] [Green Version]
- Seyed Rahmani, R.; Shi, T.; Zhang, D.; Gou, X.; Yi, J.; Miclotte, G.; Marchal, K.; Li, J. Genome-wide expression and network analyses of mutants in key brassinosteroid signaling genes. BMC Genom. 2021, 22, 465. [Google Scholar] [CrossRef]
- Huang, G.; Han, M.; Yao, W.; Wang, Y. Transcriptome analysis reveals the regulation of brassinosteroids on petal growth in Gerbera hybrida. PeerJ 2017, 5, e3382. [Google Scholar] [CrossRef]
- Wu, Q.; Wang, J.; Mao, S.; Xu, H.; Wu, Q.; Liang, M.; Yuan, Y.; Liu, M.; Huang, K. Comparative transcriptome analyses of genes involved in sulforaphane metabolism at different treatment in Chinese kale using full-length transcriptome sequencing. BMC Genom. 2019, 20, 377. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Fujioka, S.; Takatsuto, S.; Yokota, T.; Murofushi, N.; Sakurai, A. Biosynthesis of Brassinosteroids in Seedlings of Catharanthus roseus, Nicotiana tabacum, and Oryza sativa. Biosci. Biotechnol. Biochem. 1995, 59, 168–172. [Google Scholar] [CrossRef]
- Kim, S.K.; Chang, S.C.; Lee, E.J.; Chung, W.S.; Kim, Y.S.; Hwang, S.; Lee, J.S. Involvement of brassinosteroids in the gravitropic response of primary root of maize. Plant Physiol. 2000, 123, 997–1004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, Z.; Ueguchi-Tanaka, M.; Umemura, K.; Uozu, S.; Fujioka, S.; Takatsuto, S.; Yoshida, S.; Ashikari, M.; Kitano, H.; Matsuoka, M. A rice brassinosteroid-deficient mutant, ebisu dwarf (d2), is caused by a loss of function of a new member of cytochrome P450. Plant Cell 2003, 15, 2900–2910. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.-S.; Kim, T.-W.; Kim, S.-K. Brassinosteroids are inherently biosynthesized in the primary roots of maize, Zea mays L. Phytochemistry 2005, 66, 1000–1006. [Google Scholar] [CrossRef]
- Xiao, Y.; Zhang, G.; Liu, D.; Niu, M.; Tong, H.; Chu, C. GSK2 stabilizes OFP3 to suppress brassinosteroid responses in rice. Plant J. 2020, 102, 1187–1201. [Google Scholar] [CrossRef]
- Tian, X.; He, M.; Mei, E.; Zhang, B.; Tang, J.; Xu, M.; Liu, J.; Li, X.; Wang, Z.; Tang, W.; et al. WRKY53 integrates classic brassinosteroid signaling and the mitogen-activated protein kinase pathway to regulate rice architecture and seed size. Plant Cell 2021, 33, 2753–2775. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, Y.; Shi, C.; Huang, Z.; Zhang, Y.; Li, S.; Li, Y.; Ye, J.; Yu, C.; Li, Z.; et al. SOAPnuke: A MapReduce acceleration-supported software for integrated quality control and preprocessing of high-throughput sequencing data. Gigascience 2018, 7, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Cai, Z.; Xie, Z.; Huang, L.; Wang, Z.; Pan, M.; Yu, X.; Xu, S.; Luo, J. Full-length transcriptome analysis of Adiantum flabellulatum gametophyte. PeerJ 2022, 10, e13079. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, Z.; Huang, Z.; Wang, Z.; Tao, Y.; Luo, J. Identification of the related genes on the asymmetric root growth of Oryza sativa induced by ethylene through transcriptome sequencing, GO and KEGG analysis. Acta Physiol. Plant. 2021, 43, 1–11. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Iliev, E.A.; Xu, W.; Polisensky, D.H.; Oh, M.H.; Torisky, R.S.; Clouse, S.D.; Braam, J. Transcriptional and posttranscriptional regulation of Arabidopsis TCH4 expression by diverse stimuli. Roles of cis regions and brassinosteroids. Plant Physiol. 2002, 130, 770–783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nazir, F.; Fariduddin, Q.; Hussain, A.; Khan, T.A. Brassinosteroid and hydrogen peroxide improve photosynthetic machinery, stomatal movement, root morphology and cell viability and reduce Cu- triggered oxidative burst in tomato. Ecotoxicol. Environ. Saf. 2021, 207, 111081. [Google Scholar] [CrossRef]
- Jan, S.; Alyemeni, M.N.; Wijaya, L.; Alam, P.; Siddique, K.H.; Ahmad, P. Interactive effect of 24-epibrassinolide and silicon alleviates cadmium stress via the modulation of antioxidant defense and glyoxalase systems and macronutrient content in Pisum sativum L. seedlings. BMC Plant Biol. 2018, 18, 146. [Google Scholar] [CrossRef]
- Yu, X.; Li, L.; Zola, J.; Aluru, M.; Ye, H.; Foudree, A.; Guo, H.; Anderson, S.; Aluru, S.; Liu, P.; et al. A brassinosteroid transcriptional network revealed by genome-wide identification of BESI target genes in Arabidopsis thaliana. Plant J. 2011, 65, 634–646. [Google Scholar] [CrossRef]
- Bajguz, A.; Asami, T. Effects of brassinazole, an inhibitor of brassinosteroid biosynthesis, on light- and dark-grown Chlorella vulgaris. Planta 2004, 218, 869–877. [Google Scholar] [CrossRef]
- Talarek-Karwel, M.; Bajguz, A.; Piotrowska-Niczyporuk, A.; Rajewska, I. The effect of 24-epibrassinolide on the green alga Acutodesmus obliquus (Chlorophyceae). Plant Physiol. Biochem. 2018, 124, 175–183. [Google Scholar] [CrossRef]
- Xia, X.J.; Huang, L.F.; Zhou, Y.H.; Mao, W.H.; Shi, K.; Wu, J.X.; Asami, T.; Chen, Z.; Yu, J.Q. Brassinosteroids promote photosynthesis and growth by enhancing activation of Rubisco and expression of photosynthetic genes in Cucumis sativus. Planta 2009, 230, 1185–1196. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cai, Z.; Xie, Z.; Wang, X.; Zhang, S.; Wu, Q.; Yu, X.; Guo, Y.; Gao, S.; Zhang, Y.; Xu, S.; et al. Excavation of Genes Responsive to Brassinosteroids by Transcriptome Sequencing in Adiantum flabellulatum Gametophytes. Genes 2022, 13, 1061. https://doi.org/10.3390/genes13061061
Cai Z, Xie Z, Wang X, Zhang S, Wu Q, Yu X, Guo Y, Gao S, Zhang Y, Xu S, et al. Excavation of Genes Responsive to Brassinosteroids by Transcriptome Sequencing in Adiantum flabellulatum Gametophytes. Genes. 2022; 13(6):1061. https://doi.org/10.3390/genes13061061
Chicago/Turabian StyleCai, Zeping, Zhenyu Xie, Xiaochen Wang, Shuixian Zhang, Qian Wu, Xudong Yu, Yi Guo, Shuyi Gao, Yunge Zhang, Shitao Xu, and et al. 2022. "Excavation of Genes Responsive to Brassinosteroids by Transcriptome Sequencing in Adiantum flabellulatum Gametophytes" Genes 13, no. 6: 1061. https://doi.org/10.3390/genes13061061