
Genome-Wide Association Study Reveals Genetic Architecture and Candidate Genes for Yield and Related Traits under Terminal Drought, Combined Heat and Drought in Tropical Maize Germplasm

 ,
,  ,
,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Genetic Resources, Field Evaluation, and Phenotyping
2.2. Statistical Analysis
2.3. Genome-Wide Association Study Mapping
2.4. Candidate Gene Prediction and In Silico Analyses
3. Results
3.1. Phenotypic Variation, Correlation, and Heritability among the Tropical Maize Germplasm
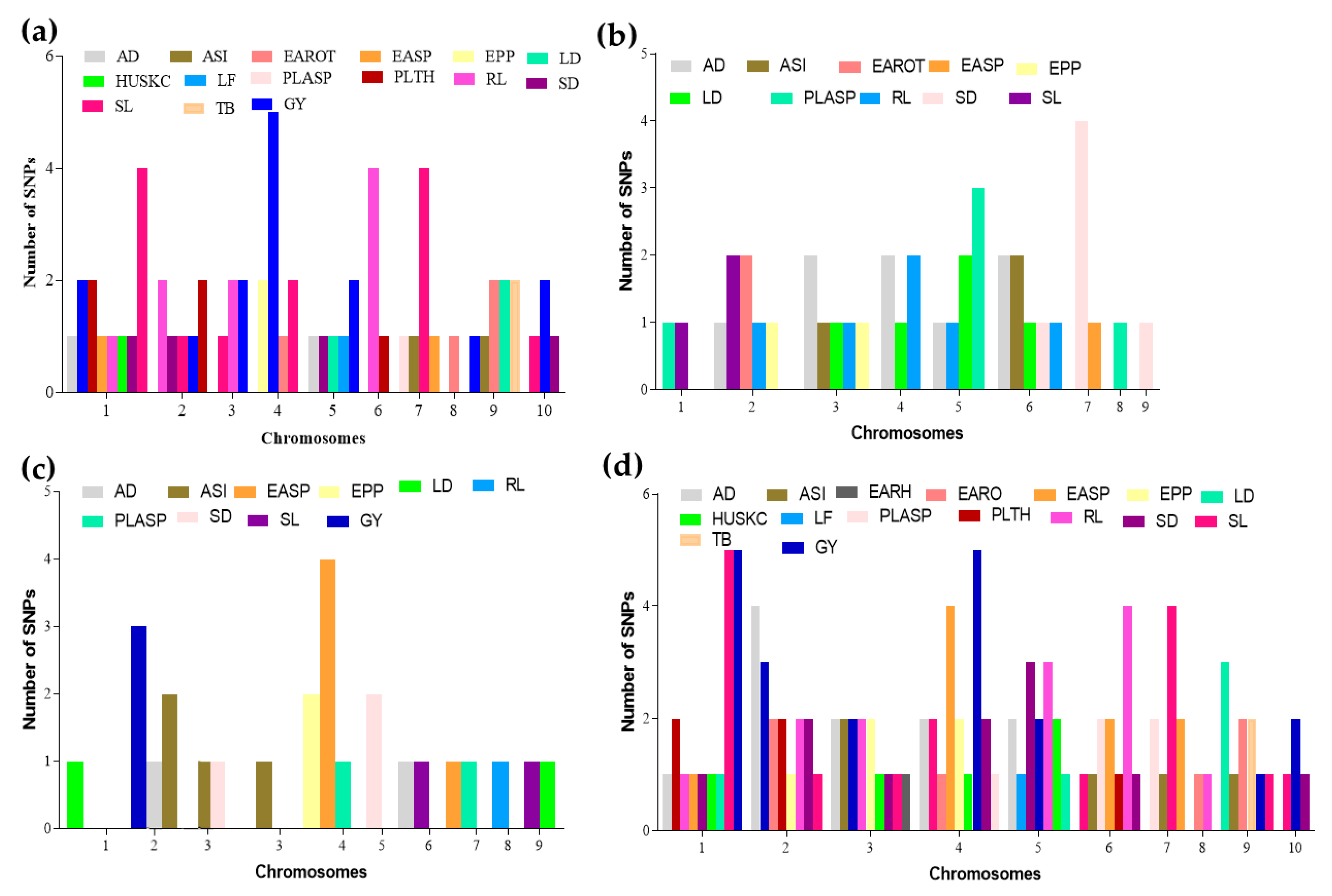
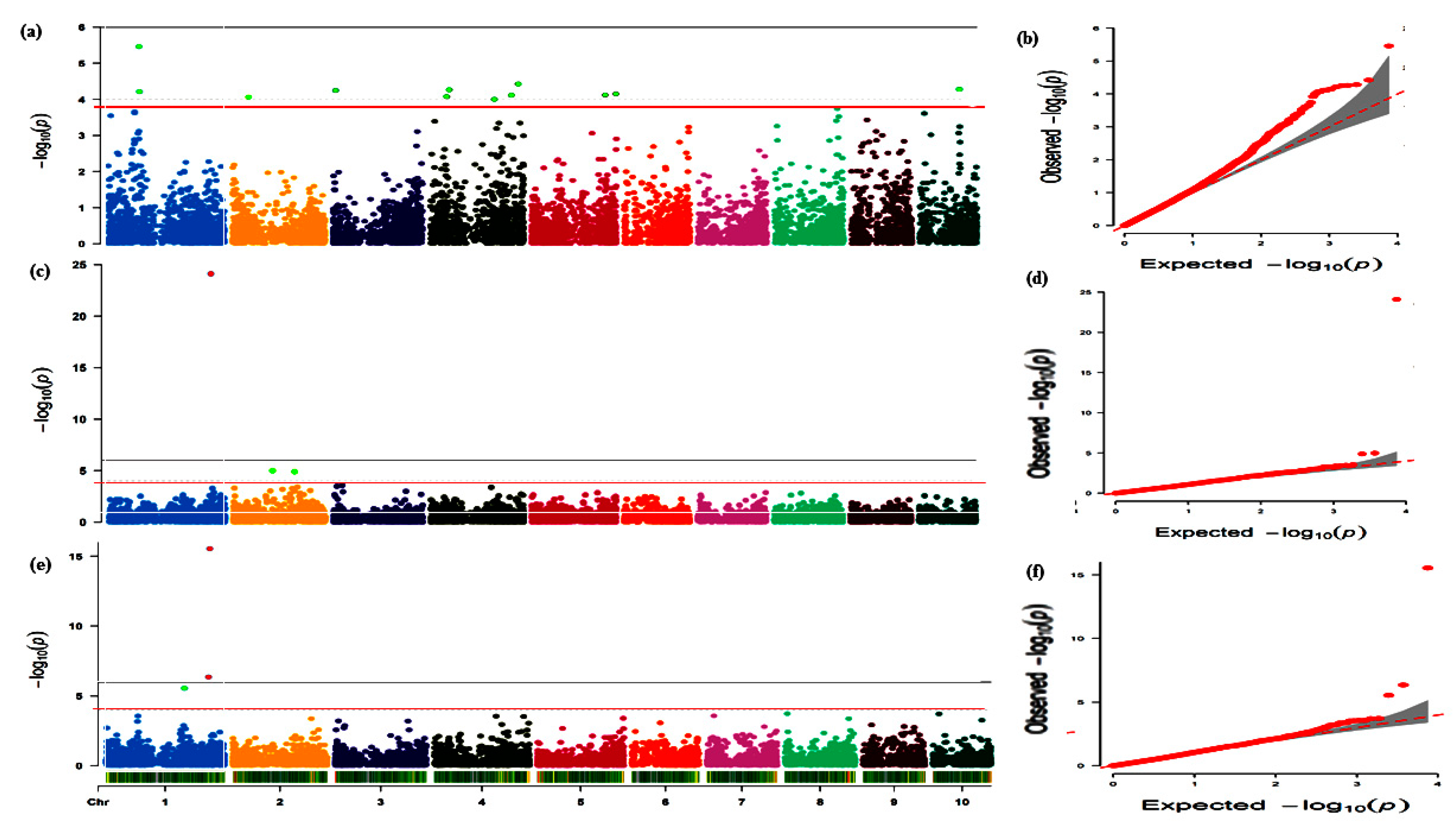
3.2. Marker–Trait Association under Combined Heat and Drought Conditions
3.3. Marker–Trait Associations under Terminal Drought Condition
3.4. Mapping Using Data Averaged across Combined Heat and Drought Conditions plus the Terminal Drought Conditions
3.5. Comparison of Mapping Results from the Three Conditions (CHD, TD, and Combined)
3.6. Candidate Gene Prediction and In Silico Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hickey, L.T.; Hafeez, A.N.; Robinson, H.; Jackson, S.A.; Leal-Bertioli, S.C.M.; Tester, M.; Gao, C.; Godwin, I.D.; Hayes, B.J.; Wulff, B.B.H. Breeding crops to feed 10 billion. Nat. Biotechnol. 2019, 37, 744–754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cairns, J.E.; Prasanna, B.M. Developing and deploying climate-resilient maize varieties in the developing world. Curr. Opin. Plant Biol. 2018, 45, 226–230. [Google Scholar] [CrossRef] [PubMed]
- Badu-Apraku, B.; Fakorede, M.A.B. Maize in Sub-Saharan Africa: Importance and production constraints. In Advances in Genetic Enhancement of Early and Extra-Early Maize for Sub-Saharan Africa; Springer: Berlin/Heidelberg, Germany, 2017; pp. 3–10. [Google Scholar]
- Mageto, E.K.; Makumbi, D.; Njoroge, K.; Nyankanga, R. Genetic analysis of early-maturing maize (Zea Mays L.) inbred lines under stress and nonstress conditions. J. Crop Improv. 2017, 31, 560–588. [Google Scholar] [CrossRef]
- Ray, D.K.; Gerber, J.S.; MacDonald, G.K.; West, P.C. Climate variation explains a third of global crop yield variability. Nat. Commun. 2015, 6, 5989. [Google Scholar] [CrossRef] [Green Version]
- Cairns, J.E.; Crossa, J.; Zaidi, P.H.; Grudloyma, P.; Sanchez, C.; Araus, J.L.; Thaitad, S.; Makumbi, D.; Magorokosho, C.; Bänziger, M. Identification of drought, heat, and combined drought and heat tolerant donors in maize. Crop Sci. 2013, 53, 1335–1346. [Google Scholar] [CrossRef] [Green Version]
- Knox, J.; Hess, T.; Daccache, A.; Wheeler, T. Climate change impacts on crop productivity in Africa and South Asia. Environ. Res. Lett. 2012, 7, 034032. [Google Scholar] [CrossRef]
- Lobell, D.B.; Hammer, G.L.; McLean, G.; Messina, C.; Roberts, M.J.; Schlenker, W. The critical role of extreme heat for maize production in the United States. Nat. Clim. Chang. 2013, 3, 497–501. [Google Scholar] [CrossRef]
- Leipner, J.; Stehli, A.; Soldati, A. Photosynthetic performance of exotic maize (Zea mays L.) germplasm from tropical highlands at low and high temperature. Angew. Bot. 1999, 73, 20–24. [Google Scholar]
- Cárcova, J.; Otegui, M.E. Ear Temperature and Pollination Timing Effects on Maize Kernel Set. Crop Sci. 2001, 41, 1809–1815. [Google Scholar] [CrossRef]
- Yuan, Y.; Cairns, J.E.; Babu, R.; Gowda, M.; Makumbi, D.; Magorokosho, C.; Zhang, A.; Liu, Y.; Wang, N.; Hao, Z.; et al. Genome-Wide Association Mapping and Genomic Prediction Analyses Reveal the Genetic Architecture of Grain Yield and Flowering Time Under Drought and Heat Stress Conditions in Maize. Front. Plant Sci. 2019, 9, 1919. [Google Scholar] [CrossRef] [Green Version]
- Wallace, J.G.; Zhang, X.; Beyene, Y.; Semagn, K.; Olsen, M.; Prasanna, B.M.; Buckler, E.S. Genome-wide Association for Plant Height and Flowering Time across 15 Tropical Maize Populations under Managed Drought Stress and Well-Watered Conditions in Sub-Saharan Africa. Crop Sci. 2016, 56, 2365–2378. [Google Scholar] [CrossRef] [Green Version]
- Millet, E.J.; Welcker, C.; Kruijer, W.; Negro, S.; Coupel-Ledru, A.; Nicolas, S.D.; Laborde, J.; Bauland, C.; Praud, S.; Ranc, N.; et al. Genome-Wide Analysis of Yield in Europe: Allelic Effects Vary with Drought and Heat Scenarios. Plant Physiol. 2016, 172, 749–764. [Google Scholar] [CrossRef] [PubMed]
- Longmei, N.; Gill, G.K.; Zaidi, P.H.; Kumar, R.; Nair, S.K.; Hindu, V.; Vinayan, M.T.; Vikal, Y. Genome wide association mapping for heat tolerance in sub-tropical maize. BMC Genom. 2021, 22, 154. [Google Scholar] [CrossRef] [PubMed]
- Thornton, P.K.; Ericksen, P.J.; Herrero, M.; Challinor, A.J. Climate variability and vulnerability to climate change: A review. Glob. Chang. Biol. 2014, 20, 3313–3328. [Google Scholar] [CrossRef] [PubMed]
- Teshome, A.; Zhang, J. Increase of Extreme Drought over Ethiopia under Climate Warming. Adv. Meteorol. 2019, 2019, 5235429. [Google Scholar] [CrossRef] [Green Version]
- Asante, F.A.; Amuakwa-Mensah, F. Climate Change and Variability in Ghana: Stocktaking. Climate 2015, 3, 78–99. [Google Scholar] [CrossRef] [Green Version]
- Nelimor, C.; Badu-Apraku, B.; Tetteh, A.Y.; Garcia-Oliveira, A.L.; N’guetta, A.S. Assessing the Potential of Extra-Early Maturing Landraces for Improving Tolerance to Drought, Heat, and Both Combined Stresses in Maize. Agronomy 2020, 10, 318. [Google Scholar] [CrossRef] [Green Version]
- Cantarero, M.G.; Cirilo, A.G.; Andrade, F.H. Night Temperature at Silking Affects Set in Maize. Crop Sci. 1999, 39, 703–710. [Google Scholar] [CrossRef]
- Rattalino, E.J.I.; Otegui, M.E. Heat stress in temperate and tropical maize hybrids: A novel approach for assessing sources of kernel loss in field conditions. Field Crops Research 2013, 142, 58–67. [Google Scholar] [CrossRef]
- Ben-Asher, J.; Garcia y Garcia, A.; Hoogenboom, G. Effect of high temperature on photosynthesis and transpiration of sweet corn (Zea mays L. var. rugosa). Photosynthetica 2008, 46, 595–603. [Google Scholar] [CrossRef]
- Ristic, Z.; Yang, G.; Martin, B.; Fullerton, S. Evidence of association between specific heat-shock protein(s) and the drought and heat tolerance phenotype in maize. J. Plant Physiol. 1998, 153, 497–505. [Google Scholar] [CrossRef]
- Ribaut, J.-M.; Betran, J.; Monneveux, P.; Setter, T. Drought tolerance in maize. In Handbook of Maize: Its Biology; Bennetzen, J.L., Hake, S.C., Eds.; Springer: New York, NY, USA, 2009; pp. 311–344. [Google Scholar]
- Barbosa, P.A.M.; Fritsche-Neto, R.; Andrade, M.C.; Petroli, C.D.; Burgueño, J.; Galli, G.; Willcox, M.C.; Sonder, K.; Vidal-Martínez, V.A.; Sifuentes-Ibarra, E.; et al. Introgression of Maize Diversity for Drought Tolerance: Subtropical Maize Landraces as Source of New Positive Variants. Front. Plant Sci. 2021, 12, 12. [Google Scholar] [CrossRef] [PubMed]
- Chávez-Arias, C.C.; Ligarreto-Moreno, G.A.; Ramírez-Godoy, A.; Restrepo-Díaz, H. Maize Responses Challenged by Drought, Elevated Daytime Temperature and Arthropod Herbivory Stresses: A Physiological, Biochemical and Molecular View. Front. Plant Sci. 2021, 12, 12. [Google Scholar] [CrossRef]
- Sattar, A.; Sher, A.; Ijaz, M.; Ul-Allah, S.; Rizwan, M.S.; Hussain, M.; Jabran, K.; Cheema, M.A. Terminal drought and heat stress alter physiological and biochemical attributes in flag leaf of bread wheat. PLoS ONE 2020, 15, e0232974. [Google Scholar] [CrossRef] [PubMed]
- Awasthi, R.; Kaushal, N.; Vadez, V.; Turner, N.C.; Berger, J.; Siddique, K.H.M.; Nayyar, H. Individual and combined effects of transient drought and heat stress on carbon assimilation and seed filling in chickpea. Funct. Plant Biol. 2014, 41, 1148–1167. [Google Scholar] [CrossRef] [Green Version]
- Urban, O.; Hlaváčová, M.; Klem, K.; Novotná, K.; Rapantová, B.; Smutná, P.; Horáková, V.; Hlavinka, P.; Škarpa, P.; Trnka, M. Combined effects of drought and high temperature on photosynthetic characteristics in four winter wheat genotypes. Field Crops Res. 2018, 223, 137–149. [Google Scholar] [CrossRef]
- Tesfaye, K.; Kruseman, G.; Cairns, J.E.; Zaman-Allah, M.; Wegary, D.; Zaidi, P.H.; Boote, K.J.; Rahut, D.; Erenstein, O. Potential benefits of drought and heat tolerance for adapting maize to climate change in tropical environments. Clim. Risk Manag. 2018, 19, 106–119. [Google Scholar] [CrossRef]
- Hussain, H.A.; Men, S.; Hussain, S.; Chen, Y.; Ali, S.; Zhang, S.; Zhang, K.; Li, Y.; Xu, Q.; Liao, C.; et al. Interactive effects of drought and heat stresses on morpho-physiological attributes, yield, nutrient uptake and oxidative status in maize hybrids. Sci. Rep. 2019, 9, 3890. [Google Scholar] [CrossRef] [Green Version]
- Azodi, C.B.; Lloyd, J.P.; Shiu, S.-H. The cis-regulatory codes of response to combined heat and drought stress in Arabidopsis thaliana. NAR Genom. Bioinform. 2020, 2, lqaa049. [Google Scholar] [CrossRef]
- Trachsel, S.; Leyva, M.; Lopez, M.; Suarez, E.A.; Mendoza, A.; Montiel, N.G.; Macias, M.S.; Burgueno, J.; San Vicente, F. Identification of Tropical Maize Germplasm with Tolerance to Drought, Nitrogen Deficiency, and Combined Heat and Drought Stresses. Crop Sci. 2016, 56, 3031–3045. [Google Scholar] [CrossRef]
- Osuman, A.S.; Badu-Apraku, B.; Ifie, B.E.; Tongoona, P.; Obeng-Bio, E.; Garcia-Oliveira, A.L. Genetic Diversity, Population Structure and Inter-Trait Relationships of Combined Heat and Drought Tolerant Early-Maturing Maize Inbred Lines from West and Central Africa. Agronomy 2020, 10, 1324. [Google Scholar] [CrossRef]
- Kamara, A.Y.; Menkir, A.; Badu-Apraku, B.; Ibikunle, O. The influence of drought stress on growth, yield and yield components of selected maize genotypes. J. Agric. Sci. 2003, 141, 43–50. [Google Scholar] [CrossRef]
- Li, M.; Zhong, W.; Yang, F.; Zhang, Z. Genetic and Molecular Mechanisms of Quantitative Trait Loci Controlling Maize Inflorescence Architecture. Plant Cell Physiol. 2018, 59, 448–457. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Liu, Y.-H.; Xu, W.; Smith, C.W.; Murray, S.C.; Zhang, H.-B. Analysis of the genes controlling three quantitative traits in three diverse plant species reveals the molecular basis of quantitative traits. Sci. Rep. 2020, 10, 10074. [Google Scholar] [CrossRef]
- Karikari, B.; Wang, Z.; Zhou, Y.; Yan, W.; Feng, J.; Zhao, T. Identification of quantitative trait nucleotides and candidate genes for soybean seed weight by multiple models of genome-wide association study. BMC Plant Biol. 2020, 20, 404. [Google Scholar] [CrossRef] [PubMed]
- Karikari, B.; Chen, S.; Xiao, Y.; Chang, F.; Zhou, Y.; Kong, J.; Bhat, J.A.; Zhao, T. Utilization of interspecific high-density genetic map of RIL population for the QTL detection and candidate gene mining for 100-seed weight in soybean. Front. Plant Sci. 2019, 10, 1001. [Google Scholar] [CrossRef]
- Alqudah, A.M.; Sallam, A.; Stephen Baenziger, P.; Börner, A. GWAS: Fast-forwarding gene identification and characterization in temperate Cereals: Lessons from Barley—A review. J. Adv. Res. 2020, 22, 119–135. [Google Scholar] [CrossRef] [PubMed]
- Varshney, R.K.; Sinha, P.; Singh, V.K.; Kumar, A.; Zhang, Q.; Bennetzen, J.L. 5Gs for crop genetic improvement. Curr. Opin. Plant Biol. 2020, 56, 190–196. [Google Scholar] [CrossRef] [PubMed]
- Brachi, B.; Morris, G.P.; Borevitz, J.O. Genome-wide association studies in plants: The missing heritability is in the field. Genome Biol. 2011, 12, 232. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.K.; Prasad, A.; Muthamilarasan, M.; Parida, S.K.; Prasad, M. Breeding and biotechnological interventions for trait improvement: Status and prospects. Planta 2020, 252, 54. [Google Scholar] [CrossRef]
- Hao, Z.; Li, X.; Xie, C.; Weng, J.; Li, M.; Zhang, D.; Liang, X.; Liu, L.; Liu, S.; Zhang, S. Identification of Functional Genetic Variations Underlying Drought Tolerance in Maize Using SNP Markers. J. Integr. Plant Biol. 2011, 53, 641–652. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Li, K.; Hu, X.; Liu, Z.; Wu, Y.; Huang, C. Genome-wide association analysis of forage quality in maize mature stalk. BMC Plant Biol. 2016, 16, 227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Wang, H.; Liu, S.; Ferjani, A.; Li, J.; Yan, J.; Yang, X.; Qin, F. Genetic variation in ZmVPP1 contributes to drought tolerance in maize seedlings. Nat. Genet. 2016, 48, 1233–1241. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.A.; Seetharam, K.; Zaidi, P.H.; Dinesh, A.; Vinayan, M.T.; Nath, U.K. Dissecting heat stress tolerance in tropical maize (Zea mays L.). Field Crops Res. 2017, 204, 110–119. [Google Scholar] [CrossRef]
- Seetharam, K.; Kuchanur, P.H.; Koirala, K.B.; Tripathi, M.P.; Patil, A.; Sudarsanam, V.; Das, R.R.; Chaurasia, R.; Pandey, K.; Vemuri, H.; et al. Genomic regions associated with heat stress tolerance in tropical maize (Zea mays L.). Sci. Rep. 2021, 11, 13730. [Google Scholar] [CrossRef]
- Farfan, I.D.B.; de la Fuente, G.N.; Murray, S.C.; Isakeit, T.; Huang, P.-C.; Warburton, M.; Williams, P.; Windham, G.L.; Kolomiets, M. Genome Wide Association Study for Drought, Aflatoxin Resistance, and Important Agronomic Traits of Maize Hybrids in the Sub-Tropics. PLoS ONE 2015, 10, e0117737. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Qin, F. Genome-Wide Association Study Reveals Natural Variations Contributing to Drought Resistance in Crops. Front. Plant Sci. 2017, 8, 1110. [Google Scholar] [CrossRef] [Green Version]
- Moose, S.P.; Mumm, R.H. Molecular plant breeding as the foundation for 21st century crop improvement. Plant Physiol. 2008, 147, 969–977. [Google Scholar] [CrossRef] [Green Version]
- Peterson, B.G.; Carl, P.; Boudt, K.; Bennett, R.; Ulrich, J.; Zivot, E.; Cornilly, D.; Hung, E.; Lestel, M.; Balkissoon, K. Package ‘performanceanalytics’. R Team Coop. 2018, 3, 13–14. [Google Scholar]
- Nyquist, W.E.; Baker, R.J. Estimation of heritability and prediction of selection response in plant populations. Crit. Rev. Plant Sci. 1991, 10, 235–322. [Google Scholar] [CrossRef]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.W.; Daly, M.J. PLINK: A tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef]
- Yu, J.; Pressoir, G.; Briggs, W.H.; Vroh Bi, I.; Yamasaki, M.; Doebley, J.F.; McMullen, M.D.; Gaut, B.S.; Nielsen, D.M.; Holland, J.B.; et al. A unified mixed-model method for association mapping that accounts for multiple levels of relatedness. Nat. Genet. 2006, 38, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Woodhouse, M.R.; Sen, S.; Schott, D.; Portwood, J.L.; Freeling, M.; Walley, J.W.; Andorf, C.M.; Schnable, J.C. qTeller: A tool for comparative multi-genomic gene expression analysis. Bioinformatics 2021, 38, 236–242. [Google Scholar] [CrossRef]
- Forestan, C.; Aiese Cigliano, R.; Farinati, S.; Lunardon, A.; Sanseverino, W.; Varotto, S. Stress-induced and epigenetic-mediated maize transcriptome regulation study by means of transcriptome reannotation and differential expression analysis. Sci. Rep. 2016, 6, 30446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waters, A.J.; Makarevitch, I.; Noshay, J.; Burghardt, L.T.; Hirsch, C.N.; Hirsch, C.D.; Springer, N.M. Natural variation for gene expression responses to abiotic stress in maize. Plant J. 2017, 89, 706–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene Ontology: Tool for the unification of biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [Green Version]
- Finn, R.D.; Bateman, A.; Clements, J.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Heger, A.; Hetherington, K.; Holm, L.; Mistry, J.; et al. Pfam: The protein families database. Nucleic Acids Res. 2014, 42, D222–D230. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M.; Goto, S. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Goodstein, D.M.; Shu, S.; Howson, R.; Neupane, R.; Hayes, R.D.; Fazo, J.; Mitros, T.; Dirks, W.; Hellsten, U.; Putnam, N. Phytozome: A comparative platform for green plant genomics. Nucleic Acids Res. 2012, 40, D1178–D1186. [Google Scholar] [CrossRef] [PubMed]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Falcone, E.; Grandoni, L.; Garibaldi, F.; Manni, I.; Filligoi, G.; Piaggio, G.; Gurtner, A. Infinity: An In-Silico Tool for Genome-Wide Prediction of Specific DNA Matrices in miRNA Genomic Loci. PLoS ONE 2016, 11, e0153658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuteja, N.; Singh, S.; Tuteja, R. Helicases in improving abiotic stress tolerance in crop plants. Improv. Crop Resist. Abiotic Stress 2012, 435–449. [Google Scholar]
- Sami, A.A.; Arabia, S.; Sarker, R.H.; Islam, T. Deciphering the role of helicases and translocases: A multifunctional gene family safeguarding plants from diverse environmental adversities. Curr. Plant Biol. 2021, 26, 100204. [Google Scholar] [CrossRef]
- Bruce, W.B.; Edmeades, G.O.; Barker, T.C. Molecular and physiological approaches to maize improvement for drought tolerance. J. Exp. Bot. 2002, 53, 13–25. [Google Scholar] [CrossRef]
- Campos, H.; Cooper, M.; Habben, J.E.; Edmeades, G.O.; Schussler, J.R. Improving drought tolerance in maize: A view from industry. Field Crops Res. 2004, 90, 19–34. [Google Scholar] [CrossRef]
- Gedil, M.; Menkir, A. An Integrated Molecular and Conventional Breeding Scheme for Enhancing Genetic Gain in Maize in Africa. Front. Plant Sci. 2019, 10, 1430. [Google Scholar] [CrossRef]
- Lunduka, R.W.; Mateva, K.I.; Magorokosho, C.; Manjeru, P. Impact of adoption of drought-tolerant maize varieties on total maize production in south Eastern Zimbabwe. Clim. Dev. 2019, 11, 35–46. [Google Scholar] [CrossRef] [Green Version]
- Cerrudo, D.; Cao, S.; Yuan, Y.; Martinez, C.; Suarez, E.A.; Babu, R.; Zhang, X.; Trachsel, S. Genomic Selection Outperforms Marker Assisted Selection for Grain Yield and Physiological Traits in a Maize Doubled Haploid Population Across Water Treatments. Front. Plant Sci. 2018, 9, 366. [Google Scholar] [CrossRef] [Green Version]
- Semagn, K.; Magorokosho, C.; Vivek, B.S.; Makumbi, D.; Beyene, Y.; Mugo, S.; Prasanna, B.M.; Warburton, M.L. Molecular characterization of diverse CIMMYT maize inbred lines from eastern and southern Africa using single nucleotide polymorphic markers. BMC Genom. 2012, 13, 113. [Google Scholar] [CrossRef] [Green Version]
- Chapman, S.C.; Edmeades, G.O. Selection improves tolerance to mid/late season drought in tropical maize populations. II. Direct and correlated responses among secondary traits. Crop Sci. 1999, 39, 1315–1324. [Google Scholar] [CrossRef]
- Benchikh-Lehocine, M.; Revilla, P.; Malvar, R.A.; Djemel, A. Response to Selection for Reduced Anthesis-Silking Interval in Four Algerian Maize Populations. Agronomy 2021, 11, 382. [Google Scholar] [CrossRef]
- Xu, Y.; Liu, X.; Fu, J.; Wang, H.; Wang, J.; Huang, C.; Prasanna, B.M.; Olsen, M.S.; Wang, G.; Zhang, A. Enhancing Genetic Gain through Genomic Selection: From Livestock to Plants. Plant Commun. 2020, 1, 100005. [Google Scholar] [CrossRef] [PubMed]
- Tuberosa, R.; Salvi, S.; Sanguineti, M.C.; Landi, P.; Maccaferri, M.; Conti, S. Mapping QTLs regulating morpho-physiological traits and yield: Case studies, shortcomings and perspectives in drought-stressed maize. Ann. Bot. 2002, 89, 941–963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Awata, L.A.O.; Beyene, Y.; Gowda, M.; Surech, M.L.; Jumbo, M.B.; Tongoona, P.; Danquah, E.; Ifie, B.E.; Marchelo-Dragga, P.W.; Olsen, M.; et al. Genetic Analysis of QTL for Resistance to Maize Lethal Necrosis in Multiple Mapping Populations. Genes 2020, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maitra Majee, S.; Sharma, E.; Singh, B.; Khurana, J.P. Drought-induced protein (Di19-3) plays a role in auxin signaling by interacting with IAA14 in Arabidopsis. Plant Direct 2020, 4, e00234. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Liu, Y.; Li, M.; Lamin-Samu, A.T.; Yang, D.; Yu, X.; Izhar, M.; Jan, I.; Ali, M.; Lu, G. The Arabidopsis SMALL AUXIN UP RNA32 Protein Regulates ABA-Mediated Responses to Drought Stress. Front. Plant Sci. 2021, 12, 259. [Google Scholar] [CrossRef]
- Grundy, J.; Stoker, C.; Carré, I.A. Circadian regulation of abiotic stress tolerance in plants. Front. Plant Sci. 2015, 6, 648. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Marker a | Chr. b | Pos. c | AD | EARH | EARO | EASP | EPP | HUSKC | RL | SD | SL | GY |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SNP_91146889 | 1 | 91146889 | - | - | - | - | - | - | 4.00 (14.58) | - | - | - |
| SNP_269120178 | 1 | 269120178 | - | - | - | - | - | - | - | - | - | 24.11 (64.79) |
| SNP_3121382 | 2 | 3121382 | 4.00 (13.77) | - | - | - | - | - | - | 3.90 (13.29) | - | - |
| SNP_24521844 | 2 | 24521844 | - | - | 5.30 (18.09) | - | - | - | - | - | - | - |
| SNP_27502516 | 2 | 27502516 | - | - | 6.74 (23.99) | - | - | - | - | - | - | - |
| SNP_51109816 | 2 | 51109816 | - | - | - | - | 4.00 (14.71) | - | - | - | - | - |
| SNP_104170418 | 2 | 104170418 | - | - | - | - | - | - | - | - | - | 4.99 (16.77) |
| SNP_162101608 | 2 | 162101608 | - | - | - | - | - | - | - | - | - | 4.89 (16.74) |
| SNP_75795682 | 3 | 75795682 | - | 3.96 (15.12) | - | - | - | - | - | - | - | - |
| SNP_93411827 | 3 | 93411827 | 3.97 (12.84) | - | - | - | - | - | - | - | - | - |
| SNP_99059711 | 3 | 99059711 | - | - | - | - | 3.98 (13.45) | - | - | - | - | - |
| SNP_138825271 | 3 | 138825271 | 5.62 (18.79) | - | - | - | - | 4.19 (13.84) | - | 5.23 (17.32) | - | - |
| SNP_199184898 | 4 | 199184898 | 3.92 (12.53) | - | - | - | - | - | - | 4.26 (13.57) | - | - |
| SNP_244895453 | 4 | 244895453 | 5.62 (18.79) | - | - | - | - | 4.19 (13.85) | - | 5.23 (17.32) | - | - |
| SNP_168561609 | 5 | 168561609 | 5.60 (19.79) | - | - | - | - | 4.39 (14.75) | - | 5.26 (18.43) | - | - |
| SNP_82875264 | 5 | 82875264 | - | - | - | - | - | - | 4.08 (13.86) | - | - | - |
| SNP_164264714 | 5 | 164264714 | - | - | - | - | - | - | 5.21 (18.97) | - | - | - |
| SNP_181875996 | 5 | 181875996 | - | - | - | - | - | - | 4.30 (14.52) | - | - | - |
| SNP_204485620 | 5 | 204485620 | - | - | - | - | - | 4.17 (13.22) | - | - | - | - |
| SNP_62970998 | 6 | 62970998 | 5.38 (19.96) | - | - | - | - | 4.18 (15.44) | - | 5.12 (18.90) | - | - |
| SNP_93721476 | 6 | 93721476 | - | 5.25 (18.10) | - | - | - | - | - | - | - | - |
| SNP_125871560 | 6 | 125871560 | - | - | - | - | - | - | - | - | 4.39 (14.72) | - |
| SNP_128658596 | 6 | 128658596 | 4.01 (13.60) | - | - | - | - | - | - | - | - | - |
| SNP_165897715 | 6 | 165897715 | - | 4.34 (15.42) | - | - | - | - | - | - | - | - |
| SNP_158184157 | 7 | 158184157 | - | - | - | 4.07 (13.84) | - | - | - | - | - | - |
| SNP_43475091 | 8 | 43475091 | - | - | - | - | - | - | 4.08 (13.86) | - | - | - |
| SNP_89731392 | 9 | 89731392 | - | - | - | - | - | - | 5.71 (19.26) | - | - | - |
| Marker a | Chr. b | Pos. c | AD | ASI | EARO | EASP | EPP | LD | PLASP | RL | SD | SL | GY |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SNP_26125481 | 1 | 26125481 | - | - | - | - | - | 3.94 (13.84) | - | - | - | - | - |
| SNP_203396231 | 1 | 203396231 | - | - | - | - | - | - | - | - | - | - | 5.55 (19.17) |
| SNP_265278865 | 1 | 265278865 | - | - | - | - | - | - | - | - | - | - | 6.36 (22.12) |
| SNP_269120178 | 1 | 269120178 | - | - | - | - | - | - | - | - | - | - | 15.55 (47.77) |
| SNP_11473743 | 2 | 11473743 | 5.11 (17.76) | - | - | - | - | - | - | - | 4.15 (14.52) | - | - |
| SNP_24521844 | 2 | 24521844 | - | - | 5.11 (17.42) | - | - | - | - | - | - | - | - |
| SNP_27502516 | 2 | 27502516 | - | - | 6.38 (22.72) | - | - | - | - | - | - | - | - |
| SNP_1001688 | 3 | 1001688 | - | 4.22 (15.56) | - | - | - | - | - | - | - | - | - |
| SNP_99059711 | 3 | 99059711 | - | - | - | - | 5.62 (19.08) | - | - | - | - | - | - |
| SNP_112491022 | 3 | 112491022 | - | - | - | - | 4.59 (17.42) | - | - | - | - | - | - |
| SNP_41272646 | 4 | 41272646 | - | - | - | 4.24 (13.99) | - | - | - | - | - | - | - |
| SNP_137415099 | 4 | 137415099 | - | - | - | 5.35 (17.88) | - | - | - | - | - | - | - |
| SNP_166608728 | 4 | 166608728 | - | - | - | - | - | - | 4.18 (14.24) | - | - | - | - |
| SNP_176971048 | 4 | 176971048 | - | - | - | 3.99 (13.17) | - | - | - | - | - | - | - |
| SNP_218713681 | 4 | 218713681 | - | - | - | 4.22 (14.08) | - | - | - | - | - | - | - |
| SNP_82875264 | 5 | 82875264 | - | - | - | - | - | - | - | 4.09 (13.94) | - | - | - |
| SNP_164264714 | 5 | 164264714 | - | - | - | - | - | - | - | 5.12 (18.43) | - | - | - |
| SNP_101458697 | 6 | 101458697 | 4.07 (13.05) | - | - | - | - | - | - | - | - | - | - |
| SNP_125871560 | 6 | 125871560 | - | - | - | - | - | - | - | - | - | 4.69 (15.87) | - |
| SNP_169223617 | 6 | 169223617 | - | - | - | 4.36 (15.10) | - | - | - | - | - | - | - |
| SNP_51772182 | 7 | 51772182 | - | - | - | - | - | - | 4.55 (16.22) | - | - | - | - |
| SNP_43475091 | 8 | 43475091 | - | - | - | - | - | - | - | 4.09 (13.94) | - | - | - |
| SNP_89731392 | 9 | 89731392 | - | - | - | - | - | - | - | - | - | 5.44 (18.44) | - |
| SNP_158056460 | 9 | 158056460 | - | - | - | - | - | 4.30 (15.33) | - | - | - | - | - |
| SNPs a | Chr. b | Conditions c | ||
|---|---|---|---|---|
| CHD | TD | Combined Effects | ||
| SNP_269120178 | 1 | - | YIELD | YIELD |
| SNP_11473743 | 2 | AD | AD and SD | - |
| SNP_24521844 | 2 | - | EARO | EARO |
| SNP_27502516 | 2 | - | EARO | EARO |
| SNP_99059711 | 3 | - | EPP | EPP |
| SNP_82875264 | 5 | - | RL | RL |
| SNP_164264714 | 5 | - | RL | RL |
| SNP_125871560 | 6 | - | SL | SL |
| SNP_51772182 | 7 | EASP and PLASP | PLASP | - |
| SNP_43475091 | 8 | - | RL | RL |
| SNP_89731392 | 9 | - | SL | RL |
| SNP_158056460 | 9 | LD | LD | - |
| SNPs a | Chr. b | Conditions c | Gene: Position (bp) d | Description e | ||
|---|---|---|---|---|---|---|
| CHD | TD | Combined Effects | ||||
| SNP_269120178 | 1 | - | GY | GY | Zm00001d033620: 269012383–269014829 | Peptide α-N-acetyltransferase/Protein N-terminal acetyltransferase |
| SNP_11473743 | 2 | AD | - | AD & SD | Zm00001d002374: 11368789–11369293 | Auxin induced-like protein |
| SNP_24521844 | 2 | - | EARO | EARO | Zm00001d002847: 24476397–24480168 | - (+)-neomenthol dehydrogenase/Monoterpenoid dehydrogenase |
| SNP_27502516 | 2 | - | EARO | EARO | Zm00001d002937: 27420577–27426388 | 11-oxo-β-amyrin 30-oxidase/CYP72A154//Secologanin synthase |
| SNP_99059711 | 3 | - | EPP | EPP | Zm00001d041124: 99054551–99065304 | Unknown |
| SNP_82875264 | 5 | - | RL | RL | Zm00001d015290: 82874339–82879720 | Clock-associated PAS protein ZTL (ZTL) |
| SNP_164264714 | 5 | - | RL | RL | Zm00001d016478: 164218445–164234669 | Glutathione-disulfide reductase/NADPH:oxidized-glutathione oxidoreductase//Thioredoxin-disulfide reductase/Thioredoxin reductase |
| SNP_125871560 | 6 | - | SL | SL | Zm00001d037455: 125931589–125933956 | Phosphoribosylanthranilate isomerase |
| SNP_51772182 | 7 | EASP & PLASP | - | PLASP | Zm00001d019699: 51748841–51764687 | Protein-serine/threonine phosphatase/Serine/threonine specific protein phosphatase |
| SNP_43475091 | 8 | - | RL | RL | Zm00001d009212: 43471862–43480271 | Dihydrolipoyl dehydrogenase/Lipoyl dehydrogenase |
| SNP_89731392 | 9 | - | RL | SL | Zm00001d046434: 89628666–89634933 | Acyl CoA binding protein (ACBP)//Kelch motif (Kelch_1)//Galactose oxidase, central domain (Kelch_3)//Kelch motif (Kelch_5) |
| SNP_158056460 | 9 | LD | - | LD | Zm00001d048531: 157939096–157942646 | RNA helicase |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Osuman, A.S.; Badu-Apraku, B.; Karikari, B.; Ifie, B.E.; Tongoona, P.; Danquah, E.Y. Genome-Wide Association Study Reveals Genetic Architecture and Candidate Genes for Yield and Related Traits under Terminal Drought, Combined Heat and Drought in Tropical Maize Germplasm. Genes 2022, 13, 349. https://doi.org/10.3390/genes13020349
Osuman AS, Badu-Apraku B, Karikari B, Ifie BE, Tongoona P, Danquah EY. Genome-Wide Association Study Reveals Genetic Architecture and Candidate Genes for Yield and Related Traits under Terminal Drought, Combined Heat and Drought in Tropical Maize Germplasm. Genes. 2022; 13(2):349. https://doi.org/10.3390/genes13020349
Chicago/Turabian StyleOsuman, Alimatu Sadia, Baffour Badu-Apraku, Benjamin Karikari, Beatrice Elohor Ifie, Pangirayi Tongoona, and Eric Yirenkyi Danquah. 2022. "Genome-Wide Association Study Reveals Genetic Architecture and Candidate Genes for Yield and Related Traits under Terminal Drought, Combined Heat and Drought in Tropical Maize Germplasm" Genes 13, no. 2: 349. https://doi.org/10.3390/genes13020349