
Genome-Wide Analysis in Drosophila Reveals the Genetic Basis of Variation in Age-Specific Physical Performance and Response to ACE Inhibition

 ,
,
Abstract
:1. Introduction
2. Methods
2.1. Drosophila Stocks, Maintenance, and Drug Treatment
2.2. Physical Performance Assays
2.3. Genome-Wide Association Tests of Physical Performance and Sensitivity to Lisinopril
2.4. Network Analyses of Physical Performance and Sensitivity to Lisinopril
2.5. Validation of Genes Associated with Physical Performance and Sensitivity to Lisinopril Using Muscle-Specific RNAi
3. Results
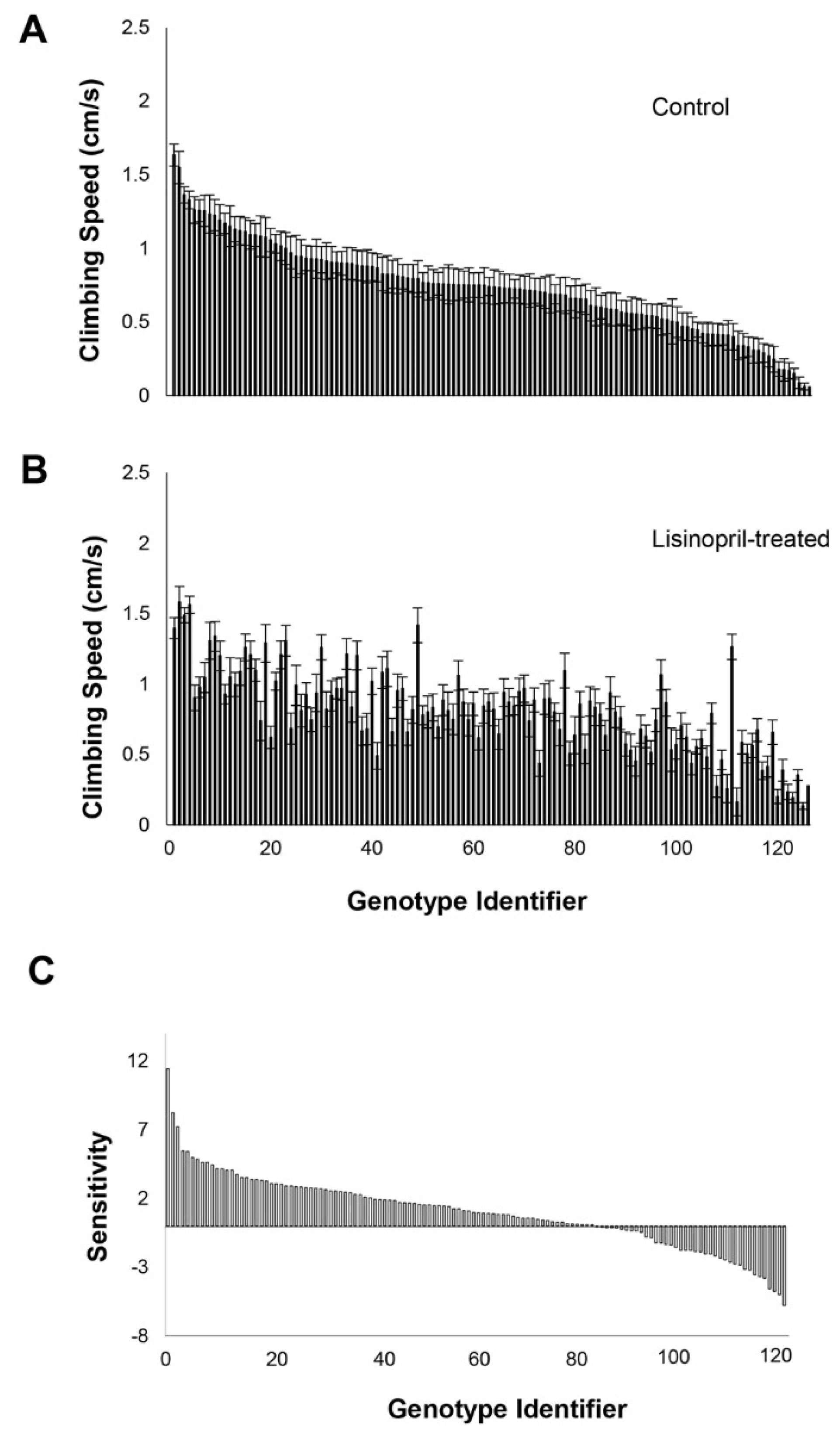
3.1. Genetic Variation in Response and Sensitivity to Lisinopril Treatment—Climbing Speed
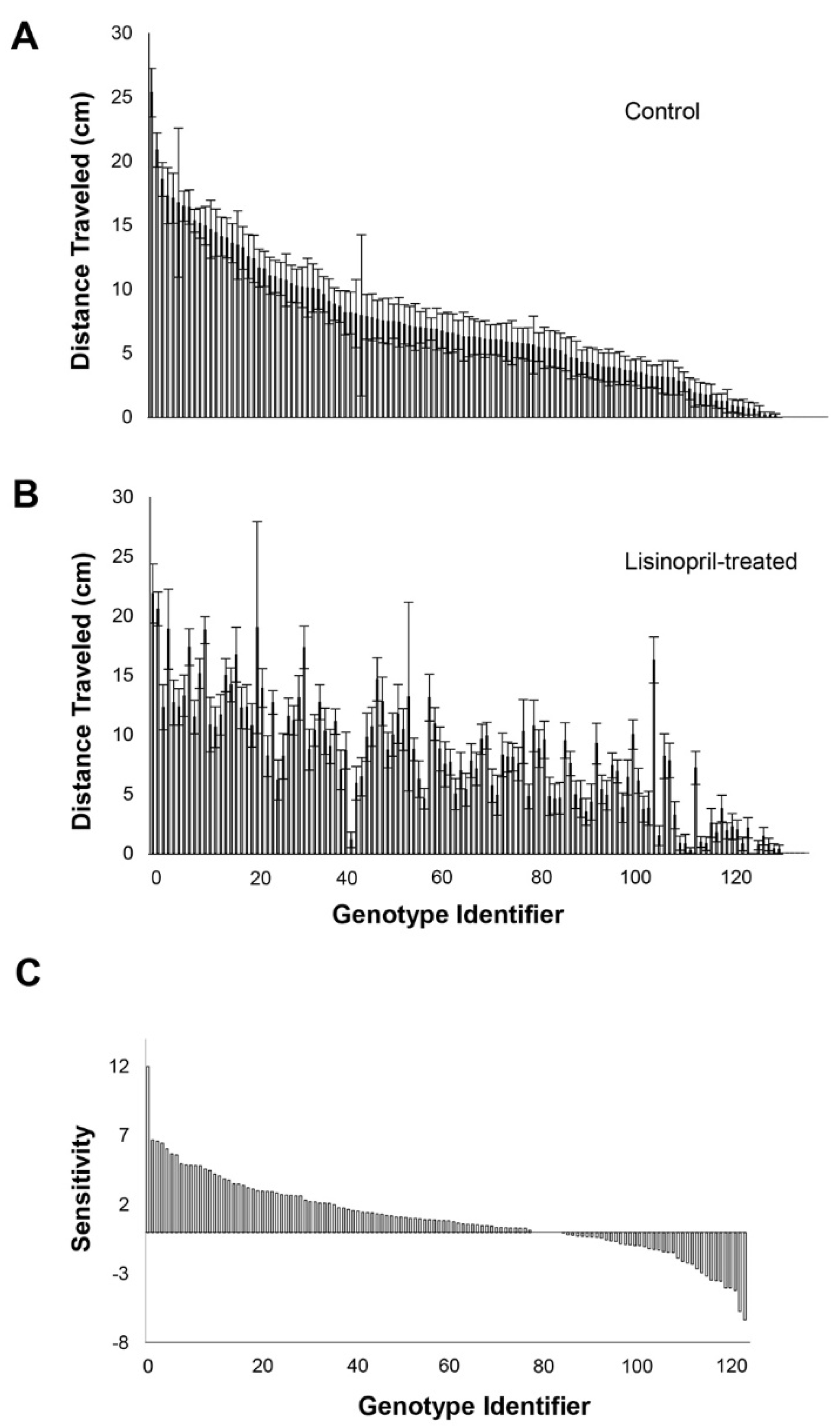
3.2. Genetic Variation in Response and Sensitivity to Lisinopril Treatment—Endurance
3.3. GWA of Effect of Lisinopril Treatment on Physical Performance and Sensitivity
3.4. Overlap of Candidate Genes Influencing Physical Performance and Sensitivity
3.5. Network Analysis of Climbing Speed
3.6. Network Analysis of Endurance
3.7. Network Analysis of Sensitivity to Lisinopril
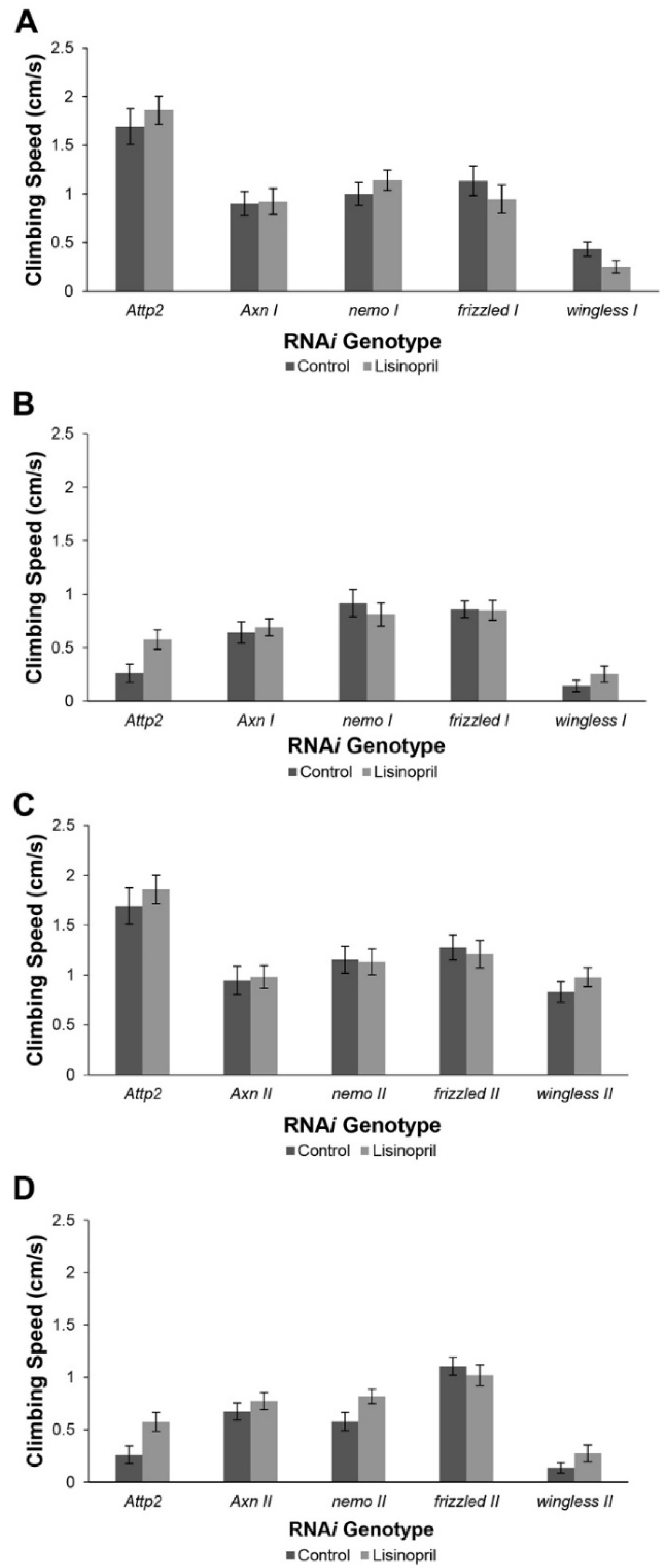
3.8. Muscle-Specific RNAi Implicates Genes in the WNT Signaling Pathway as Mediating the Effects of Lisinopril on Physical Performance
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Montgomery, H.E.; Marshall, R.; Hemingway, H.; Myerson, S.; Clarkson, P.; Dollery, C.; Hayward, M.; Holliman, D.E.; Jubb, M.; World, M.; et al. Human gene for physical performance. Nature 1998, 393, 221–222. [Google Scholar] [CrossRef]
- Wang, P.; Fedoruk, M.N.; Rupert, J.L. Keeping pace with ACE: Are ACE inhibitors and angiotensin II type 1 receptor antagonists potential doping agents? Sports Med. 2008, 38, 1065–1079. [Google Scholar] [CrossRef] [PubMed]
- Keogh, J.W.; Palmer, B.R.; Taylor, D.; Kilding, A.E. ACE and UCP2 gene polymorphisms and their association with baseline and exercise-related changes in the functional performance of older adults. PeerJ 2015, 3, e980. [Google Scholar] [CrossRef] [Green Version]
- Buford, T.W.; Hsu, F.C.B.; Brinkley, T.E. The angiotensin-converting enzyme I/D polymorphism and exercise-induced changes in physical function among Caucasian older adults. Med. Sci. Sports Exerc. 2014, 46, 598. [Google Scholar] [CrossRef]
- Abadir, P.M.; Walston, J.D.; Carey, R.M. Subcellular characteristics of functional intracellular renin-angiotensin systems. Peptides 2012, 38, 437–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buford, T.W.; Manini, T.M.; Hsu, F.C.; Cesari, M.; Anton, S.D.; Nayfield, S.; Stafford, R.S.; Church, T.S.; Pahor, M.; Carter, C.S. Angiotensin-converting enzyme inhibitor use by older adults is associated with greater functional responses to exercise. J. Am. Geriatr. Soc. 2012, 60, 1244–1252. [Google Scholar] [CrossRef] [PubMed]
- Carter, C.S.; Onder, G.; Kritchevsky, S.B.; Pahor, M. Angiotensin-converting enzyme inhibition intervention in elderly persons: Effects on body composition and physical performance. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2005, 60, 1437–1446. [Google Scholar] [CrossRef] [Green Version]
- Cesari, M.; Pedone, C.; Incalzi, R.A.; Pahor, M. ACE-inhibition and physical function: Results from the Trial of Angiotensin-Converting Enzyme Inhibition and Novel Cardiovascular Risk Factors (TRAIN) study. J. Am. Med. Dir. Assoc. 2010, 11, 26–32. [Google Scholar] [CrossRef] [Green Version]
- Cabello-Verrugio, C.; Morales, M.G.; Rivera, J.C.; Cabrera, D.; Simon, F. Renin-angiotensin system: An old player with novel functions in skeletal muscle. Med. Res. Rev. 2015, 35, 437–463. [Google Scholar] [CrossRef]
- Gray, S.L.; LaCroix, A.Z.; Aragaki, A.K.; McDermott, M.; Cochrane, B.B.; Kooperberg, C.L.; Murray, A.M.; Rodriguez, B.; Black, H.; Woods, N.F. Angiotensin-converting enzyme inhibitor use and incident frailty in women aged 65 and older: Prospective findings from the Women’s Health Initiative Observational Study. J. Am. Geriatr. Soc. 2009, 57, 297–303. [Google Scholar] [CrossRef] [Green Version]
- Shrikrishna, D.; Tanner, R.J.; Lee, J.Y.; Natanek, A.; Lewis, A.; Murphy, P.B.; Hart, N.; Moxham, J.; Montgomery, H.E.; Kemp, P.R.; et al. A randomized controlled trial of angiotensin-converting enzyme inhibition for skeletal muscle dysfunction in COPD. Chest 2014, 146, 932–940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sumukadas, D.; Band, M.; Miller, S.; Cvoro, V.; Witham, M.; Struthers, A.; McConnachie, A.; Lloyd, S.M.; McMurdo, M. Do ACE inhibitors improve the response to exercise training in functionally impaired older adults? A randomized controlled trial. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2014, 69, 736–743. [Google Scholar] [CrossRef] [PubMed]
- Witham, M.D.; Syddall, H.E.; Dennison, E.; Cooper, C.; McMurdo, M.E.; Sayer, A.A. ACE inhibitors, statins and thiazides: No association with change in grip strength among community dwelling older men and women from the Hertfordshire Cohort Study. Age Ageing 2014, 43, 661–666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tinetti, M.E.; Han, L.; Lee, D.S.; McAvay, G.J.; Peduzzi, P.; Gross, C.P.; Zhou, B.; Lin, H. Antihypertensive medications and serious fall injuries in a nationally representative sample of older adults. JAMA Intern. Med. 2014, 174, 588–595. [Google Scholar] [CrossRef]
- George, C.; Verghese, J. Polypharmacy and Gait Performance in Community-dwelling Older Adults. J. Am. Geriatr. Soc. 2017, 65, 2082–2087. [Google Scholar] [CrossRef]
- Chung, C.M.; Wang, R.Y.; Chen, J.W.; Fann, C.S.; Leu, H.B.; Ho, H.Y.; Ting, C.T.; Lin, T.H.; Sheu, S.H.; Tsai, W.C.; et al. A genome-wide association study identifies new loci for ACE activity: Potential implications for response to ACE inhibitor. Pharmacogenom. J. 2010, 10, 537–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brugts, J.J.; Isaacs, A.; Boersma, E.; van Duijn, C.M.; Uitterlinden, A.G.; Remme, W.; Bertrand, M.; Ninomiya, T.; Ceconi, C.; Chalmers, J.; et al. Genetic determinants of treatment benefit of the angiotensin-converting enzyme-inhibitor perindopril in patients with stable coronary artery disease. Eur. Heart J. 2010, 31, 1854–1864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coates, D.; Isaac, R.E.; Cotton, J.; Siviter, R.; Williams, T.A.; Shirras, A.; Corvol, P.; Dive, V. Functional conservation of the active sites of human and Drosophila angiotensin I-converting enzyme. Biochemistry 2000, 39, 8963–8969. [Google Scholar] [CrossRef]
- Demontis, F.; Piccirillo, R.; Goldberg, A.L.; Perrimon, N. Mechanisms of skeletal muscle aging: Insights from Drosophila and mammalian models. Dis. Models Mech. 2013, 6, 1339–1352. [Google Scholar] [CrossRef] [Green Version]
- Akif, M.; Georgiadis, D.; Mahajan, A.; Dive, V.; Sturrock, E.D.; Isaac, R.E.; Acharya, K.R. High-resolution crystal structures of Drosophila melanogaster angiotensin-converting enzyme in complex with novel inhibitors and antihypertensive drugs. J. Mol. Biol. 2010, 400, 502–517. [Google Scholar] [CrossRef]
- Gabrawy, M.M.; Campbell, S.; Carbone, M.A.; Morozova, T.V.; Arya, G.H.; Turlapati, B.; Walston, J.D.; Starz-Gaiano, M.; Everett, L.; Mackay, T.F.C.; et al. Lisinopril preserves physical resilience and extends life span in a genotype-specific manner in Drosophila melanogaster. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2019, 74, 1844–1852. [Google Scholar] [CrossRef]
- Fried, L.P.; Tangen, C.M.; Walston, J.; Newman, A.B.; Hirsch, C.; Gottdiener, J.; Seeman, T.; Tracy, R.; Kop, W.J.; Burke, G.; et al. Frailty in older adults: Evidence for a phenotype. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2001, 56, M146–M156. [Google Scholar] [CrossRef]
- Mackay, T.F.; Richards, S.; Stone, E.A.; Barbadilla, A.; Ayroles, J.F.; Zhu, D.; Casillas, S.; Han, Y.; Magwire, M.M.; Cridland, J.M.; et al. The Drosophila melanogaster Genetic Reference Panel. Nature 2012, 482, 173–178. [Google Scholar] [CrossRef] [Green Version]
- Falconer, D.S. Selection in different environments: Effects on environmental sensitivity (reaction norm) and on mean performance. Genet. Res. 1990, 56, 57–70. [Google Scholar] [CrossRef] [Green Version]
- Mackay, T.F.C.; Huang, W. Charting the genotype-phenotype map: Lessons from the Drosophila melanogaster Genetic Reference Panel. Wiley Interdiscip. Rev. Dev. Biol. 2018, 7, e289. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Massouras, A.; Inoue, Y.; Peiffer, J.; Ramia, M.; Tarone, A.M.; Turlapati, L.; Zichner, T.; Zhu, D.; Lyman, R.F.; et al. Natural variation in genome architecture among 205 Drosophila melanogaster Genetic Reference Panel lines. Genome Res. 2014, 24, 1193–1208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vonesch, S.C.; Lamparter, D.; Mackay, T.F.; Bergmann, S.; Hafen, E. Genome-Wide Analysis Reveals Novel Regulators of Growth in Drosophila melanogaster. PLoS Genet. 2016, 12, e1005616. [Google Scholar] [CrossRef] [Green Version]
- Robertson, A. The sampling variance of the genetic correlation coefficient. Biometrics 1959, 15, 469–485. [Google Scholar] [CrossRef]
- Fochler, S.; Morozova, T.V.; Davis, M.R.; Gearhart, A.W.; Huang, W.; Mackay, T.F.C.; Anholt, R.R.H. Genetics of alcohol consumption in Drosophila melanogaster. Genes Brain Behav. 2017, 16, 675–685. [Google Scholar] [CrossRef] [Green Version]
- Antonov, A.V. BioProfiling: Analytical web portal for high-throughput cell biology. Nucleic Acids Res. 2011, 39, 323–327. [Google Scholar] [CrossRef] [Green Version]
- Seroude, L.; Brummel, T.; Kapahi, P.; Benzer, S. Spatio-temporal analysis of gene expression during aging in Drosophila melanogaster. Aging Cell 2002, 1, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Brack, A.S.; Conboy, M.J.; Roy, S.; Lee, M.; Kuo, C.J.; Keller, C.; Rando, T.A. Increased Wnt signaling during aging alters muscle stem cell fate and increases fibrosis. Science 2007, 317, 807–810. [Google Scholar] [CrossRef]
- Eliazer, S.; Brack, A.S. Stem cells: Cause and consequence in aged-muscle decline. Nature 2016, 540, 349–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marquilly, C.; Busto, G.U.; Leger, B.S.; Boulanger, A.; Giniger, E.; Walker, J.A.; Fradkin, L.G.; Dura, J.M. Htt is a repressor of Abl activity required for APP-induced axonal growth. PLoS Genet. 2021, 17, e1009287. [Google Scholar] [CrossRef] [PubMed]
- Pocas, G.M.; Branco-Santos, J.; Herrera, F.; Outeiro, T.F.; Domingos, P.M. α-Synuclein modifies mutant huntingtin aggregation and neurotoxicity in Drosophila. Hum. Mol. Genet. 2015, 24, 1898–1907. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Feany, M.B.; Saraswati, S.; Littleton, J.T.; Perrimon, N. Inactivation of Drosophila Huntingtin affects long-term adult functioning and the pathogenesis of a Huntington’s disease model. Dis. Model. Mech. 2009, 2, 247–266. [Google Scholar] [CrossRef] [Green Version]
- Lovato, T.L.; Nguyen, T.P.; Molina, M.R.; Cripps, R.M. The HOX gene abdominal-A specifies heart cell fate in the Drosophila dorsal vessel. Development 2002, 129, 5019–5027. [Google Scholar] [CrossRef]
- Enriquez, J.; Boukhatmi, H.; Dubois, L.; Philippakis, A.A.; Bulyk, M.L.; Michelson, A.M.; Crozatier, M.; Vincent, A. Multi-step control of muscle diversity by HOX proteins in the Drosophila embryo. Development 2010, 137, 457–466. [Google Scholar] [CrossRef] [Green Version]
- Bhalerao, S.; Berdnik, D.; Torok, T.; Knoblich, J.A. Localization-dependent and -independent roles of numb contribute to cell-fate specification in Drosophila. Curr. Biol. 2005, 15, 1583–1590. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Kuromi, H.; Briggs, L.; Green, D.B.; Rocha, J.J.; Sweeney, S.T.; Bullock, S.L. Bicaudal-D binds clathrin heavy chain to promote its transport and augments synaptic vesicle recycling. EMBO J. 2010, 29, 992–1006. [Google Scholar] [CrossRef] [Green Version]
- Baird, D.H.; Koto, M.; Wyman, R.J. Dendritic reduction in Passover, a Drosophila mutant with a defective giant fiber neuronal pathway. J. Neurobiol. 1993, 24, 971–984. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, S.N.; Frei, E.; Swain, G.P.; Wyman, R.J. Passover: A gene required for synaptic connectivity in the giant fiber system of Drosophila. Cell 1993, 73, 967–977. [Google Scholar] [CrossRef]
- Trimarchi, J.R.; Murphey, R.K. The shaking-B2 mutation disrupts electrical synapses in a flight circuit in adult Drosophila. J. Neurosci. 1997, 17, 4700–4710. [Google Scholar] [CrossRef] [Green Version]
- Gilsohn, E.; Volk, T. Slowdown promotes muscle integrity by modulating integrin-mediated adhesion at the myotendinous junction. Development 2010, 137, 785–794. [Google Scholar] [CrossRef] [Green Version]
- Jordan, K.W.; Craver, K.L.; Magwire, M.M.; Cubilla, C.E.; Mackay, T.F.; Anholt, R.R. Genome-wide association for sensitivity to chronic oxidative stress in Drosophila melanogaster. PLoS ONE 2012, 7, e38722. [Google Scholar] [CrossRef]
- Packard, M.; Koo, E.S.; Gorczyca, M.; Sharpe, J.; Cumberledge, S.; Budnik, V. The Drosophila Wnt, wingless, provides an essential signal for pre- and postsynaptic differentiation. Cell 2002, 111, 319–330. [Google Scholar] [CrossRef] [Green Version]
- Von Maltzahn, J.; Chang, N.C.; Bentzinger, C.F.; Rudnicki, M.A. Wnt signaling in myogenesis. Trends Cell Biol. 2012, 22, 602–609. [Google Scholar] [CrossRef] [Green Version]
- Rosso, S.B.; Inestrosa, N.C. WNT signaling in neuronal maturation and synaptogenesis. Front. Cell Neurosci. 2013, 7, 103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Chen, C.; Wu, C.; Yang, Y.; Li, W.; Xue, L. The canonical Wg signaling modulates Bsk-mediated cell death in Drosophila. Cell Death Dis. 2015, 6, 1713. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Page-McCaw, A. A matrix metalloproteinase mediates long-distance attenuation of stem cell proliferation. J. Cell Biol. 2014, 206, 923–936. [Google Scholar] [CrossRef] [Green Version]
- Merino, C.; Penney, J.; González, M.; Tsurudome, K.; Moujahidine, M.; O’Connor, M.B.; Verheyen, E.M.; Haghighi, P. Nemo kinase interacts with Mad to coordinate synaptic growth at the Drosophila neuromuscular junction. J. Cell Biol. 2009, 185, 713–725. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.H.; Imai, Y.; Shiseki, M.; Tanaka, J.; Motoji, T. Knockdown of the Wnt receptor Frizzled-1 (FZD1) reduces MDR1/P-glycoprotein expression in multidrug resistant leukemic cells and inhibits leukemic cell proliferation. Leuk Res. 2018, 67, 99–108. [Google Scholar] [CrossRef]
- Thorpe, C.J.; Moon, R.T. Nemo-like kinase is an essential co-activator of Wnt signaling during early zebrafish development. Development 2004, 131, 2899–2909. [Google Scholar] [CrossRef] [Green Version]
- Ishitani, T.; Hirao, T.; Suzuki, M.; Isoda, M.; Ishitani, S.; Harigaya, K.; Kitagawa, M.; Matsumoto, K.; Itoh, M. Nemo-like kinase suppresses Notch signalling by interfering with formation of the Notch active transcriptional complex. Nat. Cell Biol. 2010, 12, 278–285. [Google Scholar] [CrossRef]
- Yabumoto, C.; Akazawa, H.; Yamamoto, R.; Yano, M.; Kudo-Sakamoto, Y.; Sumida, T.; Kamo, T.; Yagi, H.; Shimizu, Y.; Saga-Kamo, A.; et al. Angiotensin II receptor blockade promotes repair of skeletal muscle through down-regulation of aging-promoting C1q expression. Sci. Rep. 2015, 5, 14453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, W.; Campbell, T.; Carbone, M.A.; Jones, W.E.; Unselt, D.; Anholt, R.R.H.; Mackay, T.F.C. Context-dependent genetic architecture of Drosophila life span. PLoS Biol. 2020, 18, e3000645. [Google Scholar] [CrossRef] [PubMed]
- Yanagawa, A.; Huang, W.; Yamamoto, A.; Wada-Katsumata, A.; Schal, C.; Mackay, T.F.C. Genetic basis of natural variation in spontaneous grooming in Drosophila melanogaster. G3 Genes Genomes Genet. 2020, 10, 3453–3460. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Gene Name and Stock | Stock Number | FlyBase Genotype | Human Ortholog |
|---|---|---|---|
| Axin stock 1 Axin primers | 31705 | y1 v1; P(TRiP.HM04012)attP2 F:AATGAGTGTAGTGGCCCACG R:TCTGCTACCCCTTCGGTCAT | AXIN1 |
| Axin stock 2 | 62434 | y1 v1; P(TRiP.HMJ23888)attP40/CyO | AXIN1 |
| frizzled stock 1 frizzled primers | 31036 | y1 v1; P(TRiP.JF01481)attP2 F:TCTGGGACCGAACTAGATGGA R:CACGACCGGAGCAAACTGAT | FZD1 |
| frizzled stock 2 | 34321 | y1 sc * v1; P(TRiP.HMS01308)attP2 | FZD1 |
| nemo stock 1 nemo primers | 41586 | y1 v1; P(TRiP.GL00703)attP2 F:CTCCCTACTATCAACCGC R:GCTCCATAGCCGATAGGACGA | NLK |
| nemo stock 2 | 25793 | y1 v1; P(TRiP.JF01799)attP2 | NLK |
| wingless stock 1 | 31310 | y1 v1; P(TRiP.JF01257)attP2 F:CCAAGTCGAGGGCAAACAGAA R:TGGATCGCTGGGTCCATGTA | WNT1 |
| wingless stock 2 | 31249 | y1 v1; P(TRiP.JF01480)attP2 | WNT1 |
| GWA Phenotype | Age (Weeks) | Treatment | Number of Indels | Number of SNPs | Number of Genes |
|---|---|---|---|---|---|
| Speed | 1 | Control | 9 | 76 | 47 |
| Speed | 1 | Lisinopril | 8 | 127 | 67 |
| Speed | 5 | Control | 11 | 201 | 99 |
| Speed | 5 | Lisinopril | 1 | 51 | 29 |
| Endurance | 1 | Control | 2 | 27 | 9 |
| Endurance | 1 | Lisinopril | 2 | 26 | 9 |
| Endurance | 5 | Control | 23 | 215 | 72 |
| Endurance | 5 | Lisinopril | 5 | 39 | 14 |
| Sensitivity of Climbing Speed | 1 | Lisinopril | 2 | 23 | 16 |
| Sensitivity of Climbing Speed | 5 | Lisinopril | 6 | 68 | 27 |
| Sensitivity of Endurance | 1 | Lisinopril | 5 | 32 | 24 |
| Sensitivity of Endurance | 5 | Lisinopril | 1 | 40 | 26 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gabrawy, M.M.; Khosravian, N.; Morcos, G.S.; Morozova, T.V.; Jezek, M.; Walston, J.D.; Huang, W.; Abadir, P.M.; Leips, J. Genome-Wide Analysis in Drosophila Reveals the Genetic Basis of Variation in Age-Specific Physical Performance and Response to ACE Inhibition. Genes 2022, 13, 143. https://doi.org/10.3390/genes13010143
Gabrawy MM, Khosravian N, Morcos GS, Morozova TV, Jezek M, Walston JD, Huang W, Abadir PM, Leips J. Genome-Wide Analysis in Drosophila Reveals the Genetic Basis of Variation in Age-Specific Physical Performance and Response to ACE Inhibition. Genes. 2022; 13(1):143. https://doi.org/10.3390/genes13010143
Chicago/Turabian StyleGabrawy, Mariann M., Nick Khosravian, George S. Morcos, Tatiana V. Morozova, Meagan Jezek, Jeremy D. Walston, Wen Huang, Peter M. Abadir, and Jeff Leips. 2022. "Genome-Wide Analysis in Drosophila Reveals the Genetic Basis of Variation in Age-Specific Physical Performance and Response to ACE Inhibition" Genes 13, no. 1: 143. https://doi.org/10.3390/genes13010143