
Comprehensive miRNA and DNA Microarray Analyses Reveal the Response of Hepatic miR-203 and Its Target Gene to Protein Malnutrition in Rats

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Biochemical Tests
2.3. RNA Extraction
2.4. miRNA Microarray
2.5. DNA Microarray
2.6. Ingenuity Pathway Analysis (IPA)
2.7. miRNA Reverse Transcription Polymerase Chain Reaction (RT-PCR)
2.8. mRNA RT-PCR
2.9. Cell Culture
2.10. Luciferase Assay
2.11. miRNA Overexpression and Knockdown
2.12. Statistical Analyses
3. Results
3.1. LP Diet Affected Lipid Metabolism
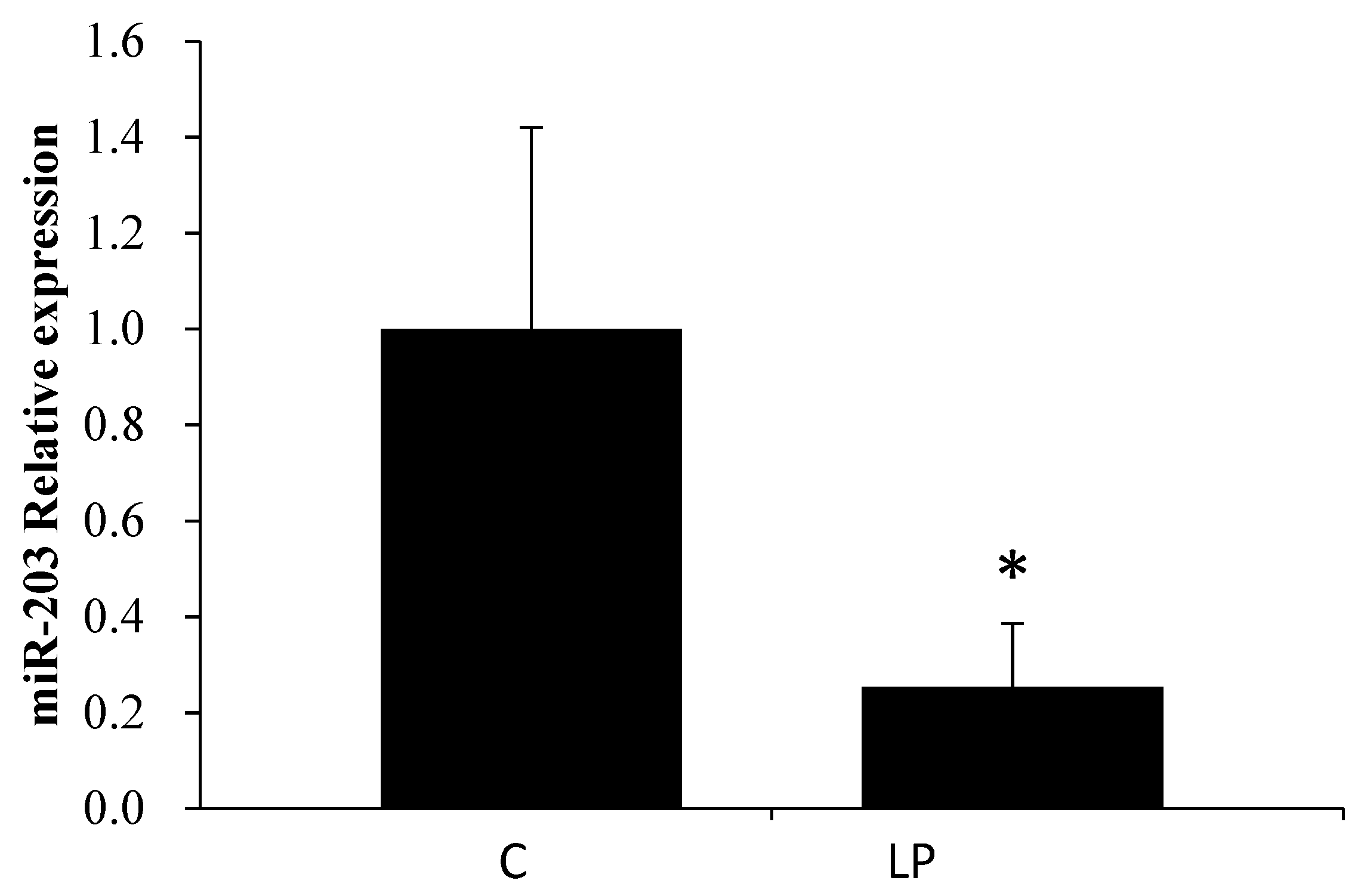
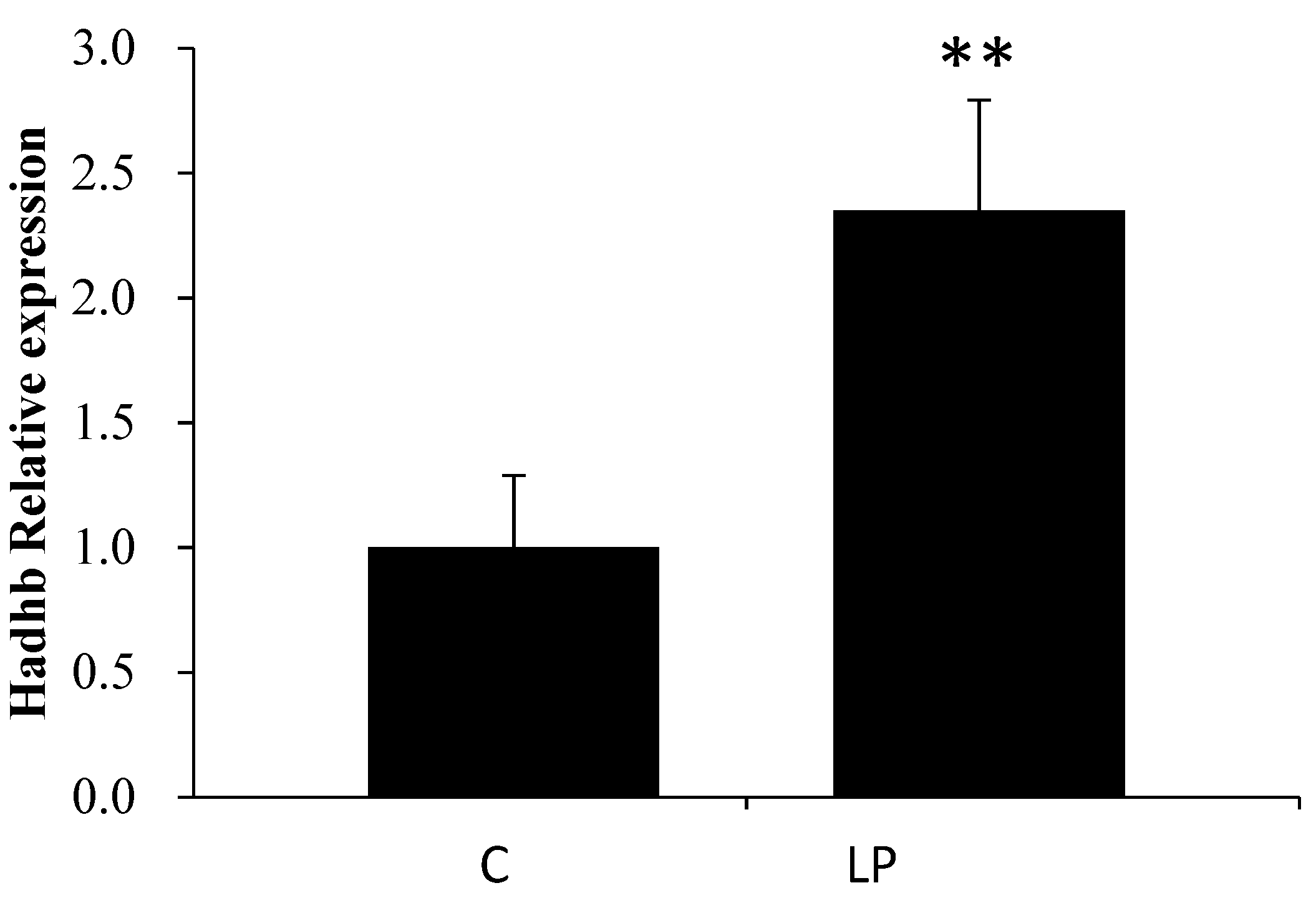
3.2. LP Diet Regulated miR-203 and Hadhb mRNA Expression in the Rat Liver
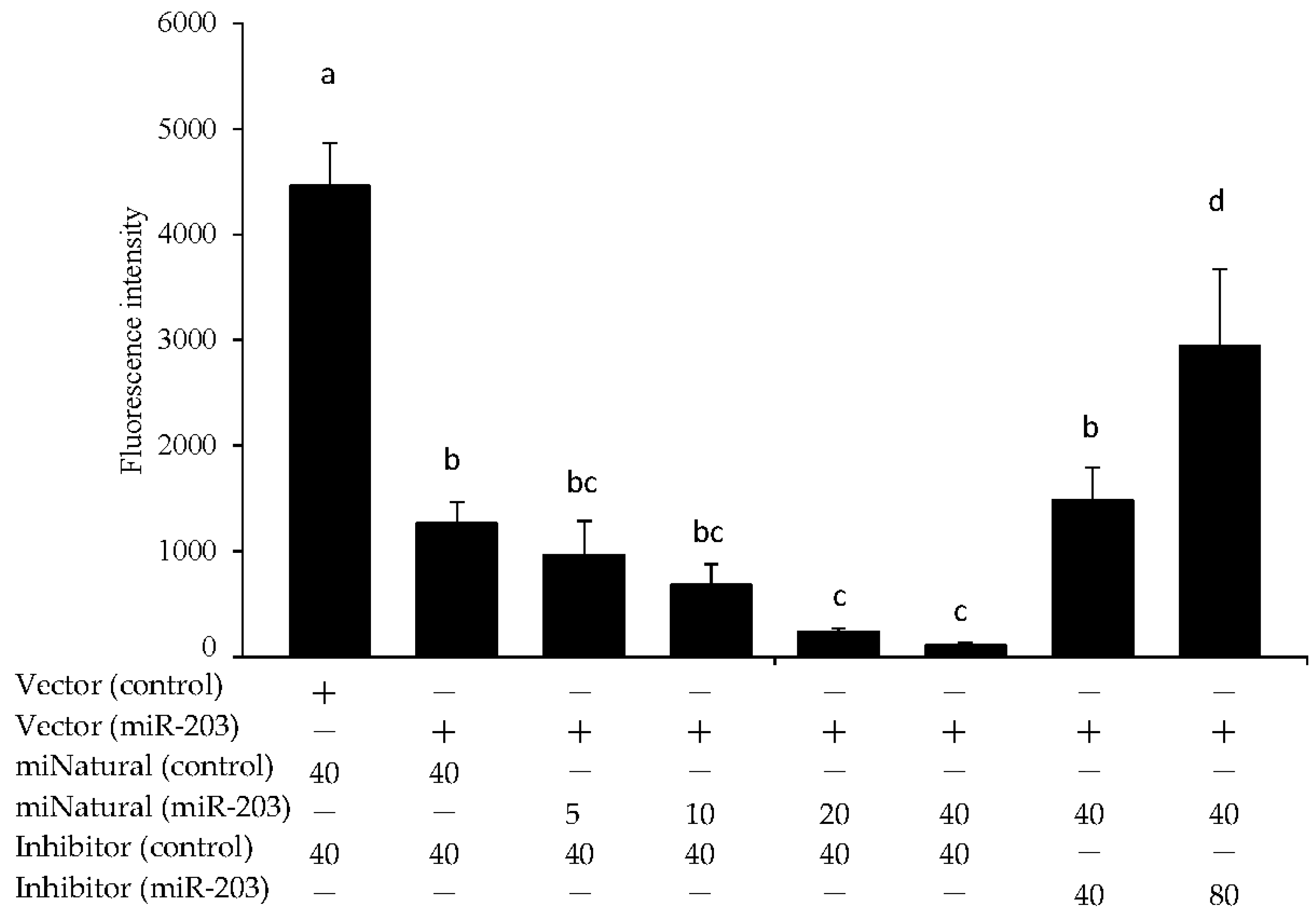
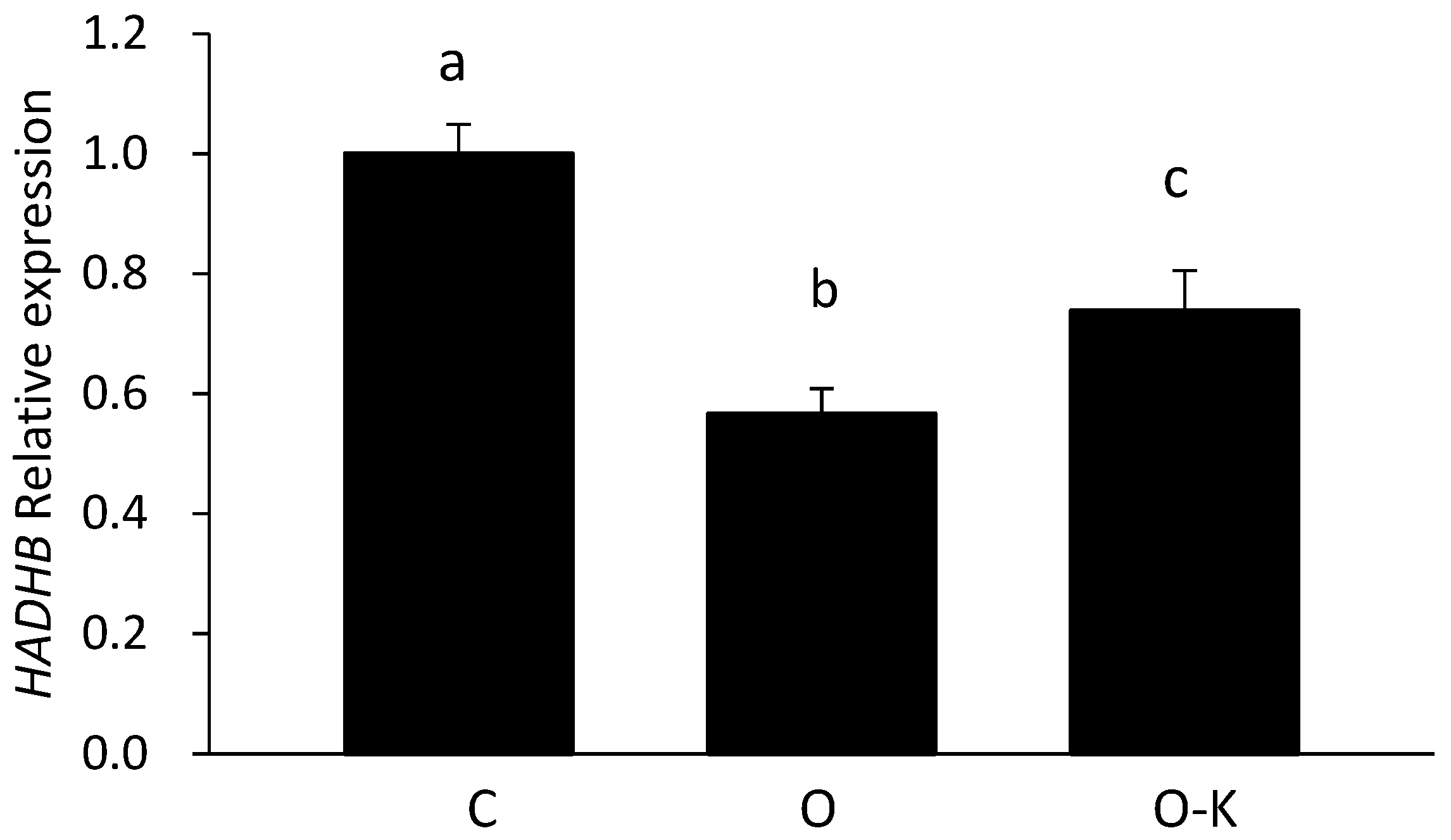
3.3. miR-203 Regulated the mRNA Expression of Hadhb
4. Discussion
Author Contributions
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Weigle, D.S.; Breen, P.A.; Matthys, C.C.; Callahan, H.S.; Meeuws, K.E.; Burden, V.R.; Purnell, J.Q. A high-protein diet induces sustained reductions in appetite, ad libitum caloric intake, and body weight despite compensatory changes in diurnal plasma leptin and ghrelin concentrations. Am. J. Clin. Nutr. 2005, 82, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Engelmann, D.T.; Sie, T.L.; Draper, H.H.; Bell, R.R. Effect of a high protein intake on calcium metabolism in the rat. J. Nutr. 1975, 105, 475–483. [Google Scholar] [CrossRef]
- Flores, H.; Sierralta, W.; Monckeberg, F. Triglyceride transport in protein-depleted rats. J. Nutr. 1970, 100, 375–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toyoshima, Y.; Tokita, R.; Ohne, Y.; Hakuno, F.; Noguchi, T.; Minami, S.; Kato, H.; Takahashi, S. Dietary protein deprivation upregulates insulin signaling and inhibits gluconeogenesis in rat liver. J. Mol. Endocrinol. 2010, 45, 329–340. [Google Scholar] [CrossRef]
- Takahashi, S.; Kajikawa, M.; Umezawa, T.; Takahashi, S.; Kato, H.; Miura, Y.; Nam, T.J.; Noguchi, T.; Naito, H. Effect of dietary proteins on the plasma immunoreactive insulin-like growth factor-1/somatomedin C concentration in the rat. Br. J. Nutr. 1990, 63, 521–534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takenaka, A.; Hirosawa, M.; Mori, M.; Yamada, S.; Miura, Y.; Kato, H.; Takahashi, S.; Noguchi, T. Effect of protein nutrition on the mRNA content of insulin-like growth factor-binding protein-1 in liver and kidney of rats. Br. J. Nutr. 1993, 69, 73–82. [Google Scholar] [CrossRef] [Green Version]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- Wightman, B.; Ha, I.; Ruvkun, G. Posttranscriptional regulation of the heterochronic gene lin-14 by lin-4 mediates temporal pattern formation in C. elegans. Cell 1993, 75, 855–862. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Lai, E.C. Micro RNAs are complementary to 3’ UTR sequence motifs that mediate negative post-transcriptional regulation. Nat. Genet. 2002, 30, 363–364. [Google Scholar] [CrossRef]
- Guo, H.; Ingolia, N.T.; Weissman, J.S.; Bartel, D.P. Mammalian microRNAs predominantly act to decrease target mRNA levels. Nature 2010, 466, 835–840. [Google Scholar] [CrossRef] [Green Version]
- Hatziapostolou, M.; Polytarchou, C.; Aggelidou, E.; Drakaki, A.; Poultsides, G.A.; Jaeger, S.A.; Ogata, H.; Karin, M.; Struhl, K.; Hadzopoulou-Cladaras, M.; et al. An HNF4alpha-miRNA inflammatory feedback circuit regulates hepatocellular oncogenesis. Cell 2011, 147, 1233–1247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hau, A.; Ceppi, P.; Peter, M.E. CD95 is part of a let-7/p53/miR-34 regulatory network. PLoS ONE 2012, 7, e49636. [Google Scholar] [CrossRef] [PubMed]
- Zong, M.; Feng, W.; Wan, L.; Yu, X.; Yu, W. miR-203 affects esophageal cancer cell proliferation, apoptosis and invasion by targeting MAP3K1. Oncol. Lett. 2020, 20, 751–757. [Google Scholar] [CrossRef]
- Editors, P.O. Retraction: MiR-203 Suppresses ZNF217 Upregulation in Colorectal Cancer and Its Oncogenicity. PLoS ONE 2020, 15, e0244268. [Google Scholar] [CrossRef]
- Chen, L.Z.; Ding, Z.; Zhang, Y.; He, S.T.; Wang, X.H. MiR-203 over-expression promotes prostate cancer cell apoptosis and reduces ADM resistance. Eur. Rev. Med. Pharm. Sci. 2020, 24, 2784. [Google Scholar] [CrossRef]
- Liu, H.P.; Zhang, Y.; Liu, Z.T.; Qi, H.; Zheng, X.M.; Qi, L.H.; Wang, J.Y. MiR-203 regulates proliferation and apoptosis of ovarian cancer cells by targeting SOCS3. Eur. Rev. Med. Pharm. Sci. 2020, 24, 7914. [Google Scholar] [CrossRef]
- Moore, K.J.; Rayner, K.J.; Suarez, Y.; Fernandez-Hernando, C. The role of microRNAs in cholesterol efflux and hepatic lipid metabolism. Annu. Rev. Nutr. 2011, 31, 49–63. [Google Scholar] [CrossRef] [Green Version]
- Wilfred, B.R.; Wang, W.X.; Nelson, P.T. Energizing miRNA research: A review of the role of miRNAs in lipid metabolism, with a prediction that miR-103/107 regulates human metabolic pathways. Mol. Genet. Metab. 2007, 91, 209–217. [Google Scholar] [CrossRef] [Green Version]
- Trajkovski, M.; Hausser, J.; Soutschek, J.; Bhat, B.; Akin, A.; Zavolan, M.; Heim, M.H.; Stoffel, M. MicroRNAs 103 and 107 regulate insulin sensitivity. Nature 2011, 474, 649–653. [Google Scholar] [CrossRef] [Green Version]
- Davalos, A.; Goedeke, L.; Smibert, P.; Ramirez, C.M.; Warrier, N.P.; Andreo, U.; Cirera-Salinas, D.; Rayner, K.; Suresh, U.; Pastor-Pareja, J.C.; et al. miR-33a/b contribute to the regulation of fatty acid metabolism and insulin signaling. Proc. Natl. Acad. Sci. USA 2011, 108, 9232–9237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wijesekara, N.; Zhang, L.H.; Kang, M.H.; Abraham, T.; Bhattacharjee, A.; Warnock, G.L.; Verchere, C.B.; Hayden, M.R. miR-33a modulates ABCA1 expression, cholesterol accumulation, and insulin secretion in pancreatic islets. Diabetes 2012, 61, 653–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinelli, R.; Nardelli, C.; Pilone, V.; Buonomo, T.; Liguori, R.; Castano, I.; Buono, P.; Masone, S.; Persico, G.; Forestieri, P.; et al. miR-519d overexpression is associated with human obesity. Obesity 2010, 18, 2170–2176. [Google Scholar] [CrossRef]
- Chen, J.W.; Cunningham, M.D.; Galton, N.; Michaelis, E.K. Immune labeling and purification of a 71-kDa glutamate-binding protein from brain synaptic membranes. Possible relationship of this protein to physiologic glutamate receptors. J. Biol. Chem. 1988, 263, 417–426. [Google Scholar] [CrossRef]
- Jia, Y.; Cong, R.; Li, R.; Yang, X.; Sun, Q.; Parvizi, N.; Zhao, R. Maternal low-protein diet induces gender-dependent changes in epigenetic regulation of the glucose-6-phosphatase gene in newborn piglet liver. J. Nutr. 2012, 142, 1659–1665. [Google Scholar] [CrossRef] [Green Version]
- Cong, R.; Jia, Y.; Li, R.; Ni, Y.; Yang, X.; Sun, Q.; Parvizi, N.; Zhao, R. Maternal low-protein diet causes epigenetic deregulation of HMGCR and CYP7alpha1 in the liver of weaning piglets. J. Nutr. Biochem. 2012, 23, 1647–1654. [Google Scholar] [CrossRef]
- Sun, M.; Estrov, Z.; Ji, Y.; Coombes, K.R.; Harris, D.H.; Kurzrock, R. Curcumin (diferuloylmethane) alters the expression profiles of microRNAs in human pancreatic cancer cells. Mol. Cancer Ther. 2008, 7, 464–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garzon, R.; Pichiorri, F.; Palumbo, T.; Visentini, M.; Aqeilan, R.; Cimmino, A.; Wang, H.; Sun, H.; Volinia, S.; Alder, H.; et al. MicroRNA gene expression during retinoic acid-induced differentiation of human acute promyelocytic leukemia. Oncogene 2007, 26, 4148–4157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Widdowson, E.M.; McCance, R.A. Effect of a low-protein diet on the chemical composition of the bodies and tissues of young rats. Br. J. Nutr. 1957, 11, 198–206. [Google Scholar] [CrossRef]
- Endo, Y.; Fu, Z.; Abe, K.; Arai, S.; Kato, H. Dietary protein quantity and quality affect rat hepatic gene expression. J. Nutr. 2002, 132, 3632–3637. [Google Scholar] [CrossRef]
- Joyal, J.S.; Sun, Y.; Gantner, M.L.; Shao, Z.; Evans, L.P.; Saba, N.; Fredrick, T.; Burnim, S.; Kim, J.S.; Patel, G.; et al. Retinal lipid and glucose metabolism dictates angiogenesis through the lipid sensor Ffar1. Nat. Med. 2016, 22, 439–445. [Google Scholar] [CrossRef] [Green Version]
- Fould, B.; Garlatti, V.; Neumann, E.; Fenel, D.; Gaboriaud, C.; Arlaud, G.J. Structural and functional characterization of the recombinant human mitochondrial trifunctional protein. Biochemistry 2010, 49, 8608–8617. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, S.; Mori, T.; Otsuka, Y.; Yamada, N.; Yasui, Y.; Iwasaki, J.; Kumazaki, M.; Maruo, K.; Akao, Y. Anti-oncogenic microRNA-203 induces senescence by targeting E2F3 protein in human melanoma cells. J. Biol. Chem. 2012, 287, 11769–11777. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Y.; Zeng, Z.Y.; Liu, X.H.; Gong, D.J.; Tao, J.; Cheng, H.Z.; Huang, S.D. MicroRNA-203 inhibits cell proliferation by repressing DeltaNp63 expression in human esophageal squamous cell carcinoma. BMC Cancer 2011, 11, 57. [Google Scholar] [CrossRef] [PubMed]
- Chim, C.S.; Wong, K.Y.; Leung, C.Y.; Chung, L.P.; Hui, P.K.; Chan, S.Y.; Yu, L. Epigenetic inactivation of the hsa-miR-203 in haematological malignancies. J. Cell. Mol. Med. 2011, 15, 2760–2767. [Google Scholar] [CrossRef] [Green Version]
- Viticchie, G.; Lena, A.M.; Latina, A.; Formosa, A.; Gregersen, L.H.; Lund, A.H.; Bernardini, S.; Mauriello, A.; Miano, R.; Spagnoli, L.G.; et al. MiR-203 controls proliferation, migration and invasive potential of prostate cancer cell lines. Cell Cycle 2011, 10, 1121–1131. [Google Scholar] [CrossRef] [Green Version]
- Ikenaga, N.; Ohuchida, K.; Mizumoto, K.; Yu, J.; Kayashima, T.; Sakai, H.; Fujita, H.; Nakata, K.; Tanaka, M. MicroRNA-203 expression as a new prognostic marker of pancreatic adenocarcinoma. Ann. Surg. Oncol. 2010, 17, 3120–3128. [Google Scholar] [CrossRef]
- Furuta, M.; Kozaki, K.I.; Tanaka, S.; Arii, S.; Imoto, I.; Inazawa, J. miR-124 and miR-203 are epigenetically silenced tumor-suppressive microRNAs in hepatocellular carcinoma. Carcinogenesis 2010, 31, 766–776. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
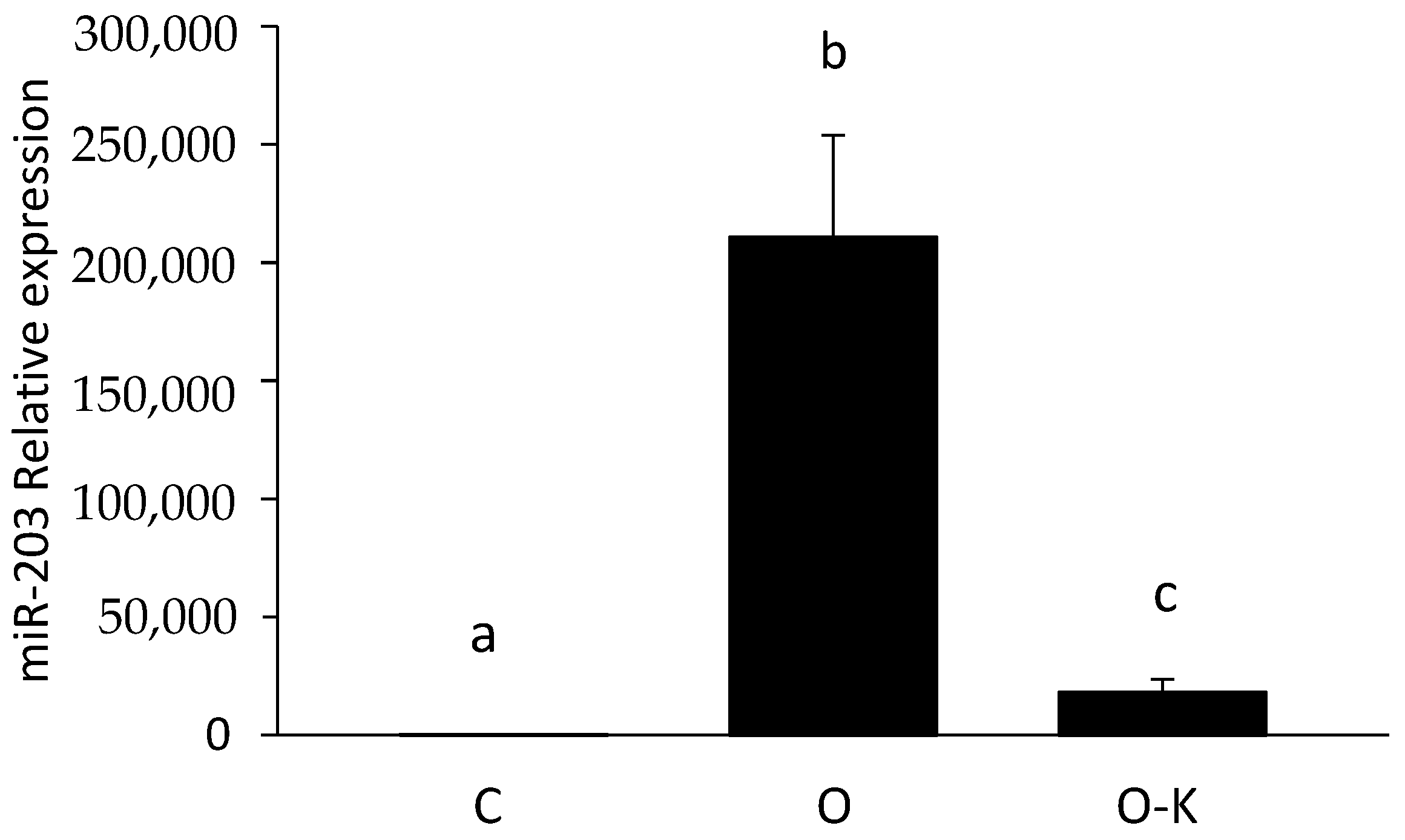{kind=link}
{kind=link}
| 20% Casein Diet (AIN-93G) | 5% Casein Diet | |
|---|---|---|
| Cornstarch (g) | 397.5 | 547.5 |
| Casein (g) | 200 | 50 |
| Dextrinized cornstarch (g) | 132 | 132 |
| Sucrose (g) | 102.5 | 102.5 |
| Soybean oil (g) | 70 | 70 |
| Fiber (g) | 50 | 50 |
| Mineral mix (g) | 35 | 35 |
| Vitamin mix (g) | 10 | 10 |
| L-cysteine (g) | 3 | 3 |
| Total (g) | 1000 | 1000 |
| C | LP | |
|---|---|---|
| Final body weight (g) | 341.3 ± 22.2 | 289.2 ± 12.9 ** |
| Liver weight (g/BW) | 4.3 ± 0.2 | 4.7 ± 0.4 |
| TG (mg/g tissue) | 11.3 ± 3.1 | 59.4 ± 13.0 ** |
| TC (mg/g tissue) | 4.5 ± 0.3 | 7.4 ± 1.1 ** |
| FFA (mEq/g tissue) | 21.4 ± 2.3 | 119.4 ± 52.7 ** |
| Plasma TG (mg/dL) | 206.0 ± 32.0 | 178.0 ± 66.2 |
| TC (mg/dL) | 63.6 ± 15.0 | 47.9 ± 6.4 |
| FFA (mEq/L) | 0.23 ± 0.02 | 0.14 ± 0.06 * |
| HDL-C (mg/dL) | 31.0 ± 6.3 | 21.6 ± 3.2 * |
| miRNA | miRNA-Log Ratio |
|---|---|
| upregulated | |
| rno-miR-200b | 1.1 |
| rno-miR-429 | 0.5 |
| downregulated | |
| rno-miR-203 | −1.1 |
| rno-miR-193 | −1.1 |
| rno-miR-182 | −0.9 |
| rno-miR-210 | −0.6 |
| miRNA | Log Ratio | Putative Target Genes | Log Ratio |
|---|---|---|---|
| rno-miR-200b | 1.1 | Fhl1 Runx1t1 Ccna2, Slc6a6 Mmd2 Kiaa0101, Lox Mpdz CIted2, Gpm6a, Mbnl3, Xkr8 Ptprd | −4.6 −3.5 −2.3 −1.9 −1.8 −1.3 −1.2 −1.0 |
| rno-miR-203 | −1.1 | Prickle2 Fkbp5 Hadhb Rpl23a | 4.0 1.3 1.2 1.0 |
| rno-miR-193 | −1.1 | Ccnd1 Agpat3, Atf5 | 1.3 1.0 |
| rno-miR-182 | −0.9 | Foxq1 Vldlr Trib3 Shc4 Glra3 Tsku Lpar4 Arhgef2, Camkk2, Fam118a, Otub2 Taf4b Npm1 Tmem86a | 4.9 4.2 3.6 3.2 3.0 1.8 1.7 1.6 1.5 1.1 1.0 |
| rno-miR-210 | −0.6 | Slc7a11 | 2.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Takahashi, K.; Jia, H.; Takahashi, S.; Kato, H. Comprehensive miRNA and DNA Microarray Analyses Reveal the Response of Hepatic miR-203 and Its Target Gene to Protein Malnutrition in Rats. Genes 2022, 13, 75. https://doi.org/10.3390/genes13010075
Takahashi K, Jia H, Takahashi S, Kato H. Comprehensive miRNA and DNA Microarray Analyses Reveal the Response of Hepatic miR-203 and Its Target Gene to Protein Malnutrition in Rats. Genes. 2022; 13(1):75. https://doi.org/10.3390/genes13010075
Chicago/Turabian StyleTakahashi, Kaoru, Huijuan Jia, Shoko Takahashi, and Hisanori Kato. 2022. "Comprehensive miRNA and DNA Microarray Analyses Reveal the Response of Hepatic miR-203 and Its Target Gene to Protein Malnutrition in Rats" Genes 13, no. 1: 75. https://doi.org/10.3390/genes13010075