
miRNA Profiling in the Chicken Liver under the Influence of Early Microbiota Stimulation with Probiotic, Prebiotic, and Synbiotic


Abstract
:1. Introduction
2. Materials and Methods
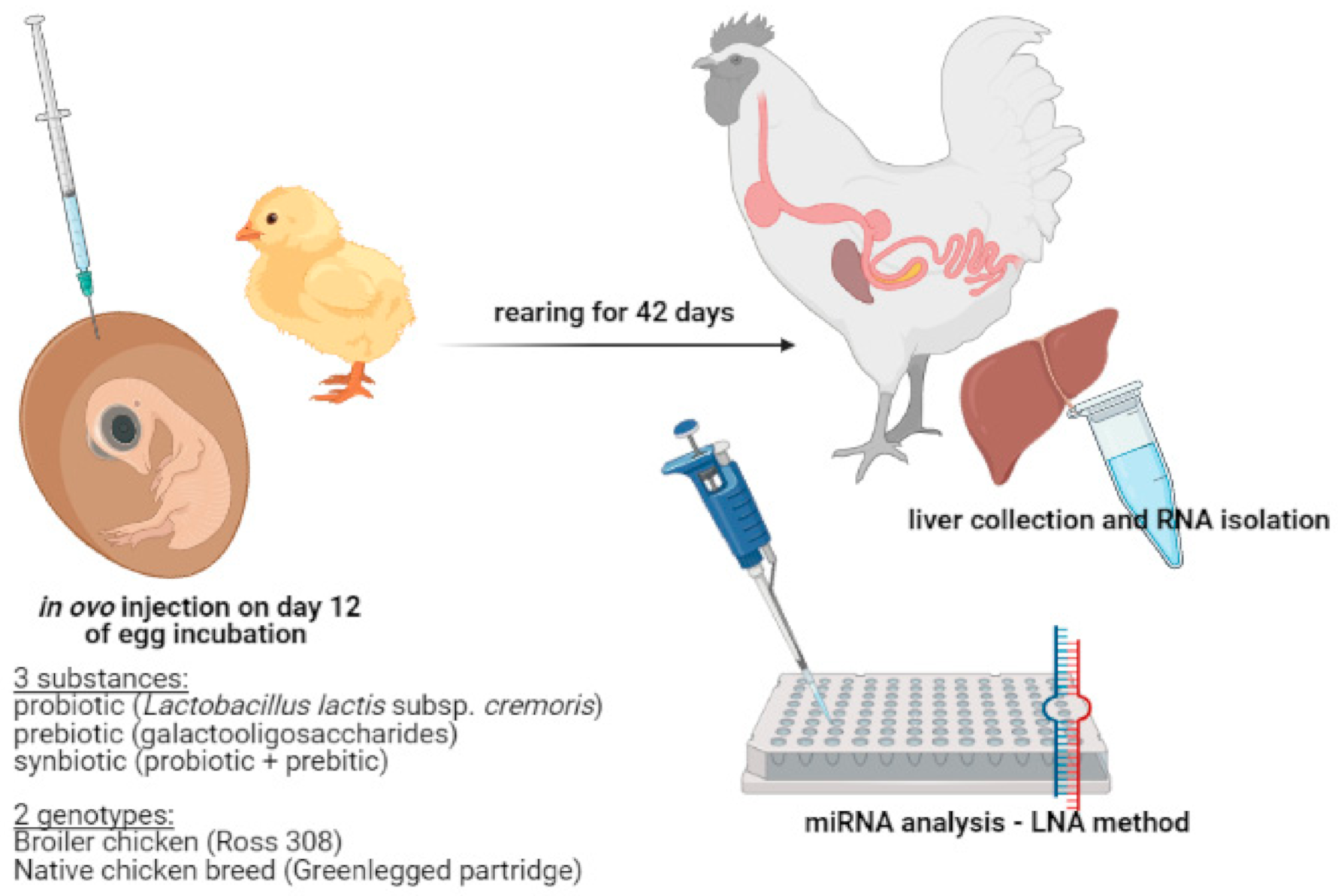
2.1. The Experimental Design
2.2. Isolation of RNA from Liver
2.3. miRNA Selection
2.4. LNA Method
2.5. Data Analysis
3. Results
miRNA Profiling
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marchix, J.; Goddard, G.; Helmrath, M.A. Host-Gut Microbiota Crosstalk in Intestinal Adaptation. Cell. Mol. Gastroenterol. Hepatol. 2018, 6, 149–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunislawska, A.; Slawinska, A.; Stadnicka, K.; Bednarczyk, M.; Gulewicz, P.; Jozefiak, D.; Siwek, M. Synbiotics for Broiler Chickens—In Vitro Design and Evaluation of the Influence on Host and Selected Microbiota Populations following In Ovo Delivery. PLoS ONE 2017, 12, e0168587. [Google Scholar]
- Dunislawska, A.; Slawinska, A.; Siwek, M. Hepatic DNA Methylation in Response to Early Stimulation of Microbiota with Lactobacillus Synbiotics in Broiler Chickens. Genes 2020, 11, 579. [Google Scholar] [CrossRef] [PubMed]
- Guerrero-Bosagna, C.; Morisson, M.; Liaubet, L.; Rodenburg, T.B.; de Haas, E.N.; Košťál, Ľ.; Pitel, F. Transgenerational epigenetic inheritance in birds. Environ. Epigenet. 2018, 4, dvy008. [Google Scholar] [CrossRef] [Green Version]
- Bednarczyk, M.; Dunislawska, A.; Stadnicka, K.; Grochowska, E. Chicken embryo as a model in epigenetic research. Poult. Sci. 2021, 101164. [Google Scholar] [CrossRef]
- Moore, L.D.; Le, T.; Fan, G. DNA methylation and its basic function. Neuropsychopharmacology 2013, 38, 23–38. [Google Scholar] [CrossRef] [Green Version]
- Taganov, K.D.; Boldin, M.P.; Baltimore, D. MicroRNAs and Immunity: Tiny Players in a Big Field. Immunity 2007, 26, 133–137. [Google Scholar] [CrossRef] [Green Version]
- Yao, Q.; Chen, Y.; Zhou, X. The roles of microRNAs in epigenetic regulation. Curr. Opin. Chem. Biol. 2019, 51, 11–17. [Google Scholar] [CrossRef]
- Chuang, J.C.; Jones, P.A. Epigenetics and MicroRNAs. Pediatr. Res. 2007, 61, 24R–29R. [Google Scholar] [CrossRef]
- Piletič, K.; Kunej, T. MicroRNA epigenetic signatures in human disease. Arch. Toxicol. 2016, 90, 2405–2419. [Google Scholar] [CrossRef]
- Ambros, V. The functions of animal microRNAs. Nature 2004, 431, 350–355. [Google Scholar] [CrossRef]
- Wienholds, E.; Kloosterman, W.P.; Miska, E.; Alvarez-Saavedra, E.; Berezikov, E.; De Bruijn, E.; Horvitz, H.R.; Kauppinen, S.; Plasterk, R.H.A. MicroRNA expression in zebrafish embryonic development. Science 2005, 309, 310–311. [Google Scholar] [CrossRef] [Green Version]
- Hicks, J.A.; Tembhurne, P.; Liu, H.C. MicroRNA expression in chicken embryos. Poult. Sci. 2008, 87, 2335–2343. [Google Scholar] [CrossRef]
- Siwek, M.; Slawinska, A.; Stadnicka, K.; Bogucka, J.; Dunislawska, A.; Bednarczyk, M. Prebiotics and synbiotics–in ovo delivery for improved lifespan condition in chicken. BMC Vet. Res. 2018, 14, 402. [Google Scholar] [CrossRef]
- Madej, J.P.; Skonieczna, J.; Siwek, M.; Kowalczyk, A.; Łukaszewicz, E.; Slawinska, A. Genotype-dependent development of cellular and humoral immunity in spleen and cecal tonsils of chickens stimulated in ovo with bioactive compounds. Poult. Sci. 2020, 99, 4343–4350. [Google Scholar] [CrossRef]
- Zehava, U.; Peter, R.F. Enhancement of Development of Iviparous Species by In Ovo Feeding. U.S. Patent US6592878B2, 15 July 2003. [Google Scholar]
- Dunislawska, A.; Slawinska, A.; Bednarczyk, M.; Siwek, M. Transcriptome modulation by in ovo delivered Lactobacillus synbiotics in a range of chicken tissues. Gene 2019, 698, 27–33. [Google Scholar] [CrossRef]
- Spinler, J.K.; Karri, V.; Hirschi, K.D. Planting the Microbiome. Trends Microbiol. 2019, 27, 90–93. [Google Scholar] [CrossRef]
- Slawinska, A.; Plowiec, A.; Siwek, M.; Jaroszewski, M.; Bednarczyk, M. Long-Term Transcriptomic Effects of Prebiotics and Synbiotics Delivered In Ovo in Broiler Chickens. PLoS ONE 2016, 11, e0168899. [Google Scholar] [CrossRef] [Green Version]
- Trefts, E.; Gannon, M.; Wasserman, D.H. The liver. Curr. Biol. 2017, 27, R1147–R1151. [Google Scholar] [CrossRef]
- Krenkel, O.; Tacke, F. Liver macrophages in tissue homeostasis and disease. Nat. Rev. Immunol. 2017, 17, 306–321. [Google Scholar] [CrossRef]
- Kubes, P.; Jenne, C. Immune Responses in the Liver. Annu. Rev. Immunol. 2018, 36, 247–277. [Google Scholar] [CrossRef]
- Stärkel, P.; Schnabl, B. Bidirectional Communication between Liver and Gut during Alcoholic Liver Disease. Semin. Liver Dis. 2016, 36, 331–339. [Google Scholar] [CrossRef]
- Tripathi, A.; Debelius, J.; Brenner, D.A.; Karin, M.; Loomba, R.; Schnabl, B.; Knight, R. The gut-liver axis and the intersection with the microbiome. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 397–411. [Google Scholar] [CrossRef]
- Siwek, M.; Wragg, D.; Sławińska, A.; Malek, M.; Hanotte, O.; Mwacharo, J.M. Insights into the genetic history of Green-legged Partridgelike fowl: MtDNA and genome-wide SNP analysis. Anim. Genet. 2013, 44, 522–532. [Google Scholar] [CrossRef] [Green Version]
- Lian, L.; Qu, L.; Chen, Y.; Lamont, S.J.; Yang, N. A Systematic Analysis of miRNA Transcriptome in Marek’s Disease Virus-Induced Lymphoma Reveals Novel and Differentially Expressed miRNAs. PLoS ONE 2012, 7, e51003. [Google Scholar]
- Glazov, E.A.; Cottee, P.A.; Barris, W.C.; Moore, R.J.; Dalrymple, B.P.; Tizard, M.L. A microRNA catalog of the developing chicken embryo identified by a deep sequencing approach. Genome Res. 2008, 18, 957–964. [Google Scholar] [CrossRef] [Green Version]
- Yu, S.; Wang, G.; Liao, J.; Tang, M.; Chen, J. Identifying and profiling the microRNAs associated with skin colour in the Muchuan black-bone chicken. Ital. J. Anim. Sci. 2020, 19, 468–476. [Google Scholar] [CrossRef]
- Parveen, A. Chasing the Genetics of Ascites in Broilers Using Whole Genome Resequencing. Ph.D. Thesis, University of Arkansas, Fayetteville, AR, USA, 2019. [Google Scholar]
- Lee, S.I.; Lee, B.R.; Hwang, Y.S.; Lee, H.C.; Rengaraj, D.; Song, G.; Park, T.S.; Han, J.Y. MicroRNA-mediated posttranscriptional regulation is required for maintaining undifferentiated properties of blastoderm and primordial germ cells in chickens. Proc. Natl. Acad. Sci. USA 2011, 108, 10426–10431. [Google Scholar] [CrossRef] [Green Version]
- Hong, Y.H.; Dinh, H.; Lillehoj, H.S.; Song, K.D.; Oh, J.D. Differential regulation of microRNA transcriptome in chicken lines resistant and susceptible to necrotic enteritis disease. Poult. Sci. 2014, 93, 1383–1395. [Google Scholar] [CrossRef]
- Liu, S.; Da Cunha, A.P.; Rezende, R.M.; Cialic, R.; Wei, Z.; Bry, L.; Comstock, L.E.; Gandhi, R.; Weiner, H.L. The Host Shapes the Gut Microbiota via Fecal MicroRNA. Cell Host Microbe 2016, 19, 32–43. [Google Scholar] [CrossRef] [Green Version]
- Dalmasso, G.; Nguyen, H.T.T.; Yan, Y.; Laroui, H.; Charania, M.A.; Ayyadurai, S.; Sitaraman, S.V.; Merlin, D. Microbiota Modulate Host Gene Expression via MicroRNAs. PLoS ONE 2011, 6, e19293. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-Y.; Jeong, W.; Lim, W.; Lim, C.-H.; Bae, S.-M.; Kim, J.; Bazer, F.W.; Song, G. Hypermethylation and Post-Transcriptional Regulation of DNA Methyltransferases in the Ovarian Carcinomas of the Laying Hen. PLoS ONE 2013, 8, e61658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kellermayer, R.; Dowd, S.E.; Harris, R.A.; Balasa, A.; Schaible, T.D.; Wolcott, R.D.; Tatevian, N.; Szigeti, R.; Li, Z.; Versalovic, J.; et al. Colonic mucosal DNA methylation, immune response, and microbiome patterns in Toll-like receptor 2-knockout mice. FASEB J. 2011, 25, 1449–1460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasan, N.; Yang, H. Factors affecting the composition of the gut microbiota, and its modulation. PeerJ 2019, 7, e7502. [Google Scholar] [CrossRef] [Green Version]
- Teng, Y.; Ren, Y.; Sayed, M.; Hu, X.; Lei, C.; Kumar, A.; Hutchins, E.; Mu, J.; Deng, Z.; Luo, C.; et al. Plant-Derived Exosomal MicroRNAs Shape the Gut Microbiota. Cell Host Microbe 2018, 24, 637–652.e8. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Nogales, A.; Algieri, F.; Garrido-Mesa, J.; Vezza, T.; Utrilla, M.P.; Chueca, N.; García, F.; Rodríguez-Cabezas, M.E.; Gálvez, J. Intestinal anti-inflammatory effect of the probiotic Saccharomyces boulardii in DSS-induced colitis in mice: Impact on microRNAs expression and gut microbiota composition. J. Nutr. Biochem. 2018, 61, 129–139. [Google Scholar] [CrossRef]
- Heydari, Z.; Rahaie, M.; Alizadeh, A.M.; Agah, S.; Khalighfard, S.; Bahmani, S. Effects of Lactobacillus acidophilus and Bifidobacterium bifidum Probiotics on the Expression of MicroRNAs 135b, 26b, 18a and 155, and Their Involving Genes in Mice Colon Cancer. Probiotics Antimicrob. Proteins 2019, 11, 1155–1162. [Google Scholar] [CrossRef]
- Giahi, L.; Aumueller, E.; Elmadfa, I.; Haslberger, A.G. Regulation of TLR4, p38 MAPkinase, IκB and mirnas by inactivated strains of lactobacilli in human dendritic cells. Benef. Microbes 2012, 3, 91–98. [Google Scholar] [CrossRef]
- Chen, Q.; Tong, C.; Ma, S.; Zhou, L.; Zhao, L.; Zhao, X. Involvement of microRNAs in probiotics-induced reduction of the cecal inflammation by Salmonella typhimurium. Front. Immunol. 2017, 8, 704. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| miRNA | Number of Regulated Genes |
|---|---|
| miR1598 | 464 |
| miR199B | 467 |
| miR1580 | 750 |
| miR1708 | 565 |
| miR1674 | 348 |
| miR1739 | 993 |
| miR211 | 241 |
| miR1807 | 1166 |
| miR1652 | 734 |
| miR1612 | 1876 |
| Name | Accession in mirBase | Catalog No. |
|---|---|---|
| gga-miR-1580 | MI0007306 | YP02116472 |
| gga-miR-1612 | MI0007340 | YP02104755 |
| hsa-miR-204-5p | MI0000284 | YP00206072 |
| gga-miR-1708 | MI0007444 | YP02101847 |
| gga-miR-1807 | MI0007552 | YP02116728 |
| gga-miR-1674 | MI0007408 | YP02110404 |
| gga-miR-1652 | MI0007384 | YP02118325 |
| gga-miR-1739 | MI0007478 | YP02102609 |
| gga-miR-1598 | MI0007325 | YP02105156 |
| gga-miR-199b | MI0007426 | YP02107667 |
| miR | Genotype | Substance | Genotype × Substance |
|---|---|---|---|
| miR-199b | 0.0001 | 0.0002 | 0.0038 |
| miR-1598 | Ns | 0.045 | 0.044 |
| miR-204-5p | 0.0001 | 0.004 | Ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sikorska, M.; Siwek, M.; Slawinska, A.; Dunislawska, A. miRNA Profiling in the Chicken Liver under the Influence of Early Microbiota Stimulation with Probiotic, Prebiotic, and Synbiotic. Genes 2021, 12, 685. https://doi.org/10.3390/genes12050685
Sikorska M, Siwek M, Slawinska A, Dunislawska A. miRNA Profiling in the Chicken Liver under the Influence of Early Microbiota Stimulation with Probiotic, Prebiotic, and Synbiotic. Genes. 2021; 12(5):685. https://doi.org/10.3390/genes12050685
Chicago/Turabian StyleSikorska, Michalina, Maria Siwek, Anna Slawinska, and Aleksandra Dunislawska. 2021. "miRNA Profiling in the Chicken Liver under the Influence of Early Microbiota Stimulation with Probiotic, Prebiotic, and Synbiotic" Genes 12, no. 5: 685. https://doi.org/10.3390/genes12050685