
Genes and SNPs Involved with Scrotal and Umbilical Hernia in Pigs


 , , , , ,
, , , , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Sample Collection
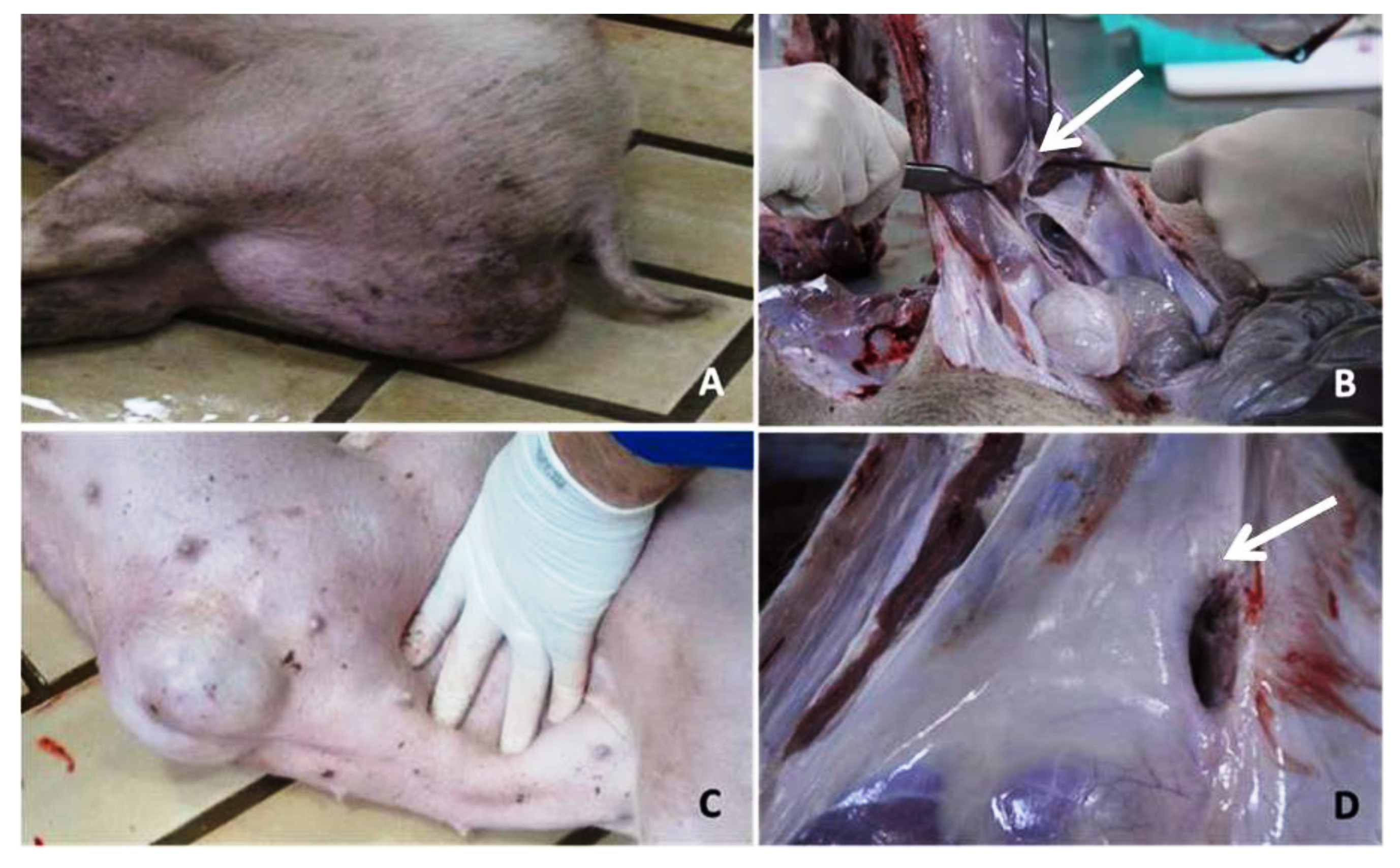
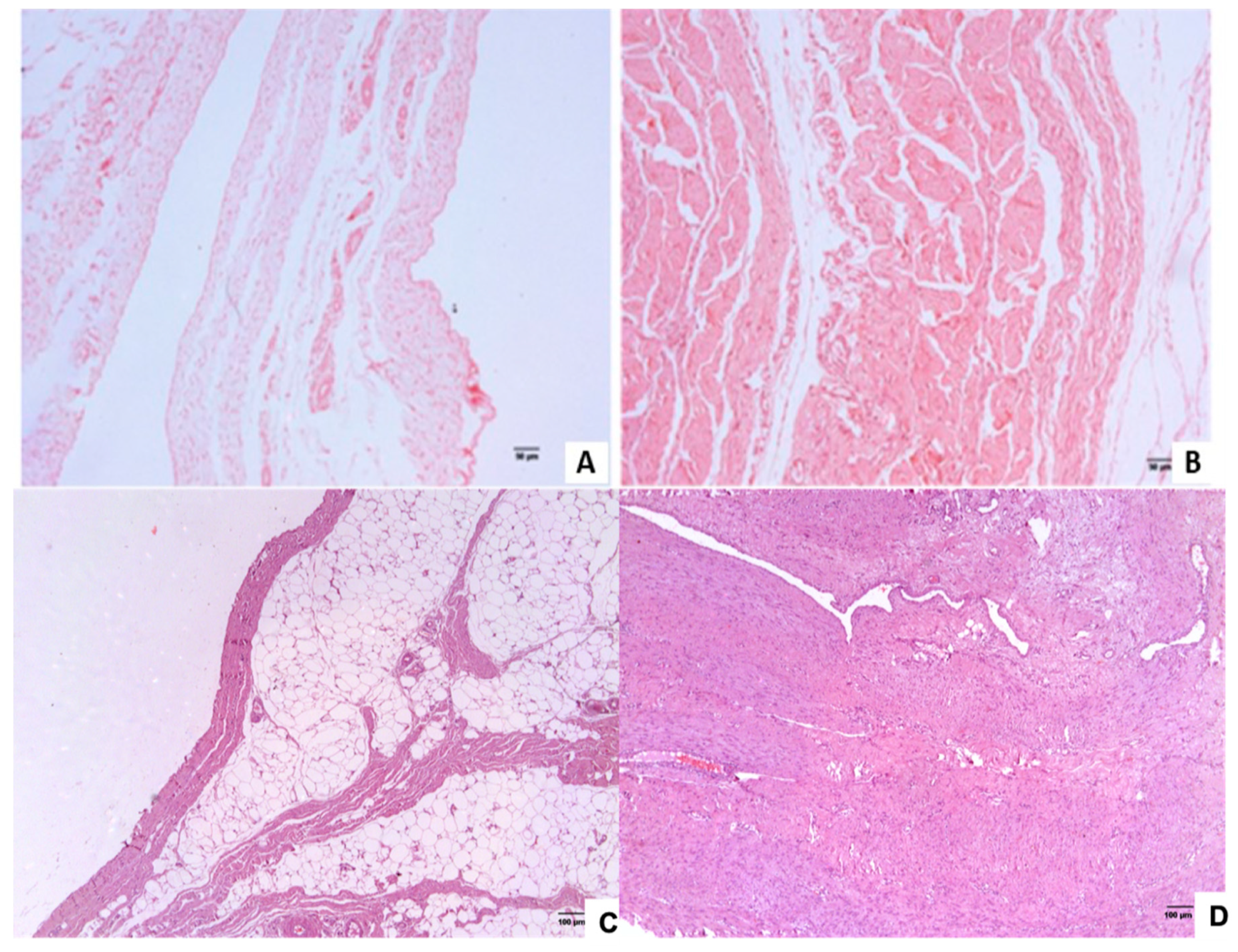
2.2. Histopathological Analysis of the Inguinal and Umbilical Ring Tissues
2.3. RNA Extraction, RNA-Seq Libraries Preparation and Sequencing
2.4. Quality Control and Differentially Expressed Genes
2.5. Transcriptomes Characterization of Scrotal and Umbilical Hernia
2.6. Comparison of Differentially Expressed Genes and In Silico Functional Analysis
2.7. Identification of Polymorphisms
3. Results
3.1. Histopathological Analysis of the Inguinal and Umbilical Ring Tissues
3.2. Sequencing and Mapping
3.3. Characterization of the Scrotal and Umbilical Hernia Transcriptomes
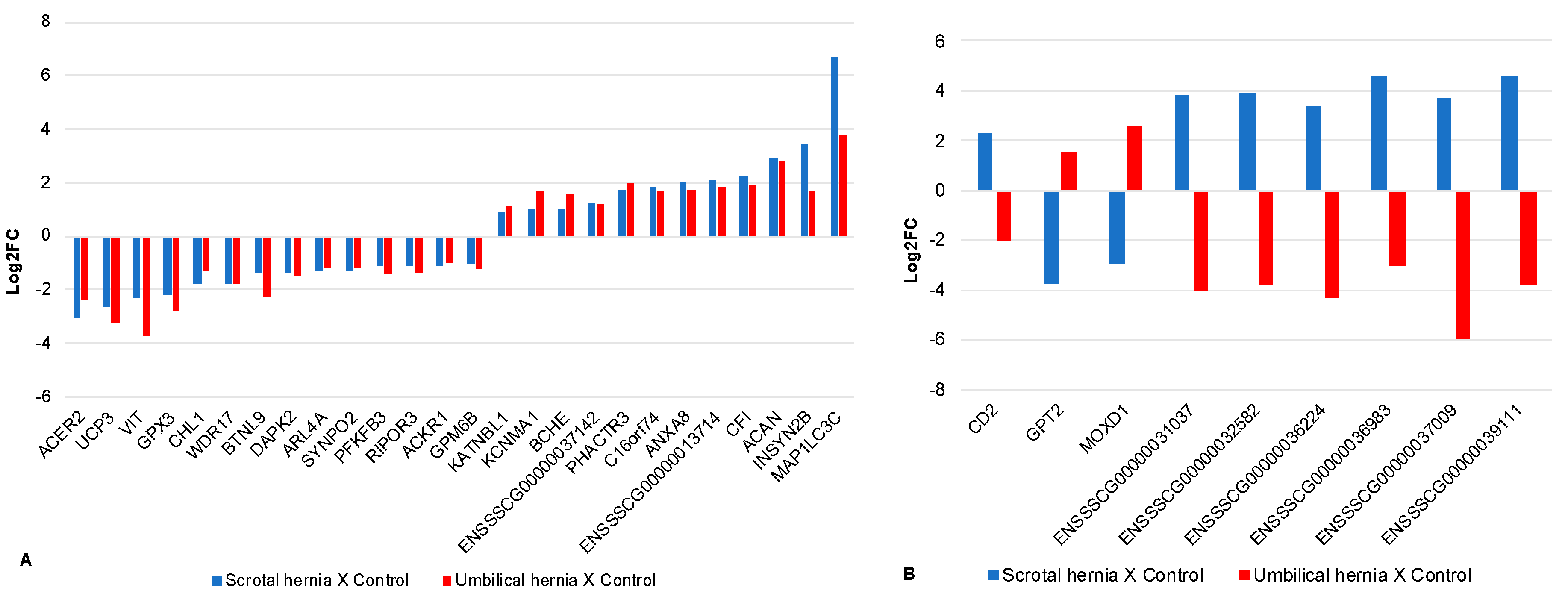
3.4. Differentially Expressed Genes
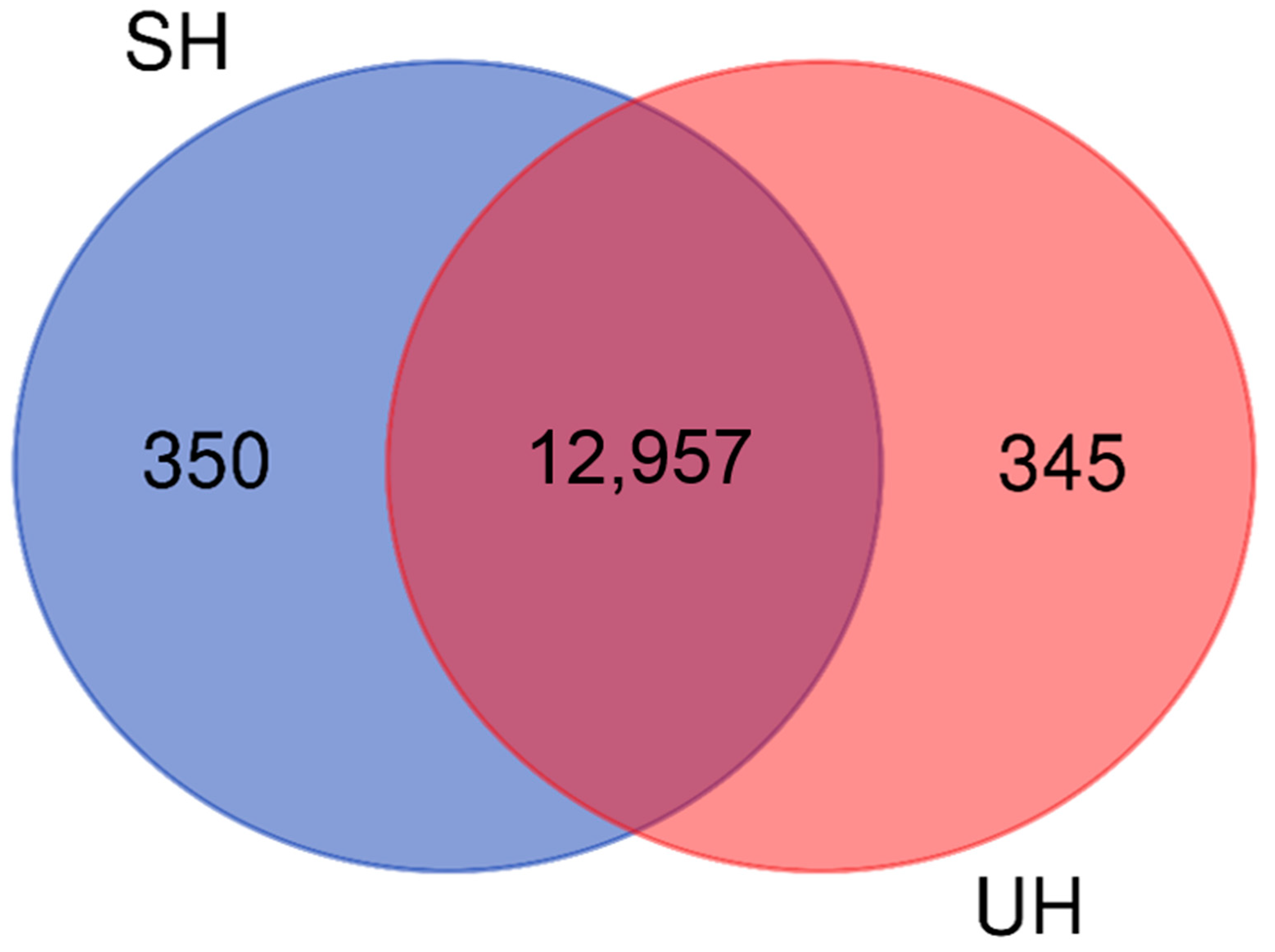
3.5. Differentially Expressed Genes Common to Both SH and UH Transcriptomes
3.6. Identification of Polymorphisms
4. Discussion
4.1. Transcriptome Characterization
4.2. Common Differentially Expressed Genes in Scrotal and Umbilical Hernias
4.3. Enriched Biological Processes
4.4. DE Genes Located in QTL Regions for Hernias and Polymorphisms Characterization
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BP | Biological process |
| DE | Differentially expressed |
| GATK | Genome analysis Tool Kit |
| GO | Gene ontology |
| HE | Hematoxylin and eosin |
| IH | Inguinal hernia |
| MDS | Multidimensional scaling |
| QTL | Quantitative trait loci |
| SH | Scrotal hernia |
| SIFT | Sorting intolerant from tolerant |
| SNP | Single nucleotide polymorphism |
| UH | Umbilical hernia |
| VEP | Variant effect predictor |
References
- Heck, A. Fatores que influenciam o desenvolvimento dos leitões na recria e terminação. Acta Sci. Vet. 2009, 37 (Suppl. 1), 211–218. [Google Scholar]
- Duarte, D.A.S.; Fortes, M.R.S.; Duarte, M.D.S.; Guimarães, S.E.F.; Verardo, L.L.; Veroneze, R.; Ribeiro, A.M.F.; Lopes, P.S.; De Resende, M.D.V.; E Silva, F.F. Genome-wide association studies, meta-analyses and derived gene network for meat quality and carcass traits in pigs. Anim. Prod. Sci. 2018, 58, 1100. [Google Scholar] [CrossRef]
- González-Prendes, R.; Quintanilla, R.; Cánovas, A.; Manunza, A.; Cardoso, T.F.; Jordana, J.; Noguera, J.L.; Pena, R.N.; Amills, M. Joint QTL mapping and gene expression analysis identify positional candidate genes influencing pork quality traits. Sci. Rep. 2017, 7, 39830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, R.; Quan, J.; Yang, M.; Wang, X.; Zheng, E.; Yang, H.; Fu, D.; Yang, Y.; Yang, L.; Li, Z.; et al. Genome-wide association analysis reveals genetic loci and candidate genes for feeding behavior and eating efficiency in Duroc boars. PLoS ONE 2017, 12, e0183244. [Google Scholar] [CrossRef] [Green Version]
- Lorenzetti, W.R. Análise da Expressão Gênica em Suínos Normais e Afetados com Hérnia Escrotal. Available online: http://sistemabu.udesc.br/pergamumweb/vinculos/000047/00004770.pdf (2018) (accessed on 29 January 2020).
- Romano, G.D.S.; Ibelli, A.M.G.; Lorenzetti, W.R.; Weber, T.; Peixoto, J.D.O.; Cantão, M.E.; Mores, M.A.Z.; Morés, N.; Pedrosa, V.B.; Cesar, A.S.M.; et al. Inguinal Ring RNA Sequencing Reveals Downregulation of Muscular Genes Related to Scrotal Hernia in Pigs. Genes 2020, 11, 117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Souza, M.R.; Ibelli, A.M.G.; Savoldi, I.R.; Cantão, M.E.; Peixoto, J.D.O.; Mores, M.A.Z.; Lopes, J.S.; Coutinho, L.L.; Ledur, M.C. Transcriptome analysis identifies genes involved with the development of umbilical hernias in pigs. PLoS ONE 2020, 15, e0232542. [Google Scholar] [CrossRef] [PubMed]
- Nietfeld, F.; Höltig, D.; Willems, H.; Valentin-Weigand, P.; Wurmser, C.; Waldmann, K.-H.; Fries, R.; Reiner, G. Candidate genes and gene markers for the resistance to porcine pleuropneumonia. Mamm. Genome 2020, 31, 54–67. [Google Scholar] [CrossRef] [Green Version]
- Yang, T.; Zhang, F.; Zhai, L.; He, W.; Tan, Z.; Sun, Y.; Wang, Y.; Liu, L.; Ning, C.; Zhou, W.; et al. Transcriptome of Porcine PBMCs over Two Generations Reveals Key Genes and Pathways Associated with Variable Antibody Responses post PRRSV Vaccination. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Mattsson, P. Prevalence of Congenital Defects in Swedish Hampshire, Landrace and Yorkshire Pig Breeds and Opinions on Their Prevalence in Swedish Commercial Herds. 2011. Available online: https://stud.epsilon.slu.se/2390/1/mattsson_p_110330.pdf (accessed on 29 January 2020).
- Grindflek, E.; Moe, M.; Taubert, H.; Simianer, H.; Lien, S.; Moen, T. Genome-wide linkage analysis of inguinal hernia in pigs using affected sib pairs. BMC Genet. 2006, 7, 25. [Google Scholar] [CrossRef] [Green Version]
- Miller, P.A.; Mezwa, D.G.; Feczko, P.J.; Jafri, Z.H.; Madrazo, B.L. Imaging of abdominal hernias. RadioGraphics 1995. [Google Scholar] [CrossRef] [Green Version]
- Straw, B.; Bates, R.; May, G. Anatomical abnormalities in a group of finishing pigs: Prevalence and pig performance. J. Swine Health Prod. 2009, 17, 28–31. [Google Scholar]
- Clarnette, T.D.; Lam, S.K.; Hutson, J.M. Ventriculo-peritoneal shunts in children reveal the natural history of closure of the processus vaginalis. J. Pediatr. Surg. 1998, 33, 413–416. [Google Scholar] [CrossRef]
- Amato, G.; Agrusa, A.; Romano, G.; Salamone, G.; Gulotta, G.; Silvestri, F.; Bussani, R. Muscle degeneration in inguinal hernia specimens. Hernia 2011, 16, 327–331. [Google Scholar] [CrossRef]
- Clarnette, T.D.; Hutson, J.M. Is the ascending testis actually? stationary?? Normal elongation of the spermatic cord is prevented by a fibrous remnant of the processus vaginalis. Pediatr. Surg. Int. 1997, 12, 155–157. [Google Scholar] [CrossRef] [PubMed]
- Beuermann, C.; Beck, J.; Schmelz, U.; Dunkelberg, H.; Schütz, E.; Brenig, B.; Knorr, C. Tissue Calcium Content in Piglets with Inguinal or Scrotal Hernias or Cryptorchidism. J. Comp. Pathol. 2009, 140, 182–186. [Google Scholar] [CrossRef] [PubMed]
- Bendavid, R. The Unified Theory of hernia formation. Hernia 2004, 8, 171–176. [Google Scholar] [CrossRef]
- Franz, M.G. The Biology of Hernia Formation. Surg. Clin. N. Am. 2008, 88, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Sevillano, C.A.; Lopes, M.S.; Harlizius, B.; Hanenberg, E.H.A.T.; Knol, E.F.; Bastiaansen, J.W.M. Genome-wide association study using deregressed breeding values for cryptorchidism and scrotal/inguinal hernia in two pig lines. Genet. Sel. Evol. 2015, 47, 18. [Google Scholar] [CrossRef] [Green Version]
- Petersen, H.H.; Nielsen, E.O.; Hassing, A.-G.; Ersboll, A.K.; Nielsen, J.P. Prevalence of clinical signs of disease in Danish finisher pigs. Veter Rec. 2008, 162, 377–382. [Google Scholar] [CrossRef]
- Warren, T.R.; Atkeson, F.W. Inheritance of Hernia. J. Hered. 1931, 22, 345–352. [Google Scholar] [CrossRef]
- Searcy-Bernal, R.; Gardner, I.A.; Hird, D.W. Effects of and factors associated with umbilical hernias in a swine herd. J. Am. Vet. Med. Assoc. 1994, 204, 1660–1664. [Google Scholar] [PubMed]
- Young, G.B.; Angus, K. A note on the genetics of umbilical hernia. Veter Rec. 1972, 90, 245–247. [Google Scholar] [CrossRef] [PubMed]
- Grindflek, E.; Hansen, M.H.S.; Lien, S.; Van Son, M. Genome-wide association study reveals a QTL and strong candidate genes for umbilical hernia in pigs on SSC14. BMC Genom. 2018, 19, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Sobestiansky, J.; Carvalho, L.F.O.S. Malformações. In Doenças dos Suínos; Sobestiansky, J., Barcellos, D.J., Eds.; Cânone Editorial: Goiânia, Brazil, 2007; pp. 527–538. [Google Scholar]
- Sutradhar, B.C.; Hossain, M.F.; Das, B.C.; Kim, G.; Hossain, M.A. Comparison between open and closed methods of herniorrhaphy in calves affected with umbilical hernia. J. Vet. Sci. 2009, 10, 343–347. [Google Scholar] [CrossRef] [Green Version]
- Thaller, G.; Dempfle, L.; Hoeschele, I. Maximum likelihood analysis of rare binary traits under different modes of inheritance. Genetics 1996, 143, 1819–1829. [Google Scholar] [CrossRef] [PubMed]
- Ding, N.S.; Mao, H.R.; Guo, Y.M.; Ren, J.; Xiao, S.J.; Wu, G.Z.; Shen, H.Q.; Wu, L.H.; Ruan, G.F.; Brenig, B.; et al. A genome-wide scan reveals candidate susceptibility loci for pig hernias in an intercross between White Duroc and Erhualian. J. Anim. Sci. 2009, 87, 2469–2474. [Google Scholar] [CrossRef] [PubMed]
- Pig QTL Database. Available online: https://www.animalgenome.org/cgi-bin/QTLdb/SS/index (accessed on 4 February 2020).
- Li, X.; Xu, P.; Zhang, C.; Sun, C.; Han, X.; Li, M.; Qiao, R. Genome-wide association study identifies variants in the CAPN9 gene associated with umbilical hernia in pigs. Anim. Genet. 2019, 50, 162–165. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, L.T.; Ono, R.K.; Ibelli, A.M.G.; Lagos, E.B.; Morés, M.A.Z.; Cantão, M.E.; Lorenzetti, W.R.; Peixoto, J.D.O.; Pedrosa, V.B.; Ledur, M.C. Novel putative candidate genes associated with umbilical hernia in pigs. In 11th World Congress on Genetics Applied to Livestock Production; WCGALP, Massey University: Auckland, New Zealand, 2018; Available online: https://www.alice.cnptia.embrapa.br/alice/bitstream/doc/1093239/1/final8696.pdf (accessed on 29 January 2020).
- Github Repository. Available online: https://github.com/hanielcedraz/BAQCOM (accessed on 6 November 2019).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Ensembl. Available online: http://www.ensembl.org/index.html (accessed on 4 February 2020).
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-Seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Team, RStudio. RStudio: Integrated Development for R; RStudio, Inc.: Boston, MA, USA; Available online: http://www. rstudio.com/ (accessed on 16 March 2020).
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. Available online: https://www.R-project.org/ (accessed on 25 April 2020).
- Robinson, M.D.; Mccarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. Royal Stat. Soc. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- DAVID Bioinformatics Resources 6.8. Available online: https://david.ncifcrf.gov/tools.jsp (accessed on 4 February 2020).
- UniProt. Available online: https://www.uniprot.org/ (accessed on 4 February 2020).
- NCBI. Available online: https://www.ncbi.nlm.nih.gov/ (accessed on 4 February 2020).
- NCBI (BLAST Glossary). Available online: https://www.ncbi.nlm.nih.gov/books/NBK62051/ (accessed on 23 March 2020).
- String. Available online: https://string-db.org/cgi/input.pl?sessionId=3nHd3fJD9mAv&input_page_active_form=multiple_identifiers (accessed on 4 February 2020).
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2018, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panther. Available online: https://www.pantherdb.org/ (accessed on 4 September 2019).
- REVIGO. Available online: http://revigo.irb.hr/ (accessed on 4 September 2019).
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.B.; Daly, M.J.; et al. The genome analysis toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef] [Green Version]
- Wysoker, A.; Tibbetts, K.; Fennel, T. Picard Tools Version 2.5. 2013. Available online: http://broadinstitute.github.io/picard/ (accessed on 6 January 2020).
- Depristo, M.A.; Banks, E.; Poplin, R.; Garimella, K.V.; Maguire, J.R.; Hartl, C.; Philippakis, A.A.; Del Angel, G.; Rivas, M.A.; Hanna, M.; et al. A framework for variation discovery and genotyping using next-generation DNA sequencing data. Nat. Genet. 2011, 43, 491–498. [Google Scholar] [CrossRef]
- Van Der Auwera, G.A.; Carneiro, M.O.; Hartl, C.; Poplin, R.; Del Angel, G.; Levy-Moonshine, A.; Jordan, T.; Shakir, K.; Roazen, D.; Thibault, J.; et al. From FastQ Data to High-Confidence Variant Calls: The Genome Analysis Toolkit Best Practices Pipeline. Curr. Protoc. Bioinform. 2013, 43, 1–33. [Google Scholar] [CrossRef]
- McLaren, W.; Gil, L.; Hunt, S.E.; Riat, H.S.; Ritchie, G.R.S.; Thormann, A.; Flicek, P.; Cunningham, F. The Ensembl Variant Effect Predictor. Genome Biol. 2016, 17, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- KEGG Pathway Database. Available online: https://www.genome.jp/kegg/pathway.html (accessed on 4 February 2020).
- miRbase. Available online: http://www.mirbase.org/ (accessed on 4 February 2020).
- Griffiths-Jones, S.; Grocock, R.J.; Van Dongen, S.; Bateman, A.; Enright, A.J. miRBase: microRNA sequences, targets and gene nomenclature. Nucleic Acids Res. 2006, 34, D140–D144. [Google Scholar] [CrossRef] [PubMed]
- Lago, L.V.; Da Silva, A.N.; Zanella, E.L.; Marques, M.G.; Peixoto, J.D.O.; Da Silva, M.V.G.; Ledur, M.C.; Zanella, R. Identification of Genetic Regions Associated with Scrotal Hernias in a Commercial Swine Herd. Vet. Sci. 2018, 5, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joaquim, L.B.; Chud, T.C.S.; Marchesi, J.A.P.; Savegnago, R.P.; Buzanskas, M.E.; Zanella, R.; Cantão, M.E.; Peixoto, J.O.; Ledur, M.C.; Irgang, R.; et al. Genomic structure of a crossbred Landrace pig population. PLoS ONE 2019, 14, e0212266. [Google Scholar] [CrossRef]
- Liao, X.J.; Li, L.; Zhang, Z.Y.; Long, Y.; Yang, B.; Ruan, G.R.; Su, Y.; Ai, H.S.; Zhang, W.C.; Deng, W.Y.; et al. Susceptibility loci for umbilical hernia in swine detected by genome-wide association. Russ. J. Genet. 2015, 51, 1000–1006. [Google Scholar] [CrossRef]
- Long, Y.; Su, Y.; Ai, H.; Zhang, Z.; Yang, B.; Ruan, G.; Xiao, S.; Liao, X.; Ren, J.; Huang, L.; et al. A genome-wide association study of copy number variations with umbilical hernia in swine. Anim. Genet. 2016, 47, 298–305. [Google Scholar] [CrossRef]
- QuickGO. Available online: https://www.ebi.ac.uk/QuickGO/ (accessed on 4 February 2020).
- Levine, B.; Mizushima, N.; Virgin, H.W. Autophagy in immunity and inflammation. Nat. Cell Biol. 2011, 469, 323–335. [Google Scholar] [CrossRef] [PubMed]
- Levine, B.; Klionsky, D.J. Development by Self-Digestion. Dev. Cell 2004, 6, 463–477. [Google Scholar] [CrossRef]
- Kroemer, G. Autophagy: A druggable process that is deregulated in aging and human disease. J. Clin. Investig. 2015, 125, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Baehrecke, E.H. Eaten alive: Novel insights into autophagy from multicellular model systems. Trends Cell Biol. 2015, 25, 376–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z.; Klionsky, D.J. Eaten alive: A history of macroautophagy. Nat. Cell Biol. 2010, 12, 814–822. [Google Scholar] [CrossRef] [Green Version]
- Marcelino, D.E.P.; Souza, M.R.; Auler, M.E.; Savoldi, I.R.; Ibelli, A.M.G.; Peixoto, J.D.O. Expressão dos genes MAP1LC3C e EPYC em suínos normais e afetados com hérnia umbilical. In Anais da 13thJornada de Iniciação Científica; Embrapa Suínos e Aves: Concórdia, UNC, 2019; pp. 34–35. [Google Scholar]
- Van Limbergent, J.; Stevens, C.; Nimmo, E.R.; Wilson, D.C.; Satsangi, J. Autophagy: From basic science to clinical application. Mucosal Immunol. 2009, 2, 315–330. [Google Scholar] [CrossRef] [Green Version]
- Carlos, M.M.L.; Freitas, P.D.F.S. Estudo da cascata de coagulação sanguínea e seus valores de referência. Acta Vet. Bras. 2007. [Google Scholar] [CrossRef]
- Iturry-Yamamoto, G.; Portinho, C. Sistema complemento: Ativação, regulação e deficiências congênitas e adquiridas. Revista Associação Médica Brasileira 2001, 47, 41–51. [Google Scholar] [CrossRef] [Green Version]
- Xue, X.; Wu, J.; Ricklin, D.; Forneris, F.; Di Crescenzio, P.; Schmidt, C.Q.; Granneman, J.; Sharp, T.H.; Lambris, J.D.; Gros, P. Regulator-dependent mechanisms of C3b processing by factor I allow differentiation of immune responses. Nat. Struct. Mol. Biol. 2017, 24, 643–651. [Google Scholar] [CrossRef]
- Winterfeld, G.A.; Khodair, A.I.; Schmidt, R.R. O-Glycosyl Amino Acids by 2-Nitrogalactal Concatenation − Synthesis of a Mucin-Type O-Glycan. Eur. J. Org. Chem. 2003, 2003, 1009–1021. [Google Scholar] [CrossRef]
- Brockhausen, I. Pathways of O-glycan biosynthesis in cancer cells. Biochim. Biophys. Acta (BBA) Gen. Subj. 1999, 1473, 67–95. [Google Scholar] [CrossRef]
- Blalock, T.D.; Spurr-Michaud, S.J.; Tisdale, A.S.; Heimer, S.R.; Gilmore, M.S.; Ramesh, V.; Gipson, I.K. Functions of MUC16 in Corneal Epithelial Cells. Investig. Opthalmol. Vis. Sci. 2007, 48, 4509–4518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alberts, B.; Johnson, A.; Lewis, J.; Morgan, D.; Raff, M.; Roberts, K.; Walter, P. Molecular Biology of the Cell, 6th ed.; Garland Science: New York, NY, USA, 2014. [Google Scholar]
- Huang, S.; Ingber, D.E. The structural and mechanical complexity of cell-growth control. Nat. Cell Biol. 1999, 1, E131–E138. [Google Scholar] [CrossRef] [PubMed]
- Khalili, A.A.; Ahmad, M.R. A review of cell adhesion studies for biomedical and biological applications. Int. J. Mol. Sci. 2015, 16, 18149–18184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grivicich, I.; Regner, A.; Rocha, A.B. Morte celular por apoptose. Rev. Bras. Cancerol. 2007, 53, 335–343. [Google Scholar]
- Puetz, S.; Lubomirov, L.T.; Pfitzer, G. Regulation of smooth muscle contraction by small GTPases. Physiology 2009, 24, 342–356. [Google Scholar] [CrossRef] [Green Version]
- Sato, T.; Konomi, K.; Yamasaki, S.; Aratani, S.; Tsuchimochi, K.; Yokouchi, M.; Masuko-Hongo, K.; Yagishita, N.; Nakamura, H.; Komiya, S.; et al. Comparative analysis of gene expression profiles in intact and damaged regions of human osteoarthritic cartilage. Arthritis Rheum. 2006, 54, 808–817. [Google Scholar] [CrossRef]
- Zhao, X.; Du, Z.-Q.; Vukasinovic, N.; Rodriguez, F.; Clutter, A.C.; Rothschild, M.F. Association of HOXA10, ZFPM2, and MMP2 genes with scrotal hernias evaluated via biological candidate gene analyses in pigs. Am. J. Veter Res. 2009, 70, 1006–1012. [Google Scholar] [CrossRef]
- Du, Z.-Q.; Zhao, X.; Vukasinovic, N.; Rodríguez, F.; Clutter, A.C.; Rothschild, M.F. Association and Haplotype Analyses of Positional Candidate Genes in Five Genomic Regions Linked to Scrotal Hernia in Commercial Pig Lines. PLoS ONE 2009, 4, e4837. [Google Scholar] [CrossRef] [Green Version]
- Piórkowska, K.; Żukowski, K.; Ropka-Molik, K.; Tyra, M. Detection of genetic variants between different Polish Landrace and Puławska pigs by means of RNA-seq analysis. Anim. Genet. 2018, 49, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Lazarides, E. Intermediate Filaments: A Chemically Heterogeneous, Developmentally Regulated Class of Proteins. Annu. Rev. Biochem. 1982, 51, 219–250. [Google Scholar] [CrossRef] [PubMed]
- Onteru, S.K.; Fan, B.; Du, Z.-Q.; Garrick, D.; Stalder, K.J.; Rothschild, M.F. A whole-genome association study for pig reproductive traits. Anim. Genet. 2011, 43, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Moore, K.L.; Persaud, T.V.N.; Torchia, M.G. Embriologia Básica, 9th ed.; Elsevier: Rio de Janeiro, Brazil, 2016; p. 463. Available online: https://ia801004.us.archive.org/30/items/Embriologia_bsica_9._ed._-_www.meulivro.biz/Embriologia_bsica_9._ed._-_www.meulivro.biz.pdf (accessed on 16 December 2019).
- Stachowiak, M.; Nowacka-Woszuk, J.; Szydlowski, M.; Switonski, M. The ACACA and SREBF1 genes are promising markers for pig carcass and performance traits, but not for fatty acid content in the longissimus dorsi muscle and adipose tissue. Meat Sci. 2013, 95, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Tian, M.; Zhang, X.; Ye, P.; Tao, Q.; Zhang, L.; Ding, Y.; Chu, M.; Zhang, X.; Yin, Z. MicroRNA-21andmicroRNA-214play important role in reproduction regulation during porcine estrous. Anim. Sci. J. 2018, 89, 1398–1405. [Google Scholar] [CrossRef]
- Guo, Y.; Chen, Y.; Zhang, Y.; Zhang, Y.; Chen, L.; Mo, D. Up-regulated miR-145 Expression Inhibits Porcine Preadipocytes Differentiation by Targeting IRS1. Int. J. Biol. Sci. 2012, 8, 1408–1417. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Annotated Transcripts | SH | UH | ||
|---|---|---|---|---|
| LncRNA | 68 | 0.53% | 77 | 0.60% |
| MiRNA | 5 | 0.04% | 4 | 0.03% |
| Mt rRNA | 2 | 0.02% | 2 | 0.02% |
| Mt tRNA | 1 | 0.01% | 1 | 0.01% |
| Processed pseudogene | 1 | 0.01% | 0 | 0% |
| Protein-coding | 12,601 | 98.55% | 12,598 | 98.50% |
| Pseudogene | 90 | 0.70% | 91 | 0.71% |
| Ribozime | 1 | 0.01% | 1 | 0.01% |
| ScaRNA | 1 | 0.01% | 1 | 0.01% |
| SnoRNA | 13 | 0.10% | 14 | 0.11% |
| SnRNA | 1 | 0.01% | 0 | 0% |
| Y RNA | 2 | 0.02% | 1 | 0.01% |
| Total annotated transcripts | 12,786 | 12,790 | ||
| ENSEMBL ID | Gene Symbol | Chromosome | Gene Name | SH-logFC | SH-FDR | UH-logFC | UH-FDR |
|---|---|---|---|---|---|---|---|
| ENSSSCG00000001832 | ACAN | 7 | Aggrecan | 2.913 | 0.001 | 2.788 | 0.040 |
| ENSSSCG00000034213 | ACER2 | 1 | Alkaline ceramidase 2 | −3.066 | 0.001 | −2.373 | 0.004 |
| ENSSSCG00000036223 | ACKR1 | 4 | Atypical chemokine receptor 1 (Duffy blood group) | −1.119 | 0.030 | −1.023 | 0.034 |
| ENSSSCG00000010370 | ANXA8 | 14 | Annexin A8 | 2.026 | 0.000 | 1.744 | 0.004 |
| ENSSSCG00000032709 | ARL4A | 9 | ADP ribosylation factor like GTPase 4A | −1.308 | 0.001 | −1.199 | 0.031 |
| ENSSSCG00000033350 | BCHE | 13 | Butyrylcholinesterase | 1.011 | 0.028 | 1.557 | 0.009 |
| ENSSSCG00000028567 | BTNL9 | 2 | Butyrophilin like 9 | −1.356 | 0.010 | −2.268 | 0.016 |
| ENSSSCG00000002662 | C16orf74 | 6 | Chromosome 16 open reading frame 74 | 1.818 | 0.025 | 1.661 | 0.008 |
| ENSSSCG00000006736 | CD2 | 4 | CD2 molecule | 2.275 | 0.028 | −1.990 | 0.002 |
| ENSSSCG00000009138 | CFI | 8 | Complement factor I | 2.254 | 0.000 | 1.904 | 0.029 |
| ENSSSCG00000011524 | CHL1 | 13 | Cell adhesion molecule L1 like | −1.776 | 0.004 | −1.321 | 0.009 |
| ENSSSCG00000021588 | DAPK2 | 1 | Death associated protein kinase 2 | −1.347 | 0.026 | −1.473 | 0.013 |
| ENSSSCG00000012126 | GPM6B | X | Glycoprotein M6B | −1.047 | 0.009 | −1.244 | 0.022 |
| ENSSSCG00000002847 | GPT2 | 6 | Glutamic--pyruvic transaminase 2 | −3.752 | 0.002 | 1.571 | 0.029 |
| ENSSSCG00000036438 | GPX3 | 16 | Glutathione peroxidase 3 | −2.184 | 0.000 | −2.762 | 0.006 |
| ENSSSCG00000017010 | INSYN2B | 16 | Inhibitory synaptic factor family member 2B | 3.436 | 0.000 | 1.667 | 0.020 |
| ENSSSCG00000002245 | KATNBL1 | 7 | Katanin regulatory subunit B1 like 1 | 0.877 | 0.040 | 1.147 | 0.002 |
| ENSSSCG00000010325 | KCNMA1 | 14 | Potassium calcium-activated channel subfamily M α 1 | 0.982 | 0.042 | 1.652 | 0.001 |
| ENSSSCG00000034838 | MAP1LC3C | 10 | Microtubule associated protein 1 light chain 3 γ | 6.715 | 0.000 | 3.819 | 0.002 |
| ENSSSCG00000004191 | MOXD1 | 1 | Monooxygenase DBH like 1 | −2.980 | 0.010 | 2.570 | 0.033 |
| ENSSSCG00000011133 | PFKFB3 | 10 | 6-phosphofructo-2-kinase/fructose-2,6-biphosphatase 3 | −1.135 | 0.008 | −1.450 | 0.010 |
| ENSSSCG00000007528 | PHACTR3 | 17 | Phosphatase and actin regulator 3 | 1.727 | 0.012 | 1.986 | 0.028 |
| ENSSSCG00000007470 | RIPOR3 | 17 | RIPOR family member 3 | −1.119 | 0.049 | −1.377 | 0.042 |
| ENSSSCG00000009111 | SYNPO2 | 8 | Synaptopodin 2 | −1.293 | 0.006 | −1.194 | 0.045 |
| ENSSSCG00000014834 | UCP3 | 9 | Uncoupling protein 3 | −2.693 | 0.024 | −3.262 | 0.016 |
| ENSSSCG00000008501 | VIT | 3 | Vitrin | −2.317 | 0.004 | −3.709 | 0.029 |
| ENSSSCG00000015766 | WDR17 | 15 | WD repeat domain 17 | −1.761 | 0.000 | −1.773 | 0.019 |
| ENSSSCG00000013714 | 2 | 2.087 | 0.048 | 1.813 | 0.019 | ||
| ENSSSC00000037142 | AEMK02000452.1 | Cysteine-rich protein 1 | 1.222 | 0.000 | −4.322 | 0.010 | |
| ENSSSCG00000031037 | 14 | 3.813 | 0.000 | −4.021 | 0.029 | ||
| ENSSSCG00000032582 | 14 | 3.873 | 0.000 | −3.822 | 0.012 | ||
| ENSSSCG00000036224 | 3 | 3.369 | 0.001 | −3.010 | 0.045 | ||
| ENSSSCG00000036983 | AEMK02000452.1 | 4.568 | 0.001 | −5.951 | 0.001 | ||
| ENSSSCG00000037009 | AEMK02000452.1 | 3.679 | 0.024 | −3.785 | 0.031 | ||
| ENSSSCG00000039111 | AEMK02000452.1 | 4.596 | 0.000 | 1.180 | 0.041 |
| Gene ID | Description | e-Value | Query Cover (%) | Identity (%) | Accession (RefSeq) |
|---|---|---|---|---|---|
| ENSSSCG00000013714 | Mucin-16 [Sus scrofa] | 8 × 10−157 | 100 | 93.1 | XP_020940777.1 |
| ENSSSCG00000036224 | Ig kappa chain V-C region (PLC18) [Sus scrofa domesticus] | 5 × 10−89 | 82 | 85 | PT0219 |
| ENSSSCG00000031037 | Immunoglobulin lambda-like polypeptide 5 precursor [Sus scrofa] | 3 × 10−71 | 99 | 99.09 | NP_001230248.1 |
| ENSSSCG00000032582 | Immunoglobulin lambda-like polypeptide 5 precursor [Sus scrofa] | 7 × 10−68 | 94 | 92.04 | NP_001230248.1 |
| ENSSSCG00000036983 | IgG heavy chain precursor [Sus scrofa] | 0.0 | 74 | 79.78 | BAM75547.1 |
| ENSSSCG00000037009 | IgG heavy chain precursor [Sus scrofa] | 0.0 | 100 | 100 | BAM75542.1 |
| ENSSSCG00000039111 | IgG heavy chain constant region [Sus scrofa] | 1 × 10−74 | 100 | 100 | BAM66306.1 |
| ENSSSCG00000037142 | Cysteine-rich protein 1 [Camelus dromedarius] | 3 × 10−41 | 36 | 94.37 | KAB1277051.1 |
| Superclusters | Genes | ||
|---|---|---|---|
| Upregulated in Both Groups | Downregulated in Both Groups | Opposite Expression Profile | |
| Cell adhesion (GO:0022610) | CHL1 | CD2 | |
| Biological regulation (GO:0065007) | ANXA8 | ACKR1, SYNPO2 | MOXD1, ENSSSCG00000036224, ENSSSCG00000039111 |
| Cellular process (GO:0009987) | PHACTR3, ANXA8, MAP1LC3C | VIT, SYNPO2, BTNL9 | GPT2, MOXD1, CD2, ENSSSCG00000039111, ENSSSCG00000036224 |
| Development (GO:0032502) | PHACTR3 | VIT | |
| Immune system process (GO:0002376) | ACER2, BTNL9 | CD2, ENSSSCG00000032582, ENSSSCG00000039111, ENSSSCG00000031037, ENSSSCG00000036224 | |
| Location (GO:0051179) | ANXA8, MAP1LC3C | ARL4A, UCP3 | ENSSSCG00000036224, ENSSSCG00000039111 |
| Metabolic process (GO:0008152) | UCP3, PFKFB3 | GPT2, MOXD1, ENSSSCG00000036224, ENSSSCG00000039111 | |
| Multicellular organismal process (GO:0032501) | ACAN, PHACTR3 | ACKR1, CHL1, GPM6B, VIT | |
| Response to stimulus (GO:0050896) | ACKR1, GPX3, UCP3 | ENSSSCG00000037009, ENSSSCG00000036983, ENSSSCG00000036224, ENSSSCG00000039111 | |
| Polymorphism Type | Scrotal Hernia | Umbilical Hernia | ||
|---|---|---|---|---|
| N° | (%) | N° | (%) | |
| Insertion | 10 | 14.93 | 6 | 7.90 |
| Deletion | 4 | 5.97 | 3 | 5.26 |
| SNP | 53 | 79.10 | 67 | 86.84 |
| Total | 67 | 100 | 76 | 100 |
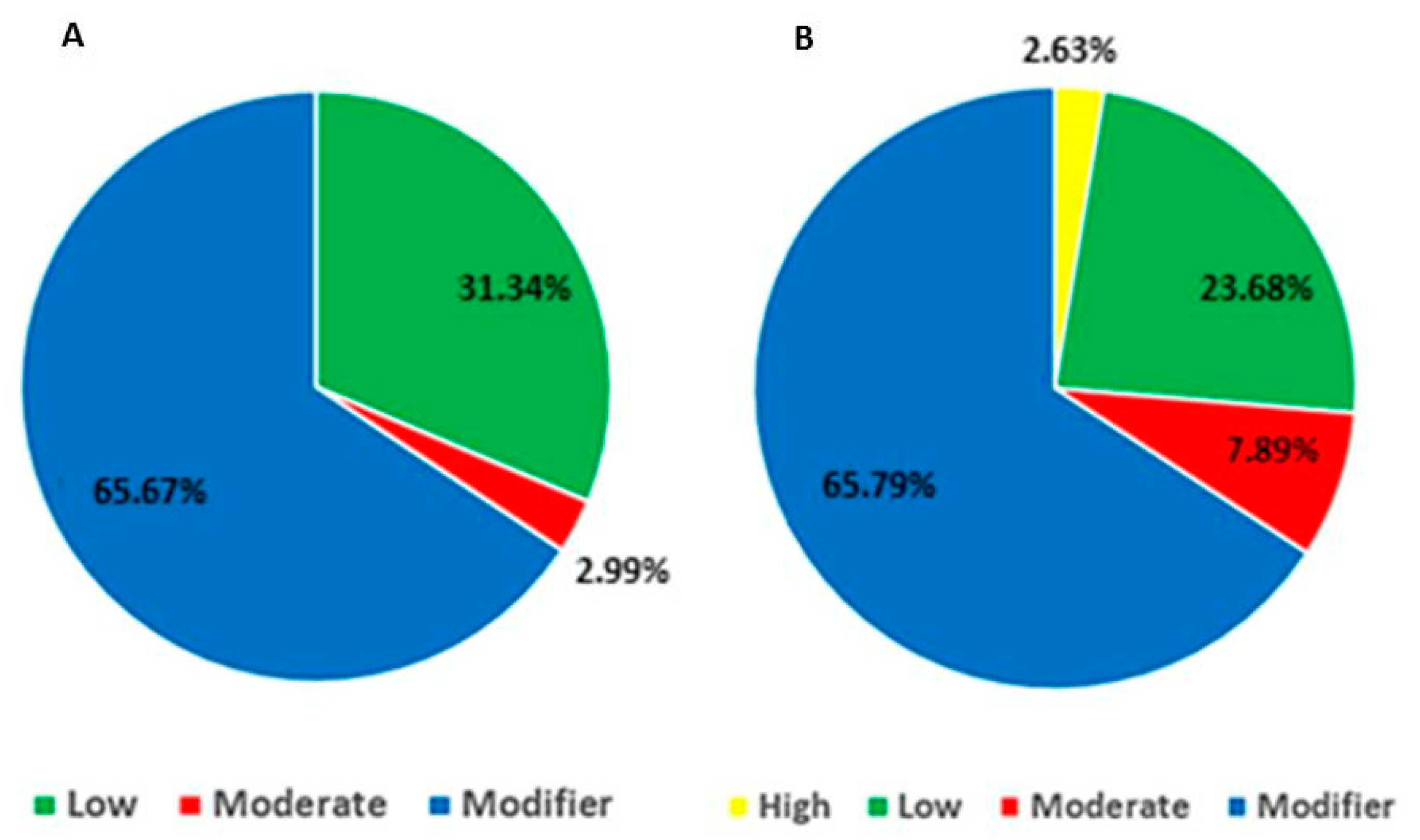
| Variant Type | Scrotal Hernia (%) | Umbilical Hernia (%) |
|---|---|---|
| Intronic | 23.88 | 5.26 |
| Synonym | 29.85 | 23.68 |
| Missense | 2.99 | 7.89 |
| Splicing | 1.49 | - |
| UTR5′ | 5.97 | 6.58 |
| UTR3′ | 34.33 | 47.37 |
| Downstream | 1.49 | 6.58 |
| Frameshift | - | 2.63 |
| Group | Variant | Location | Impact | Gene | SIFT |
|---|---|---|---|---|---|
| Scrotal hernia | rs325370594 | 16:20418972-20418972 | Moderate | RAI14 | Tolerant (1) |
| - | 7:64303141-64303141 | Moderate | RALGAPA1 | Tolerant (0.63) | |
| Umbilical hernia | rs325089032 | 6:81571496-81571496 | Moderate | ELOA | Tolerant (0.1) |
| rs327289001 | 3:17254444-17254444 | Moderate | ITGAM | Deleterious (0.01) | |
| rs789266896 | 3:17628688-17628688 | Moderate | RNF40 | Tolerant (0.6) | |
| rs330957838 | 3:17468302-17468302 | Moderate | SETD1A | Tolerant low confidence (0.34) | |
| rs337670844 | 3:17399477-17399477 | Moderate | ZNF646 | Tolerant (0.08) | |
| rs323115420 | 3:16964045-16964045 | Moderate | ZNF713 | Tolerant (0.65) |
| Variant | Gene | Pathway (ssc ¹) |
|---|---|---|
| New (Frameshift) | SEC62 | Protein exports (ssc03060); Protein processing in the endoplasmic reticulum (ssc04141). |
| rs327289001 (Missense) | ITGAM | Rap1 signaling path (ssc04015); Phagosome (ssc04145); Cell adhesion molecules (CAMs) (ssc04514); Hematopoietic cell line (ssc04640); Transendothelial migration of leukocytes (ssc04670); Regulation of the actin cytoskeleton (ssc04810); Whooping cough (ssc05133); Legionellosis (ssc05134); Leishmaniasis (ssc05140); Amebiasis (ssc05146); Infection by Staphylococcus aureus (ssc05150); Tuberculosis (ssc05152); Incorrect regulation of transcription in cancer (ssc05202) |
| rs330957838 (Missense) | SETD1A | Lysine degradation (ssc00310) |
| David Term | Biological Process | Enriched Genes |
|---|---|---|
| GO:0010604 | Positive regulation of the metabolic process of macromolecules | TBX3, MYRF, MYLIP, PARP3 |
| GO:0055088 | Lipid homeostasis | ACACA, MYYLIP |
| GO:0009893 | Positive regulation of the metabolic process | TBX3, MYRF, MYLIP, PARP3 |
| GO:0043170 | Metabolic process of the macromolecule | TBX3, MYRF, MYLIP, PARP3, ACACA, DDB2 |
| GO:0065008 | Regulation of biological quality | TBX3, ACACA, MYLIP, PARP3 |
| GO:0045935 | Positive regulation of the compound metabolic process containing nucleobase | TBX3, MYRF, PARP3 |
| GO:0051173 | Positive regulation of the metabolic process of nitrogen compounds | TBX3, MYRF, PARP3 |
| GO:0042592 | Homeostatic process | ACACA, MYLIP, PARP3 |
| GO:0033554 | Cellular stress response | TBX3, DDB2, PARP3 |
| GO:0060249 | Anatomical structure homeostasis | ACACA, PARP3 |
| GO Term | Biological Process | Enriched Genes |
|---|---|---|
| GO:0010468 | Regulation of gene expression | RPRD1A, RNF40, ELOA, VIM, ZNF629, ZNF713 |
| GO:0010977 | Negative regulation of the development of neuron projection | EPHB2, VIM |
| GO:0060255 | Regulation of the metabolic process of macromolecules | ITGB2, RPRD1A, RNF40, ELOA, VIM, ZNF629, ZNF713 |
| GO:0031345 | Negative regulation of cellular projection organization | EPHB2 e VIM |
| GO:0019222 | Regulation of the metabolic process | ITGB2, RPRD1A, RNF40, ELOA, VIM, ZNF629, ZNF713 |
| GO:0045665 | Negative regulation of neuron differentiation | EPHB2, VIM |
| ENSEMBL ID | Name/Symbol | miRBase | Location | Group |
|---|---|---|---|---|
| ENSSSCG00000018513 | ssc-mir-145 (MIR145) | MI0002417 | 2: 150.580.126-150.580.211 | SH |
| ENSSSCG00000018758 | ssc-mir-214 (MIR214) | MI0002441 | 9: 114.527.990-114.528.101 | SH/UH |
| ENSSSCG00000019065 | ssc-mir-186 (MIR186) | MI0002456 | 6: 141.943.328-141.943.409 | SH |
| ENSSSCG00000034634 | ssc-mir-6782 | MI0031620 | AEMK02000489.1: 40.305-40.379 | UH |
| ENSSSCG00000035742 | - | - | 12: 14.578.144-14.578.210 | SH/UH |
| ENSSSCG00000037094 | ssc-mir-9810 | MI0031577 | 4: 83.070.363-83.070.457 | SH/UH |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodrigues, A.F.G.; Ibelli, A.M.G.; Peixoto, J.d.O.; Cantão, M.E.; Oliveira, H.C.d.; Savoldi, I.R.; Souza, M.R.; Mores, M.A.Z.; Carreño, L.O.D.; Ledur, M.C. Genes and SNPs Involved with Scrotal and Umbilical Hernia in Pigs. Genes 2021, 12, 166. https://doi.org/10.3390/genes12020166
Rodrigues AFG, Ibelli AMG, Peixoto JdO, Cantão ME, Oliveira HCd, Savoldi IR, Souza MR, Mores MAZ, Carreño LOD, Ledur MC. Genes and SNPs Involved with Scrotal and Umbilical Hernia in Pigs. Genes. 2021; 12(2):166. https://doi.org/10.3390/genes12020166
Chicago/Turabian StyleRodrigues, Ariene Fernanda Grando, Adriana Mércia Guaratini Ibelli, Jane de Oliveira Peixoto, Maurício Egídio Cantão, Haniel Cedraz de Oliveira, Igor Ricardo Savoldi, Mayla Regina Souza, Marcos Antônio Zanella Mores, Luis Orlando Duitama Carreño, and Mônica Corrêa Ledur. 2021. "Genes and SNPs Involved with Scrotal and Umbilical Hernia in Pigs" Genes 12, no. 2: 166. https://doi.org/10.3390/genes12020166