A Comparative Transcriptome Analysis of Volvariella volvacea Identified the Candidate Genes Involved in Fast Growth at the Mycelial Growth Stage

Abstract
:1. Introduction
2. Results
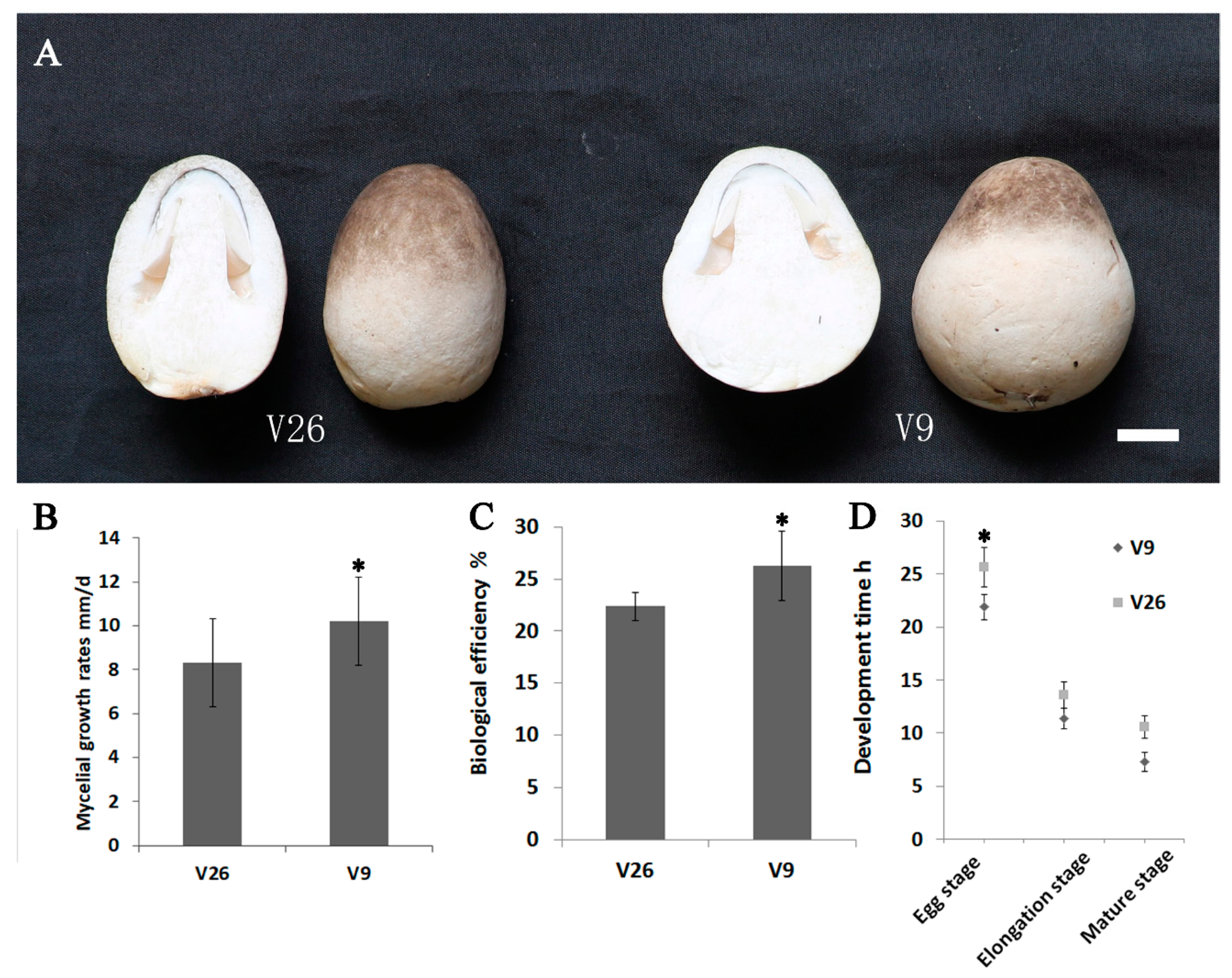
2.1. Comparations of the Growth Rates between V. volvacea Strain v9 and Strain v26
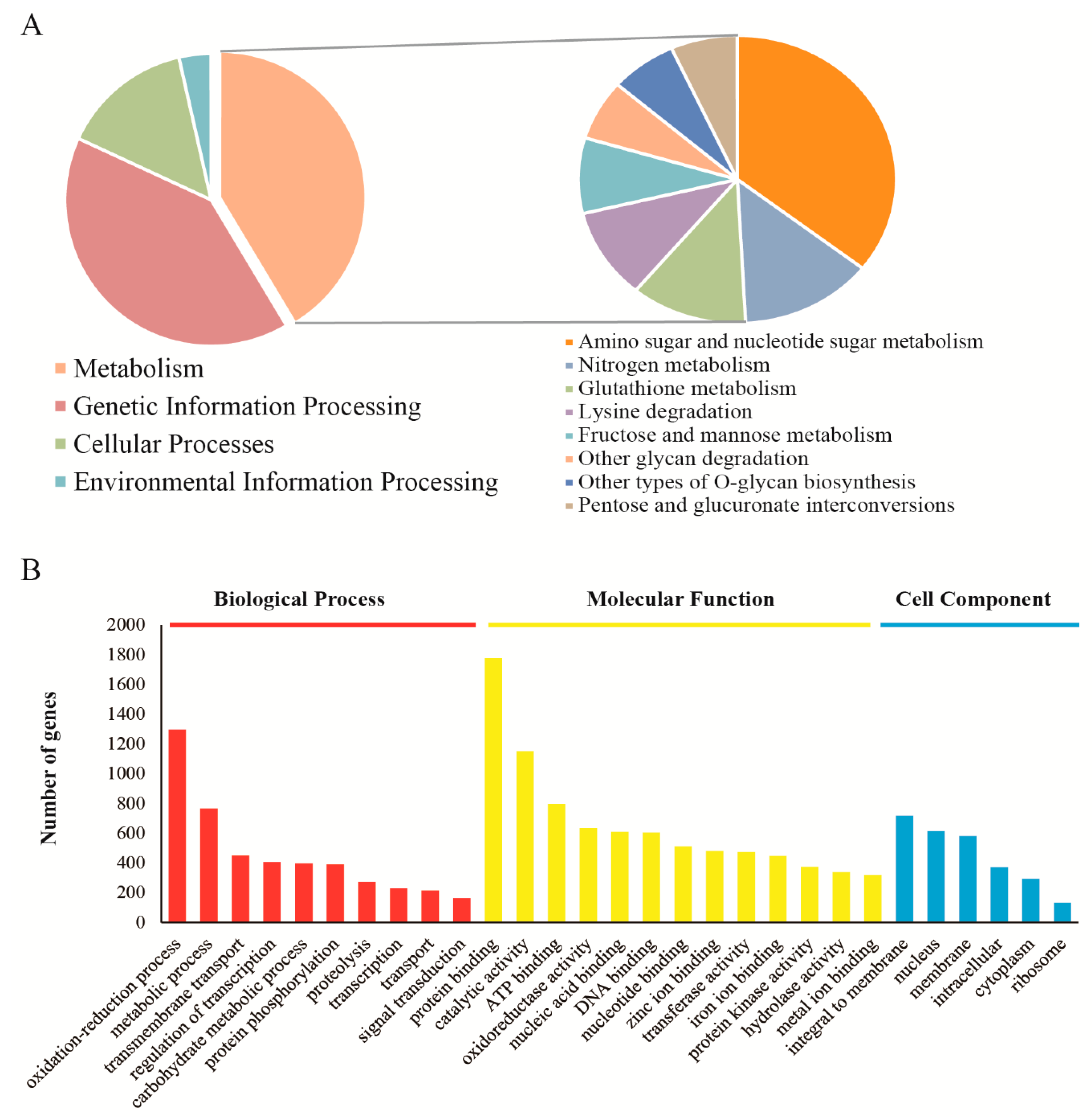
2.2. Sequencing, Assembly and Functional Annotation of V. volvacea
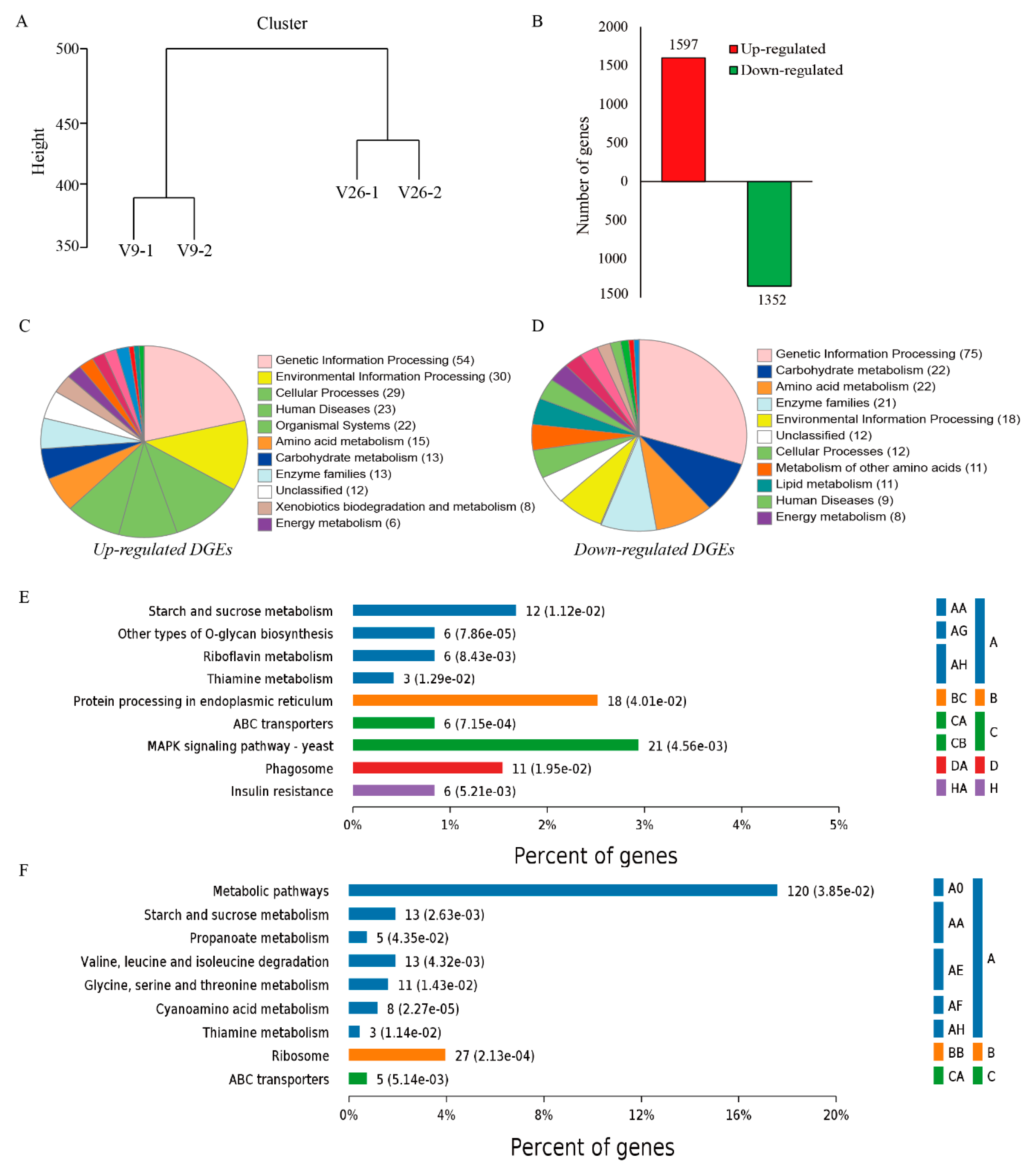
2.3. Gene Expression and Identification of DEGs
2.4. Genes Involved in MAPK Signaling Pathway
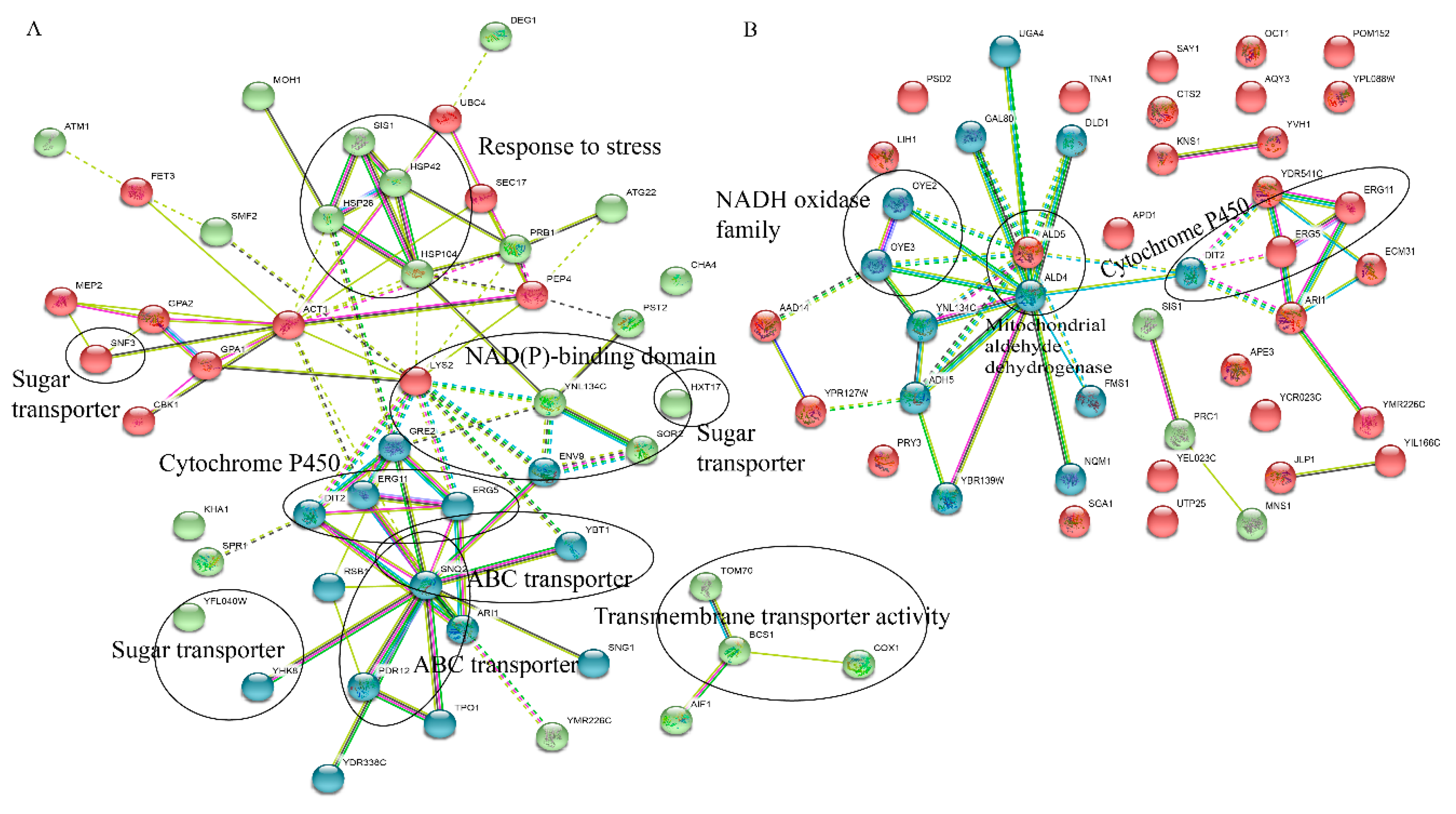
2.5. String Analysis
3. Discussion
4. Materials and Methods
4.1. Strains and Culture Conditions
4.2. RNA Extraction and cDNA Library Construction and Sequencing
4.3. Sequence Processing and Bioinformatics Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chang, S.; Miles, P. Edible Mushroom and Their Cultivation; CRC Press: New York, NY, USA, 1989; Volume 120, pp. 307–312. [Google Scholar]
- Wang, R.; Cao, H.; Zhang, G.; Tan, Q. Scientific explorations and commercial sales of the straw mushroom Volvariella volvacea (Bull.) Singer in Republican China: A brief review. Indian J. Hist. Sci. 2017, 52, 95–101. [Google Scholar]
- Bao, D.; Gong, M.; Zheng, H.; Chen, M.; Zhang, L.; Wang, H.; Jiang, J.; Wu, L.; Zhu, Y.; Zhu, G. Sequencing and comparative analysis of the straw mushroom (Volvariella volvacea) genome. PLoS ONE 2013, 8, e58294. [Google Scholar] [CrossRef] [PubMed]
- Kishida, E.; Kinoshita, C.; Sone, Y.; Misaki, A. Structures and antitumor activities of polysaccharides isolated from mycelium of Volvariella volvacea. Biosci. Biotechnol. Biochem. 1992, 56, 1308–1309. [Google Scholar] [CrossRef] [PubMed]
- Roy, A.; Prasad, P.; Gupta, N. Volvariella volvacea: A macrofungus having nutritional and health potential. Asian J. Pharm. Technol. 2014, 4, 110–113. [Google Scholar]
- Chang, S. Production of the straw-mushroom (Volvariella volvacea) from cotton wastes. Mushroom J. 1974, 98, 1019–1024. [Google Scholar]
- Ohm, R.A.; De Jong, J.F.; Lugones, L.G.; Aerts, A.; Kothe, E.; Stajich, J.E.; De Vries, R.P.; Record, E.; Levasseur, A.; Baker, S.E. Genome sequence of the model mushroom Schizophyllum commune. Nat. Biotechnol. 2010, 28, 957. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Xu, J.; Liu, C.; Zhu, Y.; Nelson, D.R.; Zhou, S.; Li, C.; Wang, L.; Guo, X.; Sun, Y. Genome sequence of the model medicinal mushroom Ganoderma lucidum. Nat. Commun. 2012, 3, 913. [Google Scholar] [CrossRef] [Green Version]
- Morin, E.; Kohler, A.; Baker, A.R.; Foulongne-Oriol, M.; Lombard, V.; Nagye, L.G.; Ohm, R.A.; Patyshakuliyeva, A.; Brun, A.; Aerts, A.L. Genome sequence of the button mushroom Agaricus bisporus reveals mechanisms governing adaptation to a humic-rich ecological niche. Proc. Natl. Acad. Sci. USA 2012, 109, 17501–17506. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Gong, Y.; Cai, Y.; Liu, W.; Zhou, Y.; Xiao, Y.; Xu, Z.; Liu, Y.; Lei, X.; Wang, G. Genome sequence of the edible cultivated mushroom Lentinula edodes (Shiitake) reveals insights into lignocellulose degradation. PLoS ONE 2016, 11, e0160336. [Google Scholar] [CrossRef]
- Chen, B.; Gui, F.; Xie, B.; Deng, Y.; Sun, X.; Lin, M.; Tao, Y.; Li, S. Composition and expression of genes encoding carbohydrate-active enzymes in the straw-degrading mushroom Volvariella volvacea. PLoS ONE 2013, 8, e58780. [Google Scholar]
- Wang, W.; Wang, L.; Chen, B.; Mukhtar, I.; Xie, B.; Li, Z.; Meng, L. Characterization and expression pattern of homeobox transcription factors in fruiting body development of straw mushroom Volvariella volvacea. Fungal Biol. 2019, 123, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Yu, C.; Zhao, Y.; Liu, S.; Wang, H.; Wang, C.; Guo, L.; Chen, M. Changes in Mannitol Content, Regulation of Genes Involved in Mannitol Metabolism, and the Protective Effect of Mannitol on Volvariella volvacea at Low Temperature. Biomed. Res. Int. 2019, 2019, 1493721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Z.; Wu, X.; Lv, B.; Wu, G.; Wang, J.; Jiang, W.; Li, P.; He, J.; Chen, J.; Chen, M. A new approach for breeding low-temperature-resistant Volvariella volvacea strains: Genome shuffling in edible fungi. Biotechnol. Appl. Biochem. 2016, 63, 605–615. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; van Peer, A.F.; Yan, J.; Li, X.; Xie, B.; Miao, J.; Huang, Q.; Zhang, L.; Wang, W.; Fu, J. Fruiting body formation in Volvariella volvacea can occur independently of its MAT-A-controlled bipolar mating system, enabling homothallic and heterothallic life cycles. G3: Genesgenomesgenetics 2016, 6, 2135–2146. [Google Scholar] [CrossRef]
- Ahlawat, O.; Ahlawat, K.; Dhar, B. Influence of lignocellulolytic enzymes on substrate colonization and yield in monosporous isolates and parent strains of Volvariella volvacea (Bull. Fr.) Sing. Indian J. Microbiol. 2005, 45, 205. [Google Scholar]
- Ahlawat, O.; Gupta, P.; Dhar, B.; Sagar, T.; Rajendranath, R.; Rathnam, K. Profile of the extracellular lignocellulolytic enzymes activities as a tool to select the promising strains of Volvariella volvacea (Bull. ex Fr.) sing. Indian J. Microbiol. 2008, 48, 389–396. [Google Scholar] [CrossRef] [Green Version]
- Chang, S.T.; Yau, C.K. Volvariella volvacea and its life history. Am. J. Bot. 1971, 58, 552–561. [Google Scholar] [CrossRef]
- Li, Z.; Chen, M.; Yu, C.; Li, Q.; Zhou, F.; Li, Y. Five Steps to Cultivate Volvariella volvacea. Agric. Sci. Technol. 2017, 18, 1593–1594. [Google Scholar]
- Sakinah, M.; Misran, A.; Mahmud, T.; Abdullah, S. A review: Production and postharvest management of Volvariella volvacea. Int. Food Res. J. 2019, 26, 367–376. [Google Scholar]
- Tiwari, S.; Thakur, R.; Shankar, J. Role of heat-shock proteins in cellular function and in the biology of fungi. Biotechnol. Res. Int. 2015, 2015, 132635. [Google Scholar] [CrossRef] [Green Version]
- Aguilera, J.; Randez-Gil, F.; Prieto, J.A. Cold response in Saccharomyces cerevisiae: New functions for old mechanisms. Fems Microbiol. Rev. 2007, 31, 327–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, B.; Feder, M.E.; Kang, L. Evolution of heat-shock protein expression underlying adaptive responses to environmental stress. Mol. Ecol. 2018, 27, 3040–3054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, S.T.; Hayes, W.A. The Biology and Cultivation of Edible Mushrooms; Academic Press: Cambridge, MA, USA, 2013. [Google Scholar]
- Ahlawat, O.; Gupta, P.; Kamal, S.; Dhar, B. Development of molecular and biochemical markers for selecting a potential high yielding strain of paddy straw mushroom (Volvariella volvacea). J. Plant Biochem. Biotechnol. 2008, 17, 57–63. [Google Scholar] [CrossRef]
- Chen, S.; Ma, D.; Ge, W.; Buswell, J.A. Induction of laccase activity in the edible straw mushroom, Volvariella volvacea. FEMS Microbiol. Lett. 2003, 218, 143–148. [Google Scholar] [CrossRef]
- Cai, Y.J.; Buswell, J.A.; Chang, S.T. Production of cellulases and hemicellulases by the straw mushroom, Volvariella volvacea. Mycol. Res. 1994, 98, 1019–1024. [Google Scholar] [CrossRef]
- Brady, S.K.; Sreelatha, S.; Feng, Y.; Chundawat, S.P.; Lang, M.J. Cellobiohydrolase 1 from Trichoderma reesei degrades cellulose in single cellobiose steps. Nat. Commun. 2015, 6, 10149. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.; Liu, X.; Long, L.; Ding, S. Cellobiose dehydrogenase from Volvariella volvacea and its effect on the saccharification of cellulose. Process Biochem. 2017, 60, 52–58. [Google Scholar] [CrossRef]
- Tandrup, T.; Frandsen, K.E.H.; Johansen, K.S.; Berrin, J.-G.; Lo Leggio, L. Recent insights into lytic polysaccharide monooxygenases (LPMOs). Biochem. Soc. Trans. 2018, 46, 1431–1447. [Google Scholar] [CrossRef]
- Mach, R.; Zeilinger, S. Regulation of gene expression in industrial fungi: Trichoderma. Appl. Microbiol. Biotechnol. 2003, 60, 515–522. [Google Scholar] [CrossRef]
- Stricker, A.R.; Mach, R.L.; De Graaff, L.H. Regulation of transcription of cellulases-and hemicellulases-encoding genes in Aspergillus niger and Hypocrea jecorina (Trichoderma reesei). Appl. Microbiol. Biotechnol. 2008, 78, 211. [Google Scholar] [CrossRef]
- Saito, H. Regulation of cross-talk in yeast MAPK signaling pathways. Curr. Opin. Microbiol. 2010, 13, 677–683. [Google Scholar] [CrossRef] [PubMed]
- Stajich, J.E.; Wilke, S.K.; Ahrén, D.; Au, C.H.; Birren, B.W.; Borodovsky, M.; Burns, C.; Canbäck, B.; Casselton, L.A.; Cheng, C. Insights into evolution of multicellular fungi from the assembled chromosomes of the mushroom Coprinopsis cinerea (Coprinus cinereus). Proc. Natl. Acad. Sci. USA 2010, 107, 11889–11894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, P.; Weinzierl, G.; Brachmann, A.; Feldbrügge, M.; Kahmann, R. Mating and pathogenic development of the smut fungus Ustilago maydis are regulated by one mitogen-activated protein kinase cascade. Eukaryot. Cell 2003, 2, 1187–1199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, L.H.; Jian, H.; Song, C.Y.; Bao, D.P.; Shang, X.D.; Wu, D.Q.; Tan, Q.; Zhang, X.H. Transcriptome analysis of candidate genes and signaling pathways associated with light-induced brown film formation in Lentinula edodes. Appl. Microbiol. Biotechnol. 2013, 97, 4977–4989. [Google Scholar] [CrossRef]
- Patel, R.K.; Jain, M. NGS QC Toolkit: A toolkit for quality control of next generation sequencing data. PLoS ONE 2012, 7, e30619. [Google Scholar] [CrossRef]
- Haas, B.J.; Papanicolaou, A.; Yassour, M.; Grabherr, M.; Blood, P.D.; Bowden, J.; Couger, M.B.; Eccles, D.; Li, B.; Lieber, M. De novo transcript sequence reconstruction from RNA-seq using the Trinity platform for reference generation and analysis. Nat. Protoc. 2013, 8, 1494. [Google Scholar] [CrossRef]
- Langmead, B.; Trapnell, C.; Pop, M.; Salzberg, S.L. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 2009, 10, R25. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]
- Xie, C.; Mao, X.; Huang, J.; Ding, Y.; Wu, J.; Dong, S.; Kong, L.; Gao, G.; Li, C.-Y.; Wei, L. KOBAS 2.0: A web server for annotation and identification of enriched pathways and diseases. Nucleic Acids Res. 2011, 39 (Suppl. 2), W316–W322. [Google Scholar] [CrossRef] [Green Version]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P. STRING v11: Protein–protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2018, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sequencing | Raw Reads | 67,399,626 |
| Clean reads | 65,013,746 | |
| Assembly | Unigenes | 14,556 |
| N50 | 2461 | |
| Max length | 16,297 | |
| Min length | 201 | |
| Average length | 1410 | |
| Total assembled bases | 20,524,297 | |
| Annotation | NR | 10,890 (86%) |
| KEGG | 10,714 (85%) |
| Unigene ID | Expression | KEGG | qPCR | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| v9-1 | v9-2 | v26-1 | v26-2 | Log2FoldChange | FDR | Status | KO | Description | Gene Name | ||
| ctg4916_g1 | 98.51 | 90.22 | 20.4 | 27.54 | 2.02721 | 3.76 × 10−31 | up-regulated | K00844 | hexokinase [EC:2.7.1.1] | HK | 4.51/1 |
| ctg7672_g1 | 20.81 | 19.75 | 8.6 | 8.55 | 1.28427 | 1.04 × 10−36 | up-regulated | K01178 | glucoamylase [EC:3.2.1.3] | SGA1 | |
| ctg164_g1 | 0.99 | 0.78 | 23.19 | 23 | −4.7117 | 1.13 × 10−147 | down-regulated | K01178 | glucoamylase [EC:3.2.1.3] | SGA1 | |
| ctg2341_g1 | 3.76 | 2.34 | 1.03 | 1.36 | 1.34333 | 9.35 × 10−2 | up-regulated | K01178 | glucoamylase [EC:3.2.1.3] | SGA1 | 2.23/1 |
| ctg9698_g1 | 39.85 | 34.04 | 15.43 | 16.83 | 1.17589 | 3.42 × 10−20 | up-regulated | K01176 | alpha-amylase [EC:3.2.1.1] | AMY | |
| ctg11581_g1 | 271.23 | 249.79 | 117.41 | 125.9 | 1.0762 | 7.15 × 10−95 | up-regulated | K01176 | alpha-amylase [EC:3.2.1.1] | AMY | 1.99/1 |
| ctg8677_g2 | 3.57 | 3.94 | 11.08 | 10.41 | −1.5319 | 7.84 × 10−18 | down-regulated | K01176 | alpha-amylase [EC:3.2.1.1] | AMY | |
| ctg3960_g1 | 6.87 | 6.38 | 2.19 | 1.84 | 1.69059 | 1.47 × 10−14 | up-regulated | K01225 | cellulose 1,4-beta-cellobiosidase [EC:3.2.1.91] | CBH1 | 2.29/1 |
| ctg10469_g1 | 15.91 | 16.43 | 7.61 | 5.71 | 1.34346 | 9.80 × 10−19 | up-regulated | K19668 | cellulose 1,4-beta-cellobiosidase [EC:3.2.1.91] | CBH2 | |
| ctg7134_g2 | 5.73 | 6.07 | 0.43 | 4.8 | 2.21162 | 1.50 × 10−5 | up-regulated | K19668 | cellulose 1,4-beta-cellobiosidase [EC:3.2.1.91] | CBH2 | 1.30/1 |
| ctg7443_g2 | 4.9 | 3.55 | 1.59 | 0.74 | 1.79673 | 8.55 × 10−2 | up-regulated | K19356 | lytic cellulose monooxygenase (C1-hydroxylating) [EC:1.14.99.54] | E1.14.99.54 | 1.44/1 |
| ctg2646_g1 | 3.45 | 2.9 | 0.18 | 0 | 4.69409 | 6.75 × 10−8 | up-regulated | K19356 | lytic cellulose monooxygenase (C1-hydroxylating) [EC:1.14.99.54] | E1.14.99.54 | 2.43/1 |
| ctg2256_g1 | 0 | 0 | 2.88 | 2 | −5.995 | 1.02 × 10−23 | down-regulated | K19356 | lytic cellulose monooxygenase (C1-hydroxylating) [EC:1.14.99.54] | E1.14.99.54 | |
| ctg10311_g1 | 2.74 | 3.01 | 17.87 | 17.93 | −2.6469 | 6.07 × 10−55 | down-regulated | K01187 | alpha-glucosidase [EC:3.2.1.20] | malZ | |
| ctg7514_g1 | 3.49 | 1.64 | 1.08 | 1.04 | 1.31816 | 4.9 × 10−2 | up-regulated | K05349 | beta-glucosidase [EC:3.2.1.21] | bglX | 1.86/1 |
| ctg5449_g1 | 1.38 | 1.25 | 3.45 | 2.69 | −1.241 | 1.35 × 10−3 | down-regulated | K05349 | beta-glucosidase [EC:3.2.1.21] | bglX | |
| ctg1206_g1 | 1.69 | 0 | 3.87 | 8.25 | −2.6145 | 1.35 × 10−2 | down-regulated | K05349 | beta-glucosidase [EC:3.2.1.21] | bglX | |
| ctg1654_g1 | 0.85 | 0 | 2.33 | 2.26 | −2.2696 | 4.85 × 10−2 | down-regulated | K05349 | beta-glucosidase [EC:3.2.1.21] | bglX | |
| ctg2108_g1 | 0 | 1.01 | 2.4 | 3.48 | −2.4362 | 4.4 × 10−2 | down-regulated | K05349 | beta-glucosidase [EC:3.2.1.21] | bglX | |
| ctg9569_g1 | 5.18 | 5.16 | 11.9 | 12.34 | −1.2461 | 5.84 × 10−26 | down-regulated | K05349 | beta-glucosidase [EC:3.2.1.21] | bglX | |
| ctg6015_g3 | 1.97 | 1.19 | 5.98 | 6.25 | −1.9476 | 2.98 × 10−11 | down-regulated | K01188 | beta-glucosidase [EC:3.2.1.21] | E3.2.1.21 | |
| ctg7541_g1 | 5.74 | 5.46 | 15.47 | 18.25 | −1.5991 | 3.44 × 10−32 | down-regulated | K01188 | beta-glucosidase [EC:3.2.1.21] | E3.2.1.21 | |
| ctg9797_g3 | 391.26 | 395.33 | 0.1 | 0 | 12.679 | 0 | up-regulated | K01194 | alpha,alpha-trehalase [EC:3.2.1.28] | TREH | 1.59/1 |
| ctg1603_g1 | 0 | 0.62 | 5.59 | 7.89 | −4.1037 | 2.06 × 10−5 | down-regulated | K15920 | beta-D-xylosidase 4 [EC:3.2.1.37] | XYL4 | |
| Unigene ID | Expression | KEGG | MAPK–Yeast Pathway | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| v9-1 | v9-2 | v26-1 | v26-2 | Log2FC | FDR | Status | KO | Description | Gene Name | ||
| ctg12452_g1 | 2.42 | 4.83 | 0 | 0 | 5.237512 | 1.08 × 10−2 | up-regulated | K02218 | casein kinase 1 [EC:2.7.11.1] | CSNK1 | pheromone to mating |
| ctg12155_g1 | 38.43 | 38.24 | 226.93 | 221.38 | −2.55246 | 1.56 × 10−247 | down-regulated | K04627 | pheromone a factor receptor | STE3 | |
| ctg11329_g1 | 16.3 | 14.63 | 0.91 | 1.3 | 3.79796 | 1.08 × 10−51 | up-regulated | K19838 | GTPase-activating protein SST2 | SST2 | |
| ctg7267_g1 | 10 | 8.02 | 4.22 | 2.29 | 1.629585 | 1.66 × 10−15 | up-regulated | K19860 | guanine nucleotide-binding protein alpha-1 subunit | GPA1 | |
| ctg9995_g1 | 13.75 | 15.65 | 4.19 | 4.95 | 2.564613 | 2.47 × 10−97 | up-regulated | ||||
| ctg7524_g1 | 15.49 | 16.67 | 2.44 | 2.49 | 2.650679 | 2.99 × 10−13 | up-regulated | ||||
| ctg7524_g2 | 10.46 | 11.13 | 1.34 | 1.17 | 3.036185 | 2.17 × 10−10 | up-regulated | ||||
| ctg12751_g1 | 3.75 | 2.25 | 0 | 0 | 5.071841 | 1.86 × 10−2 | up-regulated | K07973 | guanine nucleotide-binding protein subunit gamma | GNG | |
| ctg8861_g1 | 83.56 | 78.85 | 24.95 | 28.65 | 1.378415 | 3.00 × 10−76 | up-regulated | K19839 | Rho-type GTPase-activating protein 1/2 | RGA1_2 | |
| ctg7642_g1 | 6.45 | 6.09 | 34.3 | 36.89 | −2.33243 | 3.27 × 10−101 | down-regulated | ||||
| ctg4782_g1 | 2.79 | 2.23 | 7.26 | 5.49 | −1.3039 | 4.97 × 10−3 | down-regulated | K04393 | cell division control protein 42 | CDC42 | |
| ctg3200_g1 | 7.21 | 5.98 | 1.42 | 1.82 | 2.014258 | 1.20 × 10−20 | up-regulated | K11237 | bud emergence protein 1 | BEM1 | |
| ctg2668_g1 | 2.6 | 4.31 | 0 | 0 | 5.071842 | 1.86 × 10−2 | up-regulated | K04409 | p21-activated kinase 1 [EC:2.7.11.1] | PAK1 | |
| ctg7848_g1 | 8 | 6.52 | 2.17 | 1.32 | 1.930004 | 2.49 × 10−7 | up-regulated | K19833 | serine/threonine-protein kinase CLA4 [EC:2.7.11.1] | CLA4 | |
| ctg8443_g1 | 40.03 | 38.71 | 16.05 | 20.1 | 1.123039 | 2.04 × 10−16 | up-regulated | K19842 | RHO1 GDP-GTP exchange protein 1/2 | ROM1_2 | cell wall stress to remodeling |
| ctg6770_g1 | 31.02 | 33.24 | 0.88 | 1.03 | 5.014033 | 1.57 × 10−33 | up-regulated | ||||
| ctg9497_g3 | 4.38 | 3.34 | 1.49 | 1.15 | 1.517745 | 1.26 × 10−2 | up-regulated | ||||
| ctg293_g1 | 1.57 | 3.72 | 0.95 | 0.27 | 2.007466 | 1.79 × 10−2 | up-regulated | K19844 | GTPase-activating protein BEM2 | BEM2 | |
| ctg4687_g1 | 3.52 | 2.64 | 6.67 | 7.29 | −1.19529 | 6.18 × 10−24 | down-regulated | K00888 | phosphatidylinositol 4-kinase A [EC:2.7.1.67] | PI4KA | |
| ctg5599_g1 | 4.81 | 4.57 | 20.91 | 18.6 | −2.09223 | 6.61 × 10−47 | down-regulated | K11229 | mitogen-activated protein kinase kinase kinase [EC:2.7.11.25] | BCK1 | |
| ctg5657_g4 | 0.92 | 0 | 3.77 | 2.93 | −2.70834 | 6.18 × 10−2 | down-regulated | K19806 | tyrosine-protein phosphatase 2/3 [EC:3.1.3.48] | PTP2_3 | |
| ctg380_g1 | 0.62 | 0.62 | 3.93 | 4.62 | −2.61052 | 1.05 × 10−2 | down-regulated | K06276 | 3-phosphoinositide dependent protein kinase-1 [EC:2.7.11.1] | Pkh1 | |
| ctg8570_g3 | 3.61 | 3.22 | 1.16 | 0.84 | 1.729908 | 5.36 × 10−6 | up-regulated | ||||
| ctg1020_g1 | 11.05 | 11.08 | 4.74 | 5.05 | 1.171861 | 2.94 × 10−6 | up-regulated | K11246 | SHO1 osmosensor | SHO1 | high osmolarity to osmolyte synthesis |
| ctg9076_g2 | 2.22 | 1.33 | 5.92 | 5.44 | −1.68952 | 7.79 × 10−11 | down-regulated | K11233 | osomolarity two-component system, response regulator SSK1 | SSK1 | |
| ctg10627_g1 | 79.42 | 69.73 | 33.43 | 35.06 | 1.139585 | 3.06 × 10−71 | up-regulated | ||||
| ctg5925_g2 | 3.98 | 4.16 | 0.56 | 0.25 | 3.106293 | 7.58 × 10−7 | up-regulated | K06666 | general transcriptional corepressor TUP1 | TUP1 | |
| ctg8863_g1 | 27.22 | 28.43 | 5.57 | 9.01 | 2.427738 | 4.45 × 10−73 | up-regulated | K03114 | mitosis inhibitor protein kinase SWE1 [EC:2.7.11.1] | SWE1 | |
| ctg8944_g1 | 60.33 | 48.42 | 125.54 | 116.41 | −1.17943 | 7.93 × 10−52 | down-regulated | K03781 | catalase [EC:1.11.1.6] | Ctt1 | |
| ctg5701_g1 | 43.23 | 42.24 | 423.49 | 411.39 | −3.30667 | 0 | down-regulated | K09448 | transcriptional enhancer factor | TEAD | starvation to filamentation |
| ctg6107_g1 | 22.31 | 23.76 | 4.29 | 2.92 | 2.639995 | 1.36 × 10−46 | up-regulated | ||||
| Unigene ID | Expression | KEGG | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| v9-1 | v9-2 | v26-1 | v26-2 | Log 2-FoldChange | FDR | Status | KO | Description | Gene Name | |
| ctg2182_g1 | 547.5 | 525.1 | 237.3 | 243 | 1.139132 | 4.86 × 10−202 | up-regulated | K05658 | ATP-binding cassette, subfamily B (MDR/TAP), member 1 (EC:3.6.3.44) xenobiotic-exporting ATPase | ABCB1 |
| ctg11186_g1 | 266.3 | 259.7 | 57.76 | 60.5 | 2.134296 | 0 | up-regulated | |||
| ctg11186_g2 | 274.3 | 252.4 | 50.93 | 61.7 | 2.235691 | 2.83 × 10−179 | up-regulated | |||
| ctg10041_g1 | 53.11 | 54.03 | 18.96 | 19.3 | 1.450944 | 1.56 × 10−152 | up-regulated | |||
| ctg11158_g1 | 29.92 | 26.85 | 11.12 | 11.8 | 1.290507 | 5.02 × 10−79 | up-regulated | |||
| ctg1141_g1 | 2.79 | 5.54 | 0 | 0 | 5.237513 | 1.08 × 10−2 | up-regulated | |||
| ctg3266_g2 | 1.73 | 0.58 | 3.16 | 8.03 | −2.13418 | 1.2 × 10−2 | down-regulated | |||
| ctg4538_g1 | 0.81 | 0.41 | 4.22 | 4.44 | −2.76898 | 2.43 × 10−7 | down-regulated | |||
| ctg8579_g1 | 111.7 | 102.4 | 289.9 | 290 | −1.45923 | 3.27 × 10−141 | down-regulated | |||
| ctg11112_g1 | 30.84 | 25.83 | 71.78 | 75.5 | −1.4194 | 2.98 × 10−157 | down-regulated | |||
| ctg7752_g1 | 26.95 | 26.32 | 53.58 | 54.9 | −1.05833 | 5.22 × 10−67 | down-regulated | |||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, M.; Yu, T.; Singh, P.K.; Liu, Q.; Liu, H.; Zhu, Q.; Xiao, Z.; Xu, J.; Peng, Y.; Fu, S.; et al. A Comparative Transcriptome Analysis of Volvariella volvacea Identified the Candidate Genes Involved in Fast Growth at the Mycelial Growth Stage. Genes 2020, 11, 161. https://doi.org/10.3390/genes11020161
Liu M, Yu T, Singh PK, Liu Q, Liu H, Zhu Q, Xiao Z, Xu J, Peng Y, Fu S, et al. A Comparative Transcriptome Analysis of Volvariella volvacea Identified the Candidate Genes Involved in Fast Growth at the Mycelial Growth Stage. Genes. 2020; 11(2):161. https://doi.org/10.3390/genes11020161
Chicago/Turabian StyleLiu, Ming, Ting Yu, Puneet Kumar Singh, Qinjian Liu, Hao Liu, Qingfeng Zhu, Zitian Xiao, Jiang Xu, Yangyang Peng, Shiyu Fu, and et al. 2020. "A Comparative Transcriptome Analysis of Volvariella volvacea Identified the Candidate Genes Involved in Fast Growth at the Mycelial Growth Stage" Genes 11, no. 2: 161. https://doi.org/10.3390/genes11020161