
Milk Bioactive Compounds and Gut Microbiota Modulation: The Role of Whey Proteins and Milk Oligosaccharides

1
Department of Science, Roma Tre University, Viale Guglielmo Marconi 446, 00146 Rome, Italy
2
National Institute of Biostructures and Biosystems (INBB), Viale delle Medaglie d’Oro 305, 00136 Rome, Italy
*
Author to whom correspondence should be addressed.
Foods 2024, 13(6), 907; https://doi.org/10.3390/foods13060907
Submission received: 12 February 2024
/
Revised: 13 March 2024
/
Accepted: 14 March 2024
/
Published: 16 March 2024
(This article belongs to the Section Nutraceuticals, Functional Foods, and Novel Foods)
Abstract
:A strong correlation between the occurrence of various pathological conditions and intestinal dysbiosis is supported by a range of strong evidence. Vice versa, many pathologies have been shown, in turn, to be responsible for alterations in the gut microbiota, a condition that can worsen illness outcomes and response to therapies. For these reasons, great efforts have been made, and studies are still ongoing, to elucidate the mechanisms underlying gut microbiota alterations and to search for pharmacologic or other strategies that can effectively restore the gut microbiota. In this narrative review, we examined the most significant literature on the role of some milk bioactive compounds, such as milk oligosaccharides and whey proteins, in modulating the composition of the gut microbiota and the underlying mechanisms of action, with the aim of investigating the impact of the microbiota changes mediated by these milk bioactive molecules on human health, and their potential use as therapeutics to treat or adjuvate the treatment of gut dysbiosis and associated pathologies.
1. Introduction
The term microbiota defines the entire microbial community (including commensal, mutualistic or pathogenic) that inhabit a given habitat; in this sense, gut microbiota refers to the range of microorganisms (i.e., Archea, Bacteria, Eukarya, and viruses) which reside in the gastrointestinal (GI) tract. Billions of symbiotic microorganisms populate the human gut, presiding over structural [1,2], protective [3,4,5], neurological, and metabolic functions [6,7,8], within such an intimate and intricate relationship that it gives rise to a unit called the holobiont [9]. The gut microbiota is acquired at birth and is characterized by great instability in terms of structure and composition throughout childhood [10]. After childhood, the gut microbiota, still maintaining very dynamic behaviour, begins to differentiate into a more defined structure [11,12,13]. Indeed, adult gut microbiota is characterized by a quasi-constant composition of those microbial species which are more favoured in the gut microenvironment [14,15]. For example, the most represented gut microbiota bacteria belong to Firmicutes, Bacteroidetes, Actinobacteria, Fusobacteria, Proteobacteria, Verrucomicrobia, and Cyanobacteria phyla, where Firmicutes and Bacteroidetes constitute more than 90% [16]. It is noteworthy that the Firmicutes/Bacteroidetes ratio plays a key role in influencing intestinal homeostasis. Indeed, variation in the Firmicutes/Bacteroides ratio is strongly associated with gut dysbiosis and associated diseases, including obesity and bowel inflammatory disorders [17]. However, the relative abundance of bacterial phyla and/or colonisation by other microorganisms may vary greatly depending on all the factors that can influence the gut microenvironment parameters (e.g., pH, oxygen levels, temperature, availability of nutrients). These factors include dietary habits [18], age [19], pharmacological therapies (e.g., antibiotics) [20,21,22], host genetics [23,24], host geographic location [25], pathologies, lifestyle, and environmental stress [26,27], and account for high intra-individual time-related variations and for inter-individual diversification [28,29,30]. Compared to the microbiota which colonise other body sites (e.g., oral mucosa, vagina, and skin), gut microbiota is the major focus of researchers’ interest due to its higher clinical significance [31]. Indeed, studies have shown that the equilibrium between health status and the arising of a disease state is strongly influenced by the composition of microbiota, particularly by the equilibrium between the different species and the predominance of some of them [32]. Despite the difficulties in defining a standard health promoting microbiota composition, due to the great variability related to host and environmental factors, several studies report a strong correlation between gut dysbiosis, which leads to a decrease in microbial diversity, and the occurrence of various pathological conditions [33,34,35]. These include inflammatory and metabolic disorders, obesity, and type II diabetes [7,36], but gut dysbiosis has also been associated with the promotion of some types of cancer, including colorectal cancer [37,38], as well as neurological and neuropsychiatric disorders, including Alzheimer’s disease [39] and major depression [40,41], through the impairment of the gut-brain axis homeostasis [42]. Conversely, pathologies are often associated with the development of gut dysbiosis, which exacerbates the consequences of disease [43].
Great efforts to define the factors affecting the highly dynamic composition of the human gut microbiota and to better understand its role in health and disease have been made and are ongoing [44]. Based on these, therapeutic interventions aimed at restoring the establishment of a health-promoting gut microbiota could be useful for the prevention and treatment of gut dysbiosis. Diverse strategies, ranging from the use of probiotics, prebiotics, and pharmaceutical compounds to non-pharmacological approaches, such as faecal microbiota transplantation, have been shown to be effective in positively modulating the microbiota [30,45,46,47,48]. Importantly, one area that has gained increasing interest in recent years is the role of prebiotics in these processes. Prebiotics are a non-digestible group of molecules derived from food sources that are selectively fermented by probiotics, health beneficial gut microorganisms (e.g., Bifidobacterium and Lactobacillus genera), stimulating their growth and inducing beneficial changes in the gut microbiota composition; importantly, their effects on maintaining an optimal Firmicutes/Bacteroides ratio are well documented [49].
In this context, milk and its bioactive components have long been studied for their potential as prebiotics. Indeed, due to its content of bioactive molecules [50], including proteins, lipids, and oligosaccharides, milk is considered as a functional food whose antimicrobial [51], immunological [52], and antitumorigenic activities have been widely studied and exploited in nutraceutical and biomedical fields to support the treatment and prevention of diseases [50,53,54,55,56,57,58].
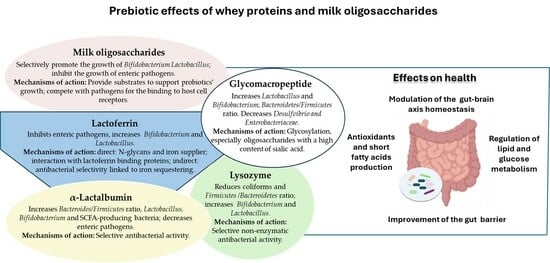
Infant gut microbiota develops at birth and breast milk plays a key role in this process [59]. Several studies have linked changes in dietary habits during pregnancy to alterations in the maternal and in the infant gut microbiota [60,61], and a marked difference between gut microbiota of breast-fed and formula-fed infants has been demonstrated in metagenomic studies [11], which indicate that the breast-fed infants’ microbiota is less diversified but consists mainly of health-beneficial bacteria such as bifidobacteria, lactobacilli, and staphylococci [62,63,64]. Apart from the role exerted by its own microbiota, which contributes to a healthy microbial diversification of the gut microbiota of newborns [65,66], the bioactive components of milk, especially oligosaccharides and whey proteins such as lactoferrin, lysozyme, and α-lactalbumin, have been shown to play a crucial role in shaping the gut microbiota from birth to adulthood [67,68]. Indeed, studies have reported the role of human and cow’s milk constituents in promoting the growth of beneficial probiotic bacteria, including B. infantis, B. Pennsylvanicus, B. longum, B. bifidum, and B. breve [69,70,71,72].
In this work, we reviewed the most significant literature on the role of milk bioactive compounds in influencing and modulating the composition of gut microbiota in newborns and throughout life, focusing on oligosaccharides and whey proteins. We have also evaluated the impact of the microbiota changes mediated by these milk bioactive molecules on health. This article aims to provide and spread up-to-date knowledge on this promising and evolving field of research and to raise awareness of the main current limitations. This may be helpful to stimulate further research aimed at improving our understanding of the prebiotic effects of these molecules, of the underlying mechanisms of action and of their potential beneficial effects on health, and to open the possibility of using these molecules, alone or in synergy with other drugs, as therapeutics to treat and/or adjuvate the treatment of gut dysbiosis and related diseases.
2. Prebiotic Activity of Milk Oligosaccharides
2.1. Mechanisms of Action
Milk consists of three main structural classes of oligosaccharides: fucosylated, non-fucosylated neutral, and sialylated, which are composed of five sugar residues with a variable degree of polymerization. Human milk has a higher content of oligosaccharides than cow’s milk, which is about 20-fold lower; furthermore, fucosylated and neutral species are predominant in human milk, whereas sialylated species are most abundant in cow’s milk [73,74,75]. Two main mechanisms have been associated with the modulation of the gut microbiota by milk oligosaccharides (MOs) [76]. The first mechanism is related to a direct prebiotic activity of MOs, which have been shown to selectively promote the growth of certain Bifidobacterium strains, including B. infantis, B. breve, and B. bifidum, as well as some Lactobacillus species [77,78,79,80,81]. Furthermore, in vitro studies have shown that structural differences in MOs account for their selective utilization by gut microbiota bacteria [82,83]. The second mechanism is related to the inhibition of colonisation by various enteric pathogens, including Vibrio cholerae, Salmonella fyris, Campylobactor jejuni, Clostridioides difficile, and various Escherichia coli strains, by preventing epithelial adhesion (Table 1). Indeed, studies demonstrated the capability of MOs, especially those belonging to the fucosylated class, to compete with pathogens in binding to host cell receptors and to sequester pathogens by acting as free analogues of host cell receptors [84,85,86,87].
2.2. Effects of MOs in Modulating Gut Microbiota and Health: In Vitro Studies and Clinical Trials
In addition to a protective role against diverse enteric pathogens, studies suggest that the effects of MOs on gut microbiota can lead to a modulation of the immune system, reducing the risk of allergies, asthma, and inflammatory diseases [88]. Indeed, a deficiency in bifidobacteria, and thus in the genes required for the metabolism of MOs, has been shown to correlate with systemic inflammation and immune system dysregulation in infants [89,90,91]. Furthermore, MOs have been demonstrated to be important substrates for short-chain fatty acids production, particularly butyrate, which is mediated by an interplay between bifidobacteria and butyrate-producing bacteria [92,93]. This has deep health implications, since butyrate plays a critical role in maintaining the gut mucosal barrier and modulating the immune system of infants and adults. In addition, the bifidogenic effect of MOs is important for the production of other health-promoting metabolites, including the neurotransmitter GABA [94] and indole-3-lactic acid, which is involved in the activation of the aryl hydrocarbon receptor, a regulator of the gut-brain axis, intestinal homeostasis, and immune response [95]. However, in adults, the levels of bifidobacteria decrease greatly (up to 90%) and this has been demonstrated to be related to the insurgence of various diseases and GI disorders, including those associated with changes in intestinal permeability (leaky gut). In this context, in vitro studies and clinical trials to evaluate the effects of MOs in modulating adult gut microbiota have been made in an attempt to exploit these molecules as therapeutics for the restoration of a healthier gut [96,97,98]. In vitro studies by Šuligoj and colleagues investigated the effects of human MOs (HMOs) on the gut microbiota using Caco2 cell lines, human intestinal gut organoid-on-chips, and the Simulator of the Human Intestinal Microbial Ecosystem (SHIME®) as models. This study showed that the treatment with 2′-O-fucosyllactose (2′FL) and/or lacto-N-neotetraose (LNnT) fermented HMOs led to a significant upregulation of cytokines, permeability, and tight junction proteins, including claudin-8 and claudin-5, suggesting a role of HMOs fermented by gut microbiota in positively promoting gut barrier functionality [99]. Elison et al. performed a clinical trial based on oral supplementation of HMOs in 100 healthy, adult volunteers, providing sets of data on safety, tolerance, and impact of HMOs on the adult gut microbiota. In this study, chemically produced 2′FL and/or LNnT at various daily doses and mixes, or placebo, were administered for 2 weeks. The obtained results showed that the uptake of 2′FL and LNnT is sufficient to modulate the adult microbiota within two weeks, with an increase in relative abundance of bifidobacteria, to >25% in some individuals, and a reduction in the relative abundance of Firmicutes and Proteobacteria phyla [100]. In another study, the effects of oral supplementation of 2′-O-fucosyllactose and lacto-N-neotetraose HMOs on gut microbiota were evaluated in patients with irritable bowel syndrome; a modulation of the gut microbiota, and of faecal and plasma metabolite profiles, associated with the HMOs bifidogenic effect, was observed [101].
All these studies highlight the beneficial effects of HMOs beyond childhood, suggesting their potential in supporting the restoration of a wholesome adult gut microbiota. In this context, interesting results came from a more recent study which investigated the age-dependent impact of HMOs on gut microbiota. In this study, 6-year-old children and adults were enrolled for ex vivo testing using systemic intestinal fermentation (SIFR®) technology. Single types of HMOs, including 2′FL, LNnT, 3′Sialyllactose, 6′Sialyllactose, and combinations of these were used for this study. The results showed that age-dependent differences in microbiota composition, especially in terms of Bifidobacterium strains, strongly impact the utilization of HMOs and the individual response to the treatment, suggesting that the development of age-specific nutritional supplements using HMOs could maximize their beneficial outcomes [102].

{kind=link}
{kind=link}
Table 1.
Prebiotic activity of milk oligosaccharides (MOs), mechanisms of action and potential health implications.
Table 1.
Prebiotic activity of milk oligosaccharides (MOs), mechanisms of action and potential health implications.
| Activity | Mechanism of Action | MOs | Effects on Gut Microbiota | Potential Health Implications | Ref. |
|---|---|---|---|---|---|
| Direct prebiotic activity | Selective utilization by the bacteria of the gut microbiota of structurally different classes of MOs. | HMOs (2′-O-fucosyllactose; Lacto-N-neotetraose); sialic acid. | Promotes the growth of probiotics, including Bifidobacterium (i.e., B. infantis, B. breve, B. bifidum) and some Lactobacillus species. | Modulation of the immune system, reducing the risk of allergies, asthma, and systemic inflammatory conditions. | [78,79,80,82,83,89,90,91] |
| MOs provide substrates which support the metabolism of diverse probiotics, especially Bifidobacterium, thus stimulating the production of health beneficial metabolites. | Promote the production of the neurotransmitter GABA and indole-3-lactic acid, a regulator of gut-brain axis by Bifidobacterium (B. adolescentis). | Modulation of the gut-brain axis response, intestinal homeostasis and immune response. | [69,94] | ||
| Promote the production of short-chain fatty acids, especially butyrate, mediated by an interplay between bifidobacteria and butyrate-producing bacteria. | Maintenance of the gut mucosal barrier by limiting intestinal permeability (leaky gut); modulation of the immune system. | [92,93,99] | |||
| Indirect prebiotic activity | MOs, especially fucosylated classes, compete with pathogens in binding to host cells by acting as free analogues of cell receptors. | Prevent epithelial adhesion thus inhibiting colonisation by diverse enteric pathogens (i.e., V. cholerae, S. fyris, C. jejuni, C. difficile and E. coli). | Protective role against enteric diseases. | [84,85,86,87] |
3. Prebiotic Activity of Whey Proteins
Milk contains two main classes of proteins: caseins and whey proteins. Human milk’s whey proteins include α-lactalbumin, immunoglobulins, serum albumin, lactoferrin, glycomacropeptide, lactoperoxidase, and lysozyme [103]. Bovine milk differs from human milk in the presence of β-lactoglobulin and lower levels of α-lactalbumin [104].
Whey proteins exert several functions including antioxidant, anticancer, antimicrobial, anti-inflammatory, and immunomodulatory [105,106,107,108,109,110,111]. Furthermore, evidence suggests a beneficial role of whey proteins in positively modulating the gut microbiota in both infants and adults [112,113,114].
3.1. Lactoferrin
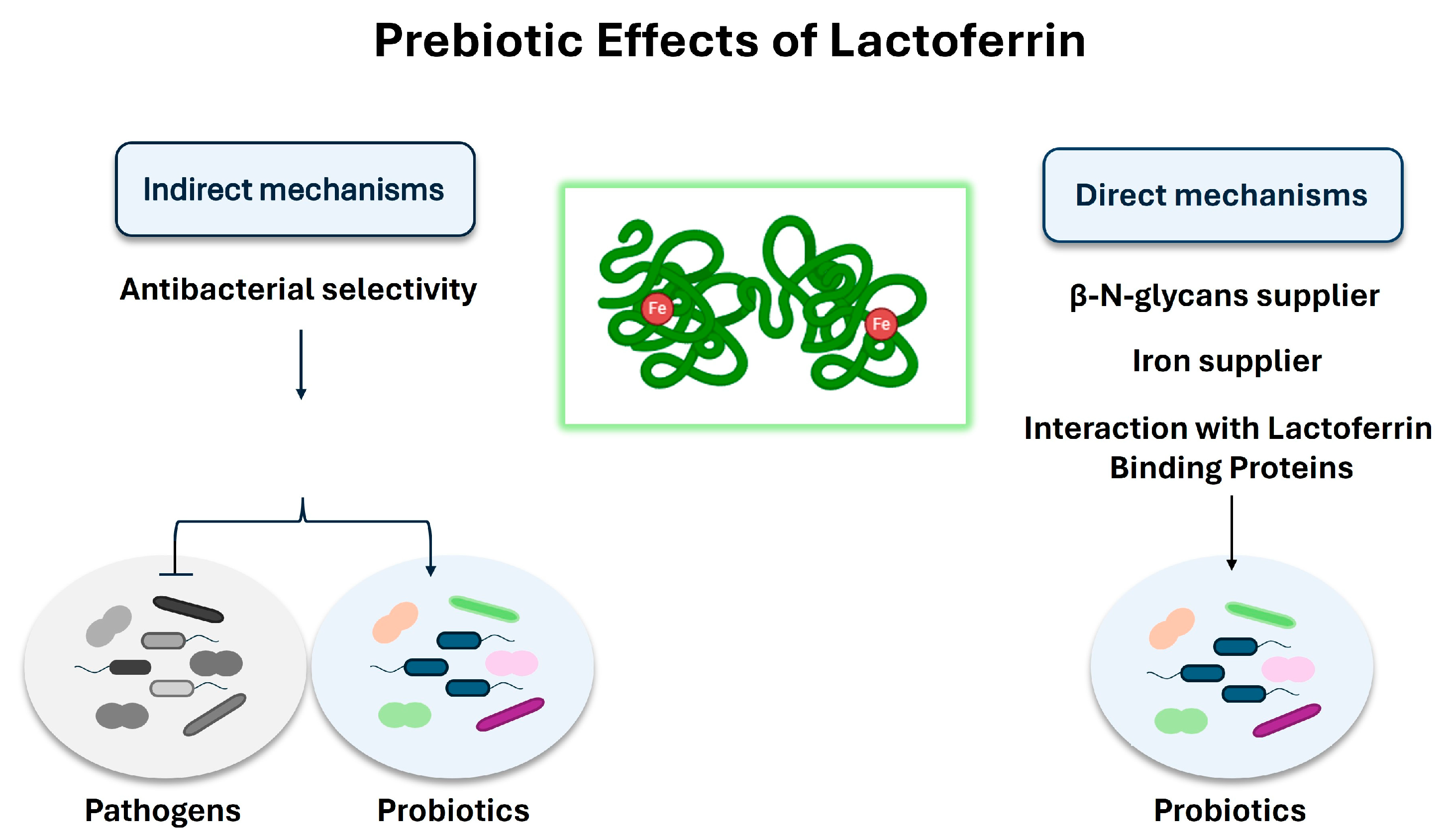
Lactoferrin is an 80 kDa positively charged iron-binding glycoprotein, that belongs to the transferrin superfamily and is present at high levels in various biological fluids, especially in milk and colostrum [115,116]. Depending on the degree of iron saturation, it exists in two forms: the iron-depleted (apo) and the iron-saturated (holo). Both the apo and holo forms of lactoferrin and its peptide derivatives, including lactoferricin, play an important role in promoting and maintaining a functional gut microbiota and in inhibiting gut barrier impairment [117,118,119,120]. Experimental evidence suggests that these activities are mainly due to its strong antimicrobial activity, including bacteriostatic (related to the iron sequestering function) and bactericidal (linked to the binding and the consequent neutralization of some anionic bacterial surface components, such as lipopolysaccharide), as well as immunomodulatory functions [121,122,123], which help to preserve the integrity of the gut barrier [124,125].
3.1.1. In Vitro Studies
Various in vitro studies suggest a prebiotic role of lactoferrin and its peptide derivative, lactoferricin, on the gut microbiota [126,127] (Figure 1). Though the mechanisms still need to be better clarified, indirect prebiotic effects have been observed, mainly related to a selective inhibition of bacterial growth. Indeed, some studies have shown that instead of directly promoting the growth of probiotics (i.e., Bifidobacteria and Lactobacilli) [128], lactoferrin acts mainly by inhibiting the growth of pathogens, which are less resistant to its antibacterial activity [129]. Lactoferrin-mediated iron sequestering could be one explanation for this antibacterial selectivity; indeed, contrary to many pathogens, such as E. coli, various probiotics, including many Bifidobacterial strains, do not necessarily need iron to grow and are able to predominate over pathogens under conditions of iron deprivation [130,131,132]. However, various studies have also shown direct prebiotic effects; in this context, different mechanisms of action have been postulated from the results of in vitro studies. Some studies suggest a role for lactoferrin as a supplier of β-N-glycans to stimulate bifidobacterial growth [133], while other in vitro studies have shown that the holo form of lactoferrin promotes the growth of Bifidobacterium breve by acting as an iron supplier [134]. In addition, some studies reported the presence of lactoferrin-binding proteins on the surface of some probiotics which are sensitive to the prebiotic action of lactoferrin, suggesting that the interaction between the cationic surface of lactoferrin and some anionic cell surface components may be involved in mediating this process [135,136]. Indeed, studies have shown that these interactions favour the cytosolic internalization of lactoferrin which, in the presence of ATP, dissociates to the N-lobe and C-lobe [137]; it has been suggested that the internalization of the C-lobe in the nucleus mediates the modulation of genes involved in DNA replication and cell growth [127].
However, regardless of the mechanisms involved, the results obtained from testing different experimental conditions indicated that the direct prebiotic activity of lactoferrin may vary depending on factors such as bacterial strain, temperature, oxygen levels, lactoferrin iron-saturation levels, and dosage [138,139,140,141,142]. Chen et al., while exploring in vitro the prebiotic potential of bovine lactoferrin in 14 probiotics, including B. breve, L. coryniformis, L. delbrueckii, L. acidophilus, B. angulatum, B. catenulatum, and L. paraplantarum, observed a negligible effect on probiotic growth at 37 °C, but a strong lactoferrin-mediated dose-dependent restart of growth at lower temperatures (i.e., 22 °C), suggesting that the direct prebiotic effects of lactoferrin become significant when the probiotic growth is retarded by cold stress conditions [143]. These findings could be of great significance, especially in all those environmental conditions or clinical features which cause probiotic growth inhibition or retardation; however, further studies, including in vivo studies and clinical trials, are needed to translate these results into a clinically relevant context. More recently, the same research group investigated the transcriptome profiles of Lactobacillus rhamnosus GG, treated with 1 mg/mL bovine lactoferrin at 22 °C, to elucidate the molecular mechanism involved in the prebiotic effect of lactoferrin on L. rhamnosus GG. The results showed that lactoferrin supplementation is associated with the transcriptional modulation of several genes that are involved in many central metabolic pathways. Increased transcription levels have been observed for genes encoding transporter proteins, including ABC transporter permease and ABC transported related proteins; genes involved in amino acid synthesis, DNA replication, and peptidoglycan biosynthesis; and genes encoding for proteins involved in carbohydrate metabolism and stress responses, including CspA, LytR, and XRE. On the contrary, decreased transcription levels have been found for some genes involved in purine and pyrimidine metabolism, biosynthesis of antibiotics, and secondary metabolites. However, since the prebiotic activity of lactoferrin is strongly strain-dependent, further studies on other probiotic strains could be useful to better understand the underlying mechanisms [144].
3.1.2. In Vivo Studies and Clinical Trials
Several in vivo studies and clinical trials have investigated the effects of lactoferrin supplementation on the gut microbiota and its potential health-promoting effects (Table 2).
Many in vivo studies have been conducted using mice as animal models, documenting the role of lactoferrin supplementation in the positive modulation of the gut microbiota [120]. In experiments on vitamin D deficient mice, Wang and colleagues showed that oral administration of 100 to 1000 mg/kg body weight (BW) of bovine lactoferrin induced a significant change in the gut microbial community, favouring the growth of lactobacillus through a direct mechanism [145]. In another study, performed in high-fat diet-induced obese mice, the oral administration of 100 mg/kg BW of bovine lactoferrin for 12 weeks was shown to induce a significant increase of Bifidobacterium spp. in the faeces and a decrease of Enterobacterales, compared to controls. Furthermore, this treatment was shown to reduce inflammation and regulate lipid and glucose metabolism, highlighting the potential of bovine lactoferrin to improve microbiota dysbiosis and associated health conditions induced by a high-fat diet [146]. The in vivo prebiotic activity of lactoferrin has been demonstrated in other similar studies, which suggest a correlation between the lactoferrin-mediated improvement of dysbiosis and the amelioration of diverse pathological conditions, including obesity, inflammation, and metabolic disorders [125,147,148]. Studies also showed the potential of native and iron-saturated bovine lactoferrin in restoring the normal levels of some anti-inflammatory bacteria (i.e., Bacteroidaceae, Prevotellaceae and Rikenellaceae) in a mouse model of antibiotic-induced dysbiosis [149]. In addition, a recent study demonstrated a role of lactoferrin in alleviating cognitive impairment in western-diet induced obese mice by improving the gut-microbiota-brain axis, mainly inducing an increase in Bacteroidetes (i.e., Roseburia), which led to an inhibition of microglial activation and neuroinflammation [150].
Several studies have also been performed on piglets, as they have a greater biological similarity to humans compared to rodents [120,127]. In a study conducted on weaning piglets, the supplementation of bovine lactoferrin increased the amount of Lactobacillus and Bifidobacterium and decreased the abundance of Escherichia coli in the cecum, leading to an associated improvement of intestinal immunity and the gut barrier [151]. In another study, piglets administered lactoferrin orally (0.5 g per kg body weight per day) showed increased levels of Lactobacillus and a lower abundance of Veillonella and Escherichia-Shigella in the jejunum, and of Actinobacillus in the ileum [152]. Other similar studies confirmed the role of bovine and human native full-length lactoferrin in promoting a health beneficial modulation of the gut microbiota [153,154]. In one of these studies, healthy full-term piglets were employed as newborn models and administered with a combination of lactoferrin and probiotics to evaluate the impact of this treatment on the development of gut microbiota. Interestingly, results showed a decrease in the abundance of Enterobacteriaceae (frequently involved in neonatal infections and sepsis), and an increase in the levels of Erysipelotrichaceae and Veillonellaceae taxa and of butyrate-producing bacteria (i.e., Faecalibacterium prausnitzii), compared to controls [155]. In addition to studies that used native full-length lactoferrin, interesting results came from a study in which a recombinant fusion peptide constituted by the two main lactoferrin peptide derivatives, namely lactoferricin and lactoferrampin, was used as a dietary supplement; results showed that weaned piglets administered with 0.1 g/kg of recombinant fusion peptide increased the amount of Lactobacillus and Bifidobacterium in the chyme of the stomach, duodenum, jejunum, ileum, colon, and caecum [156]. Importantly, in all these studies, a reduction in the diarrhoea rate and an enhancement in the piglets’ trends of growth, together with an increased intestinal cell proliferation and maturation, were observed, which were associated with the lactoferrin-mediated balancing of gut microbiota [157,158].
Several clinical trials have explored the potential of lactoferrin supplementation in both infant and adult gut microbiota and its beneficial consequences on health [159]. In a double-blind, placebo-controlled, randomized trial, conducted in VLBW neonates in Italy, bovine lactoferrin supplementation, alone or in combination with the probiotic Lactobacillus rhamnosus GG, was shown to reduce the incidence of a first episode of late-onset sepsis, a severe complication which frequently affects premature neonates, by affecting the fungal progression from colonisation to infection [160]. Similar results were obtained in another randomized controlled clinical trial performed in Peruvian neonates [161]. Interestingly, another clinical trial conducted in preterm infants showed the efficacy of a recombinant human lactoferrin (talactoferrin, TLf) in reducing infection [162]. Further evidence from randomized controlled trials on enteral lactoferrin supplementation in preterm neonates has been recently reviewed by Pammi et al. [163].
The prebiotic and gut microbiota modulating effects of lactoferrin have also been evaluated in both healthy individuals and those suffering from various pathologies or undergoing therapies. Indeed, pathogenic gut dysbiosis has been observed during diverse therapies, including anticancer chemotherapy [164], and various diseases, including cancer and HIV infection, and it has often been associated with disease progression [165,166]. Results from some clinical trials suggested that lactoferrin may provide an adjunctive therapy to accelerate the process of gut microbiota restoration, thereby improving health outcomes and the response to therapy.
In a double-blind, placebo-controlled clinical trial, the impact of oral supplementation of lactoferrin on gut microbiota was evaluated in paediatric oncological patients undergoing chemotherapy. Results showed that lactoferrin, besides being well tolerated, played a role in favouring the promotion of gut microbiota homeostasis and in exerting a protective role against Enterococcus and other pathobiont colonisations [167].
In a randomized, double-blind, crossover clinical trial design, the treatment effects of oral recombinant human lactoferrin (1500 mg twice daily) in immunodeficiency virus–infected participants undergoing antiretroviral therapy and enrolled in an intestinal microbiome study, were investigated. In this study, insignificant changes to the intestinal microbiota were observed, although there was remarkable stability in the microbial community over time [168].
Results from a double-blind, placebo-controlled study conducted in healthy elderly women who were orally supplemented with bovine lactoferrin showed a significant increase in the relative abundance of Holdemanella in the faecal microbiota. Furthermore, an increase in the relative abundance of Bifidobacterium was observed when lactoferrin was administered in combination with active galactooligosaccharides [169].
Table 2.
Lactoferrin’s effects on gut microbiota: in vivo studies and clinical trials.
| Patients or Animal Models | Treatment | Effects on Gut Microbiota | Health Effects | Type of Study | Ref. |
|---|---|---|---|---|---|
| Vitamin D deficient mice. | Oral administration of bovine lactoferrin (100 and 1000 mg/kg BW). | Reduces the abundance of Oscillibater; increases the proportion of Lachnospiraceae, Faecalibaculum, and Lactobacillus. | Stimulates the expression of vitamin-D receptor by regulating gut microbiota; reduces serum levels of pro-inflammatory cytokines; enhances intestinal barrier function. | In vivo | [145] |
| Mouse models of colitis induced by dextran sulphate sodium salt. | Oral administration of bovine lactoferrin (100 mg/kg). | Phylum level: decreases Bacteroidetes and Firmicutes; increases Verrucomicrobia. Family level: decreases Muribaculaceae and Lachnospiraceae; increases Akkermansiaceae. | Alleviates colitis by improving the inflammatory response and the structure of the colon barrier in the colon. | In vivo | [125] |
| High-fat diet induced obese C57BL/6J mice. | Oral administration of bovine lactoferrin (100 mg/kg BW). | Restores the abundance of Bifidobacterium spp. | Reduces inflammation and regulates lipid and glucose metabolism. | [146] | |
| Hight-fat diet induced obese C57BL/6J mice. | Oral supplementation of bovine lactoferrin (100 mg/kg BW) for 12 weeks. | Increases faecal Bifidobacterium spp.; decreases Enterobacteriales and Bacteroidetes. | Improvement of high-fat diet induced microbiota dysbiosis, improvement of hypercholesterolaemia and hyperglycaemia. | In vivo | [147,148] |
| C7BL-6 mice with clindamycin-induced dysbiosis. | Native bovine lactoferrin; iron-saturated bovine lactoferrin. | Promotes the growth of Bacterioidaceae, Prevoellaceae and Rikenellaceae. | Reverses clinamycin-induced dysbiosis. | In vivo | [149] |
| C57BL/6J mice models of western diet-induced cognitive impairment. | Oral supplementation of lactoferrin (50 mg/kg BW) for 16 weeks. | Increases Bacteroidetes (i.e., Roseburia). | Alleviates cognitive impairment by improving gut-microbiota-brain axis. | In vivo | [150] |
| Weaning piglets. | Oral supplementation of bovine lactoferrin (1 to 3 g/kg). | Increases Lactobacillus and Bifidobacterium; decreases the abundance of Escherichia coli in the cecum. | Enhances the growth performance; reduces diarrhoea rate by improving gut barrier and balancing intestinal microbiota. | In vivo | [151] |
| Suckling piglets. | Oral administration of lactoferrin (0.5 g/kg BW) daily for a week. | Increases Lactobacillus and decreases Veillonella and Escherichia-Shigella in the jejunum and Actinobacillus in the ileum. | Reduction of diarrhoea incidence and enhancement in the trends of growth by promoting the development of intestinal function and modulating the microbiota in the small intestine. | In vivo | [152,158] |
| Healthy full-term piglets. | 0.5 g of probiotic (FloraBABY) and 100 mg of bovine lactoferrin. | Reduces the abundance of taxa commonly associated with sepsis in pre-term human infants (Enterobacteriaceae); increases the levels of Erysipelotrichaceae and Veillonellaceae taxa; increases butyrate producers (i.e., Faecalibacterium prausnitzii). | Improvement of gut microbiota and gut barrier. | In vivo | [155] |
| Weaned piglets. | Diet supplementation with 0.1 g/kg recombinant bovine lactoferrampin-lactoferricin fusion peptide. | Increases the number of Lactobacillus and Bifidobacterium in the chyme of the stomach, duodenum, jejunum, ileum, colon and caecum. | Improvement of intestinal microflora. | in vivo | [156] |
| Very low birth weight neonates. | Oral administration of bovine lactoferrin (100 mg day) alone or in combination with the Lactobacillus rhamnosus GG (6 × 109 colony-forming units/day). | Affects the progression from bacterial and fungal colonisation to infection. | Reduction of the incidence of a first episode of late-onset sepsis. | Clinical trial | [160] |
| Preterm infants. | Enteral administration of recombinant human lactoferrin (talactoferrin, TLf) 150 mg kg/12 h from day 1 until day 28 of life. | Alteration of faecal microbiome with a reduction of gram-positive pathogenic bacteria. | Reduction in the rate of urinary tract infections, possibly associated with enteric prophylaxis with TLf. | Clinical trial | [162] |
| Paediatric patients undergoing chemotherapy. | Oral supplementation of bovine lactoferrin (200 mg/day) for two months. | Promotes gut microbiota eubiosis by containing the growth of pathobionts (e.g., Enterococcus) and modulating the abundance of other taxa relevant to intestinal health (i.e., Akkermansia). | Counteracts the onset of dysbiosis, thus ameliorating health and the response to therapy. | Clinical trial | [167] |
| HIV–infected patients undergoing antiretroviral therapy. | Oral supplementation of recombinant human lactoferrin (1500 mg twice a day). | No significant changes in gut microbiota composition. | Promotes a remarkable stability in the gut microbial community. | Clinical trial | [168] |
| Healthy elderly women. | Oral supplementation of bovine lactoferrin (1 g/day), alone or in combination with galactooligosaccharides and vitamin D. | Increases Holdemanella in the faecal microbiota; increases Bifidobacterium in combination with active galactooligosaccharides. | Not assessed. | Clinical trial | [169] |
3.2. α-Lactalbumin
α-Lactalbumin is a 14 kDa globular protein produced by the mammary glands’ epithelial cells and involved in the regulation of lactose synthesis [170]. α-Lactalbumin is the most prevalent whey protein in human milk, constituting approximately 35%, while its quantity is lower in bovine milk, representing the second most abundant whey protein (approximately 17%) after β-lactoglobulin (which is absent in human milk). Due to its nutritional and therapeutic properties, α-lactalbumin is used as a component of infant formula and as a supplement to modulate gastrointestinal and neurological functions and ameliorate diverse disease conditions [171,172]. Even though the mechanism of action underlying these health-promoting effects of α-lactalbumin remains to be elucidated, several pieces of evidence suggest that a pivotal role is played by α-lactalbumin bioactive peptides [173]. Importantly, α-lactalbumin and some of its bioactive peptides have been shown to play a role in the modulation of the gut microbiota by acting as prebiotics (Table 3), and studies suggest this is related both to a direct promotion of probiotics’ growth, including bifidobacteria, and, indirectly, to their antimicrobial activity against diverse pathogens [172,174,175,176]. Xie et al. demonstrated that hyperuricemic mice orally administered with α-lactalbumin gastrointestinal hydrolysates showed a reduction in the levels of serum uric acid, creatinine, and urea nitrogen in association with an increased abundance of some SCFA-producing bacteria, and a decrease of the growth of hyperuricaemia- and inflammation-associated genera [177]. In a previous study, Xie and colleagues demonstrated that treatment with α-lactalbumin hydrolysates is effective in alleviating hypertension-associated intestinal microbiota dysbiosis in spontaneously hypertensive rats [178]. In a study by Chen and colleagues, the effects of treatment with the α-lactalbumin peptide Asp-Gln-Trp in high-fat diet (HFD)-induced NAFLD mice were investigated; results showed that the treatment positively modulated the gut microbiota, reducing the relative abundance of pathogenic bacteria and enhancing the relative abundance of Firmicutes and short-chain fatty acid (SCFA)-producing bacteria [179]. Li et al. demonstrated that bovine α-lactalbumin hydrolysate alters gut microbiota by modulating the Bacteroidetes/Firmicutes ratios and increasing the relative abundance of Lachnospiraceae and Blautia in HFD-induced obese mice. Additionally, this supplementation was demonstrated to reduce the levels of inflammatory cytokines, such as interleukin-6 and tumour necrosis factor-α, and of lipopolysaccharides, leading to a significant reduction of obesity-associated systematic inflammation and endotoxaemia [180]. In another study, performed on high-fat diet-fed mice, the effects of dietary α-lactalbumin on intestinal–adipose–hypothalamic control of energy balance were investigated. Results showed that the supplementation led to a decrease in cd36 and glut2 gene expression in the intestine accompanied by increased cumulative energy ingestion. This was associated with a significant modulation of the gut microbiota with a higher abundance of the Lactobacillus, Parabacteroides and Bifidobacterium genera, compared to controls [181]. In another study, the effects of α-lactalbumin supplementation were tested in preterm pigs that were used as a model for newborn infants. In this study, pigs receiving diets with a high content of α-lactalbumin had a higher abundance of Clostridiaceae, Enterobacteriaceae, Streptococcus, and Streptomyces, compared to controls, but no differences were found at the class or phylum level [182].
Other studies have demonstrated that the health-promoting properties of α-lactalbumin, associated with its effects on positively modulating gut microbiota, can be enhanced when the protein is used in combination with other drugs and/or other milk bioactive components. In a randomized controlled trial, the addition of 3.0 g/L of oligofructose to an α-lactalbumin-enriched term infant formula led to a synergic effect, resulting in a greater increase in faecal bifidobacteria compared to infants receiving the control formula without oligofructose [183]. In a pilot, open label, controlled and interventional study, the authors showed that the oral supplementation with a combination of inositols, α-lactalbumin, and Gymnema sylvestre, positively impacts insulin, glucose, lipid metabolism, and anthropometric measures in dysmetabolic and obese patients, and suggested a role of α-lactalbumin in recovering gut dysbiosis and related metabolic effects [184].
3.3. Lysozyme
Lysozyme is a 14.4 kDa antibacterial enzyme found in plants and animals [185]. In animals, lysozyme is present in diverse biological fluids, including saliva, tears, and milk [186]. Human milk, especially colostrum, has a high content of lysozyme (ranging from 0.2 to 0.9 g/L, depending on lactation stage), while its content is significantly lower in bovine colostrum, with only traces detectable in mature bovine milk [187,188].
As for lactoferrin, the antibacterial activity of lysozyme has a role in reducing the complexity of the gut microbiota, increasing the resistance to intestinal colonisation by some bacterial species, including pathogens, still favouring the growth of beneficial bacteria, and enhancing the recovery from diverse gastrointestinal pathological conditions [189,190]. The mechanisms underlying the resistance of some probiotic strains to the antibacterial action of lysozyme is not yet completely elucidated. However, results from in vitro studies on human-residential bifidobacteria indicated that the tolerance among some bifidobacteria strains is to be ascribed to the non-enzymatic antibacterial activity of lysozyme [191].
Several in vitro and in vivo studies have shown that lysozyme is efficient at modifying the gut microbiota [192,193,194,195] (Table 4). Human lysozyme was found to significantly reduce the number of coliforms and E. coli in the intestine of young pigs fed with milk from transgenic goats expressing human-comparable levels of human lysozyme, compared with pigs fed with milk from non-transgenic goats [196]. Results from similar studies showed that the presence of human lysozyme in goat milk ameliorates intestinal inflammation [197], results in the inhibition of pathogenic E. coli growth in young pig models, and accelerates recovery from E. coli-induced diarrhoea [198]. In addition to modulating pigs’ gut microbiota by decreasing disease-causing bacteria numbers, lysozyme transgenic goat milk was found to increase the ratio of beneficial bacteria, and these changes were associated with improved gut health. The analyses of faecal microbiota showed changes in the bacterial abundance at levels of Firmicutes and Bacteroidetes phyla, with a decrease of Firmicutes and an increase in Bacteroidetes, as well as a reduction of clostridia, Streptococcaceae, and disease-related bacteria such as Mycobacteriaceae and Campylobacterales, in pigs fed with lysozyme transgenic goat milk compared to controls. Furthermore, the presence of lysozyme in goat milk resulted in a gut microbiota more similar to that of breast-fed human infants, which presented a major abundance of Bifidobacteriaceae and Lactobacillaceae [199]. To support these results, other studies have evaluated the effects of lysozyme deficiency on the occurrence of intestinal dysbiosis. The production of lysozyme by Paneth cells is known to be a key factor in gut microbiota modulation and stabilisation [200]. Indeed, deficiencies in the secretion of lysozyme by Paneth cells have been shown to be related to gut microbiota disorders and to an increased vulnerability to bacterial infections and intestinal inflammation [201,202,203]. Interestingly, results from an in vivo study performed on mice with a deoxynivalenol-induced depletion of Paneth cells, demonstrated that supplementation with lysozyme was effective in improving Paneth cells’ functionality and gut microbiota restoration [204].
4. Prebiotic Activity of Glycomacropeptide
Glycomacropeptide (GMP) is a 64-amino acid glycosylated bioactive peptide derived from the C-terminal region of kappa-casein [205]. GMP is mostly found in dairy products, being released in whey by enzymatic digestion during cheese-making processes [206]. However, it is also found free in cow milk and whey, though in lower amounts [172,207].
GMP’s extensive glycosylation is responsible for most of its biological properties [208], which include antibacterial, anti-tumoral, and immunomodulatory effects [205,209]. Interestingly, studies showed that glycosylation, particularly oligosaccharide chains with a high content of sialic acid, accounts for the GMP prebiotic activity, which has been suggested to be responsible for GMP’s diverse biological activities and beneficial health effects [210]. Aside from numerous in vitro studies that demonstrated the efficacy of GMP in inducing the growth of probiotics such as B. infantis, B. breve, and B. bifidum [211,212], in vivo studies and clinical trials have also investigated the prebiotic effects of GMP (Table 5), and the first in vivo demonstrations came from studies performed in mice orally administered GMP. In a study, the treatment induced a significant decrease in faecal Enterobacteriaceae and coliforms together with an increase in Lactobacillus and Bifidobacterium [213]. In another study, it was observed that GMP induces modulation in the gut microbiota, mainly by reducing the growth of Desulfvibrio and increasing Firmicutes, and leads to an increase in caecal concentrations of SCFAs [210]. Experiments performed in allergen (ovalbumin)-sensitized rats orally treated with GMP demonstrated that GMP administration exerts a prebiotic action on allergy-protective microbiota bacteria, increasing the amount of Lactobacillus and Bifidobacterium after 3 days of treatment, and of Bacteroides after 17 days of treatment, suggesting that this, together with other GMP-induced responses, could have a role in the antiallergic activities of GMP [214]. Moreover, GMP hydrolysate was demonstrated to increase the Bacteroidetes/Firmicutes ratio and the abundance of S24-7, Ruminiclostridium, Blautia, and Allobaculum, in high-fat diet-fed and streptozotocin-induced type 2 diabetes mouse models, and these gut microbiota changes were associated with the observed antidiabetic effects [215].
Referring to the GMP effects in modulating human microbiota, a study performed using an artificial colon model of elderly gut microbiota showed that GMP sustained microbiota diversity, decreasing the abundance of Clostridium cluster IV and Ruminococcus and favouring the growth of Blautia spp. [216], suggesting that GMP could exert even in humans the strong prebiotic activity that was observed in in vitro and animal studies. This hypothesis was confirmed in some clinical trials. In a prospective, non-randomized, controlled trial, alterations of gut microbiota, consisting of a significant increase in the abundance of Bifidobacterium, were observed in very preterm infants fed with an infant formula containing a specific prebiotic mixture of 0.65 g scGOS/lcFOS (9:1) and casein GMP providing 40 mg sialic acid/100 mL; these microbiota changes resulted in a more efficient production of neuroactive compounds and energy source utilization [217]. Furthermore, in a two-week clinical trial performed on obese postmenopausal women, alteration in the faecal microbiota, consisting of a reduction of members of the genus Streptococcus and of overall α diversity, was observed after a supplementation with 15 g GMP + 10 g whey protein twice daily for one week, and three times daily for one week, respectively. This study showed that the supplementation of GMP, in combination with other whey proteins, improves satiety and regulates glucose homeostasis, suggesting its potential role as a helpful nutritional supplement for the reduction of metabolic syndrome risk in obese postmenopausal women [218]. However, in other clinical trials, no significant alterations in the gut microbiota were observed after GMP treatment [219,220]. One of these studies was performed in adult subjects with irritable bowel syndrome, and no significant changes in faecal microbiota and faecal immune markers were observed after a three-week period of daily supplementation with 30 g of GMP. The authors suggested that diverse factors, including species-specific gene expression variations, may account for the divergent results obtained from in vitro and animal studies, compared to those from clinical trials [221]. However, all the studies discussed above suffer from some limitations, including interindividual variability and sample size, and further clinical trials are needed to fully assess the role of GMP in modulating the human gut microbiota.
Table 5.
In vivo studies and clinical trials on the modulation of gut microbiota by glycomacropeptide (GMP).
Table 5.
In vivo studies and clinical trials on the modulation of gut microbiota by glycomacropeptide (GMP).
| Patients or Animal Models | Treatment | Effects on Gut Microbiota | Health Effects | Type of Study | Ref. |
|---|---|---|---|---|---|
| Male BALB/c mice. | Oral administration of GMP (0.5 mg/mL at a dose of 0.2 mL per day). | Decrease in Enterobacteriaceae and coliforms; increase in Lactobacillus and Bifidobacterium. | Establishment of a healthier intestinal microbiota. | In vivo | [213] |
| Weaning C57Bl/6 mice. | Dietary supplementation with 20% GMP. | Increases the Firmicutes levels; reduces the growth of Desulfvibrio, thus increasing the caecal concentrations of SCFAs. | Anti-inflammatory effects. | In vivo | [210] |
| High-fat diet-fed and streptozotocin-induced type 2 diabetes C57BL/6J mice. | 8-week GHP hydrolysate dietary supplementation. | Increases the Bacteroidetes/Firmicutes ratio; increases the S24-7, Ruminiclostridium, Blautia, and Allobaculum. | Exerts hypoglycaemic activity; ameliorates dyslipidaemia and inflammation. | In vivo | [215] |
| Allergen-sensitized rats. | Oral administration of GMP for 17 days. | Increases Lactobacillus and Bifidobacterium after 3 days of treatment and increases Bacteroides after 17 days of treatment. | Exerts antiallergic activity. | In vivo | [214] |
| Healthy very preterm infants. | Oral administration of infant formula containing a specific prebiotic mixture 0.65 g scGOS/lcFOS (9:1) and GMP (providing 40 mg sialic acid/100 mL). | Increases Bifidobacterium. | Induces health-beneficial microbiota changes resulting in a more efficient production of neuroactive compounds and energy source utilization. | Clinical trial | [217] |
| Obese postmenopausal women. | Supplementation with 15 g GMP plus 10 g whey protein twice daily for 1 week and thrice daily for 1 week. | Alteration in the faecal microbiota consisting of a reduction of members of the genus Streptococcus and of overall α diversity. | Improves satiety and regulates glucose homeostasis. | Clinical trial | [218] |
| Adult individuals with irritable bowel syndrome. | Three-week period of daily supplementation with 30 g of GMP. | No significant changes in faecal microbiota and faecal immune markers. | No significant effects on inflammation and symptoms of irritable bowel syndrome. | Clinical trial | [221] |
5. Conclusions
Gut microbiota dysbiosis has been shown to be triggered by various factors, including environmental factors, dietary habits, pharmacological therapies, and diverse health conditions. On the other hand, a dysregulated gut microbiota has been shown to be a crucial factor in the establishment of various health conditions, increasing susceptibility not only to chronic inflammatory bowel diseases but also to several systemic pathologies, including cancer, type II diabetes, and some neurological disorders.
Several studies suggested that a healthy gut microbiota can be restored by bioactive compounds derived from food sources, including milk. The possibility of restoring gut microbiota naturally, by exploiting food resources, is intriguing; however, dietary correction, alone, could take a long time, and a more advantageous possibility would be to use these bioactive food components, individually or in combination with other compounds, as dietary supplements. In this context, several studies have been undertaken to evaluate the effects of single milk bioactive compounds or a combination of them on gut microbiota. Evidence from in vitro and in vivo studies demonstrated the potential of milk oligosaccharides and whey proteins in positively modulating gut microbiota, which have shown to exert a strong prebiotic activity, contributing to restoration of healthy gut microbiota and associated health conditions. Interestingly, these results have been confirmed by clinical trials, performed on both infants and adults suffering from different pathologies and/or undergoing pharmacological therapies. In addition, these studies have helped to elucidate the mechanisms of action linked to the prebiotic activity of these compounds; these include direct prebiotic activity and indirect mechanisms of action mainly related to the selective antimicrobial activity exhibited by some of these bioactive milk components, both of which favour the growth of certain probiotic strains to the detriment of pathogens or other non-beneficial strains.
These findings could open up the possibility of exploiting these molecules as therapeutics for the treatment of gut microbiota disorders and associated pathologies. However, while the results of most of the studies provide plausible evidence for the efficacy of milk oligosaccharides and whey proteins in modulating and restoring a healthy gut microbiota, it must be taken into consideration that many studies have been conducted on animals and that, although studies on humans have given similar results, some of them are divergent. Therefore, further clinical trials are needed to overcome the current limitations and to designate these molecules as effective candidates for the treatment of intestinal dysbiosis in humans.
Author Contributions
Conceptualization, V.G. and G.A; resources and data curation, V.G.; writing—original draft preparation, V.G.; writing—review and editing, V.G., G.A., A.A. and F.T.; supervision, G.A. and V.G.; funding acquisition, G.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Funding for Centers of Excellence (Science Department 2023–2027, Roma Tre University, MIUR, Articolo 1, Commi 314–337, Legge 232/2016) and by Next Generation EU PNRR Rome Technopole (ECS_00000024), by MUR, PNRR, Missione 4 Componente 2, “Dalla ricerca all’impresa, Investimento 1.4”. Rome Technopole Innovation Ecosystem—CUP F83B22000040006.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
A part of the graphical abstract was made using icons from BioRender.com.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Corfield, A.P. Mucins: A biologically relevant glycan barrier in mucosal protection. Biochim. Biophys. Acta 2015, 1850, 236–252. [Google Scholar] [CrossRef]
- Heintz-Buschart, A.; Wilmes, P. Human Gut Microbiome: Function Matters. Trends Microbiol. 2018, 26, 563–574. [Google Scholar] [CrossRef]
- Belkaid, Y.; Hand, T.W. Role of the microbiota in immunity and inflammation. Cell 2014, 157, 121–141. [Google Scholar] [CrossRef]
- Chiu, L.; Bazin, T.; Truchetet, M.E.; Schaeverbeke, T.; Delhaes, L.; Pradeu, T. Protective Microbiota: From Localized to Long-Reaching Co-Immunity. Front. Immunol. 2017, 8, 1678. [Google Scholar] [CrossRef] [PubMed]
- Shandilya, S.; Kumar, S.; Kumar Jha, N.; Kumar Kesari, K.; Ruokolainen, J. Interplay of gut microbiota and oxidative stress: Perspective on neurodegeneration and neuroprotection. J. Adv. Res. 2021, 38, 223–244. [Google Scholar] [CrossRef]
- Le Chatelier, E.; Nielsen, T.; Qin, J.; Prifti, E.; Hildebrand, F.; Falony, G.; Almeida, M.; Arumugam, M.; Batto, J.M.; Kennedy, S.; et al. Richness of human gut microbiome correlates with metabolic markers. Nature 2013, 500, 541–546. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Pedersen, O. Gut microbiota in human metabolic health and disease. Nat. Rev. Microbiol. 2021, 19, 55–71. [Google Scholar] [CrossRef]
- Cai, J.; Rimal, B.; Jiang, C.; Chiang, J.Y.L.; Patterson, A.D. Bile acid metabolism and signaling, the microbiota, and metabolic disease. Pharmacol. Ther. 2022, 237, 108238. [Google Scholar] [CrossRef]
- Bordenstein, S.R.; Theis, K.R. Host Biology in Light of the Microbiome: Ten Principles of Holobionts and Hologenomes. PLoS Biol. 2015, 13, 1002226. [Google Scholar] [CrossRef] [PubMed]
- Koenig, J.E.; Spor, A.; Scalfone, N.; Fricker, A.D.; Stombaugh, J.; Knight, R.; Angenent, L.T.; Ley, R.E. Succession of microbial consortia in the developing infant gut microbiome. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. 1), 4578–4585. [Google Scholar] [CrossRef]
- Yatsunenko, T.; Rey, F.E.; Manary, M.J.; Trehan, I.; Dominguez-Bello, M.G.; Contreras, M.; Magris, M.; Hidalgo, G.; Baldassano, R.N.; Anokhin, A.P.; et al. Human gut microbiome viewed across age and geography. Nature 2012, 486, 222–227. [Google Scholar] [CrossRef]
- Bäckhed, F.; Roswall, J.; Peng, Y.; Feng, Q.; Jia, H.; Kovatcheva-Datchary, P.; Li, Y.; Xia, Y.; Xie, H.; Zhong, H.; et al. Dynamics and Stabilization of the Human Gut Microbiome during the First Year of Life. Cell Host Microbe. 2015, 17, 690–703. [Google Scholar] [CrossRef]
- Rodríguez, J.M.; Murphy, K.; Stanton, C.; Ross, R.P.; Kober, O.I.; Juge, N.; Avershina, E.; Rudi, K.; Narbad, A.; Jenmalm, M.C.; et al. The composition of the gut microbiota throughout life, with an emphasis on early life. Microb. Ecol. Health Dis. 2015, 26, 26050. [Google Scholar] [CrossRef]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. What is the Healthy Gut Microbiota Composition? A Changing Ecosystem across Age, Environment, Diet, and Diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef]
- Lozupone, C.A.; Stombaugh, J.I.; Gordon, J.I.; Jansson, J.K.; Knight, R. Diversity, stability and resilience of the human gut microbiota. Nature 2012, 489, 220–230. [Google Scholar] [CrossRef]
- Bäckhed, F.; Ley, R.E.; Sonnenburg, J.L.; Peterson, D.A.; Gordon, J.I. Host-bacterial mutualism in the human intestine. Science 2005, 307, 1915–1920. [Google Scholar] [CrossRef]
- Stojanov, S.; Berlec, A.; Štrukelj, B. The Influence of Probiotics on the Firmicutes/Bacteroidetes Ratio in the Treatment of Obesity and Inflammatory Bowel disease. Microorganisms 2020, 8, 1715. [Google Scholar] [CrossRef]
- Singh, R.K.; Chang, H.W.; Yan, D.; Lee, K.M.; Ucmak, D.; Wong, K.; Abrouk, M.; Farahnik, B.; Nakamura, M.; Zhu, T.H.; et al. Influence of diet on the gut microbiome and implications for human health. J. Transl. Med. 2017, 15, 73. [Google Scholar] [CrossRef]
- Wang, F.; Yu, T.; Huang, G.; Cai, D.; Liang, X.; Su, H.; Zhu, Z.; Li, D.; Yang, Y.; Shen, P.; et al. Gut Microbiota Community and Its Assembly Associated with Age and Diet in Chinese Centenarians. J. Microbiol. Biotechnol. 2015, 25, 1195–1204. [Google Scholar] [CrossRef]
- Mikkelsen, K.H.; Frost, M.; Bahl, M.I.; Licht, T.R.; Jensen, U.S.; Rosenberg, J.; Pedersen, O.; Hansen, T.; Rehfeld, J.F.; Holst, J.J.; et al. Effect of Antibiotics on Gut Microbiota, Gut Hormones and Glucose Metabolism. PLoS ONE 2015, 10, e0142352. [Google Scholar] [CrossRef]
- Reijnders, D.; Goossens, G.H.; Hermes, G.D.; Neis, E.P.; van der Beek, C.M.; Most, J.; Holst, J.J.; Lenaerts, K.; Kootte, R.S.; Nieuwdorp, M.; et al. Effects of Gut Microbiota Manipulation by Antibiotics on Host Metabolism in Obese Humans: A Randomized Double-Blind Placebo-Controlled Trial. Cell Metab. 2016, 24, 63–74. [Google Scholar] [CrossRef]
- Rogers, M.A.M.; Aronoff, D.M. The influence of non-steroidal anti-inflammatory drugs on the gut microbiome. Clin. Microbiol. Infect. 2016, 22, 178.e1–178.e9. [Google Scholar] [CrossRef]
- Goodrich, J.K.; Waters, J.L.; Poole, A.C.; Sutter, J.L.; Koren, O.; Blekhman, R.; Beaumont, M.; Van Treuren, W.; Knight, R.; Bell, J.T.; et al. Human genetics shape the gut microbiome. Cell 2014, 159, 789–799. [Google Scholar] [CrossRef] [PubMed]
- Davenport, E.R.; Cusanovich, D.A.; Michelini, K.; Barreiro, L.B.; Ober, C.; Gilad, Y. Genome-wide association studies of the human gut microbiota. PLoS ONE 2015, 10, e0140301. [Google Scholar] [CrossRef]
- Dehingia, M.; Sen, S.; Bhaskar, B.; Joishy, T.K.; Deka, M.; Talukdar, N.C.; Khan, M.R. Ethnicity influences gut metabolites and microbiota of the tribes of Assam, India. Metabolomics 2017, 13, 69. [Google Scholar] [CrossRef]
- Zhang, J.; Guo, Z.; Xue, Z.; Sun, Z.; Zhang, M.; Wang, L.; Wang, G.; Wang, F.; Xu, J.; Cao, H.; et al. A phylo-functional core of gut microbiota in healthy young Chinese cohorts across lifestyles, geography and ethnicities. ISME J. 2015, 9, 1979–1990. [Google Scholar] [CrossRef] [PubMed]
- Chevalier, C.; Stojanović, O.; Colin, D.J.; Suarez-Zamorano, N.; Tarallo, V.; Veyrat-Durebex, C.; Rigo, D.; Fabbiano, S.; Stevanović, A.; Hagemann, S.; et al. Gut Microbiota Orchestrates Energy Homeostasis during Cold. Cell 2015, 163, 1360–1374. [Google Scholar] [CrossRef]
- Hollister, E.B.; Gao, C.; Versalovic, J. Compositional and functional features of the gastrointestinal microbiome and their effects on human health. Gastroenterology 2014, 146, 1449–1458. [Google Scholar] [CrossRef]
- El Aidy, S.; van den Bogert, B.; Kleerebezem, M. The small intestine microbiota, nutritional modulation and relevance for health. Curr. Opin. Biotechnol. 2015, 32, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Adak, A.; Khan, M.R. An insight into gut microbiota and its functionalities. Cell Mol. Life Sci. 2019, 76, 473–493. [Google Scholar] [CrossRef]
- Gomaa, E.Z. Human gut microbiota/microbiome in health and diseases: A review. Antonie Van Leeuwenhoek 2020, 113, 2019–2040. [Google Scholar] [CrossRef] [PubMed]
- Lynch, S.V.; Pedersen, O. The Human Intestinal Microbiome in Health and Disease. N. Engl. J. Med. 2016, 375, 2369–2379. [Google Scholar] [CrossRef] [PubMed]
- Mor, A.; Antonsen, S.; Kahlert, J.; Holsteen, V.; Jørgensen, S.; Holm-Pedersen, J.; Sørensen, H.T.; Pedersen, O.; Ehrenstein, V. Prenatal exposure to systemic antibacterials and overweight and obesity in Danish schoolchildren: A prevalence study. Int. J. Obes. 2015, 39, 1450–1455. [Google Scholar] [CrossRef]
- Smits, S.A.; Leach, J.; Sonnenburg, E.D.; Gonzalez, C.G.; Lichtman, J.S.; Reid, G.; Knight, R.; Manjurano, A.; Changalucha, J.; Elias, J.E.; et al. Seasonal cycling in the gut microbiome of the Hadza hunter-gatherers of Tanzania. Science 2017, 357, 802–806. [Google Scholar] [CrossRef]
- Weiss, G.A.; Hennet, T. Mechanisms and consequences of intestinal dysbiosis. Cell Mol. Life Sci. 2017, 74, 2959–2977. [Google Scholar] [CrossRef]
- Esteve, E.; Ricart, W.; Fernández-Real, J.M. Gut microbiota interactions with obesity, insulin resistance and type 2 diabetes: Did gut microbiote co-evolve with insulin resistance? Curr. Opin. Clin. Nutr. Metab. Care 2011, 14, 483–490. [Google Scholar] [CrossRef]
- Schwabe, R.F.; Jobin, C. The microbiome and cancer. Nat. Rev. Cancer 2013, 13, 800–812. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.; Wei, H.; Liu, W.; Coker, O.O.; Gou, H.; Liu, C.; Zhao, L.; Li, C.; Zhou, Y.; Wang, G.; et al. Cigarette smoke promotes colorectal cancer through modulation of gut microbiota and related metabolites. Gut 2022, 71, 2439–2450. [Google Scholar] [CrossRef]
- Liu, S.; Gao, J.; Zhu, M.; Liu, K.; Zhang, H.L. Gut Microbiota and Dysbiosis in Alzheimer’s Disease: Implications for Pathogenesis and Treatment. Mol. Neurobiol. 2020, 57, 5026–5043. [Google Scholar] [CrossRef] [PubMed]
- Sonali, S.; Ray, B.; Ahmed Tousif, H.; Rathipriya, A.G.; Sunanda, T.; Mahalakshmi, A.M.; Rungratanawanich, W.; Essa, M.M.; Qoronfleh, M.W.; Chidambaram, S.B.; et al. Mechanistic Insights into the Link between Gut Dysbiosis and Major Depression: An Extensive Review. Cells 2022, 11, 1362. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Wang, H.; Chen, X.; Zhang, Y.; Zhang, H.; Xie, P. Gut microbiota and its metabolites in depression: From pathogenesis to treatment. EBioMedicine 2023, 90, 104527. [Google Scholar] [CrossRef]
- Fröhlich, E.E.; Farzi, A.; Mayerhofer, R.; Reichmann, F.; Jačan, A.; Wagner, B.; Zinser, E.; Bordag, N.; Magnes, C.; Fröhlich, E.; et al. Cognitive impairment by antibiotic-induced gut dysbiosis: Analysis of gut microbiota-brain communication. Brain Behav. Immun. 2016, 56, 40–55. [Google Scholar] [CrossRef]
- Yonekura, S.; Terrisse, S.; Alves Costa Silva, C.; Lafarge, A.; Iebba, V.; Ferrere, G.; Goubet, A.G.; Fahrner, J.E.; Lahmar, I.; Ueda, K.; et al. Cancer Induces a Stress Ileopathy Depending on β-Adrenergic Receptors and Promoting Dysbiosis that Contributes to Carcinogenesis. Cancer Discov. 2022, 12, 1128–1151. [Google Scholar] [CrossRef]
- Relman, D.A. The human microbiome: Ecosystem resilience and health. Nutr. Rev. 2012, 70, S2–S9. [Google Scholar] [CrossRef]
- Smits, L.P.; Bouter, K.E.; de Vos, W.M.; Borody, T.J.; Nieuwdorp, M. Therapeutic potential of fecal microbiota transplantation. Gastroenterology 2013, 145, 946–953. [Google Scholar] [CrossRef]
- Butel, M.J. Probiotics, gut microbiota and health. Med. Mal. Infect. 2014, 44, 1–8. [Google Scholar] [CrossRef]
- Neri-Numa, I.A.; Pastore, G.M. Novel insights into prebiotic properties on human health: A review. Food Res. Int. 2020, 131, 108973. [Google Scholar] [CrossRef]
- Huang, J.; Gong, C.; Zhou, A. Modulation of gut microbiota: A novel approach to enhancing the effects of immune checkpoint inhibitors. Ther. Adv. Med. Oncol. 2023, 15, 17588359231204854. [Google Scholar] [CrossRef]
- Davani-Davari, D.; Negahdaripour, M.; Karimzadeh, I.; Seifan, M.; Mohkam, M.; Masoumi, S.J.; Berenjian, A.; Ghasemi, Y. Prebiotics: Definition, Types, Sources, Mechanisms, and Clinical Applications. Foods 2019, 8, 92. [Google Scholar] [CrossRef]
- Sánchez, C.; Franco, L.; Regal, P.; Lamas, A.; Cepeda, A.; Fente, C. Breast Milk: A Source of Functional Compounds with Potential Application in Nutrition and Therapy. Nutrients 2021, 13, 1026. [Google Scholar] [CrossRef]
- Spicer, S.K.; Gaddy, J.A. Townsend SD. Recent advances on human milk oligosaccharide antimicrobial activity. Curr. Opin. Chem. Biol. 2022, 71, 102202. [Google Scholar] [CrossRef]
- Carr, L.E.; Virmani, M.D.; Rosa, F.; Munblit, D.; Matazel, K.S.; Elolimy, A.A.; Yeruva, L. Role of Human Milk Bioactives on Infants’ Gut and Immune Health. Front. Immunol. 2021, 12, 604080. [Google Scholar] [CrossRef]
- Xiao, J.; Ma, J.; Khan, M.Z.; Alugongo, G.M.; Chen, T.; Liu, S.; Li, S.; Cao, Z. Unlocking the potential of milk whey protein components in colorectal cancer prevention and therapy. Crit. Rev. Food Sci. Nutr. 2023, 17, 1–38. [Google Scholar] [CrossRef]
- Hill, D.R.; Newburg, D.S. Clinical applications of bioactive milk components. Nutr. Rev. 2015, 3, 463–476. [Google Scholar] [CrossRef]
- Davies, R.W.; Carson, B.P.; Jakeman, P.M. The Effect of Whey Protein Supplementation on the Temporal Recovery of Muscle Function Following Resistance Training: A Systematic Review and Meta-Analysis. Nutrients 2018, 10, 221. [Google Scholar] [CrossRef]
- Kim, K.U.; Kim, W.H.; Jeong, C.H.; Yi, D.Y.; Min, H. More than Nutrition: Therapeutic Potential of Breast Milk-Derived Exosomes in Cancer. Int. J. Mol. Sci. 2020, 21, 7327. [Google Scholar] [CrossRef]
- Galley, J.D.; Besner, G.E. The Therapeutic Potential of Breast Milk-Derived Extracellular Vesicles. Nutrients 2020, 12, 745. [Google Scholar] [CrossRef]
- Dybdahl, M.; Selesko, D.B.; Mikkelsen, U.R. Safety evaluation of whey derived beta-lactoglobulin, Lacprodan®® BLG. Toxicol. Rep. 2021, 8, 617–626. [Google Scholar] [CrossRef]
- Lyons, K.E.; Ryan, C.A.; Dempsey, E.M.; Ross, R.P.; Stanton, C. Breast Milk, a source of beneficial microbes and associated benefits for infant health. Nutrients 2020, 12, 1039. [Google Scholar] [CrossRef]
- Calkins, K.; Devaskar, S.U. Fetal Origins of Adult Disease. Curr. Probl. Pediatr. Adolesc. Health Care 2011, 41, 158–176. [Google Scholar] [CrossRef] [PubMed]
- Manoppo, J.I.C.; Nurkolis, F.; Gunawan, W.B.; Limen, G.A.; Rompies, R.; Heroanto, J.P.; Natanael, H.; Phan, S.; Tanjaya, K. Functional sterol improves breast milk quality by modulating the gut microbiota: A proposed opinion for breastfeeding mothers. Front. Nutr. 2022, 9, 1018153. [Google Scholar] [CrossRef]
- Adlerberth, I.; Wold, A.E. Establishment of the gut microbiota in Western infants. Acta Paediatr. 2009, 98, 229–238. [Google Scholar] [CrossRef]
- Fallani, M.; Young, D.; Scott, J.; Norin, E.; Amarri, S.; Adam, R.; Aguilera, M.; Khanna, S.; Gil, A.; Edwards, C.A.; et al. Other Members of the INFABIO Team. Intestinal microbiota of 6-week-old infants across Europe: Geographic influence beyond delivery mode, breast-feeding, and antibiotics. J. Pediatr. Gastroenterol. Nutr. 2010, 51, 77–84. [Google Scholar] [CrossRef]
- Marcobal, A.; Barboza, M.; Froehlich, J.W.; Block, D.E.; German, B.; Lebrilla, C.B.; Mills, D.A. Consumption of human milk oligosaccharides by gut-related microbes. J. Agric. Food Chem. 2010, 58, 5334–5340. [Google Scholar] [CrossRef]
- Le Doare, K.; Holder, B.; Bassett, A.; Pannaraj, P.S. Mother’s Milk: A Purposeful Contribution to the Development of the Infant Microbiota and Immunity. Front. Immunol. 2018, 9, 361. [Google Scholar] [CrossRef]
- Zimmermann, P.; Curtis, N. Breast milk microbiota: A review of the factors that influence composition. J. Infect. 2020, 81, 17–47. [Google Scholar] [CrossRef]
- Maga, E.A.; Weimer, B.C.; Murray, J.D. Dissecting the role of milk components on gut microbiota composition. Gut Microbes. 2013, 4, 136–139. [Google Scholar] [CrossRef] [PubMed]
- Gopalakrishna, K.P.; Hand, T.W. Influence of Maternal Milk on the Neonatal Intestinal Microbiome. Nutrients 2020, 12, 823. [Google Scholar] [CrossRef] [PubMed]
- Bakshani, C.R.; Crouch, L.I. Human milk oligosaccharides and Bifidobacterium species. Trends Microbiol. 2024, 32, 118–119. [Google Scholar] [CrossRef]
- Lordan, C.; Roche, A.K.; Delsing, D.; Nauta, A.; Groeneveld, A.; MacSharry, J.; Cotter, P.D.; van Sinderen, D. Linking human milk oligosaccharide metabolism and early life gut microbiota: Bifidobacteria and beyond. Microbiol. Mol. Biol. Rev. 2024, 11, e0009423. [Google Scholar] [CrossRef] [PubMed]
- Boudry, G.; Charton, E.; Le Huerou-Luron, I.; Ferret-Bernard, S.; Le Gall, S.; Even, S.; Blat, S. The Relationship Between Breast Milk Components and the Infant Gut Microbiota. Front. Nutr. 2021, 8, 629740. [Google Scholar] [CrossRef]
- Xi, M.; Liang, D.; Yan, Y.; Duan, S.; Leng, H.; Yang, H.; Shi, X.; Na, X.; Yang, Y.; Yang, C.; et al. Functional proteins in breast milk and their correlation with the development of the infant gut microbiota: A study of mother-infant pairs. Front. Microbiol. 2023, 14, 1239501. [Google Scholar] [CrossRef] [PubMed]
- Gopal, P.K.; Gill, H.S. Oligosaccharides and glycoconjugates in bovine milk and colostrum. Br. J. Nutr. 2000, 84, S69–S74. [Google Scholar] [CrossRef]
- Totten, S.M.; Zivkovic, A.M.; Wu, S.; Ngyuen, U.; Freeman, S.L.; Ruhaak, L.R.; Darboe, M.K.; German, J.B.; Prentice, A.M.; Lebrilla, C.B. Comprehensive profiles of human milk oligosaccharides yield highly sensitive and specific markers for determining secretor status in lactating mothers. J. Proteome Res. 2012, 11, 6124–6133. [Google Scholar] [CrossRef]
- Donovan, S.M.; Comstock, S.S. Human Milk Oligosaccharides Influence Neonatal Mucosal and Systemic Immunity. Ann. Nutr. Metab. 2016, 69, 42–51. [Google Scholar] [CrossRef]
- De Leoz, M.L.; Kalanetra, K.M.; Bokulich, N.A.; Strum, J.S.; Underwood, M.A.; German, J.B.; Mills, D.A.; Lebrilla, C.B. Human milk glycomics and gut microbial genomics in infant feces show a correlation between human milk oligosaccharides and gut microbiota: A proof-of-concept study. J. Proteome Res. 2015, 14, 491–502. [Google Scholar] [CrossRef]
- Sela, D.A.; Mills, D.A. Nursing our microbiota: Molecular linkages between bifidobacteria and milk oligosaccharides. Trends Microbiol. 2010, 18, 298–307. [Google Scholar] [CrossRef]
- Ward, R.E.; Ninonuevo, M.; Mills, D.A.; Lebrilla, C.B.; German, J.B. In vitro fermentability of human milk oligosaccharides by several strains of bifidobacteria. Mol. Nutr. Food Res. 2007, 51, 1398–1405. [Google Scholar] [CrossRef] [PubMed]
- Thomson, P.; Medina, D.A.; Garrido, D. Human milk oligosaccharides and infant gut bifidobacteria: Molecular strategies for their utilization. Food Microbiol. 2017, 75, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Ganzle, M.G.; Follador, R. Metabolism of oligosaccharides and starch in lactobacilli: A review. Front. Microbiol. 2012, 3, 340. [Google Scholar] [CrossRef]
- Kang, J.E.; Kim, T.J.; Moon, G.S. A Novel Lactobacillus casei LP1 Producing 1,4-Dihydroxy-2-Naphthoic Acid, a Bifidogenic Growth Stimulator. Prev. Nutr. Food Sci. 2015, 20, 78–81. [Google Scholar] [CrossRef]
- Garrido, D.; Dallas, D.C.; Mills, D.A. Consumption of human milk glycoconjugates by infant-associated bifidobacteria: Mechanisms and implications. Microbiology 2013, 159, 649–664. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Li, L.Q.; Liu, F.; Wu, J.Y. Human milk oligosaccharides and infant gut microbiota: Molecular structures, utilization strategies and immune function. Carbohydr. Polym. 2022, 276, 118738. [Google Scholar] [CrossRef]
- Ruiz-Palacios, G.M.; Cervantes, L.E.; Ramos, P.; Chavez-Munguia, B.; Newburg, D.S. Campylobacter jejuni binds intestinal H(O) antigen (Fuc alpha 1, 2Gal beta 1, 4GlcNAc), and fucosyloligosaccharides of human milk inhibit its binding and infection. J. Biol. Chem. 2003, 278, 14112–14120. [Google Scholar] [CrossRef] [PubMed]
- Morrow, A.L.; Ruiz-Palacios, G.M.; Jiang, X.; Newburg, D.S. Human-milk glycans that inhibit pathogen binding protect breast-feeding infants against infectious diarrhea. J. Nutr. 2005, 135, 1304–1307. [Google Scholar] [CrossRef] [PubMed]
- Coppa, G.V.; Zampini, L.; Galeazzi, T.; Facinelli, B.; Ferrante, L.; Capretti, R.; Orazio, G. Human milk oligosaccharides inhibit the adhesion to Caco-2 cells of diarrheal pathogens: Escherichia coli, Vibrio cholerae, and Salmonella fyris. Pediatr. Res. 2006, 59, 377–382. [Google Scholar] [CrossRef] [PubMed]
- Vigsnaes, L.K.; Ghyselinck, J.; Van den Abbeele, P.; McConnell, B.; Moens, F.; Marzorati, M.; Bajic, D. 2′FL and LNnT Exert Antipathogenic Effects against C. Difficile ATCC 9689 In vitro, Coinciding with Increased Levels of Bifidobacteriaceae and/or Secondary Bile Acids. Pathogens 2021, 10, 927. [Google Scholar] [CrossRef]
- Singh, R.P.; Niharika, J.; Kondepudi, K.K.; Bishnoi, M.; Tingirikari, J.M.R. Recent understanding of human milk oligosaccharides in establishing infant gut microbiome and roles in immune system. Food Res. Int. 2022, 151, 110884. [Google Scholar] [CrossRef]
- Vatanen, T.; Kostic, A.D.; d’Hennezel, E.; Siljander, H.; Franzosa, E.A.; Yassour, M.; Kolde, R.; Vlamakis, H.; Arthur, T.D.; Hämäläinen, A.M.; et al. Variation in Microbiome LPS Immunogenicity Contributes to Autoimmunity in Humans. Cell 2016, 165, 842–853. [Google Scholar] [CrossRef]
- Arrieta, M.C.; Arévalo, A.; Stiemsma, L.; Dimitriu, P.; Chico, M.E.; Loor, S.; Vaca, M.; Boutin, R.C.T.; Morien, E.; Jin, M.; et al. Associations between infant fungal and bacterial dysbiosis and childhood atopic wheeze in a nonindustrialized setting. J. Allergy Clin. Immunol. 2018, 142, 424–434. [Google Scholar] [CrossRef]
- Henrick, B.M.; Rodriguez, L.; Lakshmikanth, T.Y.; Pou, C.; Henckel, E.; Arzoomand, A.; Olin, A.; Wang, J.; Mikes, J.; Tan, Z.; et al. Bifidobacteria-mediated immune system imprinting early in life. Cell 2021, 184, 3884–3898. [Google Scholar] [CrossRef]
- Rivière, A.; Selak, M.; Lantin, D.; Leroy, F.; De Vuyst, L. Bifidobacteria and Butyrate-Producing Colon Bacteria: Importance and Strategies for Their Stimulation in the Human Gut. Front. Microbiol. 2016, 7, 979. [Google Scholar] [CrossRef]
- Verkhnyatskaya, S.; Ferrari, M.; de Vos, P.; Walvoort, M.T.C. Shaping the Infant Microbiome with Non-digestible Carbohydrates. Front. Microbiol. 2019, 10, 343. [Google Scholar] [CrossRef]
- Duranti, S.; Ruiz, L.; Lugli, G.A.; Tames, H.; Milani, C.; Mancabelli, L.; Mancino, W.; Longhi, G.; Carnevali, L.; Sgoifo, A.; et al. Bifidobacterium adolescentis as a key member of the human gut microbiota in the production of GABA. Sci. Rep. 2020, 10, 14112. [Google Scholar] [CrossRef]
- Madison, C.A.; Hillbrick, L.; Kuempel, J.; Albrecht, G.L.; Landrock, K.K.; Safe, S.; Chapkin, R.S.; Eitan, S. Intestinal Epithelium Aryl Hydrocarbon Receptor Is Involved in Stress Sensitivity and Maintaining Depressive Symptoms. Behav. Brain Res. 2023, 440, 114256. [Google Scholar] [CrossRef]
- Vulevic, J.; Drakoularakou, A.; Yaqoob, P.; Tzortzis, G.; Gibson, G.R. Modulation of the fecal microflora profile and immune function by a novel trans-galactooligosaccharide mixture (B-GOS) in healthy elderly volunteers. Am. J. Clin. Nutr. 2008, 88, 1438–1446. [Google Scholar] [CrossRef] [PubMed]
- Costabile, A.; Kolida, S.; Klinder, A.; Gietl, E.; Bäuerlein, M.; Frohberg, C.; Landschütze, V.; Gibson, G.R. A double-blind, placebo-controlled, cross-over study to establish the bifidogenic effect of a very-long-chain inulin extracted from globe artichoke (Cynara scolymus) in healthy human subjects. Br. J. Nutr. 2010, 104, 1007–1017. [Google Scholar] [CrossRef] [PubMed]
- Finegold, S.M.; Li, Z.; Summanen, P.H.; Downes, J.; Thames, G.; Corbett, K.; Dowd, S.; Krak, M.; Heber, D. Xylooligosaccharide increases bifidobacteria but not lactobacilli in human gut microbiota. Food Funct. 2014, 5, 436–445. [Google Scholar] [CrossRef] [PubMed]
- Šuligoj, T.; Vigsnæs, L.K.; Abbeele, P.V.D.; Apostolou, A.; Karalis, K.; Savva, G.M.; McConnell, B.; Juge, N. Effects of Human Milk Oligosaccharides on the Adult Gut Microbiota and Barrier Function. Nutrients 2020, 12, 2808. [Google Scholar] [CrossRef] [PubMed]
- Elison, E.; Vigsnaes, L.K.; Rindom Krogsgaard, L.; Rasmussen, J.; Sørensen, N.; McConnell, B.; Hennet, T.; Sommer, M.O.; Bytzer, P. Oral supplementation of healthy adults with 2′-O-fucosyllactose and lacto-N-neotetraose is well tolerated and shifts the intestinal microbiota. Br. J. Nutr. 2016, 116, 1356–1368. [Google Scholar] [CrossRef] [PubMed]
- Iribarren, C.; Magnusson, M.K.; Vigsnæs, L.K.; Aziz, I.; Amundsen, I.D.; Šuligoj, T.; Juge, N.; Patel, P.; Sapnara, M.; Johnsen, L.; et al. The Effects of Human Milk Oligosaccharides on Gut Microbiota, Metabolite Profiles and Host Mucosal Response in Patients with Irritable Bowel Syndrome. Nutrients 2021, 13, 3836. [Google Scholar] [CrossRef]
- Bajic, D.; Wiens, F.; Wintergerst, E.; Deyaert, S.; Baudot, A.; Van den Abbeele, P. HMOs Exert Marked Bifidogenic Effects on Children’s Gut Microbiota Ex Vivo, Due to Age-Related Bifidobacterium Species Composition. Nutrients 2023, 15, 1701. [Google Scholar] [CrossRef]
- Donovan, S.M. Human Milk Proteins: Composition and Physiological Significance. Nestle Nutr. Inst. Workshop Ser. 2019, 90, 93–101. [Google Scholar] [CrossRef]
- Schack, L.; Lange, A.; Kelsen, J.; Agnholt, J.; Christensen, B.; Petersen, T.E.; Sørensen, E.S. Considerable variation in the concentration of osteopontin in human milk, bovine milk, and infant formulas. J. Dairy Sci. 2009, 92, 5378–5385. [Google Scholar] [CrossRef] [PubMed]
- Valenti, P.; Marchetti, M.; Superti, F.; Amendolia, M.G.; Puddu, P.; Gessani, S.; Borghi, P.; Berardelli, F.; Antonini, G.; Seganti, L. Antiviral activity of Lactoferrin. In Advances in Lactoferrin Research. Advances in Experimental Medicine and Biology; Spik, G., Legrand, D., Mazurier, J., Pierce, A., Perraudin, J.P., Eds.; Springer: Boston, MA, USA, 1998; Volume 443. [Google Scholar] [CrossRef]
- Krissansen, G.W. Emerging health properties of whey proteins and their clinical implications. J. Am. Coll. Nutr. 2007, 26, 713S–723S. [Google Scholar] [CrossRef] [PubMed]
- Leboffe, L.; Giansanti, F.; Antonini, G. Antifungal and Antiparasitic Activities of Lactoferrin Anti-Infective Agents in Medicinal Chemistry. J. Med. Chem. 2009, 8, 114–127. [Google Scholar] [CrossRef]
- Akhavan, T.; Luhovyy, B.L.; Panahi, S.; Kubant, R.; Brown, P.H.; Anderson, G.H. Mechanism of action of pre-meal consumption of whey protein on glycemic control in young adults. J. Nutr. Biochem. 2014, 25, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, F.J.; Santos, H.O.; Howell, S.L.; Pimentel, G.D. Whey protein in cancer therapy: A narrative review. Pharmacol. Res. 2019, 144, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Gallo, V.; Giansanti, F.; Arienzo, A.; Antonini, G. Antiviral properties of whey proteins and their activity against SARS-CoV-2 infection. J. Funct. Foods 2022, 89, 104932. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Chen, N.; Ashaolu, T.J. Whey proteins and peptides in health-promoting functions-A review. Int. Dairy J. 2022, 126, 105269. [Google Scholar] [CrossRef]
- Mastromarino, P.; Capobianco, D.; Campagna, G.; Laforgia, N.; Drimaco, P.; Dileone, A.; Baldassarre, M.E. Correlation between lactoferrin and beneficial microbiota in breast milk and infant’s feces. Biometals 2014, 27, 1077–1086. [Google Scholar] [CrossRef]
- Giansanti, F.; Panella, G.; Leboffe, L.; Antonini, G. Lactoferrin from Milk: Nutraceutical and Pharmacological Properties. Pharmaceuticals 2016, 9, 61. [Google Scholar] [CrossRef]
- Boscaini, S.; Skuse, P.; Nilaweera, K.N.; Cryan, J.F.; Cotter, P.D. The ‘Whey’ to good health: Whey protein and its beneficial effect on metabolism, gut microbiota and mental health. Trends Food Sci. Technol. 2023, 133, 1–14. [Google Scholar] [CrossRef]
- Kowalczyk, P.; Kaczyńska, K.; Kleczkowska, P.; Bukowska-Ośko, I.; Kramkowski, K.; Sulejczak, D. The Lactoferrin Phenomenon-A Miracle Molecule. Molecules 2022, 27, 2941. [Google Scholar] [CrossRef]
- Wang, B.; Timilsena, Y.P.; Blanch, E.; Adhikari, B. Lactoferrin: Structure, function, denaturation and digestion. Crit. Rev. Food Sci. Nutr. 2019, 59, 580–596. [Google Scholar] [CrossRef]
- Buccigrossi, V.; de Marco, G.; Bruzzese, E.; Ombrato, L.; Bracale, I.; Polito, G.; Guarino, A. Lactoferrin induces concentration-dependent functional modulation of intestinal proliferation and differentiation. Pediatr. Res. 2007, 61, 410–414. [Google Scholar] [CrossRef]
- Giansanti, F.; Panella, G.; Arienzo, A.; Gallo, V.; Leboffe, L.; Antonini, G. Nutraceutical Peptides from Lactoferrin. J. Adv. Dairy Res. 2018, 5, 199. [Google Scholar] [CrossRef]
- Zhao, C.; Chen, N.; Ashaolu, T.J. Prebiotic and modulatory evidence of lactoferrin on gut health and function. J. Funct. Foods 2023, 108, 105741. [Google Scholar] [CrossRef]
- Conesa, C.; Bellés, A.; Grasa, L.; Sánchez, L. The Role of Lactoferrin in Intestinal Health. Pharmaceutics 2023, 15, 1569. [Google Scholar] [CrossRef] [PubMed]
- Actor, K.J.; Hwang, S.A.; Kruzel, L.M. Lactoferrin as a natural immune modulator. Curr. Pharm. Des. 2009, 15, 1956–1973. [Google Scholar] [CrossRef]
- Hu, P.; Zhao, F.; Wang, J.; Zhu, W. Lactoferrin attenuates lipopolysaccharide-stimulated inflammatory responses and barrier impairment through the modulation of NF-κB/MAPK/Nrf2 pathways in IPEC-J2 cells. Food Funct. 2020, 11, 8516–8526. [Google Scholar] [CrossRef] [PubMed]
- Dierick, M.; Vanrompay, D.; Devriendt, B.; Cox, E. Lactoferrin, a versatile natural antimicrobial glycoprotein that modulates the host’s innate immunity. Biochem. Cell Biol. 2021, 99, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Chen, J.; Wu, W.; Shi, J.; Zhong, Y.; van Tol, E.A.; Tang, Q.; Cai, W. Enteral supplementation of bovine lactoferrin improves gut barrier function in rats after massive bowel resection. Br. J. Nutr. 2014, 112, 486–492. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhou, J.; Xiao, D.; Shu, G.; Gu, L. Bovine lactoferrin protects dextran sulfate sodium salt mice against inflammation and impairment of colonic epithelial barrier by regulating gut microbial structure and metabolites. Front. Nutr. 2021, 8, 660598. [Google Scholar] [CrossRef] [PubMed]
- Oda, H.; Wakabayashi, H.; Yamauchi, K.; Abe, F. Lactoferrin and bifidobacteria. Biometals 2014, 27, 915–922. [Google Scholar] [CrossRef] [PubMed]
- Vega-Bautista, A.; de la Garza, M.; Carrero, J.C.; Campos-Rodríguez, R.; Godínez-Victoria, M.; Drago-Serrano, M.E. The Impact of Lactoferrin on the Growth of Intestinal Inhabitant Bacteria. Int. J. Mol. Sci. 2019, 20, 4707. [Google Scholar] [CrossRef] [PubMed]
- Bruni, N.; Capucchio, M.T.; Biasibetti, E.; Pessione, E.; Cirrincione, S.; Giraudo, L.; Corona, A.; Dosio, F. Antimicrobial activity of lactoferrin-related peptides and applications in human and veterinary medicine. Molecules 2016, 21, 752. [Google Scholar] [CrossRef]
- Chen, P.W.; Jheng, T.T.; Shyu, C.L.; Mao, F.C. Antimicrobial potential for the combination of bovine lactoferrin or its hydrolysate with lactoferrin-resistant probiotics against foodborne pathogens. J. Dairy Sci. 2013, 96, 1438–1446. [Google Scholar] [CrossRef]
- Seyoum, Y.; Baye, K.; Humblot, C. Iron homeostasis in host and gut bacteria—A complex interrelationship. Gut Microbes 2021, 13, 1874855. [Google Scholar] [CrossRef]
- Coppa, G.V.; Zampini, L.; Galeazzi, T.; Gabrielli, O. Prebiotics in human milk: A review. Dig. Liver Dis. 2006, 38, S291–S294. [Google Scholar] [CrossRef]
- Lanigan, N.; Bottacini, F.; Casey, P.G.; O’Connell Motherway, M.; van Sinderen, D. Genome-Wide Search for Genes Required for Bifidobacterial Growth under Iron-Limitation. Front. Microbiol. 2017, 8, 964. [Google Scholar] [CrossRef] [PubMed]
- Garrido, D.; Nwosu, C.; Ruiz-Moyano, S.; Aldredge, D.; German, J.B.; Lebrilla, C.B.; Mills, D.A. Endo-beta-N-acetylglucosaminidases from infant gut-associated bifidobacteria release complex N-glycans from human milk glycoproteins. Mol. Cell. Proteom. 2012, 11, 775–785. [Google Scholar] [CrossRef] [PubMed]
- Miller-Catchpole, R.; Kot, E.; Haloftis, G.; Furmanov, S.; Bezkorovainy, A. Lactoferrin can supply iron for the growth of Bifidobacterium breve. Nutr. Res. 1997, 17, 205–213. [Google Scholar] [CrossRef]
- Kim, W.S.; Tanaka, T.; Kumura, H.; Shimazaki, K. Lactoferrin-binding proteins in Bifidobacterium bifidum. Biochem. Cell Biol. 2002, 80, 91–94. [Google Scholar] [CrossRef]
- Rahman, M.M.; Kim, W.S.; Ito, T.; Kumura, H.; Shimazaki, K. Growth promotion and cell binding ability of bovine lactoferrin to Bifidobacterium longum. Anaerobe 2009, 15, 133–137. [Google Scholar] [CrossRef]
- Semenov, D.V.; Kanyshkova, T.G.; Buneva, V.N.; Nevinsky, G.A. Human milk lactoferrin binds ATP and dissociates into monomers. Biochem. Mol. Biol. Int. 1999, 47, 177–184. [Google Scholar] [CrossRef]
- Liu, Z.S.; Chen, P.W. Featured Prebiotic Agent: The Roles and Mechanisms of Direct and Indirect Prebiotic Activities of Lactoferrin and Its Application in Disease Control. Nutrients 2023, 15, 2759. [Google Scholar] [CrossRef]
- Griffiths, E.A.; Duffy, L.C.; Schanbacher, F.L.; Dryja, D.; Leavens, A.; Neiswander, R.L.; Qiao, H.; Di Rienzo, D.; Ogra, P. In vitro growth responses of bifidobacteria and enteropathogens to bovine and human lactoferrin. Dig. Dis. Sci. 2003, 48, 1324–1332. [Google Scholar] [CrossRef]
- Kim, W.S.; Ohashi, M.; Tanaka, T.; Kumura, H.; Kim, G.Y.; Kwon, I.K.; Goh, J.S.; Shimazaki, K. Growth-promoting effects of lactoferrin on L. acidophilus and Bifidobacterium spp. Biometals 2004, 17, 279–283. [Google Scholar] [CrossRef]
- Tian, H.; Maddox, I.S.; Ferguson, L.R.; Shu, Q. Influence of bovine lactoferrin on selected probiotic bacteria and intestinal pathogens. Biometals 2010, 23, 593–596. [Google Scholar] [CrossRef]
- Chen, P.W.; Ku, Y.W.; Chu, F.Y. Influence of bovine lactoferrin on the growth of selected probiotic bacteria under aerobic conditions. Biometals 2014, 27, 905–914. [Google Scholar] [CrossRef]
- Chen, P.W.; Liu, Z.S.; Kuo, T.C.; Hsieh, M.C.; Li, Z.W. Prebiotic effects of bovine lactoferrin on specific probiotic bacteria. Biometals 2017, 30, 237–248. [Google Scholar] [CrossRef]
- Liu, Z.S.; Lin, C.F. Chen PW. Transcriptome analysis of Lactobacillus rhamnosus GG strain treated with prebiotic—Bovine lactoferrin under a cold environment. J. Food Drug Anal. 2021, 29, 402–418. [Google Scholar] [CrossRef]
- Wang, J.; Yixuan, L.; Liang, Z.; Fazheng, R.; Huiyuan, G. Lactoferrin stimulates the expression of vitamin D receptor in vitamin D deficient mice, Journal of Functional. Foods 2019, 55, 48–56. [Google Scholar] [CrossRef]
- Sun, J.; Ren, F.; Xiong, L.; Zhao, L.; Guo, H. Bovine lactoferrin suppresses high-fat diet induced obesity and modulates gut microbiota in C57BL/6J mice. J. Funct. Foods 2016, 22, 189–200. [Google Scholar] [CrossRef]
- Li, D.M.; He, Q.; Yang, H.H.; Du, Y.F.; Yu, K.Q.; Yang, J.; Tong, X.; Guo, Y.X.; Xu, J.Y.; Qin, L.Q. Daily Dose of Bovine Lactoferrin Prevents Ethanol-Induced Liver Injury and Death in Male Mice by Regulating Hepatic Alcohol Metabolism and Modulating Gut Microbiota. Mol. Nutr. Food Res. 2021, 65, e2100253. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Ma, C.; Hurilebagen; Yuan, H.; Hu, R.; Wang, W.; Weilisi. Effects of lactoferrin on intestinal flora of metabolic disorder mice. BMC Microbiol. 2022, 22, 181. [Google Scholar] [CrossRef] [PubMed]
- Bellés, A.; Aguirre-Ramírez, D.; Abad, I.; Parras-Moltó, M.; Sánchez, L.; Grasa, L. Lactoferrin modulates gut microbiota and Toll-like receptors (TLRs) in mice with dysbiosis induced by antibiotics. Food Funct. 2022, 13, 5854–5869. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Zhang, L.L.; Li, D.; Wu, J.; Guo, Y.X.; Fan, J.; Wu, Q.; Wang, H.P.; Wan, Z.; Xu, J.Y.; et al. Lactoferrin alleviates Western diet-induced cognitive impairment through the microbiome-gut-brain axis. Curr. Res. Food Sci. 2023, 7, 100533. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Hao, Y.; Mao, R.; Yang, N.; Zheng, X.; Li, B.; Wang, Z.; Zhang, Q.; Teng, D.; Wang, J. Effects of dietary supplementation of bovine lactoferrin on growth performance, immune function and intestinal health in weaning piglets. Biometals 2023, 36, 587–601. [Google Scholar] [CrossRef] [PubMed]
- Hu, P.; Zhao, F.; Zhu, W.; Wang, J. Effects of early-life lactoferrin intervention on growth performance, small intestinal function and gut microbiota in suckling piglets. Food Funct. 2019, 10, 5361–5373. [Google Scholar] [CrossRef]
- Hu, W.; Zhao, J.; Wang, J.; Yu, T.; Wang, J.; Li, N. Transgenic milk containing recombinant human lactoferrin modulates the intestinal flora in piglets. Biochem. Cell Biol. 2012, 90, 485–496. [Google Scholar] [CrossRef]
- Yang, C.; Zhu, X.; Liu, N.; Chen, Y.; Gan, H.; Troy, F.A.; Wang, B. Lactoferrin up-regulates intestinal gene expression of brain-derived neurotrophic factors BDNF, UCHL1 and alkaline phosphatase activity to alleviate early weaning diarrhea in postnatal piglets. J. Nutr. Biochem. 2014, 25, 834–842. [Google Scholar] [CrossRef]
- Grzywacz, K.; Butcher, J.; Romain, G.; Li, J.; Stintzi, A. The impact of probiotics and lactoferrin supplementation on piglet gastrointestinal microbial communities. Biometals 2019, 32, 533–543. [Google Scholar] [CrossRef]
- Tang, X.S.; Shao, H.; Li, T.J.; Tang, Z.R.; Huang, R.L.; Wang, S.P.; Kong, X.F.; Wu, X.; Yin, Y.L. Dietary supplementation with bovine lactoferrampin-lactoferricin produced by Pichia pastoris fed-batch fermentation affects intestinal microflora in weaned piglets. Appl. Biochem. Biotechnol. 2012, 168, 887–898. [Google Scholar] [CrossRef]
- Cooper, C.A.; Nelson, K.M.; Maga, E.A.; Murray, J.D. Consumption of transgenic cows’ milk containing human lactoferrin results in beneficial changes in the gastrointestinal tract and systemic health of young pigs. Transgenic Res. 2013, 22, 571–578. [Google Scholar] [CrossRef]
- Reznikov, E.A.; Comstock, S.S.; Yi, C.; Contractor, N.; Donovan, S.M. Dietary bovine lactoferrin increases intestinal cell proliferation in neonatal piglets. J. Nutr. 2014, 144, 1401–1408. [Google Scholar] [CrossRef]
- He, Y.; Cao, L.; Yu, J. Prophylactic lactoferrin for preventing late-onset sepsis and necrotizing enterocolitis in preterm infants: A PRISMA-compliant systematic review and meta-analysis. Medicine 2018, 97, e11976. [Google Scholar] [CrossRef]
- Manzoni, P.; Rinaldi, M.; Cattani, S.; Pugni, L.; Romeo, M.G.; Messner, H.; Stolfi, I.; Decembrino, L.; Laforgia, N.; Vagnarelli, F.; et al. Bovine lactoferrin supplementation for prevention of late-onset sepsis in very low-birth-weight neonates: A randomized trial. JAMA 2009, 302, 1421–1428. [Google Scholar] [CrossRef]
- Ochoa, T.J.; Zegarra, J.; Cam, L.; Llanos, R.; Pezo, A.; Cruz, K.; Zea-Vera, A.; Cárcamo, C.; Campos, M.; Bellomo, S. Randomized controlled trial of lactoferrin for prevention of sepsis in peruvian neonates less than 2500 g. Pediatr. Infect. Dis. J. 2015, 34, 571–576. [Google Scholar] [CrossRef]
- Sherman, M.P.; Adamkin, D.H.; Niklas, V.; Radmacher, P.; Sherman, J.; Wertheimer, F.; Petrak, K. Randomized Controlled Trial of Talactoferrin Oral Solution in Preterm Infants. J. Pediatr. 2016, 175, 68–73.e3. [Google Scholar] [CrossRef]
- Pammi, M.; Preidis, G.A.; Tarnow-Mordi, W.O. Evidence from systematic reviews of randomized trials on enteral lactoferrin supplementation in preterm neonates. Biochem. Cell Biol. 2021, 99, 20–24. [Google Scholar] [CrossRef] [PubMed]
- Roggiani, S.; Mengoli, M.; Conti, G.; Fabbrini, M.; Brigidi, P.; Barone, M.; D’Amico, F.; Turroni, S. Gut microbiota resilience and recovery after anticancer chemotherapy. Microbiome Res. Rep. 2023, 2, 16. [Google Scholar] [CrossRef]
- Vujkovic-Cvijin, I.; Dunham, R.M.; Iwai, S.; Maher, M.C.; Albright, R.G.; Broadhurst, M.J.; Hernandez, R.D.; Lederman, M.M.; Huang, Y.; Somsouk, M.; et al. Dysbiosis of the gut microbiota is associated with HIV disease progression and tryptophan catabolism. Sci. Transl. Med. 2013, 5, 193ra91. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.J.; Walmsley, S.L.; Raboud, J.M.; Kovacs, C.; Coburn, B.; Rousseau, R.; Reinhard, R.; Rosenes, R.; Kaul, R. Can Probiotics Reduce Inflammation and Enhance Gut Immune Health in People Living with HIV: Study Designs for the Probiotic Visbiome for Inflammation and Translocation (PROOV IT) Pilot Trials. HIV Clin. Trials 2016, 17, 147–157. [Google Scholar] [CrossRef]
- D’Amico, F.; Decembrino, N.; Muratore, E.; Turroni, S.; Muggeo, P.; Mura, R.; Perruccio, K.; Vitale, V.; Zecca, M.; Prete, A.; et al. Oral Lactoferrin Supplementation during Induction Chemotherapy Promotes Gut Microbiome Eubiosis in Pediatric Patients with Hematologic Malignancies. Pharmaceutics 2022, 14, 1705. [Google Scholar] [CrossRef] [PubMed]
- Sortino, O.; Hullsiek, K.H.; Richards, E.; Rupert, A.; Schminke, A.; Tetekpor, N.; Quinones, M.; Prosser, R.; Schacker, T.; Sereti, I.; et al. The Effects of Recombinant Human Lactoferrin on Immune Activation and the Intestinal Microbiome Among Persons Living with Human Immunodeficiency Virus and Receiving Antiretroviral Therapy. J. Infect. Dis. 2019, 219, 1963–1968. [Google Scholar] [CrossRef]
- Konstanti, P.; van Splunter, M.; van den Brink, E.; Belzer, C.; Nauta, A.; van Neerven, R.J.; Smidt, H. The effect of nutritional intervention with lactoferrin, galactooligosacharides and vitamin D on the gut microbiota composition of healthy elderly women. Nutrients 2022, 14, 2468. [Google Scholar] [CrossRef]
- Permyakov, E.A.; Berliner, L.J. alpha-Lactalbumin: Structure and function. FEBS Lett. 2000, 473, 269–274. [Google Scholar] [CrossRef]
- Lönnerdal, B. Infant formula and infant nutrition: Bioactive proteins of human milk and implications for composition of infant formulas. Am. J. Clin. Nutr. 2014, 99, 712S–717S. [Google Scholar] [CrossRef]
- Layman, D.K.; Lönnerdal, B.; Fernstrom, J.D. Applications for α-lactalbumin in human nutrition. Nutr. Rev. 2018, 76, 444–460. [Google Scholar] [CrossRef]
- Kamau, S.M.; Cheison, S.C.; Chen, W.; Liu, X.M.; Lu, R.R. Alpha-lactalbumin: Its production technologies and bioactive peptides. Comp. Rev. Food Sci. Food Safey 2010, 9, 197–212. [Google Scholar] [CrossRef]
- Pellegrini, A.; Thomas, U.; Bramaz, N.; Hunziker, P.; von Fellenberg, R. Isolation and identification of three bactericidal domains in the bovine α-lactalbumin molecule. Biochim. Biophys. Acta 1999, 1426, 439–448. [Google Scholar] [CrossRef]
- Lajnaf, R.; Gharsallah, H.; Jridi, M.; Attia, H.; Ayadi, M.A. Antioxidant and antibacterial activities, interfacial and emulsifying properties of the apo and holo forms of purified camel and bovine α-lactalbumin. Int. J. Biol. Macromol. 2020, 165 Pt A, 205–213. [Google Scholar] [CrossRef]
- Brück, W.M.; Redgrave, M.; Tuohy, K.M.; Lönnerdal, B.; Graverholt, G.; Hernell, O.; Gibson, G.R. Effects of Bovine α-Lactalbumin and Casein Glycomacropeptide—Enriched Infant Formulae on Faecal Microbiota in Healthy Term Infants. J. Pediatr. Gastroenterol. Nutr. 2006, 43, 673–679. [Google Scholar] [CrossRef]
- Xie, D.; Shen, Y.; Su, E.; Du, L.; Xie, J.; Wei, D. Anti-Hyperuricemic, Nephroprotective, and Gut Microbiota Regulative Effects of Separated Hydrolysate of α-Lactalbumin on Potassium Oxonate- and Hypoxanthine-Induced Hyperuricemic Mice. Mol. Nutr. Food Res. 2023, 67, e2200162. [Google Scholar] [CrossRef]
- Xie, D.; Shen, Y.; Su, E.; Du, L.; Xie, J.; Wei, D. The effects of angiotensin I-converting enzyme inhibitory peptide VGINYW and the hydrolysate of α-lactalbumin on blood pressure, oxidative stress and gut microbiota of spontaneously hypertensive rats. Food Funct. 2022, 13, 2743–2755. [Google Scholar] [CrossRef]
- Chen, H.; Sun, Y.; Zhao, H.; Qi, X.; Cui, H.; Li, Q.; Ma, Y. α-Lactalbumin peptide Asp-Gln-Trp alleviates hepatic insulin resistance and modulates gut microbiota dysbiosis in high-fat diet-induced NAFLD mice. Food Funct. 2022, 13, 9878–9892. [Google Scholar] [CrossRef]
- Li, T.; Gao, J.; Duc, M.; Mao, X. Bovine α-lactalbumin hydrolysates ameliorate obesity-associated endotoxemia and inflammation in high-fat diet-fed mice through modulation of gut microbiota. Food Funct. 2019, 10, 3368–3378. [Google Scholar] [CrossRef]
- Boscaini, S.; Cabrera-Rubio, R.; Speakman, J.R.; Cotter, P.D.; Cryan, J.F.; Nilaweera, K.N. Dietary α-lactalbumin alters energy balance, gut microbiota composition and intestinal nutrient transporter expression in high-fat diet-fed mice. Br. J. Nutr. 2019, 121, 1097–1107. [Google Scholar] [CrossRef]
- Nielsen, C.H.; Hui, Y.; Nguyen, D.N.; Ahnfeldt, A.M.; Burrin, D.G.; Hartmann, B.; Heckmann, A.B.; Sangild, P.T.; Thymann, T.; Bering, S.B. Alpha-Lactalbumin Enriched Whey Protein Concentrate to Improve Gut, Immunity and Brain Development in Preterm Pigs. Nutrients 2020, 12, 245. [Google Scholar] [CrossRef]
- Wernimont, S.; Northington, R.; Kullen, M.J.; Yao, M.; Bettler, J. Effect of an α-lactalbumin-enriched infant formula supplemented with oligofructose on fecal microbiota, stool characteristics, and hydration status: A randomized, double-blind, controlled trial. Clin. Pediatr. 2015, 54, 359–370. [Google Scholar] [CrossRef] [PubMed]
- Basciani, S.; Nordio, M.; Dinicola, S.; Unfer, V.; Gnessi, L. Diet Plus Inositols, α-Lactalbumin and Gymnema sylvestre: The Successful Combo to Restore Body Weight and Metabolic Profile in Obese and Dysmetabolic Patients. Nutrients 2023, 15, 3142. [Google Scholar] [CrossRef]
- Ferraboschi, P.; Ciceri, S.; Grisenti, P. Applications of Lysozyme, an Innate Immune Defense Factor, as an Alternative Antibiotic. Antibiotics 2021, 10, 1534. [Google Scholar] [CrossRef]
- Gajda, E.; Bugla-Płoskońska, G. Lizozym—Występowanie w przyrodzie, właściwości biologiczne i możliwości zastosowań [Lysozyme—Occurrence in nature, biological properties and possible applications]. Postep. Hig. Med. Dosw. 2014, 68, 1501–1515. [Google Scholar] [CrossRef]
- Montagne, P.; Cuilliere, M.L.; Mole, C.; Bene, M.C.; Faure, G. Changes in lactoferrin and lysozyme levels in human milk during the first twelve weeks of lactation. Adv. Exp. Med. Biol. 2001, 501, 241–247. [Google Scholar]
- Piccinini, R.; Binda, E.; Belotti, M.; Casirani, G.; Zecconi, A. Comparison of blood and milk non-specific immune parameters in heifers after calving in relation to udder health. Vet. Res. 2005, 36, 747–757. [Google Scholar] [CrossRef]
- Xu, S.; Shi, J.; Shi, X.; Dong, Y.; Wu, X.; Li, Z.; Fang, Z.; Lin, Y.; Che, L.; Li, J.; et al. Effects of dietary supplementation with lysozyme during late gestation and lactation stage on the performance of sows and their offspring. J. Anim. Sci. 2018, 96, 4768–4779. [Google Scholar] [CrossRef] [PubMed]
- Larsen, I.S.; Jensen, B.A.H.; Bonazzi, E.; Choi, B.S.Y.; Kristensen, N.N.; Schmidt, E.G.W.; Süenderhauf, A.; Morin, L.; Olsen, P.B.; Hansen, L.B.S.; et al. Fungal lysozyme leverages the gut microbiota to curb DSS-induced colitis. Gut Microbes. 2021, 13, 1988836. [Google Scholar] [CrossRef] [PubMed]
- Sakurai, T.; Hashikura, N.; Minami, J.; Yamada, A.; Odamaki, T.; Xiao, J.Z. Tolerance mechanisms of human-residential bifidobacteria against lysozyme. Anaerobe 2017, 47, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Lu, D.; Li, Q.; Wu, Z.; Shang, S.; Liu, S.; Wen, X.; Li, Z.; Wu, F.; Li, N. High-level recombinant human lysozyme expressed in milk of transgenic pigs can inhibit the growth of Escherichia coli in the duodenum and influence intestinal morphology of sucking pigs. PLoS ONE 2014, 9, e89130. [Google Scholar] [CrossRef] [PubMed]
- Minami, J.; Odamaki, T.; Hashikura, N.; Abe, F.; Xiao, J.Z. Lysozyme in breast milk is a selection factor for bifidobacterial colonisation in the infant intestine. Benef. Microbes. 2016, 7, 53–60. [Google Scholar] [CrossRef]
- Zhou, J.; Xiong, X.; Yin, J.; Zou, L.; Wang, K.; Shao, Y.; Yin, Y. Dietary Lysozyme Alters Sow’s Gut Microbiota, Serum Immunity and Milk Metabolite Profile. Front. Microbiol. 2019, 10, 177. [Google Scholar] [CrossRef]
- Du, M.; Xie, X.; Yang, S.; Li, Y.; Jiang, T.; Yang, J.; Li, L.; Huang, Y.; Wu, Q.; Chen, W.; et al. Lysozyme-like Protein Produced by Bifidobacterium longum Regulates Human Gut Microbiota Using In Vitro Models. Molecules 2021, 26, 6480. [Google Scholar] [CrossRef]
- Maga, E.A.; Walker, R.L.; Anderson, G.B.; Murray, J.D. Consumption of milk from transgenic goats expressing human lysozyme in the mammary gland results in the modulation of intestinal microflora. Transgenic Res. 2006, 15, 515–519. [Google Scholar] [CrossRef]
- Cooper, C.A.; Brundige, D.R.; Reh, W.A.; Maga, E.A.; Murray, J.D. Lysozyme transgenic goats’ milk positively impacts intestinal cytokine expression and morphology. Transgenic Res. 2011, 20, 1235–1243. [Google Scholar] [CrossRef] [PubMed]
- Cooper, C.A.; Garas Klobas, L.C.; Maga, E.A.; Murray, J.D. Consuming transgenic goats’ milk containing the antimicrobial protein lysozyme helps resolve diarrhea in young pigs. PLoS ONE 2013, 8, e58409. [Google Scholar] [CrossRef]
- Maga, E.A.; Desai, P.T.; Weimer, B.C.; Dao, N.; Kültz, D.; Murray, J.D. Consumption of lysozyme-rich milk can alter microbial fecal populations. Appl. Environ. Microbiol. 2012, 78, 6153–6160. [Google Scholar] [CrossRef]
- Cui, C.; Wang, F.; Zheng, Y.; Wei, H.; Peng, J. From birth to death: The hardworking life of Paneth cell in the small intestine. Front. Immunol. 2023, 14, 1122258. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, X.; Zuo, Z.; Zhang, Q.; Pan, Y.; Zeng, B.; Li, W.; Wei, H.; Liu, Z. Rip2 Is Required for Nod2-Mediated Lysozyme Sorting in Paneth Cells. J. Immunol. 2017, 198, 3729–3736. [Google Scholar] [CrossRef]
- Strigli, A.; Gopalakrishnan, S.; Zeissig, Y.; Basic, M.; Wang, J.; Schwerd, T.; Doms, S.; Peuker, K.; Hartwig, J.; Harder, J.; et al. Deficiency in X-linked inhibitor of apoptosis protein promotes susceptibility to microbial triggers of intestinal inflammation. Sci. Immunol. 2021, 6, eabf7473. [Google Scholar] [CrossRef]
- Gaudino, S.J.; Beaupre, M.; Lin, X.; Joshi, P.; Rathi, S.; McLaughlin, P.A.; Kempen, C.; Mehta, N.; Eskiocak, O.; Yueh, B.; et al. IL-22 receptor signaling in Paneth cells is critical for their maturation, microbiota colonization, Th17-related immune responses, and anti-Salmonella immunity. Mucosal Immunol. 2021, 14, 389–401. [Google Scholar] [CrossRef]
- Cui, C.; Wang, X.; Zheng, Y.; Li, L.; Wang, F.; Wei, H.; Peng, J. Paneth cells protect intestinal stem cell niche to alleviate deoxynivalenol-induced intestinal injury. Ecotoxicol. Environ. Saf. 2023, 264, 115457. [Google Scholar] [CrossRef]
- Córdova-Dávalos, L.E.; Jiménez, M.; Salinas, E. Glycomacropeptide Bioactivity and Health: A Review Highlighting Action Mechanisms and Signaling Pathways. Nutrients 2019, 11, 598. [Google Scholar] [CrossRef]
- Thomä-Worringer, C.; Sørensen, J.; López-Fandiño, R. Health effects and technological features of caseinomacropeptide. Int. Dairy J. 2006, 16, 1324–1333. [Google Scholar] [CrossRef]
- Furlanetti, A.M.; Prata, L.F. Free and total GMP (glycomacropeptide) contents of milk during bovine lactation. Ciênc Tecnol. Aliment. 2003, 23, 121–125. [Google Scholar] [CrossRef]
- Otani, H.; Monnai, M.; Kawasaki, Y.; Kawakami, H.; Tanimoto, M. Inhibition of mitogen-induced proliferative responses of lymphocytes by bovine κ-caseinoglycopeptides having different carbohydrate chains. J. Dairy Res. 1995, 62, 349–357. [Google Scholar] [CrossRef]
- Neelima, S.R.; Rajput, Y.S.; Mann, B. Chemical and functional properties of glycomacropeptide (GMP) and its role in the detection of cheese whey adulteration in milk: A review. Dairy Sci. Technol. 2013, 93, 21–43. [Google Scholar] [CrossRef]
- Sawin, E.A.; De Wolfe, T.J.; Aktas, B.; Stroup, B.M.; Murali, S.G.; Steele, J.L.; Ney, D.M. Glycomacropeptide is a prebiotic that reduces Desulfovibrio bacteria, increases cecal short-chain fatty acids, and is anti-inflammatory in mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2015, 309, G590–G601. [Google Scholar] [CrossRef]
- Azuma, N.; Yamauchi, K.; Mitsuoka, T. Bifidus growth-promoting activity of a glycomacropeptide derived from human K-casein. Agric. Biol. Chem. 1984, 48, 2159–2162. [Google Scholar] [CrossRef]
- O’Riordan, N.; O’Callaghan, J.; Buttò, L.F.; Kilcoyne, M.; Joshi, L.; Hickey, R.M. Bovine glycomacropeptide promotes the growth of Bifidobacterium longum ssp. infantis and modulates its gene expression. J. Dairy Sci. 2018, 101, 6730–6741. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Cao, J.; Jia, Y.; Liu, X.; Yan, Y.; Pang, G. Modulation of mice fecal microbiota by administration of casein glycomacropeptide. Microbiol. Res. 2012, 3, e3. [Google Scholar] [CrossRef]
- Jiménez, M.; Cervantes-García, D.; Muñoz, Y.H.; García, A.; Haro, L.M., Jr.; Salinas, E. Novel Mechanisms Underlying the Therapeutic Effect of Glycomacropeptide on Allergy: Change in Gut Microbiota, Upregulation of TGF-β, and Inhibition of Mast Cells. Int. Arch. Allergy Immunol. 2016, 171, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Q.; Zhan, B.; Chang, R.; Du, M.; Mao, X. Antidiabetic Effect of Casein Glycomacropeptide Hydrolysates on High-Fat Diet and STZ-Induced Diabetic Mice via Regulating Insulin Signaling in Skeletal Muscle and Modulating Gut Microbiota. Nutrients 2020, 12, 220. [Google Scholar] [CrossRef]
- Ntemiri, A.; Chonchúir, F.N.; O’Callaghan, T.F.; Stanton, C.; Ross, R.P.; O’Toole, P.W. Glycomacropeptide Sustains Microbiota Diversity and Promotes Specific Taxa in an Artificial Colon Model of Elderly Gut Microbiota. J. Agric. Food Chem. 2017, 65, 1836–1846. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Xing, Y.; Liu, H.; Chang, Y.; You, Y.; Dou, Y.; Liu, B.; Wang, Q.; Ma, D.; Chen, L.; et al. Effects of a Formula with scGOS/lcFOS (9:1) and Glycomacropeptide (GMP) Supplementation on the Gut Microbiota of Very Preterm Infants. Nutrients 2022, 14, 1901. [Google Scholar] [CrossRef]
- Hansen, K.E.; Murali, S.; Chaves, I.Z.; Suen, G.; Ney, D.M. Glycomacropeptide Impacts Amylin-Mediated Satiety, Postprandial Markers of Glucose Homeostasis, and the Fecal Microbiome in Obese Postmenopausal Women. J. Nutr. 2023, 153, 1915–1929. [Google Scholar] [CrossRef]
- Wernlund, P.G.; Hvas, C.L.; Dahlerup, J.F.; Bahl, M.I.; Licht, T.R.; Knudsen, K.E.B.; Agnholt, J.S. Casein Glycomacropeptide Is Well Tolerated in Healthy Adults and Changes Neither High-Sensitive C-Reactive Protein, Gut Microbiota nor Faecal Butyrate: A Restricted Randomised Trial. Br. J. Nutr. 2021, 125, 1374–1385. [Google Scholar] [CrossRef]
- Montanari, C.; Ceccarani, C.; Corsello, A.; Zuvadelli, J.; Ottaviano, E.; Dei Cas, M.; Banderali, G.; Zuccotti, G.; Borghi, E.; Verduci, E. Glycomacropeptide Safety and Its Effect on Gut Microbiota in Patients with Phenylketonuria: A Pilot Study. Nutrients 2022, 14, 1883. [Google Scholar] [CrossRef]
- Qu, Y.; Park, S.H.; Dallas, D.C. Evaluating the Potential of Casein Glycomacropeptide in Adult Irritable Bowel Syndrome Management: A Pilot Study. Nutrients 2023, 15, 4174. [Google Scholar] [CrossRef]
Figure 1.
Prebiotic activity of lactoferrin: mechanisms of action. Both direct and indirect prebiotic effects have been described. The indirect mechanism is due to the lactoferrin-mediated iron sequestering that leads to a selective inhibition of the growth of pathogens, promoting the growth of those probiotics, such as Bifidobacterium and Lactobacillus, which do not necessarily need iron to grow. In the direct mechanism, lactoferrin promotes the growth of probiotics (especially Bifidobacterium) by acting as β-N-glycans and iron supplier, or by interacting with the lactoferrin-binding proteins present on the membrane of some probiotics, including Bifidobacterium and Lactobacillus. This interaction has been shown to promote the cytosolic internalization of lactoferrin which has been suggested to modulate genes involved in DNA replication and cell growth.
Figure 1.
Prebiotic activity of lactoferrin: mechanisms of action. Both direct and indirect prebiotic effects have been described. The indirect mechanism is due to the lactoferrin-mediated iron sequestering that leads to a selective inhibition of the growth of pathogens, promoting the growth of those probiotics, such as Bifidobacterium and Lactobacillus, which do not necessarily need iron to grow. In the direct mechanism, lactoferrin promotes the growth of probiotics (especially Bifidobacterium) by acting as β-N-glycans and iron supplier, or by interacting with the lactoferrin-binding proteins present on the membrane of some probiotics, including Bifidobacterium and Lactobacillus. This interaction has been shown to promote the cytosolic internalization of lactoferrin which has been suggested to modulate genes involved in DNA replication and cell growth.

Table 3.
In vivo studies and clinical trials on the modulation of gut microbiota by α-lactalbumin.
| Patients or Animal Models | Treatment | Effects on Gut Microbiota | Health Effects | Type of Study | Ref. |
|---|---|---|---|---|---|
| Potassium Oxonate- and hypoxanthine-induced hyperuricemic mice. | Oral supplementation of α-lactalbumin hydrolysates. | Increases the abundance of some SCFA-producing bacteria; decreases the growth of hyperuricaemia- and inflammation-associated genera. | Reduction in the levels of serum uric acid, creatinine, and urea nitrogen. | In vivo | [177] |
| Spontaneously hypertensive rats. | Oral gavage of α-lactalbumin hydrolysates under 3 kDa (100 mg/kg BW) and VGINYW (5 mg per kg BW). | Recovers the diversity of the gut microbiota and modulates short-chain fatty acid producing bacteria. | Alleviates the hypertension-associated intestinal microbiota dysbiosis. | In vivo | [178] |
| High-fat diet-induced NAFLD mice. | Treatment with the α-lactalbumin peptide Asp-Gln-Trp. | Modulates the gut microbiota increasing the ratio of Bacteroides to Firmicutes, reducing the relative abundance of pathogenic bacteria (i.e., Bacteroides, Blautia, and Alistipes) and enhancing the relative abundance of short-chain fatty acid (SCFA)-producing bacteria (i.e., Muribaculaceae, Lachnospiraceae, and Rikenellaceae). | Improves the intestinal barrier integrity and inflammation. | In vivo | [179] |
| High-fat diet-induced obese mice. | Supplementation of bovine α-lactalbumin hydrolysate. | Modulates the Bacteroidetes/Firmicutes ratios and increases the relative abundance of Lachnospiraceae and Blautia. | Reduces the levels of inflammatory cytokines, such as interleukin-6 and tumour necrosis factor-α, and of lipopolysaccharides. Reduces the obesity-associated systematic inflammation and endotoxaemia. | In vivo | [180] |
| High-fat diet-fed mice. | Supplementation of dietary α-lactalbumin. | Favours the abundance of the Lactobacillus, Parabacteroides and Bifidobacterium genera. | Decrease in cd36 and glut2 gene expression in the intestine accompanied by increased cumulative energy ingestion. | In vivo | [181] |
| Preterm pigs (model of newborn infants). | High α-lactalbumin diet. | Improves the growth of Clostridiaceae, Enterobacteriaceae, Streptococcus, and Streptomyces. | Produces beneficial effects on the growth and functional development of body and organ systems, gut, immunity, and structural brain development. | In vivo | [182] |
| Full-term infants. | α-Lactalbumin enriched term infant formula plus 3.0 g/L of oligofructose. | Increases the faecal bifidobacteria. | Improves the stool consistency without adversely affecting stool frequency or hydration. | Clinical trial | [183] |
| Obese and dysmetabolic patients. | Oral supplementation of a combination of inositols, α-lactalbumin, and Gymnema sylvestre. | Recovers gut dysbiosis. | Positive impact on insulin, glucose, lipid metabolism. | Clinical trial | [184] |
Table 4.
In vivo studies on the modulation of gut microbiota by lysozyme.
| Animal Models | Treatment | Effects on Gut Microbiota | Health Effects | Type of Study | Ref. |
|---|---|---|---|---|---|
| Young pigs. | Milk from transgenic goats expressing human-comparable levels of human lysozyme. | Reduction of coliforms and E. coli. | Reduction in the serum levels of uric acid, creatinine, and urea nitrogen. | In vivo | [196] |
| Young pigs. | Milk from transgenic goats expressing human-comparable levels of human lysozyme. | Inhibition of pathogenic E. coli. Decrease of Firmicutes and increase in Bacteroidetes; reduction of Clostridia, Streptococcaceae, Mycobacteriaceae, and Campylobacterales. | Ameliorates intestinal inflammation; accelerates recovery from E. coli-induced diarrhoea. Improved gut health. | In vivo | [197,198,199] |
| Mice with deoxynivalenol-induced depletion of Paneth cells. | Supplementation with lysozyme (200 U/day). | Reduces the Firmicutes/Bacteroidetes ratio; increases the abundance of Dubosiella and decreases the abundance of Lactobacillus. | Improved Paneth cells’ functionality and gut microbiota restoration. | In vivo | [204] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Gallo, V.; Arienzo, A.; Tomassetti, F.; Antonini, G. Milk Bioactive Compounds and Gut Microbiota Modulation: The Role of Whey Proteins and Milk Oligosaccharides. Foods 2024, 13, 907. https://doi.org/10.3390/foods13060907
AMA Style
Gallo V, Arienzo A, Tomassetti F, Antonini G. Milk Bioactive Compounds and Gut Microbiota Modulation: The Role of Whey Proteins and Milk Oligosaccharides. Foods. 2024; 13(6):907. https://doi.org/10.3390/foods13060907
Chicago/Turabian StyleGallo, Valentina, Alyexandra Arienzo, Federica Tomassetti, and Giovanni Antonini. 2024. "Milk Bioactive Compounds and Gut Microbiota Modulation: The Role of Whey Proteins and Milk Oligosaccharides" Foods 13, no. 6: 907. https://doi.org/10.3390/foods13060907
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.