Inhibitory Effects of Jiuzao Polysaccharides on Alcoholic Fatty Liver Formation in Zebrafish Larvae and Their Regulatory Impact on Intestinal Microbiota

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Chemicals
2.2. Preparation of Polysaccharides
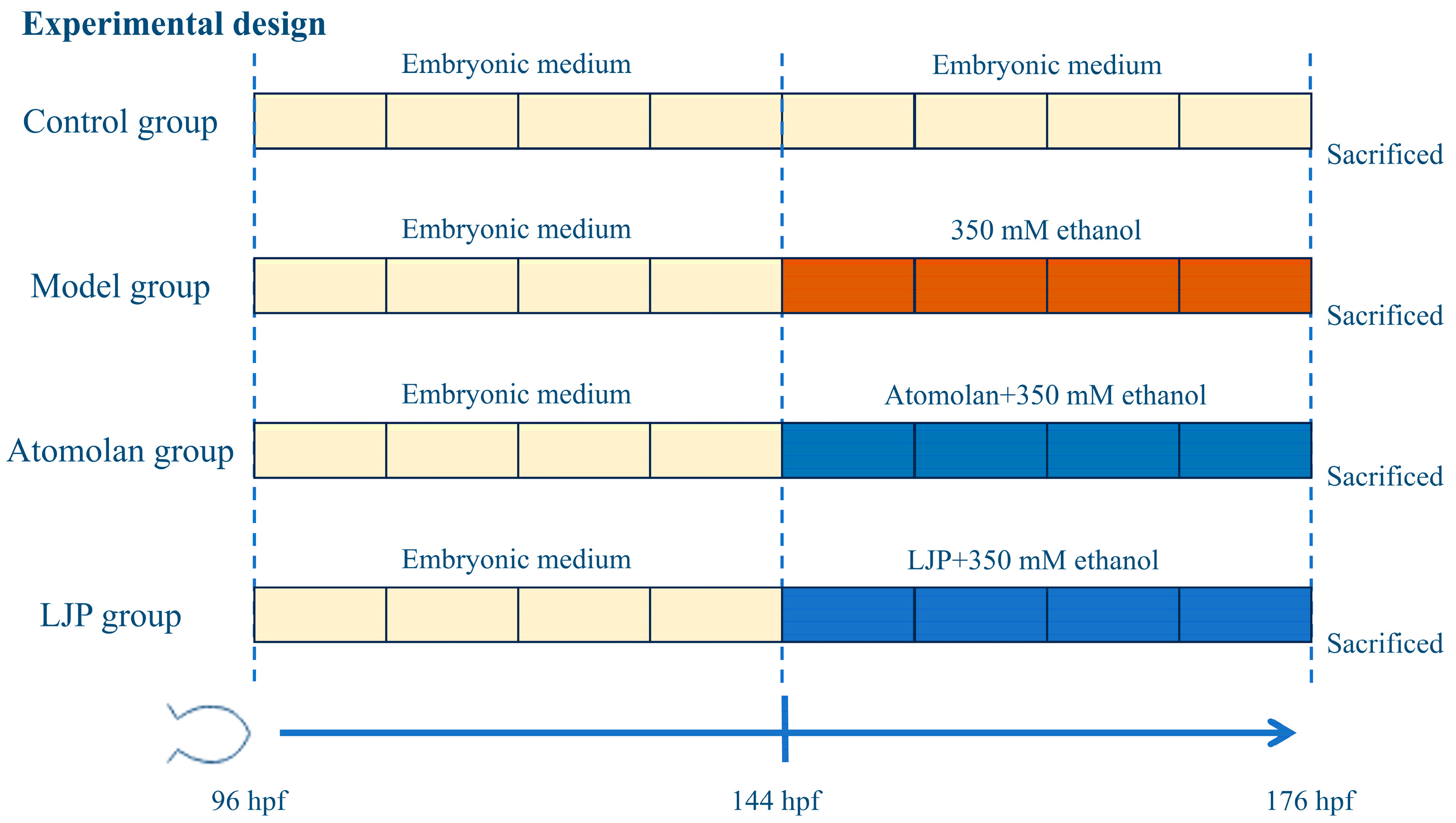
2.3. Animals and Experimental Design
2.4. Oil Red O Staining
2.5. Hematoxylin and Eosin Stainings
2.6. Nile Red and DAPI Stainings
2.7. Enzymatic Assays in Ethanol-Induced Fatty Liver Disease in Zebrafish Larvae
2.8. Tissue RNA Extraction, Library Construction, and Transcriptome Sequencing
2.9. Real-Time Quantitative Polymerase Chain Reaction (RT-qPCR) Analysis
2.10. 16 S rRNA Gene Sequencing of Intestinal Bacteria
2.11. Statistical Analysis
3. Results
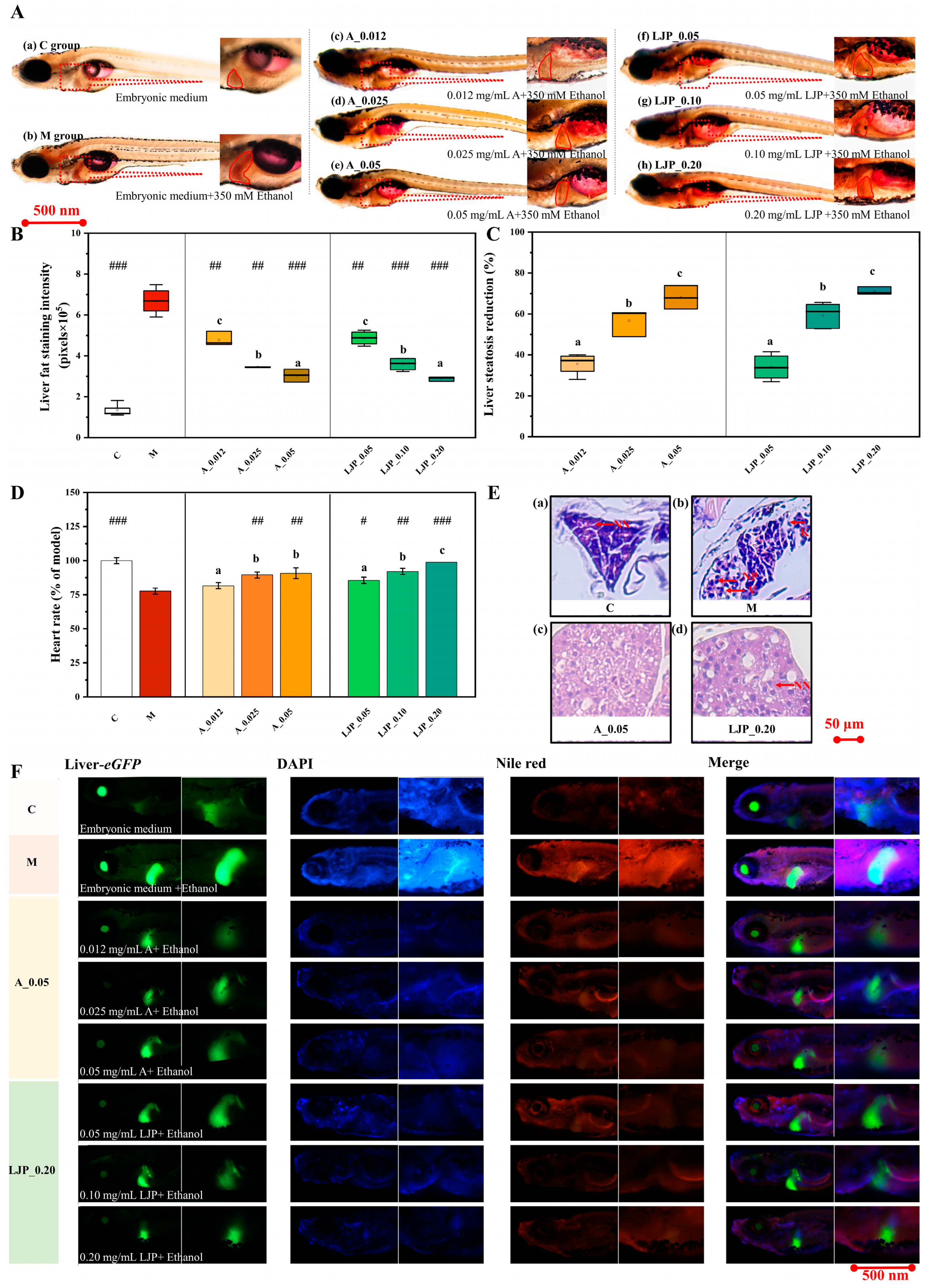
3.1. Effect of LJP on Liver Visual Phenotype in AFLD Zebrafish Larvae
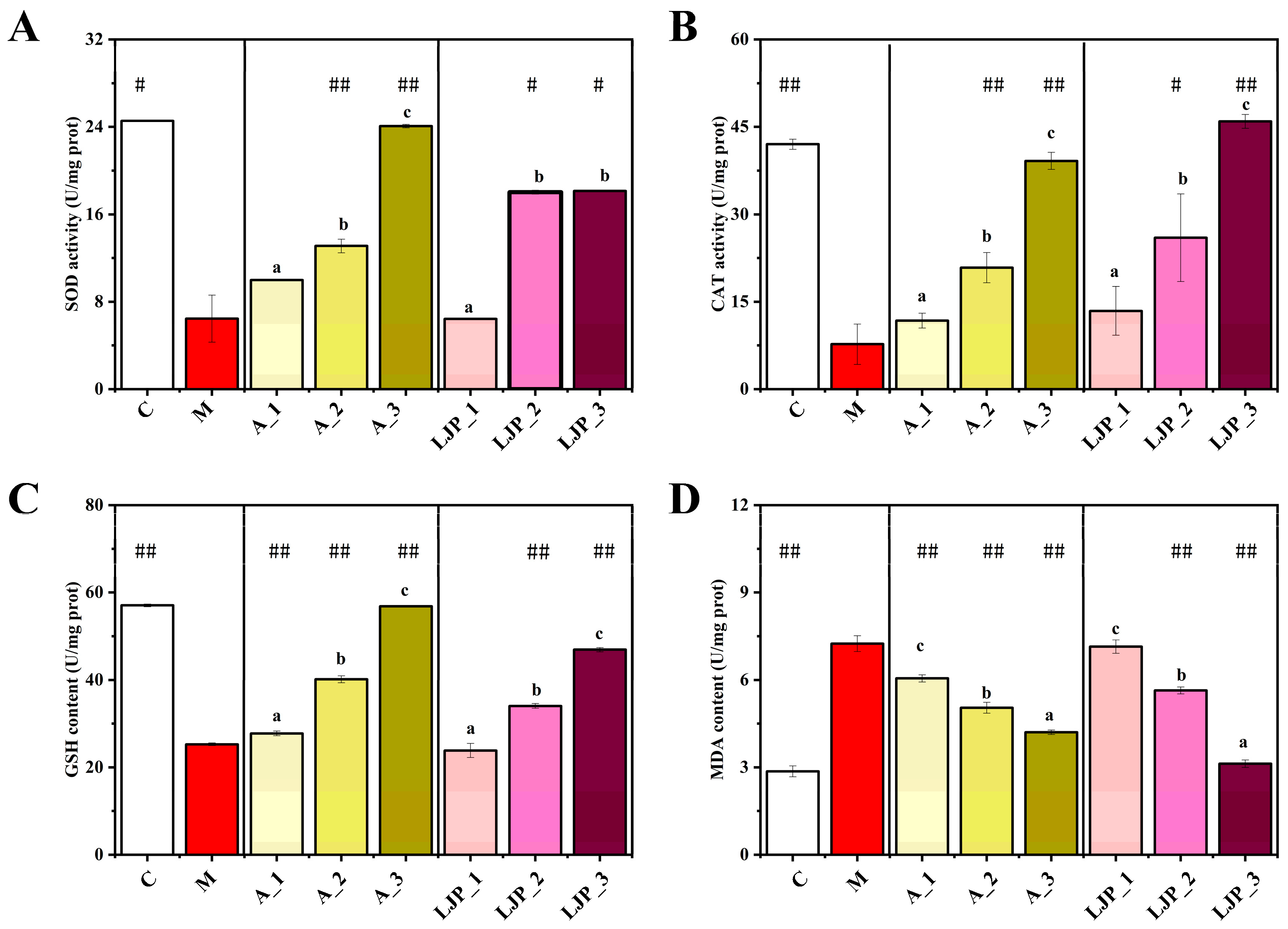
3.2. Effect of LJP on Oxidative Stress and Lipid Peroxidation Indices in AFLD Zebrafish Larvae
3.3. Effect of LJP on the Expression of Liver Lipid-Related Genes in AFLD Zebrafish Larvae
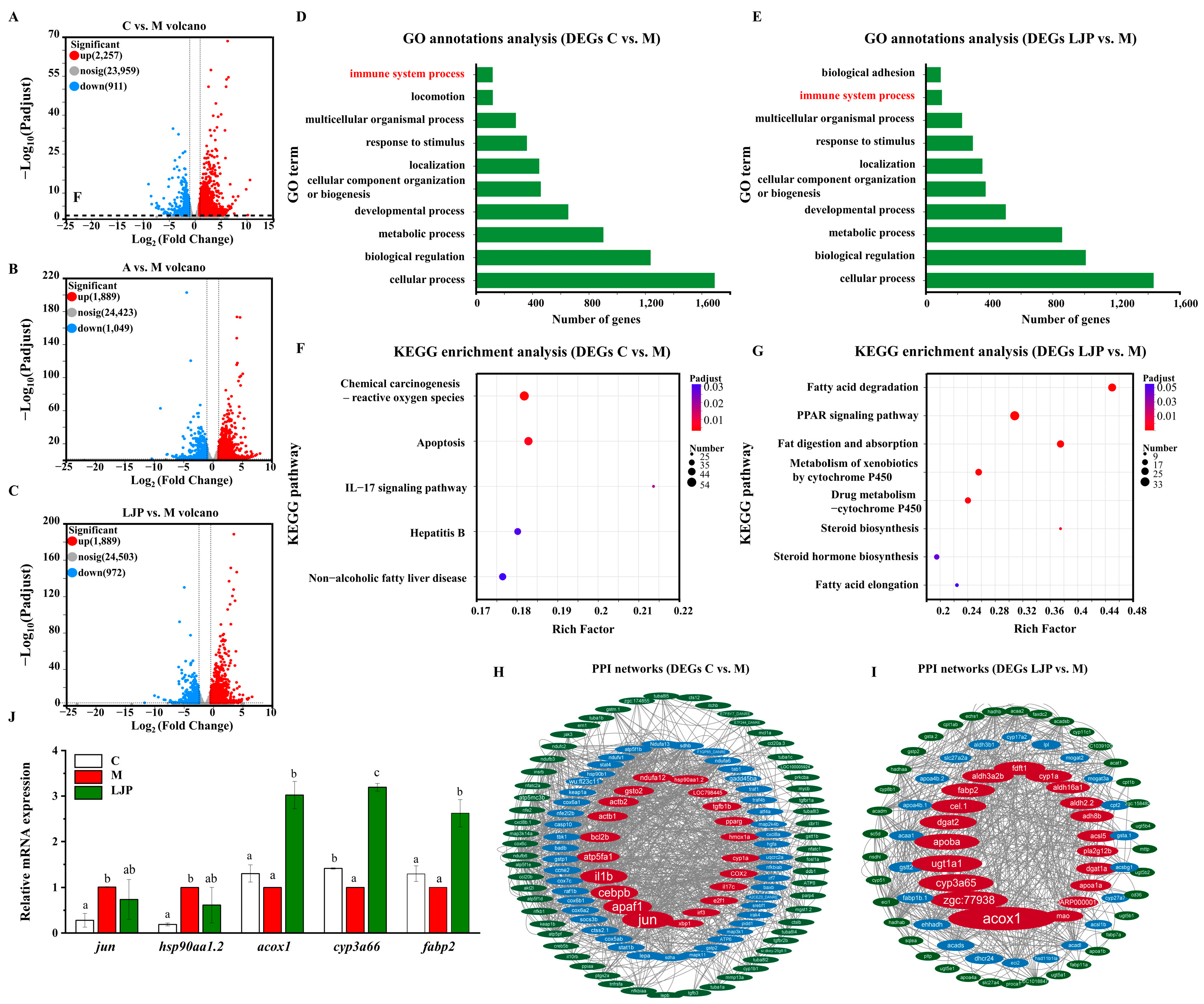
3.3.1. Differentially Expressed Gene Profiles
3.3.2. Gene Annotation and Functional Analysis
3.3.3. Protein-Protein Interaction (PPI) Network for Lipid Metabolism-Related Genes
3.3.4. qRT-PCR Validation Analysis of DEGs
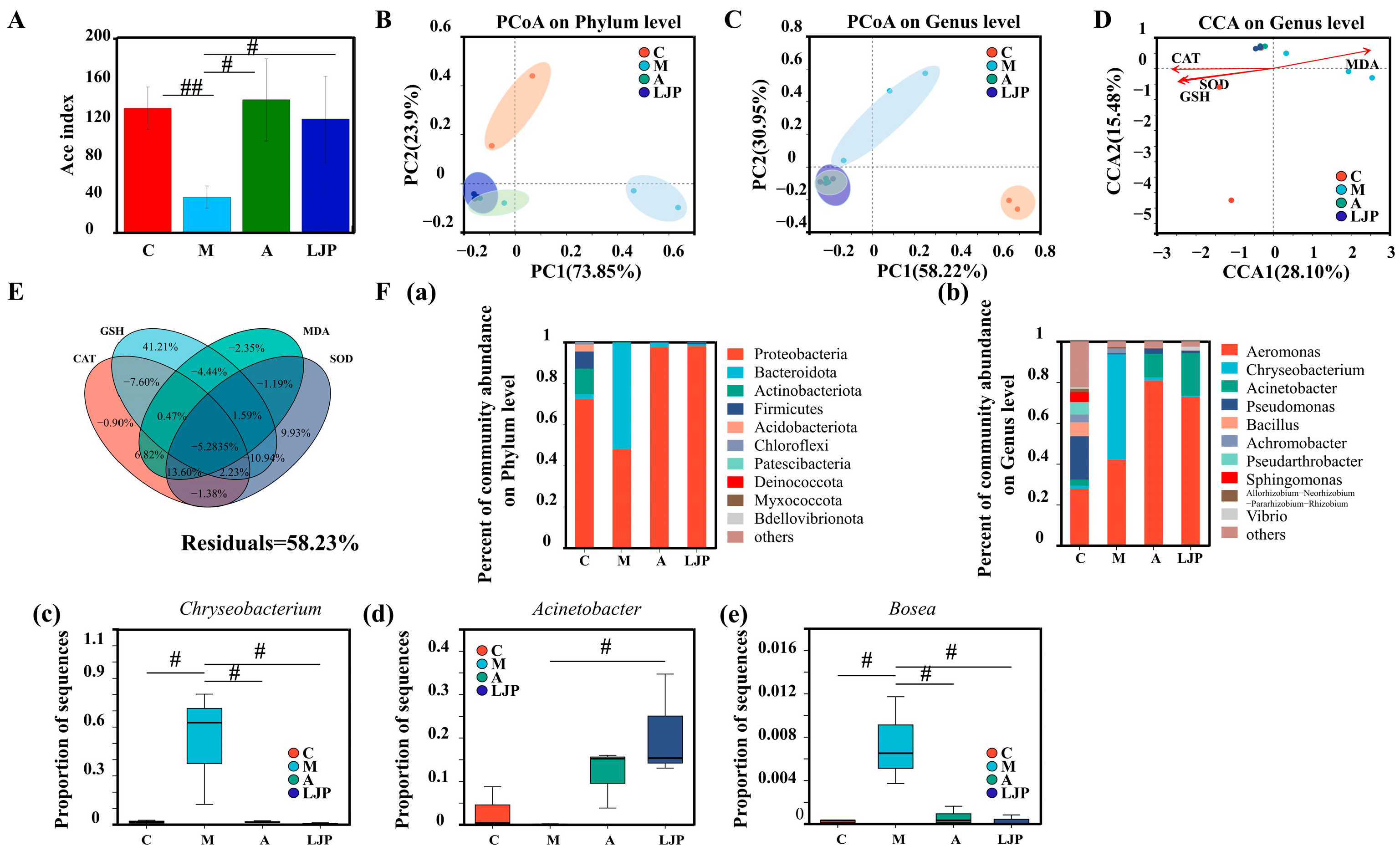
3.4. Effect of LJP on the Intestinal Bacteria of AFLD Zebrafish Larvae
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Altura, B.M.; Altura, B.T. Association of Alcohol in Brain Injury, Headaches, and Stroke with Brain-Tissue and Serum Levels of Ionized Magnesium: A Review of Recent Findings and Mechanisms of Action. Alcohol 1999, 19, 119–130. [Google Scholar] [CrossRef] [PubMed]
- Chu, L.; Wallach, J.D. Consideration of Confounding in Epidemiologic Studies Assessing Alcohol Consumption on the Risk of Breast Cancer: A Brief Report. Chem. Biol. Interact. 2020, 322, 109060. [Google Scholar] [CrossRef] [PubMed]
- Buyco, D.G.; Martin, J.; Jeon, S.; Hooks, R.; Lin, C.; Carr, R. Experimental Models of Metabolic and Alcoholic Fatty Liver Disease. World J. Gastroenterol. 2021, 126, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Ji, X.; Peng, Q.; Yuan, Y.; Shen, J.; Xie, X.; Wang, M. Isolation, Structures and Bioactivities of the Polysaccharides from Jujube Fruit (Ziziphus jujuba Mill.): A Review. Food Chem. 2017, 227, 349–357. [Google Scholar] [CrossRef] [PubMed]
- Shin, D.; Hyun Shin, C.; Tucker, J.; Ober, E.A.; Rentzsch, F.; Poss, K.D.; Hammerschmidt, M.; Mullins, M.C.; Stanier, D.Y.R. Bmp and Fgf Signaling Are Essential for Liver Specification in Zebrafish. Development 2007, 134, 2041–2050. [Google Scholar] [CrossRef]
- Ye, L.; Cheng, B.; Ma, S.; Hao, J.; Meng, G.; Fu, J.; Wu, X. Preventive Effect and Mechanism of Ganoderma Lingzhi Polysaccharides on Acute Alcoholic Liver Injury in Mice. Food Sci. 2022, 43, 103–110. [Google Scholar] [CrossRef]
- Zhang, X.; Xiao, J.; Guo, Z.; Zhong, H.; Luo, Y.; Wang, J.; Tang, Z.; Huang, T.; Li, M.; Zhu, J.; et al. Genomics Transcriptomics Integrated with Metabolomics Reveals the Effect of Lycium Barbarum Polysaccharide on Apoptosis in Nile Tilapia (Oreochromis Niloticus). Food Chem. 2022, 114, 229–240. [Google Scholar] [CrossRef]
- Zhai, X.; Zhu, C.; Zhang, Y.; Sun, J.; Alim, A.; Yang, X. Chemical Characteristics, Antioxidant Capacities and Hepatoprotection of Polysaccharides from Pomegranate Peel. Carbohydr. Polym. 2018, 202, 461–469. [Google Scholar] [CrossRef]
- Chen, Q.; Wang, Y.; Yin, N.; Wang, R.; Zheng, Y.; Yang, Y.; An, X.; Qi, J. Polysaccharides from Fermented Wheat Bran Enhanced the Growth Performance of Zebrafish (Danio rerio) through Improving Gut Microflora and Antioxidant Status. Aquac. Rep. 2022, 25, 101188. [Google Scholar] [CrossRef]
- Li, Q.; Geng, X.; Zhu, L.; Zheng, F.; Chen, E.; Wang, G.; Li, X. Structural Characterization and Antioxidant Properties of a Novel Polysaccharide Isolated from Jiuzao In Vitro and In Vivo. Food Res. Int. 2022, 162, 111940. [Google Scholar] [CrossRef]
- Passeri, M.J.; Cinaroglu, A.; Gao, C.; Sadler, K.C. Hepatic Steatosis in Response to Acute Alcohol Exposure in Zebrafish Requires Sterol Regulatory Element Binding Protein Activation. Hepatology 2009, 49, 443–452. [Google Scholar] [CrossRef] [PubMed]
- Cao, Q.; Shan, H.; Zhao, J.; Deng, J.; Xu, M.; Kang, H.; Li, T.; Zhao, Y.; Liu, H.; Jiang, J. Liver Fibrosis in Fish Research: From an Immunological Perspective. Fish Shellfish Immunol. 2023, 139, 108885. [Google Scholar] [CrossRef] [PubMed]
- Howarth, D.L.; Passeri, M.; Sadler, K.C. Drinks like a fish: Using zebrafish to understand alcoholic liver disease. Alcohol. Clin. Exp. Res. 2011, 35, 826–829. [Google Scholar] [CrossRef]
- Jang, Z.H.; Chung, H.C.; Ahn, Y.G.; Kwon, Y.K.; Kim, J.S.; Ryu, J.H.; Ryu, D.H.; Kim, C.H.; Hwang, G.S. Metabolic Profiling of an Alcoholic Fatty Liver in Zebrafish (Danio rerio). Mol. Biosyst. 2012, 8, 2001–2009. [Google Scholar] [CrossRef] [PubMed]
- Valdes, A.M.; Walter, J.; Segal, E.; Spector, T.D. Role of the Gut Microbiota in Nutrition and Health. BMJ 2018, 361, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Cesaro, C.; Tiso, A.; Del Prete, A.; Cariello, R.; Tuccillo, C.; Cotticelli, G.; Del Vecchio Blanco, C.; Loguercio, C. Gut Microbiota and Probiotics in Chronic Liver Diseases. Dig. Liver Dis. 2011, 43, 431–438. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wu, S.; Wu, R.; Ai, H.; Lu, X.; Wang, J.; Luo, Y.; Li, L.; Cao, J. Essential Oil from Artemisia argyi Alleviated Liver Disease in Zebrafish (Danio rerio) via the Gut-Liver Axis. Fish Shellfish Immunol. 2023, 140, 108962. [Google Scholar] [CrossRef] [PubMed]
- Rao, R.K. Acetaldehyde-induced barrier disruption and paracellular permeability in Caco-2 cell monolayer. Methods Mol. Biol. 2008, 447, 171–183, ISBN 9781607617372. [Google Scholar]
- Visapää, J.P.; Tillonen, J.; Salaspuro, M. Microbes and Mucosa in the Regulation of Intracolonic Acetaldehyde Concentration during Ethanol Challenge. Alcohol Alcohol. 2002, 37, 322–326. [Google Scholar] [CrossRef]
- Wang, X.; Song, X.; Zhu, L.; Zheng, F.; Sun, B.; Li, Z.; Zhang, Y. Difference and its change rule of contents of main flavor compounds in base Laobaigan Baijiu among production batches. J. Food Sci. Tech. 2021, 39, 125–134. [Google Scholar] [CrossRef]
- Fan, Q.; Wang, X.; Zheng, F.; Li, H.; Sun, B.; Zhang, Y.; Zhang, F. Analysis of volatiles of Laobaigan base baijiu fermented by Laowuzeng and Sanpaijing processes. J. Food Sci. Tech. 2019, 37, 50–63. [Google Scholar] [CrossRef]
- Zhou, Z.; Zhong, W.; Lin, H.; Huang, P.; Ma, N.; Zhang, Y.; Zhou, C.; Lai, Y.; Huang, S.; Huang, S.; et al. Hesperidin Protects against Acute Alcoholic Injury through Improving Lipid Metabolism and Cell Damage in Zebrafish Larvae. Evid.-Based Complement. Altern. Med. 2017, 2017, 7282653. [Google Scholar] [CrossRef]
- Xu, H.; Jiang, Y.; Miao, X.M.; Tao, Y.X.; Xie, L.; Li, Y. A Model Construction of Starvation Induces Hepatic Steatosis and Transcriptome Analysis in Zebrafish Larvae. Biology. 2021, 10, 92–109. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Dewey, C.N. RSEM: Accurate Transcript Quantification from RNA-Seq Data with or without a Reference Genome. BMC Bioinform. 2014, 12, 323–338. [Google Scholar] [CrossRef]
- Xie, C.; Mao, X.; Huang, J.; Ding, Y.; Wu, J.; Dong, S.; Kong, L.; Gao, G.; Li, C.Y.; Wei, L. KOBAS 2.0: A Web Server for Annotation and Identification of Enriched Pathways and Diseases. Nucleic Acids Res. 2011, 39, 316–322. [Google Scholar] [CrossRef] [PubMed]
- Okazaki, F.; Zang, L.; Nakayama, H.; Chen, Z.; Gao, Z.J.; Chiba, H.; Hui, S.P.; Aoki, T.; Nishimura, N.; Shimada, Y. Microbiome Alteration in Type 2 Diabetes Mellitus Model of Zebrafish. Sci. Rep. 2019, 9, 867. [Google Scholar] [CrossRef] [PubMed]
- Purohit, V.; Gao, B.; Song, B.J. Molecular Mechanisms of Alcoholic Fatty Liver. Alcohol. Clin. Exp. Res. 2009, 33, 191–205. [Google Scholar] [CrossRef]
- Zhang, W.; Zhou, L.; Yin, P.; Wang, J.; Lu, X.; Wang, X.; Chen, J.; Lin, X.; Xu, G. A Weighted Relative Difference Accumulation Algorithm for Dynamic Metabolomics Data: Long-Term Elevated Bile Acids Are Risk Factors for Hepatocellular Carcinoma. Sci. Rep. 2015, 5, 191–205. [Google Scholar] [CrossRef]
- Samojlik, I.; Lakić, N.; Mimica-Dukić, N.; Daković-Švajcer, K.; Božin, B. Antioxidant and Hepatoprotective Potential of Essential Oils of Coriander (Coriandrum sativum L.) and Caraway (Carum carvi L.) (Apiaceae). J. Agric. Food Chem. 2010, 58, 8848–8853. [Google Scholar] [CrossRef]
- Martin, G.; Schoonjans, K.; Lefebvre, A.M.; Staels, B.; Auwerx, J. Coordinate Regulation of the Expression of the Fatty Acid Transport Protein and Acyl-CoA Synthetase Genes by PPARα and PPARγ Activators. J. Biol. Chem. 1997, 272, 28210–28217. [Google Scholar] [CrossRef]
- Tseng, H.P.; Hseu, T.H.; Buhler, D.R.; Wang, W.D.; Hu, C.H. Constitutive and Xenobiotics-Induced Expression of a Novel CYP3A Gene from Zebrafish Larva. Toxicol. Appl. Pharmacol. 2005, 205, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Boord, J.B.; Fazio, S.; Linton, M.F. Cytoplasmic Fatty Acid-Binding Proteins: Emerging Roles in Metabolism and Atherosclerosis. Curr. Opin. Lipidol. 2002, 13, 141–147. [Google Scholar] [CrossRef]
- Berger, J.P.; Akiyama, T.E.; Meinke, P.T. PPARs: Therapeutic Targets for Metabolic Disease. Trends Pharmacol. Sci. 2005, 26, 244–251. [Google Scholar] [CrossRef] [PubMed]
- Peyrin-Biroulet, L.; Beisner, J.; Wang, G.; Nuding, S.; Oommen, S.T.; Kelly, D.; Parmentier-Decrucq, E.; Dessein, R.; Merour, E.; Chavatte, P.; et al. Peroxisome Proliferator-Activated Receptor Gamma Activation Is Required for Maintenance of Innate Antimicrobial Immunity in the Colon. Proc. Natl. Acad. Sci. USA 2010, 107, 8772–8777. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Wang, N.; Huang, L.; Zhang, X.; Feng, S.; Liu, S.; Wang, Y.; Liu, Z. Changes in the Mucosa-Associated Microbiome and Transcriptome across Gut Segments Are Associated with Obesity in a Metabolic Syndrome Porcine Model. Microbiol. Spectr. 2022, 10, e00717-22. [Google Scholar] [CrossRef]
- Liu, Y.S.; Yuan, M.H.; Zhang, C.Y.; Liu, H.M.; Liu, J.R.; Wei, A.L.; Ye, Q.; Zeng, B.; Li, M.F.; Guo, Y.P.; et al. Puerariae Lobatae Radix Flavonoids and Puerarin Alleviate Alcoholic Liver Injury in Zebrafish by Regulating Alcohol and Lipid Metabolism. Biomed. Pharmacother. 2021, 134, 111121. [Google Scholar] [CrossRef]
- Jin, Y.; Wu, S.; Zeng, Z.; Fu, Z. Effects of Environmental Pollutants on Gut Microbiota. Environ. Pollut. 2017, 222, 1–9. [Google Scholar] [CrossRef]
- Lu, L.; Wan, Z.; Luo, T.; Fu, Z.; Jin, Y. Polystyrene Microplastics Induce Gut Microbiota Dysbiosis and Hepatic Lipid Metabolism Disorder in Mice. Sci. Total Environ. 2018, 631–632, 449–458. [Google Scholar] [CrossRef]
- Gong, J.; Li, L.; Zuo, X.; Li, Y. Change of the Duodenal Mucosa-Associated Microbiota Is Related to Intestinal Metaplasia. BMC Microbiol. 2019, 19, 275–281. [Google Scholar] [CrossRef]
- Sun, S.; Wang, K.; Sun, L.; Cheng, B.; Qiao, S.; Dai, H.; Shi, W.; Ma, J.; Liu, H. Therapeutic Manipulation of Gut Microbiota by Polysaccharides of Wolfiporia Cocos Reveals the Contribution of the Gut Fungi-Induced PGE2 to Alcoholic Hepatic Steatosis. Gut Microbes 2020, 12, 1830693. [Google Scholar] [CrossRef]
- Sheng, Y.; Ren, H.; Limbu, S.M.; Sun, Y.; Qiao, F.; Zhai, W.; Du, Z.Y.; Zhang, M. The Presence or Absence of Intestinal Microbiota Affects Lipid Deposition and Related Genes Expression in Zebrafish (Danio rerio). Front. Microbiol. 2018, 9, 1124. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Qin, Z.; Huang, Z.; Bao, Z.; Luo, T.; Jin, Y. Effects of Polyethylene Microplastics on the Microbiome and Metabolism in Larval Zebrafish. Environ. Pollut. 2021, 282, 117039. [Google Scholar] [CrossRef] [PubMed]
- Nagpal, R.; Shively, C.A.; Appt, S.A.; Register, T.C.; Michalson, K.T.; Vitolins, M.Z.; Yadav, H. Gut Microbiome Composition in Non-Human Primates Consuming a Western or Mediterranean Diet. Front. Nutr. 2018, 5, 28. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Forward Primer (5′ to 3′) | Reverse Promer (5′ to 3′) |
|---|---|---|
| jun | acacaacatgacgctcaatc | gctagactggatgatgagcc |
| hsp90aa1.2 | gagagctcatctccaactcc | gctcttctttgttgggaatg |
| acox1 | ctgaggctctggtggacgtg | ttgaacagtccaacaatctc |
| cyp3a66 | gagaaagcttgccaaacagg | agaagcgtgtgaatcacagc |
| fabp2 | catgacaacctgaagatcac | ttgtccttgcgtgtgaaagt |
| rpp0 | ctgaacatctcgcccttctc | tagccgatctgcagacacac |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Q.; Wu, L.; Wang, G.; Zheng, F.; Sun, J.; Zhang, Y.; Li, Z.; Li, L.; Sun, B. Inhibitory Effects of Jiuzao Polysaccharides on Alcoholic Fatty Liver Formation in Zebrafish Larvae and Their Regulatory Impact on Intestinal Microbiota. Foods 2024, 13, 276. https://doi.org/10.3390/foods13020276
Li Q, Wu L, Wang G, Zheng F, Sun J, Zhang Y, Li Z, Li L, Sun B. Inhibitory Effects of Jiuzao Polysaccharides on Alcoholic Fatty Liver Formation in Zebrafish Larvae and Their Regulatory Impact on Intestinal Microbiota. Foods. 2024; 13(2):276. https://doi.org/10.3390/foods13020276
Chicago/Turabian StyleLi, Qing, Liling Wu, Guangnan Wang, Fuping Zheng, Jinyuan Sun, Yuhang Zhang, Zexia Li, Lianghao Li, and Baoguo Sun. 2024. "Inhibitory Effects of Jiuzao Polysaccharides on Alcoholic Fatty Liver Formation in Zebrafish Larvae and Their Regulatory Impact on Intestinal Microbiota" Foods 13, no. 2: 276. https://doi.org/10.3390/foods13020276