

Leaf Mustard (Brassica juncea) Germplasm Resources Showed Diverse Characteristics in Agro-Morphological Traits and Glucosinolate Levels
 ,
,  ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Reagents and Standard Chemicals
2.3. Agro-Morphological Characters
2.4. Sample Pretreatment and UPLC-MS/MS Analysis of Intact GSLs
2.5. Statistical Analysis
3. Results
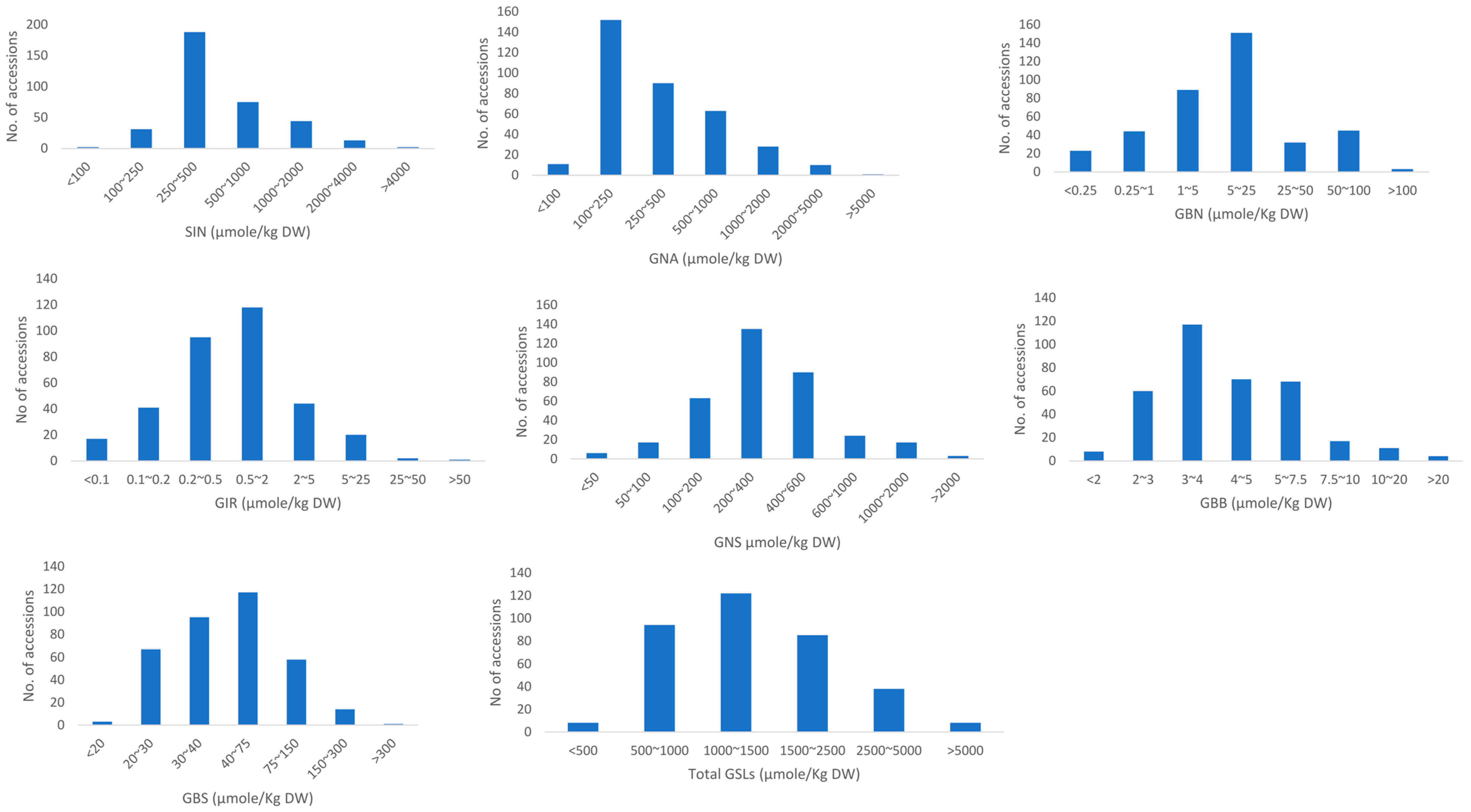
3.1. Variations in Agro-Morphological Traits among 355 Brassica juncea Accessions
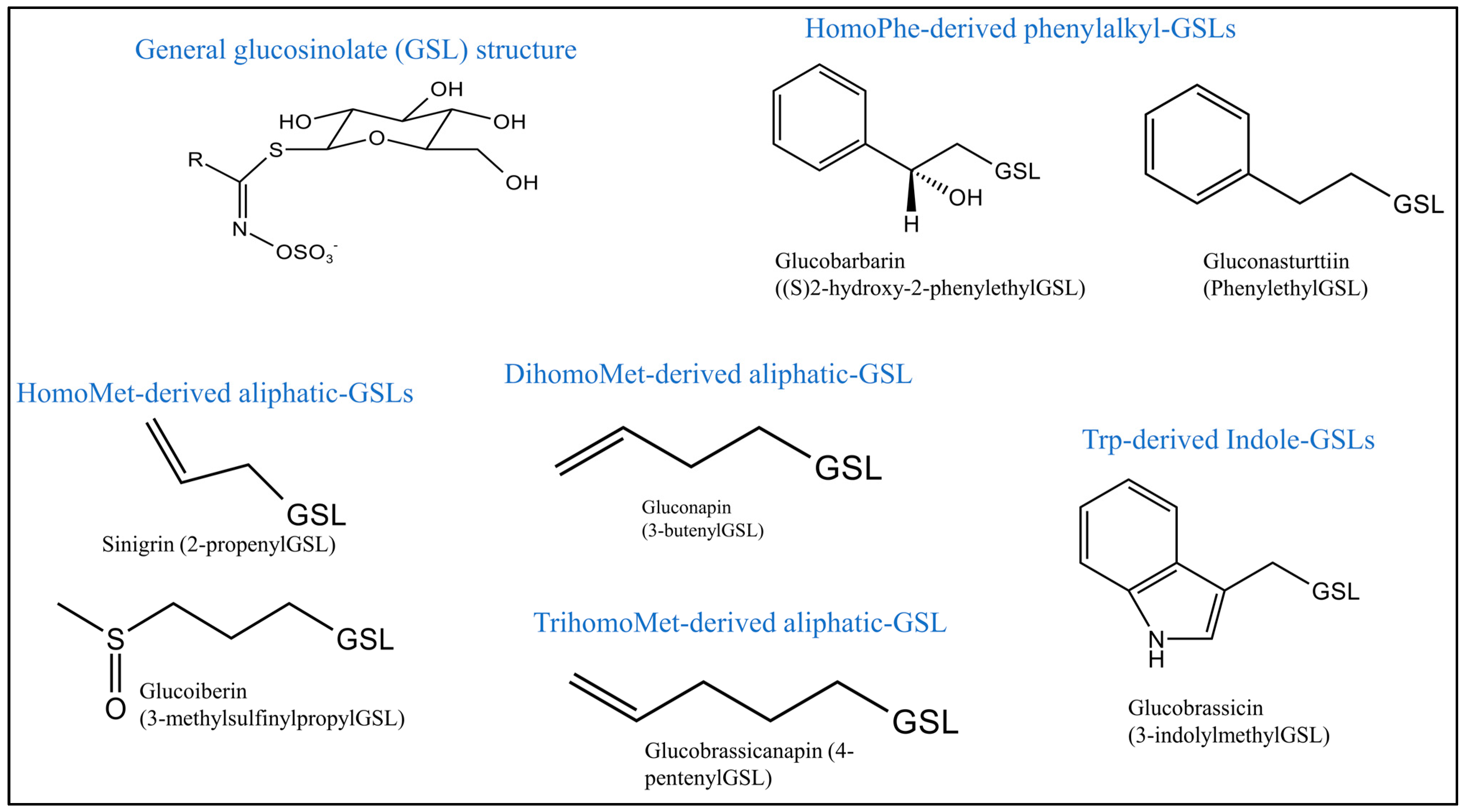
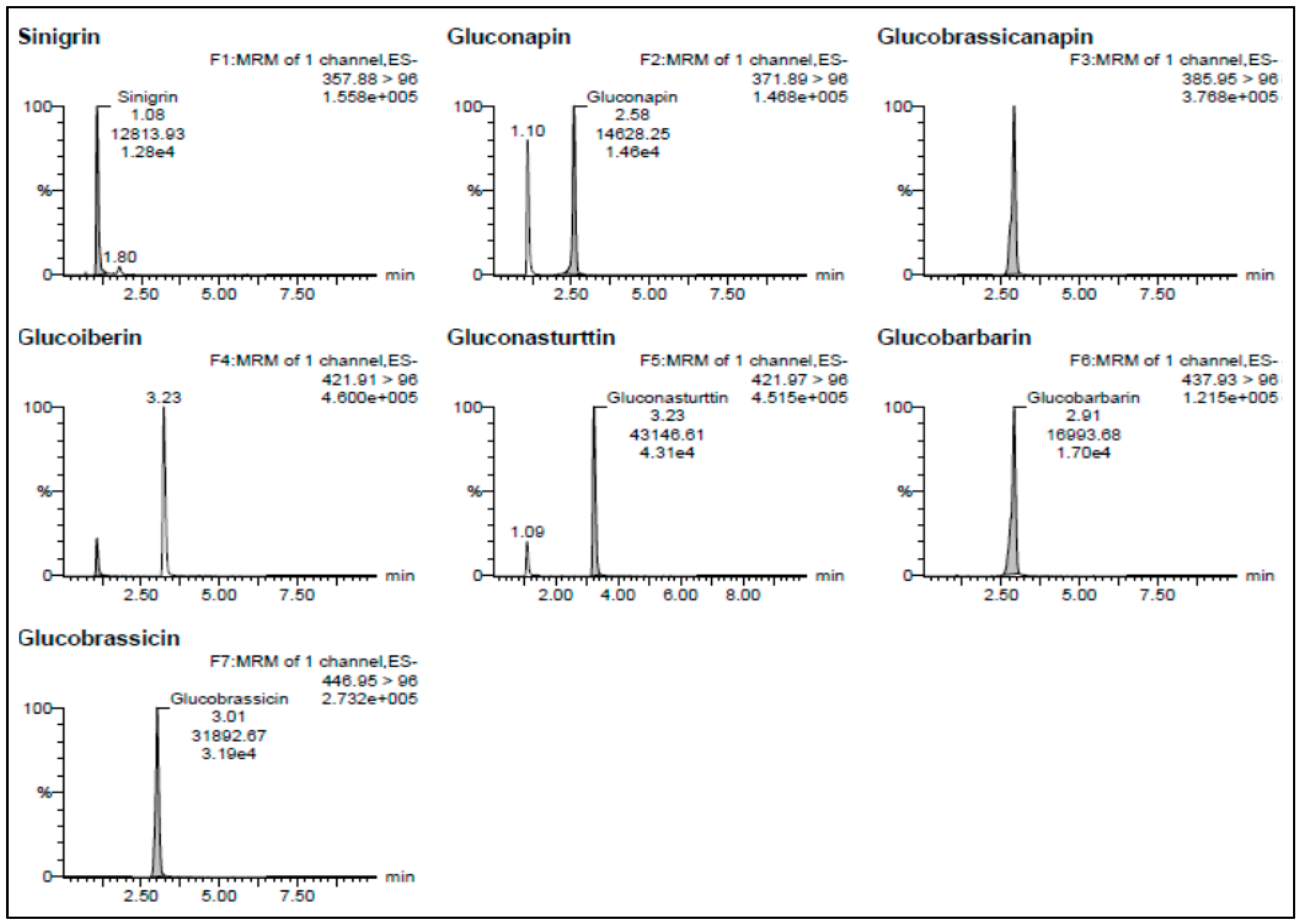
3.2. Identification and Quantification of Individual GSLs in Brassica juncea
3.3. Correlation Analysis among GSLs and Agro-Morphological Characters
3.4. Relationship between the Morphological Characters and the Levels of GSLs
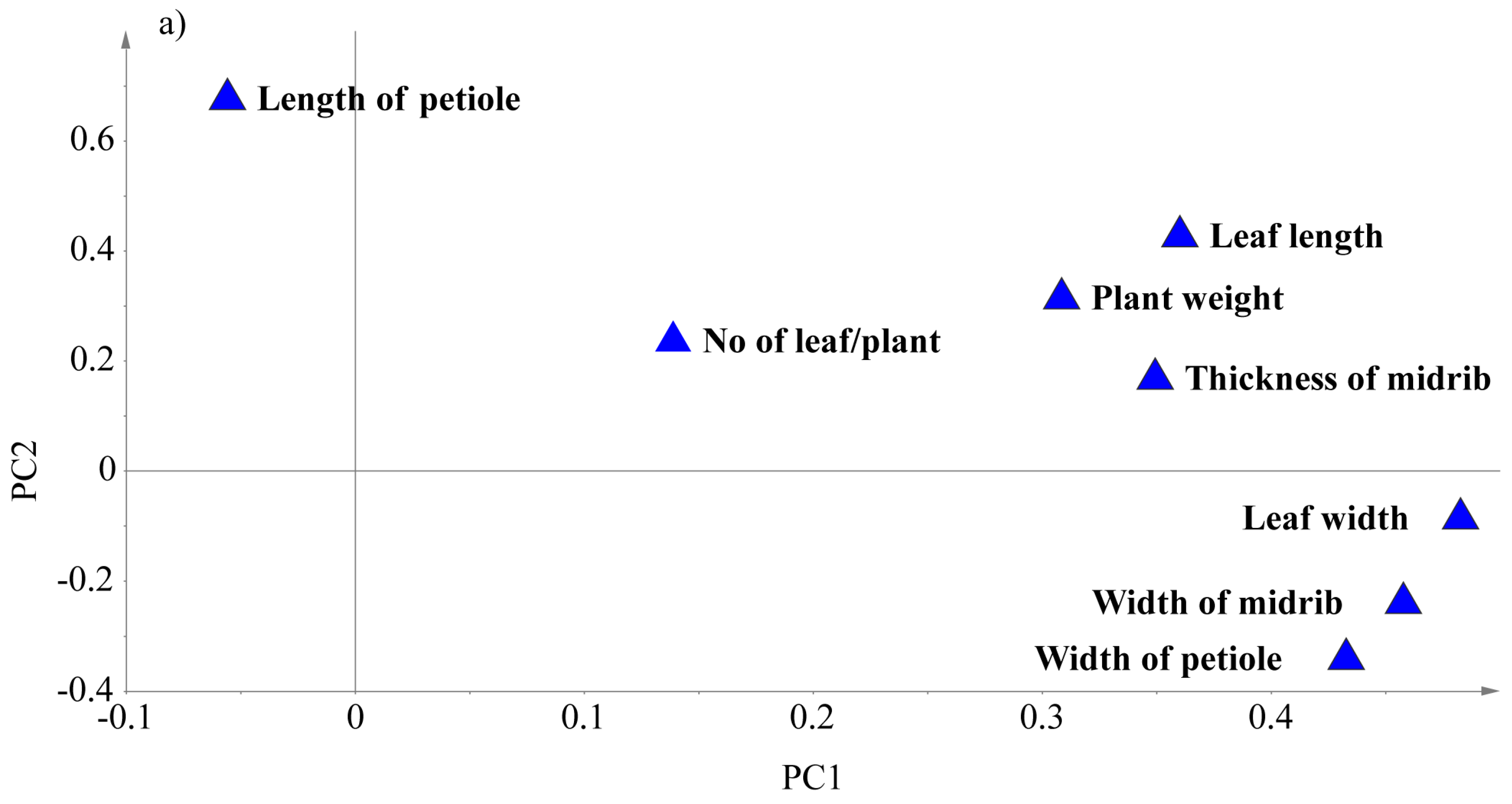
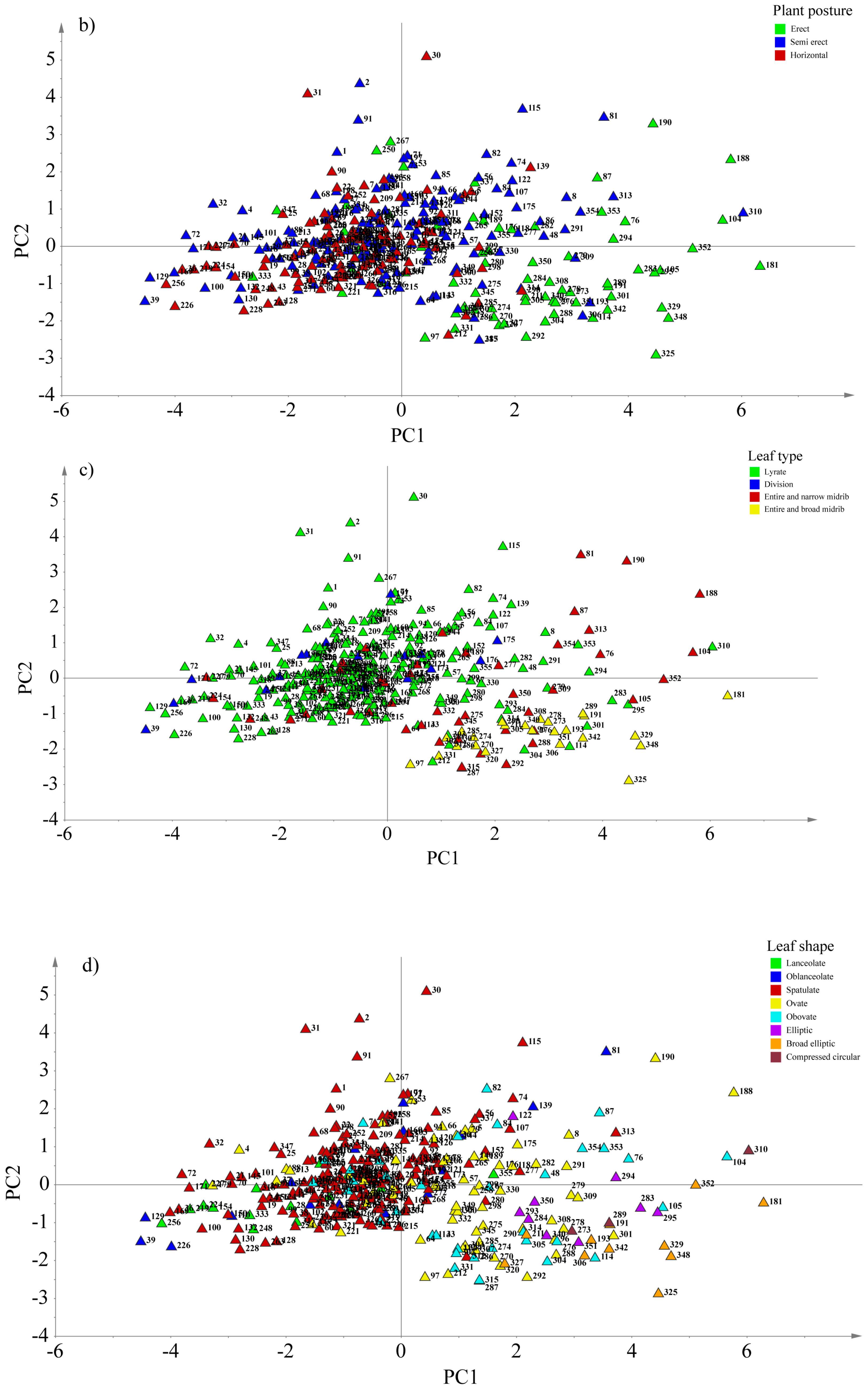
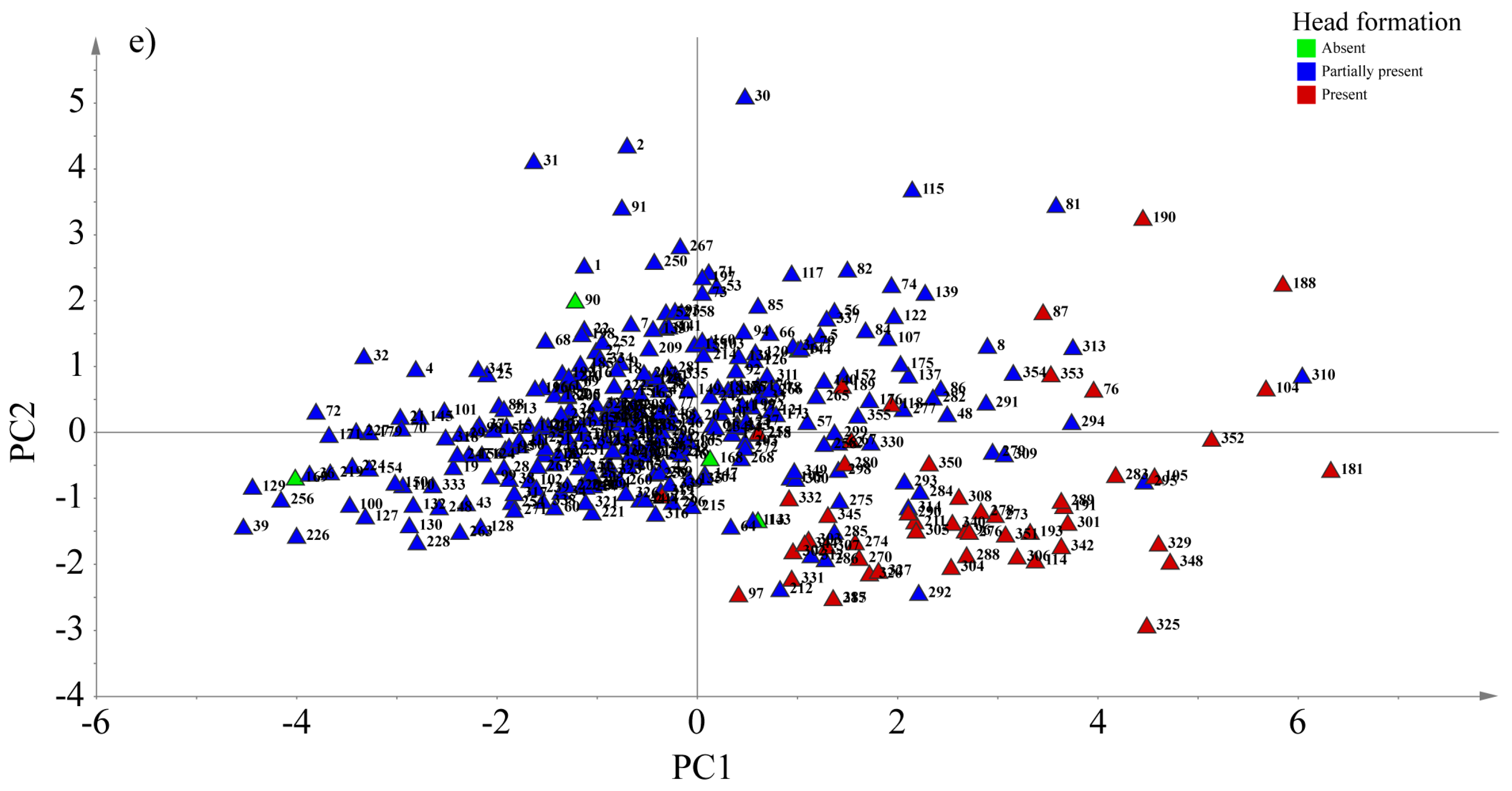
3.5. Principal Component Analysis (PCA) Based on Some Agro-Morphological Traits
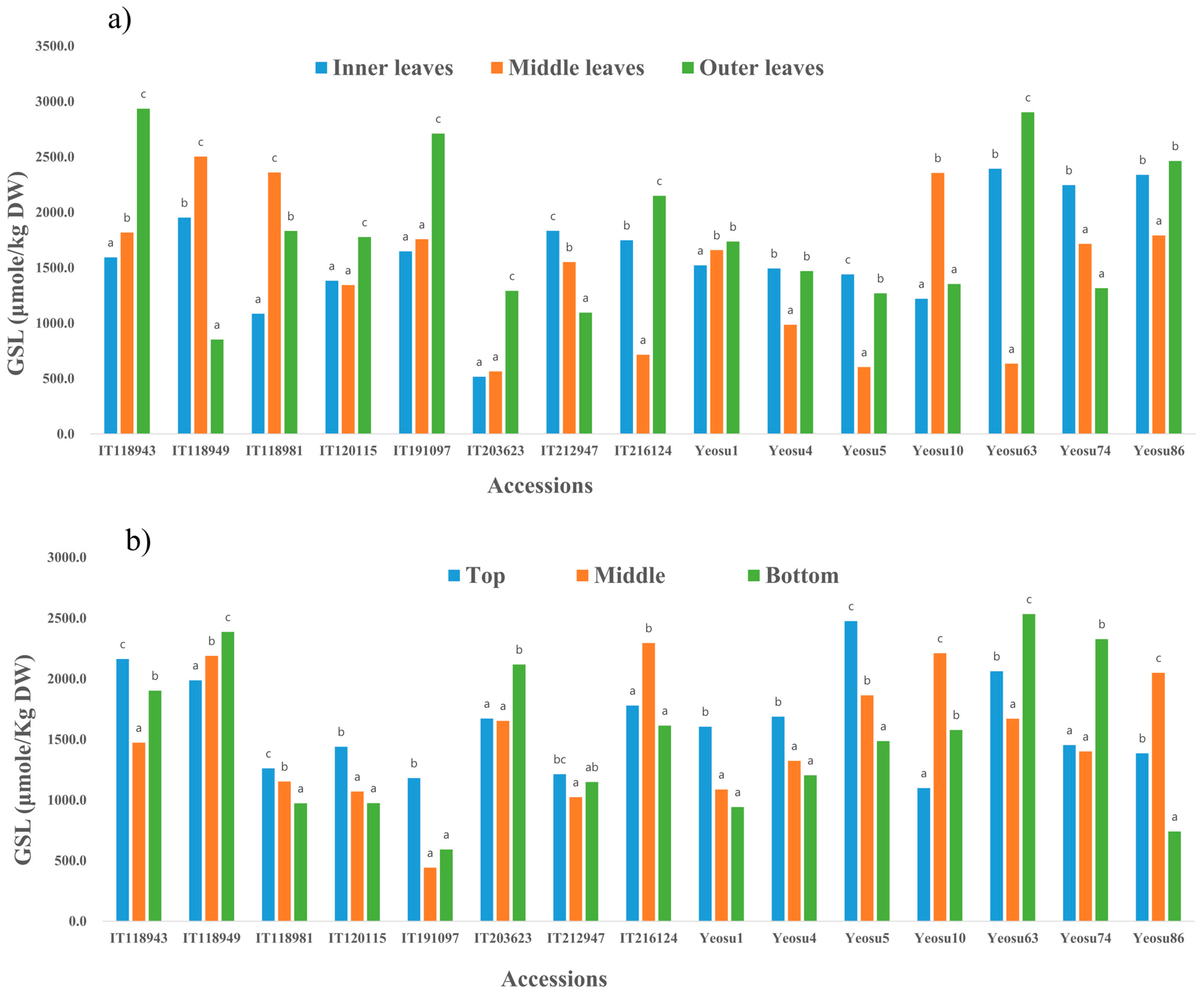
3.6. Intra- and Inter-Leaf Distribution of GSLs in Brassica juncea
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Thomas, J.; Kuruvilla, K.M.; Hrideek, T.K. Mustard. In Handbook of Herbs and Spices, 2nd ed.; Peter, K.V., Ed.; Woodhead Publishing Limited: Sawston, UK, 2012; Volume 1, pp. 388–398. [Google Scholar]
- Lietzow, J. Biologically active compounds in mustard seeds: A toxicological perspective. Foods 2021, 10, 2089. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Lee, I.H.; Kim, H.T.; Yang, S.Y.; Nou, I.S. Analysis of genetic similarity of dolsan leaf mustard based on several horticultural traits and RAPD. Korean J. Hortic. Sci. Technol. 2007, 25, 305–310. [Google Scholar]
- Kang, S.K. Antimicrobial activities in the Korean traditional leaf mustard, Brassica juncea Coss. Korean J. Plant Resour. 2005, 8, 96–102. [Google Scholar]
- Vig, A.P.; Rampal, G.; Thind, T.S.; Arora, S. 2009. Bio-protective effects of glucosinolates—A review. LWT-Food Sci. Technol. 2009, 42, 1561–1572. [Google Scholar] [CrossRef]
- Blažević, I.; Mastelić, J. Glucosinolate degradation products and other bound and free volatiles in the leaves and roots of radish (Raphanus sativus L.). Food Chem. 2009, 113, 96–102. [Google Scholar] [CrossRef]
- Kushad, M.M.; Brown, A.F.; Kurilich, A.C.; Juvik, J.A.; Klein, B.P.; Wallig, M.A.; Jeffery, E.H. Variation of glucosinolates in vegetable crops of Brassica oleracea. J. Agric. Food Chem. 1999, 47, 1541–1548. [Google Scholar] [CrossRef]
- Bennett, R.N.; Mellon, F.A.; Foidl, N.; Pratt, J.H.; Dupont, M.S.; Perkins, L.; Kroon, P.A. Profiling glucosinolates and phenolics in vegetative and reproductive tissues of the multi-purpose trees Moringa oleifera L. (Horseradish tree) and Moringa stenopetala L. J. Agric. Food Chem. 2003, 51, 3546–3553. [Google Scholar] [CrossRef]
- Bennet, R.N.; Mellon, F.A.; Kroon, P.A. Screening Crucifer seeds as sources of specific intact glucosinolates using ion-pair high-performance liquid chromatography negative ion electrospray mass spectrometry. J. Agric. Food Chem. 2004, 52, 428–438. [Google Scholar] [CrossRef]
- Clarke, D.B. Glucosinolates, structures and analysis in food. Anal. Methods 2010, 2, 310–325. [Google Scholar] [CrossRef]
- Zasada, I.A.; Ferris, H. Nematode suppression with brassicaceous amendments: Application based upon glucosinolate profiles. Soil Biol. Biochem. 2004, 36, 1017–1024. [Google Scholar] [CrossRef]
- Jones, R.B.; Frisina, C.L.; Winkler, S.; Imsic, M.; Tomkins, R.B. Cooking method significantly effects glucosinolate content and sulforaphane production in broccoli florets. Food Chem. 2010, 123, 237–242. [Google Scholar] [CrossRef]
- Pérez-Balibrea, S.; Moreno, D.A.; García-Viguera, C. Glucosinolates in broccoli sprouts (Brassica oleracea var. italica) as conditioned by sulphate supply during germination. J. Food Sci. 2010, 75, 673–677. [Google Scholar]
- Charron, C.S.; Saxton, A.M.; Sams, C.E. Relationship of climate and genotype to seasonal variation in the glucosinolate-myrosinase system. I. Glucosinolate content in ten cultivars of Brassica oleracea grown in fall and spring seasons. J. Sci. Food Agric. 2005, 85, 671–681. [Google Scholar] [CrossRef]
- Carlson, D.G.; Daxenbichler, M.E.; Vanetten, C.H.; Kwolek, W.F.; Williams, P.H. Glucosinolates in Crucifer vegetables: Broccoli, Brussels sprouts, cauliflower, collards, kale, mustard greens, and kohlrabi. Am. Soc. Hortic. Sci. 1987, 112, 173–178. [Google Scholar] [CrossRef]
- Qian, H.; Liu, T.; Deng, M.; Miao, H.; Cai, C.; Shen, W.; Wang, Q. Effects of light quality on main health-promoting compounds and antioxidant capacity of Chinese kale sprouts. Food Chem. 2016, 196, 1232–1238. [Google Scholar] [CrossRef]
- Tiedink, H.G.M.; Davies, J.A.R.; van Broekhoven, L.W.; Van der Kamp, H.J.; Jongen, W.M.F. Formation of mutagenic N-nitroso compounds in vegetable extracts upon nitrite treatment: A comparison with the glucosinolate content. Food Chem. Toxicol. 1988, 26, 947–954. [Google Scholar] [CrossRef]
- Verkerk, R.; Dekker, M. Glucosinolates and myrosinase activity in red cabbage (Brassica oleracea L. var. Capatita f. rubra DC.) after various microwave treatments. J. Agric. Food Chem. 2004, 52, 7318–7323. [Google Scholar]
- Wiesner, M.; Zrenner, R.; Krumbein, A.; Glatt, H.; Schreiner, M. Genotypic variation of the glucosinolate profile in pak choi (Brassica rapa ssp. chinensis). J. Agric. Food Chem. 2013, 61, 1943–1953. [Google Scholar] [CrossRef]
- Rhee, J.H.; Choi, S.; Lee, J.E.; Hur, O.S.; Ro, N.Y.; Hwang, A.J.; Ko, H.; Chung, Y.; Noh, J.; Assefa, A.D. Glucosinolate content in Brassica genetic resources and their distribution pattern within and between inner, middle, and outer leaves. Plants 2020, 9, 1421. [Google Scholar] [CrossRef]
- Doerr, B.; Wade, K.L.; Stephenson, K.K.; Reed, S.B.; Fahey, J.W. Cultivar effect on Moringa oleifera glucosinolate content and taste: A pilot study. Ecol. Food Nutr. 2009, 48, 199–211. [Google Scholar] [CrossRef]
- Förster, N.; Ulrichs, C.; Schreiner, M.; Müller, C.T.; Mewis, I. Development of a reliable extraction and quantification method for glucosinolates in Moringa oleifera. Food Chem. 2015, 166, 456–464. [Google Scholar] [CrossRef] [PubMed]
- Han, N.; Súudi, M.; Kim, J. The major aliphatic glucosinolate content in Korean radish during vegetative and reproductive growth. Hortic. Environ. Biotechnol. 2015, 56, 152–158. [Google Scholar] [CrossRef]
- Giallourou, N.; Oruna-Concha, M.J.; Harbourne, N. Effects of domestic processing methods on the phytochemical content of watercress (Nasturtium officinale). Food Chem. 2016, 212, 411–419. [Google Scholar] [CrossRef] [PubMed]
- Pradhan, A.K.; Pental, D. Genetics of Brassica juncea. In Genetics and Genomics of the Brassicaceae; Plant Genetics and Genomics: Crops and Models; Schmidt, R., Bancroft, I., Eds.; Springer: New York, NY, USA, 2011; Volume 9, pp. 323–345. [Google Scholar]
- Devi, T.R.; Devshini Devi, N.; Vivekananda, Y.; Sharma, P.R. Genetic diversity analysis in Indian mustard (Brassica juncea L. Czern and Coss) genotypes using agro-morphological parameters. Electron. J. Plant Breed. 2017, 8, 749–753. [Google Scholar] [CrossRef]
- Khan, M.A.; Rabbani, M.A.; Munir, M.; Ajmal, S.K.; Malik, M.A. Assessment of genetic variation within Indian mustard (Brassica juncea) germplasm using random amplified polymorphic DNA markers. J. Integr. Plant Biol. 2008, 50, 385–392. [Google Scholar] [CrossRef] [PubMed]
- Khan, W.M.; Munir, I.; Arif, M.; Iqbal, A.; Ali, I.; Ahmad, D.; Ahmad, M.; Mian, A.; Bakht, J.; Swati, Z.A. Assessment of genetic diversity of Brassica juncea germplasm using randomly amplified polymorphic DNA (RAPD) markers. Afr. J. Biotechnol. 2011, 10, 3654–3658. [Google Scholar]
- Saleem, N.; Ahmad Jan, S.; Jawaad Atif, M.; Khurshid, H.; Ali Khan, S.; Abdullah, M.; Jahanzaib, M.; Ahmed, H.; Ullah, S.F.; Iqbal, A.; et al. Multivariate based variability within diverse Indian mustard (Brassica juncea L.) genotypes. Open J. Genet. 2017, 07, 69–83. [Google Scholar] [CrossRef]
- Sharma, H.K.; Kumar, A.; Singh, V.V.; Meena, H.S.; Priyamedha; Sharma, P.; Rai, P.K. Variability and genetic diversity study based on agro- morphological traits in a diverse set of Indian mustard [Brassica juncea (L.) Czern. & Coss.] germplasm. J. Environ. Biol. 2021, 42, 22438. [Google Scholar]
- Shekhawat, N.; Jadeja, G.C.; Singh, J. Genetic variability for yield and its components in Indian mustard (Brassica juncea L. Czern & Coss). Electron. J. Plant Breed. 2014, 5, 117–119. [Google Scholar]
- Tiwari, V.K. Morphological parameters in breeding for higher seed yield in Indian mustard [Brassica juncea (L.) Czern. & Coss.]. Electron. J. Plant Breed. 2019, 10, 187–195. [Google Scholar]
- Verma, K.; Tripathi, M.K.; Tiwari, S.; Tripathi, N. Analysis of genetic diversity among Brassica juncea genotypes using morpho-physiological and SSR Markers. Int. J. Curr. Microbiol. Appl. Sci. 2021, 10, 1108–1117. [Google Scholar] [CrossRef]
- Merah, O. Genetic variability in glucosinolates in seed of Brassica juncea: Interest in mustard condiment. J. Chem. 2015, 2015, 606142. [Google Scholar] [CrossRef]
- Saikia, S.L.; Rai, G.K.; Salgotra, R.; Rai, S.; Singh, M.; Rai, P. Erucic acid and glucosinolate variability in Brassica juncea L. Int. J. Chem. Stud. 2018, 6, 1223–1226. [Google Scholar]
- Velasco, L.; Becker, H.C. Variability for seed glucosinolates in a germplasm collection of the genus Brassica. Genet. Resour. Crop Evol. 2000, 47, 231–238. [Google Scholar] [CrossRef]
- Zhang, C.; Di, H.; Lin, P.; Wang, Y.; Li, Z.; Lai, Y.; Li, H.; Sun, B.; Zhang, F. Genotypic variation of glucosinolates and their breakdown products in mustard (Brassica juncea) seeds. Sci. Hortic. 2022, 294, 110765. [Google Scholar] [CrossRef]
- Kim, H.W.; Ko, H.C.; Baek, H.J.; Cho, S.M.; Jang, H.H.; Lee, Y.M.; Kim, J.B. Identification and quantification of glucosinolates in Korean leaf mustard germplasm (Brassica juncea var. integrifolia) by liquid chromatography–electrospray ionization/tandem mass spectrometry. Eur. Food Res. Technol. 2016, 242, 1479–1484. [Google Scholar]
- Sodhi, Y.S.; Mukhopadhyay, A.; Arumugam, N.; Verma, J.K.; Gupta, V.; Pental, D.; Pradhan, A.K. Genetic analysis of total glucosinolate in crosses involving a high glucosinolate Indian variety and a low glucosinolate line of Brassica juncea. Plant Breed. 2002, 121, 508–511. [Google Scholar] [CrossRef]
- International Union for the Protection of New Varieties of Plants (UPOV). Guidelines for the Conduct of Tests for Distinctness, Uniformity and Stability of Brassica juncea L. Czern. 2017. Available online: http://www.upov.int (accessed on 30 March 2022).
- Yadav, R.; Singh, R.; Kumar, S.; Prasad, T.V.; Bharadwaj, R.; Kaur, V.; Petapadi, A.; Kumar, A. Genetic diversity among indigenous germplasm of Brassica juncea (L.) Czern and Coss, using agro-morphological and phenological traits. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2017, 87, 1125–1131. [Google Scholar] [CrossRef]
- Teklehaymanot, T.; Wang, H.; Liang, J.; Wu, J.; Lin, R.; Zhou, Z.; Cai, X.; Wang, X. Variation in plant morphology and sinigrin content in Ethiopian mustard (Brassica carinata L.). Hortic. Plant J. 2019, 5, 205–212. [Google Scholar] [CrossRef]
- Possenti, M.; Baima, S.; Raffo, A.; Durazzo, A.; Giusti, A.M.; Natella, F. Glucosinolates in food. In Glucosinolates, Reference Series in Phytochemistry; Mérillon, J.M., Ramawat, K.G., Eds.; Springer: Cham, Switzerland, 2017; pp. 87–132. [Google Scholar]
- Choi, S.H.; Park, S.; Lim, Y.P.; Kim, S.J.; Park, J.T.; An, G. Metabolite profiles of glucosinolates in cabbage varieties (Brassica oleracea var. capitata) by season, color, and tissue position. Hortic. Environ. Biotechnol. 2014, 55, 237–247. [Google Scholar] [CrossRef]
- Shelton, A.L. Within-plant variation in glucosinolate concentrations of Raphanus sativus across multiple scales. J. Chem. Ecol. 2005, 31, 1711–1732. [Google Scholar] [CrossRef] [PubMed]
- Sharma, D.; Lal, J.N.; Singh, S.S.P.; Parmar, N. Genetic diversity in leafy mustard (Brassica juncea var. rugosa) as revealed by agro-morphological traits and SSR markers. Physiol. Mol. Biol. Plants 2020, 26, 2005–2018. [Google Scholar] [PubMed]
- Singh, K.H.; Shakya, R.; Nanjundan, J.; Thakur, A.; Singh, K.; Singh, K.K. Genetic diversity assessment and characterization of Indian mustard (Brassica juncea L.) varieties using agro-morphological traits. Indian J. Plant Genet. Resour. 2018, 31, 44–50. [Google Scholar] [CrossRef]
- Singh, K.H.; Shakya, R.; Mahawar, R.K. Genetic diversity and patterns of variation among Indian mustard (Brassica juncea (L.) Czernj. & Cosson) varieties. SABRAO J. Breed. Genet. 2014, 46, 329–339. [Google Scholar]
- Rabbani, M.A.; Iwabuchi, A.; Murakami, Y.; Suzuki, T.; Takayanagi, K. Phenotypic variation and the relationships among mustard (Brassica juncea L.) germplasm from Pakistan. Euphytica 1998, 101, 357–366. [Google Scholar] [CrossRef]
- Cuong, D.M.; Kim, J.K.; Bong, S.J.; Baek, S.A.; Jeon, J.; Park, J.S.; Park, S.U. Comparative analysis of glucosinolates and metabolite profiling of green and red mustard (Brassica juncea) hairy roots. 3 Biotech 2018, 8, 382. [Google Scholar] [CrossRef]
- Abe, K.; Kido, S.; Maeda, T.; Kami, D.; Matsuura, H.; Shimura, H.; Suzuki, T. Glucosinolate profiles in Cardamine fauriei and effect of light quality on glucosinolate concentration. Sci. Hortic. 2015, 189, 12–16. [Google Scholar] [CrossRef]
- Warwick, S.I.; Gugel, R.K.; Mcdonald, T.; Falk, K.C. Genetic variation of Ethiopian mustard (Brassica carinata A. Braun) germplasm in western Canada. Genet. Resour. Crop Evol. 2006, 53, 297–312. [Google Scholar] [CrossRef]
- Bell, L.; Wagstaff, C. Enhancement of glucosinolate and isothiocyanate profiles in Brassicaceae crops: Addressing challenges in breeding for cultivation, storage, and consumer-related traits. J. Agric. Food Chem. 2017, 65, 9379–9403. [Google Scholar] [CrossRef]
- Clarke, J.D.; Dashwood, R.H.; Ho, E. Multi-targeted prevention of cancer by sulforaphane. Cancer Lett. 2008, 269, 291–304. [Google Scholar] [CrossRef]
- Zhang, Y. Cancer-preventive isothiocyanates: Measurement of human exposure and mechanism of action. Mutat. Res. 2004, 555, 173–190. [Google Scholar] [CrossRef] [PubMed]
- Blažević, I.; Montaut, S.; Burčul, F.; Rollin, P. Glucosinolates: Novel sources and biological potential. In Glucosinolates, Reference Series in Phytochemistry; Mérillon, J.M., Ramawat, K.G., Eds.; Springer: Cham, Switzerland, 2017; pp. 3–60. [Google Scholar]
- Singh, A. Glucosinolates and plant defense. In Glucosinolates, Reference Series in Phytochemistry; Mérillon, J.M., Ramawat, K.G., Eds.; Springer: Cham, Switzerland, 2017; pp. 237–246. [Google Scholar]
- Liu, Z.; Wang, H.; Xie, J.; Lv, J.; Zhang, G.; Hu, L.; Luo, S.; Li, L.; Yu, J. The roles of Cruciferae glucosinolates in disease and pest resistance. Plants 2021, 10, 1097. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Glucosinolates | tR | MRM Transition | CID | Calibration Equation |
|---|---|---|---|---|
| Sinigrin | 1.08 | 357.88 > 96 | 20 | Y = 12.8988X + 16.8399, r2 = 0.999 |
| Gluconapin | 2.58 | 371.89 > 96 | 20 | Y = 12.0411X − 13.1698, r2 = 0.998 |
| Glucobrassicanapin | 2.90 | 385.95 > 96 | 25 | Y = 44.9459X − 37.6357, r2 = 0.997 |
| Glucoiberin | 1.09 | 421.91 > 96 | 20 | Y = 8.48762X − 24.3857, r2 = 0.999 |
| Gluconasturtiin | 3.23 | 421.97 > 96 | 25 | Y = 39.0356X − 44.9008, r2 = 0.998 |
| Glucobarbarin | 2.91 | 437.93 > 96 | 20 | Y = 15.443X − 21.6364, r2 = 0.996 |
| Glucobrassicin | 3.01 | 446.95 > 96 | 20 | Y = 28.0663X − 43.3473, r2 = 0.999 |
| S/NO | Trait | Descriptions * |
|---|---|---|
| 1 | Plant habit (posture) | Erect (73), semi-erect (162), horizontal (114), mixed (6) |
| 2 | Plant: head formation | Absent (5), partially present (297), fully present (52), mixed (1) |
| 3 | Leaf attitude at the apical part | Flat (119), bend downward (215), bend upward (15), mixed (5) |
| 4 | Leaf type | Lyrate (257), division (17), entire and narrow midrib (46), entire and broad midrib (24), mixed (11) |
| 5 | Leaf shape | Lanceolate (17), oblanceolate (16), spatulate (186), ovate (67), obovate (35), elliptic (9), broad elliptic (11), compressed circular (3), mixed (11) |
| 6 | Leaf waxiness | Weak (52), medium (142), strong (148), mixed (13) |
| 7 | Lobation of leaf margins | Absent (10), lobed (103), cleft (181), parted (21), dissected (22), mixed (18) |
| 7 | Leaf blade: density of incisions of margin | Absent (8), sparse (113), medium (164), dense (29), very dense (23), mixed (18) |
| 8 | Leaf blade: size of the terminal lobe | Absent (16), small (196), medium (106), large (20), mixed (17) |
| 9 | Leaf blade: number of serrates | Absent (17), few (152), medium (127), many (47), mixed (12) |
| 11 | Leaf blade: anthocyanin coloration | Absent (27), very weak (121), weak (52), medium (65), strong (42), very strong (24), mixed (24) |
| 12 | Leaf blade: blistering | Absent/weak (80), medium (118), strong (143), mixed (14) |
| 13 | Leaf blade: pubescence on the upper side | Absent (81), present (263), mixed (11) |
| 14 | Leaf blade: pubescence on the lower side | Absent (54), few (101), medium (154), many (35), mixed (11) |
| 15 | Stem hair | Absent (59), present (291), mixed (5) |
| 16 | Midrib transection | Horizontal (41), intermediate (122), semi-circled (191) |
| 17 | Petiole anthocyanin coloration | Absent (77), present (257), mixed (18) |
| GSLs and Agro-Morphological Characters | Range | Average ± SD | Coefficient of Variation (%) |
|---|---|---|---|
| SIN | 13.0–4184.6 | 635.9 ± 586.4 | 92.2 |
| GNA | 44.1–7708.7 | 496.4 ± 628.5 | 126.6 |
| GBN | 0.1–180.2 | 12.8 ± 20.1 | 156.9 |
| GIB | 0.0–93.6 | 2.1 ± 6.3 | 299.5 |
| GBB | 0.7–33.9 | 4.7 ± 3.3 | 68.9 |
| Total aliphatic GSLs | 133.7–7738.2 | 1151.8 ± 982.6 | 85.3 |
| GNS | 27.2–3393.5 | 401.9 ± 342.9 | 85.3 |
| GBS | 17.3–351.9 | 57.9 ± 44.2 | 76.3 |
| Total GSLs | 320.4–8055.3 | 1610.8 ± 1064.8 | 66.1 |
| Leaf length (cm) | 20.8–57.0 | 36.3 ± 5.8 | 16.0 |
| Leaf width (cm) | 8.6–31.8 | 18.1 ± 4.3 | 24.0 |
| Midrib width (cm) | 0.6–3.8 | 1.8 ± 0.6 | 33.7 |
| Midrib thickness (cm) | 0.3–1.1 | 0.6 ± 0.1 | 18.9 |
| Petiole length (cm) | 0.9–12.5 | 3.1 ± 1.5 | 47.7 |
| Petiole width (cm) | 0.4–3.8 | 1.3 ± 0.6 | 48.7 |
| Number of leaves (ea) | 6.0–28.0 | 16.3 ± 6 | 36.8 |
| Plant weight (g, FW) | 91.7–1854.3 | 338.9 ± 152.7 | 45.1 |
| Leaf Length | Leaf Width | No of Leaves | Midrib Width | Midrib Thickness | Petiole Length | Petiole Width | Plant Weight | SIN | GNA | GBN | GIB | GNS | GBB | GBS | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Leaf width | 0.622 ** | ||||||||||||||
| Number of leaves | 0.303 ** | 0.167 ** | |||||||||||||
| Midrib width | 0.444 ** | 0.854 ** | 0.256 ** | ||||||||||||
| Midrib thickness | 0.475 ** | 0.581 ** | 0.161 ** | 0.455 ** | |||||||||||
| Petiole length | 0.210 ** | −0.128 * | −0.004 | −0.229 ** | 0.045 | ||||||||||
| Petiole width | 0.320 ** | 0.829 ** | 0.004 | 0.895 ** | 0.429 ** | −0.268 ** | |||||||||
| Plant weight | 0.532 ** | 0.451 ** | 0.085 | 0.348 ** | 0.352 ** | 0.122 * | 0.368 ** | ||||||||
| SIN | −0.116 * | 0.008 | −0.009 | 0.051 | −0.082 | −0.185 ** | 0.055 | −0.138 ** | |||||||
| GNA | −0.061 | 0.081 | 0.012 | 0.143 ** | 0.119 * | −0.007 | 0.119 * | −0.098 | 0.272 ** | ||||||
| GBN | −0.049 | 0.065 | 0.063 | 0.155 ** | 0.041 | −0.166 ** | 0.121 * | −0.106 * | 0.346 ** | 0.551 ** | |||||
| GIB | −0.119 * | −0.005 | 0.038 | 0.04 | −0.061 | −0.144 ** | 0.061 | −0.044 | 0.256 ** | 0.159 ** | 0.139 * | ||||
| GNS | −0.016 | −0.102 | 0.092 | −0.053 | −0.087 | −0.000 | −0.101 | −0.042 | 0.082 | −0.085 | 0.097 | −0.058 | |||
| GBB | −0.005 | −0.009 | 0.012 | 0.048 | −0.01 | −0.036 | 0.016 | −0.024 | 0.139 ** | −0.054 | 0.098 | 0.004 | 0.565 ** | ||
| GBS | −0.101 | 0.065 | −0.015 | 0.112 * | 0.006 | −0.124 * | 0.127 * | −0.105 * | 0.463 ** | 0.380 ** | 0.173 ** | 0.064 | 0.110 * | 0.083 | |
| Total GSLs | −0.111 * | 0.024 | 0.032 | 0.103 | −0.001 | −0.115* | 0.076 | −0.154 ** | 0.766 ** | 0.740 ** | 0.575 ** | 0.226 ** | 0.325 ** | 0.235 ** | 0.560 ** |
| Morphological Characters | Descriptions (N) | SIN | GNA | GBN | GIB | GNS | GBB | GBS | Total GSLs |
|---|---|---|---|---|---|---|---|---|---|
| Plant habit (posture) | Erect (73) | 659.2 ab | 614.9 a | 12.9 a | 2.5 a | 370.4 a | 5.1 a | 71.1 b | 1735.9 ab |
| Semi erect (162) | 541.2 a | 412.3 a | 11.0 a | 1.5 a | 399.5 a | 4.5 a | 48.3 a | 1418.2 a | |
| Horizontal (141) | 749.5 b | 537.7 a | 15.3 a | 2.7 a | 424.3 a | 4.8 a | 62.8 b | 1794.6 b | |
| Leaf: attitude at the apical part | Flat (119) | 680.1 a | 535.6 a | 12.2 a | 2.6 a | 412.7 a | 4.5 a | 58.8 a | 1703.9 a |
| Bend downward (215) | 607 a | 475.1 a | 12.9 a | 1.8 a | 397.8 a | 4.9 a | 58.5 a | 1557.9 a | |
| Bend upward (16) | 674.9 a | 486.5 a | 16.8 a | 2.9 a | 406.6 a | 4.4 a | 43.9 a | 1635.3 a | |
| Leaf type | Lyrate (257) | 635.8 ab | 463.2 a | 12.3 a | 1.9 a | 427.2 a | 4.8 a | 55.7 ab | 1599.7 ab |
| Division (17) | 305.1 a | 431.9 a | 7.2 a | 0.3 a | 355.6 a | 4.6 a | 33.1 a | 1137.7 a | |
| Entire, narrow midrib (46) | 678.5 ab | 631.0 a | 19.0 a | 3.2 a | 321.6 a | 4.2 a | 65.8 bc | 1723.1 ab | |
| Entire, broad midrib (24) | 762.3 b | 668.8 a | 13.0 a | 3.9 a | 341.7 a | 5.0 a | 86.2 c | 1880.9 b | |
| Leaf shape | Lanceolate (17) | 788.8 a | 400.8 a | 8.6 a | 1.7 a | 327.7 a | 4.9 a | 63.1 ab | 1595.5 a |
| Oblanceolate (16) | 524 a | 343.1 a | 7.7 a | 1.3 a | 398.7 a | 4.0 a | 55.4 ab | 1334.1 a | |
| Spatulate (186) | 633.6 a | 467.9 a | 11.5 a | 2.1 a | 441.0 a | 4.9 a | 56.3 ab | 1615.6 a | |
| Ovate (67) | 665.5 a | 688.7 a | 20.5 a | 2.1 a | 378.8 a | 4.6 a | 57.4 ab | 1817.3 a | |
| Obovate (35) | 510.0 a | 476.4 a | 13.9 a | 1.4 a | 286.0 a | 3.6 a | 64.5 ab | 1355.7 a | |
| Elliptic (9) | 590.9 a | 383.8 a | 4.2 a | 2.0 a | 409.5 a | 5.0 a | 43.9 a | 1439.4 a | |
| Broad elliptic (11) | 714.1 a | 261.0 a | 4.0 a | 5.3 a | 374.2 a | 5.7 a | 65.3 ab | 1429.4 a | |
| Compressed circular (3) | 694.8 a | 676.6 a | 13.0 a | 1.5 a | 395.5 a | 7.1 a | 109 b | 1897.5 a | |
| Lobation of leaf margins | Absent (10) | 600.1 a | 571.5 a | 21.1 b | 2.1 a | 298.0 a | 4.0 a | 60.1 ab | 1556.6 ab |
| Lobed (103) | 718.9 a | 565.0 a | 16.6 ab | 2.4 a | 397.0 a | 4.9 a | 64.2 ab | 1768.9 ab | |
| Cleft (181) | 592.5 a | 474.7 a | 11.0 ab | 2.0 a | 418.2 a | 4.8 a | 56.3 ab | 1557.6 ab | |
| Parted (21) | 840.5 a | 668.7 a | 16.2 ab | 3.1 a | 387.5 a | 4.1 a | 72.8 b | 1993.0 b | |
| Dissected (22) | 416.4 a | 199.5 a | 3.6 a | 0.6 a | 351.9 a | 3.6 a | 37.4 a | 1012.8 a | |
| Leaf blade: density of incisions of margins | Absent (8) | 614.3 a | 600.5 a | 24.4 b | 2.2 a | 335.1 a | 4.1 a | 67.2 a | 1647.8 a |
| Sparse (113) | 726.9 a | 543.3 a | 16.5 ab | 2.3 a | 403.4 a | 4.8 a | 65.1 a | 1762.1 a | |
| Medium (164) | 591.7 a | 474.4 a | 10.9 ab | 2.2 a | 415.0 a | 4.9 a | 55.8 a | 1552.8 a | |
| Dense (29) | 680.1 a | 557.5 a | 12.9 ab | 2.1 a | 377.0 a | 4.2 a | 64.9 a | 1698.7 a | |
| Very dense (23) | 447.3 a | 351.5 a | 4.3 a | 0.8 a | 355.8 a | 3.7 a | 36.2 a | 1199.6 a | |
| Leaf blade: size of terminal lobe | Absent (16) | 755.3 a | 560.7 a | 17.9 b | 1.9 a | 296.7 a | 4.4 a | 70.5 a | 1707.3 a |
| Small (196) | 623.0 a | 529.6 a | 14.2 ab | 2.3 a | 408.9 a | 4.6 a | 60.5 a | 1642.5 a | |
| Medium (106) | 660.9 a | 477.4 a | 11.0 ab | 2.1 a | 385.7 a | 4.9 a | 52.9 a | 1592.7 a | |
| Large (20) | 510.2 a | 253.0 a | 4.7 a | 0.7 a | 497.0 a | 5.0 a | 59.7 a | 1330.2 a | |
| Leaf blade: number of serrates | Absent (17) | 906.1 a | 514.2 a | 17.4 a | 4.8 a | 373.4 a | 5.5 a | 77.0 b | 1898.3 a |
| Few (152) | 596.2 a | 490.8 a | 13.8 a | 1.6 a | 405.2 a | 4.7 a | 55.6 ab | 1566.2 a | |
| Medium (127) | 670.2 a | 556.9 a | 12.6 a | 1.9 a | 394.3 a | 4.7 a | 62.4 ab | 1702.9 a | |
| Many (47) | 595.6 a | 383.8 a | 9.8 a | 3.4 a | 423.6 a | 4.4 a | 49.4 a | 1468.3 a | |
| Leaf blade: anthocyanin coloration | Absent (27) | 453.7 a | 296.3 a | 5.6 a | 1.5 a | 320.4 a | 3.9 a | 38.5 a | 1119.7 a |
| Very weak (121) | 649.0 a | 645.4 a | 16.7 a | 2.7 a | 362.0 a | 4.7 a | 64.9 a | 1744.8 a | |
| Weak (52) | 704.1 a | 410.5 a | 13.2 a | 1.6 a | 513.3 a | 5.3 a | 63.9 a | 1707.3 a | |
| Medium (65) | 742.5 a | 568.3 a | 15.4 a | 2.8 a | 367.8 a | 4.4 a | 59.3 a | 1760.3 a | |
| Strong (42) | 510.0 a | 342.0 a | 6.3 a | 1.3 a | 420.5 a | 4.5 a | 50.9 a | 1335.2 a | |
| Very strong (24) | 497.5 a | 342.2 a | 8.0 a | 0.5 a | 515.3 a | 5.8 a | 49.8 a | 1418.9 a | |
| Leaf waxiness | Weak (52) | 651.4 a | 367.6 a | 11.3 ab | 1.1 a | 449.4 a | 4.4 a | 63.6 b | 1548.6 ab |
| Medium (142) | 528.9 a | 417.0 ab | 9.6 a | 2.0 a | 377.2 a | 4.6 a | 45.5 a | 1382.4 a | |
| Strong (148) | 721.9 a | 619.7 b | 16.6 b | 2.5 a | 410.1 a | 5.0 a | 68.9 b | 1844.6 b | |
| Leaf blade: blistering | Absent/weak (80) | 663.2 a | 465.3 a | 13.4 a | 1.9 a | 415.5 a | 4.6 a | 61.9 a | 1625.8 a |
| Medium (118) | 590.0 a | 518.3 a | 13.1 a | 1.7 a | 368.9 a | 4.9 a | 57.5 a | 1552.3 a | |
| Strong (143) | 647.6 a | 496.0 a | 12.2 a | 2.5 a | 419.8 a | 4.6 a | 57.1 a | 1639.1 a | |
| Leaf blade: pubescence on the upper side | Absent (81) | 668.8 | 653.7 | 20.1 | 2.8 | 319.9 | 4.9 | 68.8 | 1738.9 |
| Present (263) | 633.5 | 449.3 | 10.8 | 1.9 | 427.2 | 4.7 | 55.3 | 1581.5 | |
| Leaf blade: pubescence on the lower side | Absent (54) | 677.9 b | 661.5 b | 20.5 b | 3.2 a | 310.7 a | 5.0 a | 74.1 b | 1752.8 b |
| Few (101) | 687.0 b | 483.3 ab | 14.2 ab | 1.7 a | 385.5 a | 4.9 a | 61.2 b | 1637.7 ab | |
| Medium (154) | 659.5 b | 478.9 ab | 11.0 a | 2.4 a | 449.0 a | 4.8 a | 55.9 ab | 1659.3 ab | |
| Many (35) | 378.1 a | 366.5 a | 7.2 a | 0.6 a | 383.0 a | 3.7 a | 37.9 a | 1177.0 a | |
| Stem hair | Absent (59) | 670.9 | 652.4 | 20.2 | 3.2 | 321.2 | 5.0 | 73.8 | 1746.5 |
| Present (291) | 630.8 | 461.0 | 11.4 | 1.9 | 417.9 | 4.7 | 55.0 | 1581.6 | |
| Midrib transection | Horizontal (41) | 837.8 b | 684.4 b | 22.8 b | 2.4 a | 434.0 a | 5.2 a | 73.1 b | 2059.7 b |
| Intermediate (122) | 589.1 a | 541.8 ab | 12.8 a | 2.0 a | 420.8 a | 4.8 a | 55.3 a | 1626.6 a | |
| Semi-circled (191) | 604.4 a | 422.8 a | 10.6 a | 2.0 a | 383.3 a | 4.6 a | 56.2 a | 1482.1 a | |
| Petiole anthocyanin coloration | Absent (77) | 468.6 | 418.8 | 8.7 | 1.0 | 385.1 | 4.3 | 43.2 | 1329.7 |
| Present (259) | 683.5 | 531.9 | 14.4 | 2.5 | 411.9 | 4.8 | 62.7 | 1710.5 | |
| Plant head formation | Absent (5) | 263.5 a | 216.4 a | 5.5 a | 0.2 a | 326.0 a | 3.5 a | 30.9 a | 846.0 a |
| Partially present (297) | 635.1 a | 483.5 a | 12.6 a | 2.0 a | 421.4 a | 4.8 a | 55.7 ab | 1613.9 a | |
| Fully present (52) | 697.7 a | 619.4 a | 15.6 a | 3.1 a | 301.8 a | 4.5 a | 75.0 b | 1716.9 a |
| Traits | PC1 | PC2 | PC3 | PC4 | PC5 |
|---|---|---|---|---|---|
| Leaf length | 0.36066 | 0.42513 | 0.050563 | −0.12065 | −0.014741 |
| Leaf width | 0.48364 | −0.08839 | −0.053683 | 0.099252 | 0.13102 |
| No of leaves | 0.14067 | 0.23187 | 0.90222 | −0.06798 | 0.070513 |
| Midrib width | 0.45814 | −0.24269 | 0.090063 | 0.072238 | 0.3238 |
| Midrib thickness | 0.35029 | 0.16403 | −0.055367 | 0.54633 | −0.70066 |
| Petiole length | −0.055534 | 0.67376 | −0.25685 | 0.37261 | 0.51955 |
| Petiole width | 0.43236 | −0.34385 | −0.16308 | 0.053491 | 0.27198 |
| Plant weight | 0.30598 | 0.31576 | −0.27717 | −0.72497 | −0.19486 |
| Eigenvalue | 3.78061 | 1.43425 | 0.997487 | 0.572543 | 0.398334 |
| % Variance | 47.258 | 17.928 | 12.469 | 7.1568 | 4.9792 |
| S/No | Traits | Top Ten Accessions * |
|---|---|---|
| 1 | Leaf length (≥50 cm) | IT120115, IT118972, IT118979, IT203623, IT228987, IT102888, Yesou86, IT215792, and IT120114 |
| 2 | Leaf width (≥28 cm) | Yeosu84, Yeosu 42, Yeosu61, IT228223, IT204152, Yeosu27, Yeosu 57, IT203623, Yeosu26, and Yeosu20 |
| 3 | No of leaves per plant (≥26 leaves) | IT228222, IT204153, IT215792, IT218388, IT235416, IT216873, IT118948, IT235418, IT180994, IT235344, IT216872, IT218451, IT218458, IT118955, and IT218449 |
| 4 | Midrib width (≥3.15 cm) | Yeosu80, IT228223, IT203623, Yeosu57, Yeosu15, IT204152, Yeosu84, IT228990, IT228988, and Yeosu61 |
| 5 | Midrib thickness (≥0.75 cm) | IT218357, Yeosu42, Yeosu27, IT141424, Yeosu41, IT102888, IT208801, Yeosu6, Yeosu15, IT118974, Yeosu11, and IT259499 |
| 6 | Petiole length (≥6.3 cm) | IT102941, IT102942, IT259517, IT100949, IT141423, IT228987, IT259499, IT100945, IT218383, and IT141422 |
| 7 | Petiole width (≥2.9 cm) | Yeosu57, Yeosu80, IT228223, Yeosu61, IT191096, IT228988, IT228988, IT204152, Yeosu21, and Yeosu24 |
| 8 | Plant weight (≥650 g) | IT228984, Yeosu45, IT118979, IT215792, IT228223, IT100949, IT248039, Yeosu42, Yeosu64, IT228987 |
| 9 | Sinigrin (≥2295) | IT237840, IT259503, IT248036, IT259487, IT236762, IT248037, IT250121, IT247853, IT235418, and Yeosu17 |
| 10 | Gluconapin (≥2000) | Yeosu66, Yeosu31, IT102941, Yeosu17, IT218460, IT191097, Yeosu41, Yeosu32, Yeosu2, IT250121, and IT259487 |
| 11 | Glucobrassicanapin (≥70) | Yeosu31, Yeosu37, Yeosu53, Yeosu41, Yeosu23, Yeosu17, Yeosu32, Yeosu44, Yeosu52, and Yeosu75 |
| 12 | Glucoiberin (≥16) | IT102894, Yeosu74, Yeosu2, IT259503, IT250121, Yeosu63, Yeosu52, IT259514, IT259492, and Yeosu38 |
| 13 | Gluconasturtiin (≥1340) | IT236761, IT248035, IT259491, IT248614, IT237839, IT235624, IT235803, Yeosu16, IT259513, and Yeosu23 |
| 14 | Glucobarbarin (≥13) | IT102937, IT102950, IT236761, IT109154, IT218453, IT248035, Yeosu43, IT235415, Yeosu80, and IT237839 |
| 15 | Glucobrassicin (≥180) | IT248036, IT248037, Yeosu66, Yeosu20, Yeosu19, Yeosu80, Yeosu21, IT259487, Yeosu18, and IT259509 |
| 16 | Total Glucosinolates (≥4600) | Yeosu66, IT259487, IT259503, IT237840, Yeosu17, IT248036, IT248035, IT250121, IT248037, and Yeosu31 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Assefa, A.D.; Kim, S.-H.; Ko, H.C.; Ro, N.; Subramanian, P.; Chung, Y.-J.; Lee, Y.-H.; Hahn, B.-S.; Rhee, J.-H. Leaf Mustard (Brassica juncea) Germplasm Resources Showed Diverse Characteristics in Agro-Morphological Traits and Glucosinolate Levels. Foods 2023, 12, 4374. https://doi.org/10.3390/foods12234374
Assefa AD, Kim S-H, Ko HC, Ro N, Subramanian P, Chung Y-J, Lee Y-H, Hahn B-S, Rhee J-H. Leaf Mustard (Brassica juncea) Germplasm Resources Showed Diverse Characteristics in Agro-Morphological Traits and Glucosinolate Levels. Foods. 2023; 12(23):4374. https://doi.org/10.3390/foods12234374
Chicago/Turabian StyleAssefa, Awraris Derbie, Seong-Hoon Kim, Ho Chul Ko, Nayoung Ro, Parthiban Subramanian, Yun-Jo Chung, Yong-Hyuk Lee, Bum-Soo Hahn, and Ju-Hee Rhee. 2023. "Leaf Mustard (Brassica juncea) Germplasm Resources Showed Diverse Characteristics in Agro-Morphological Traits and Glucosinolate Levels" Foods 12, no. 23: 4374. https://doi.org/10.3390/foods12234374