

Alfalfa Silage Diet Improves Meat Quality by Remodeling the Intestinal Microbes of Fattening Pigs
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
2.2. Assessment of Growth Performance
2.3. Assessment of Pork Quality and Fatty Acid Quantification
2.4. Amino Acid Profiling
2.5. Assessment of SCFAs in the Colon
2.6. 16S rRNA Sequencing
2.7. Statistical Analysis
3. Results
3.1. Growth Performance
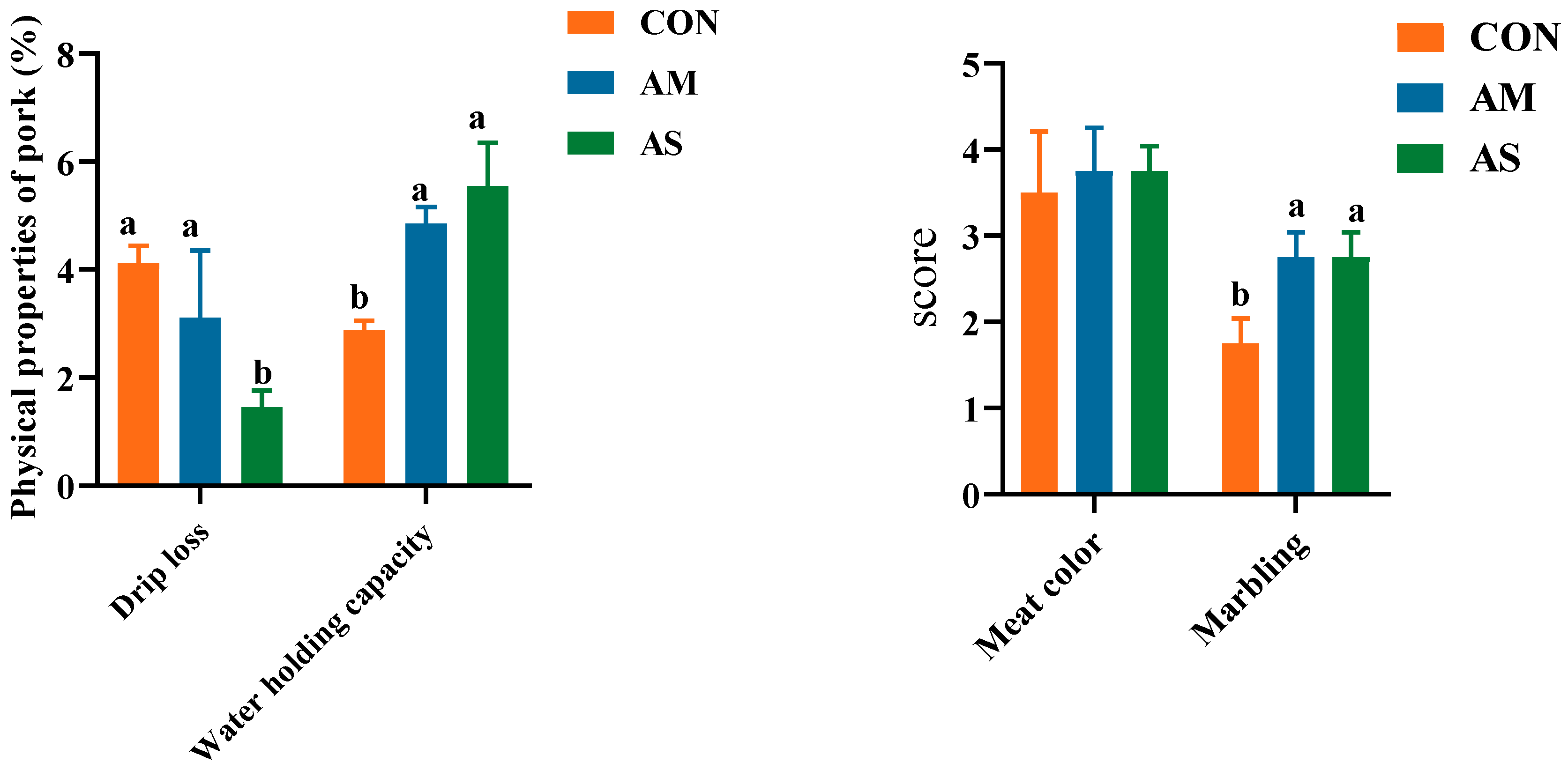
3.2. Meat Quality of Fattening Pigs
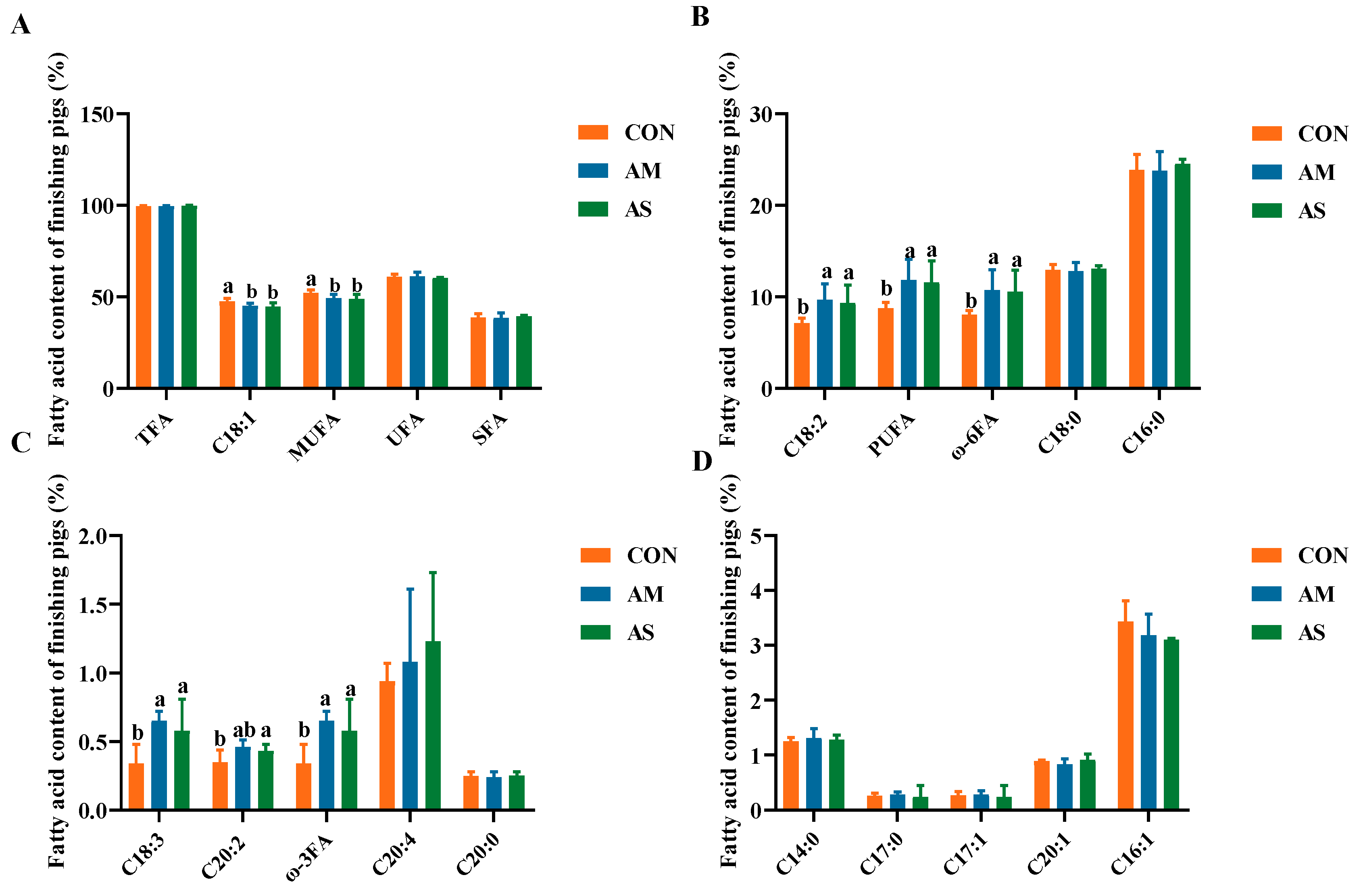
3.3. Muscle Fatty Acid Profile
3.4. Muscle Amino Acid Profile
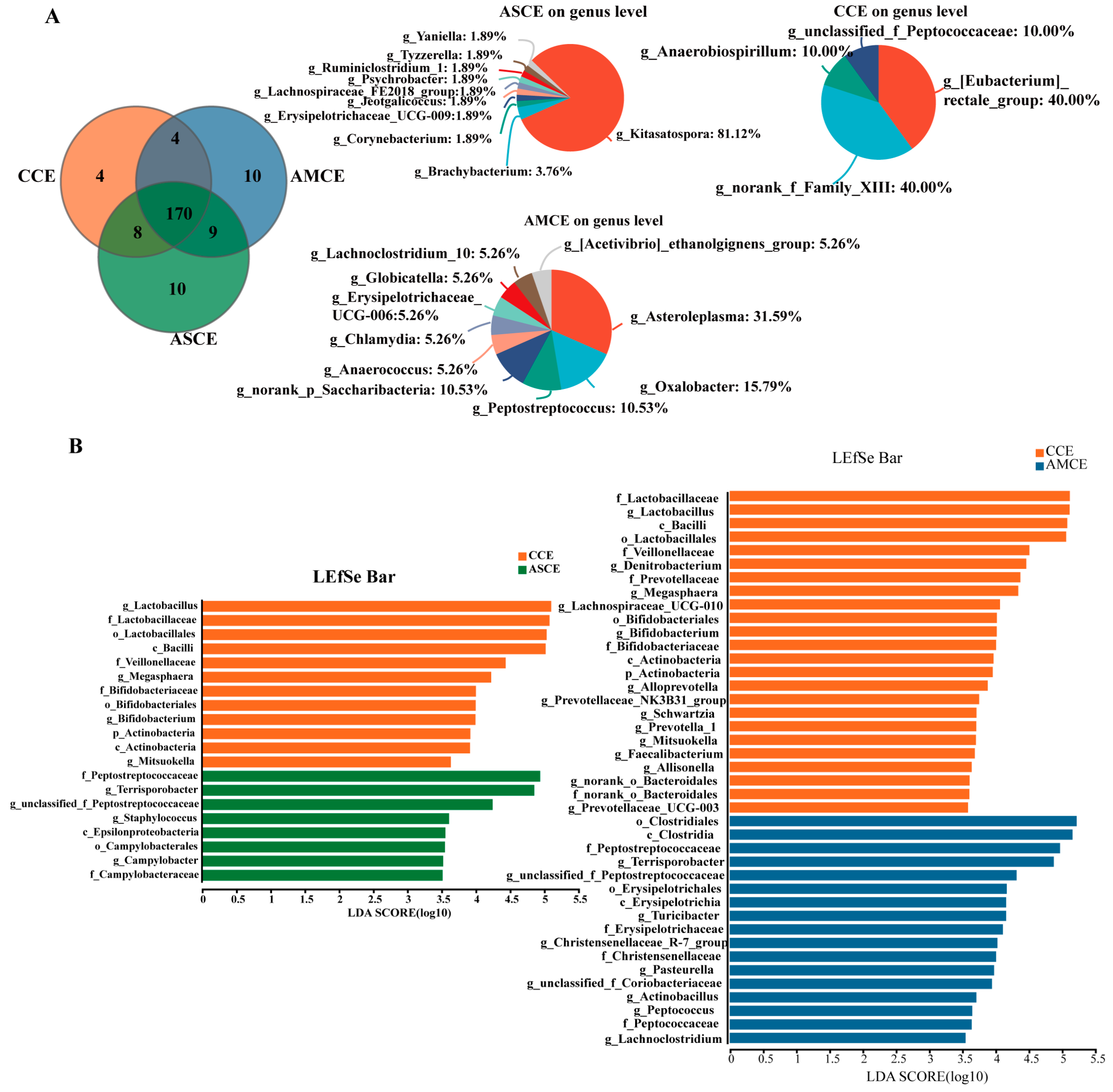
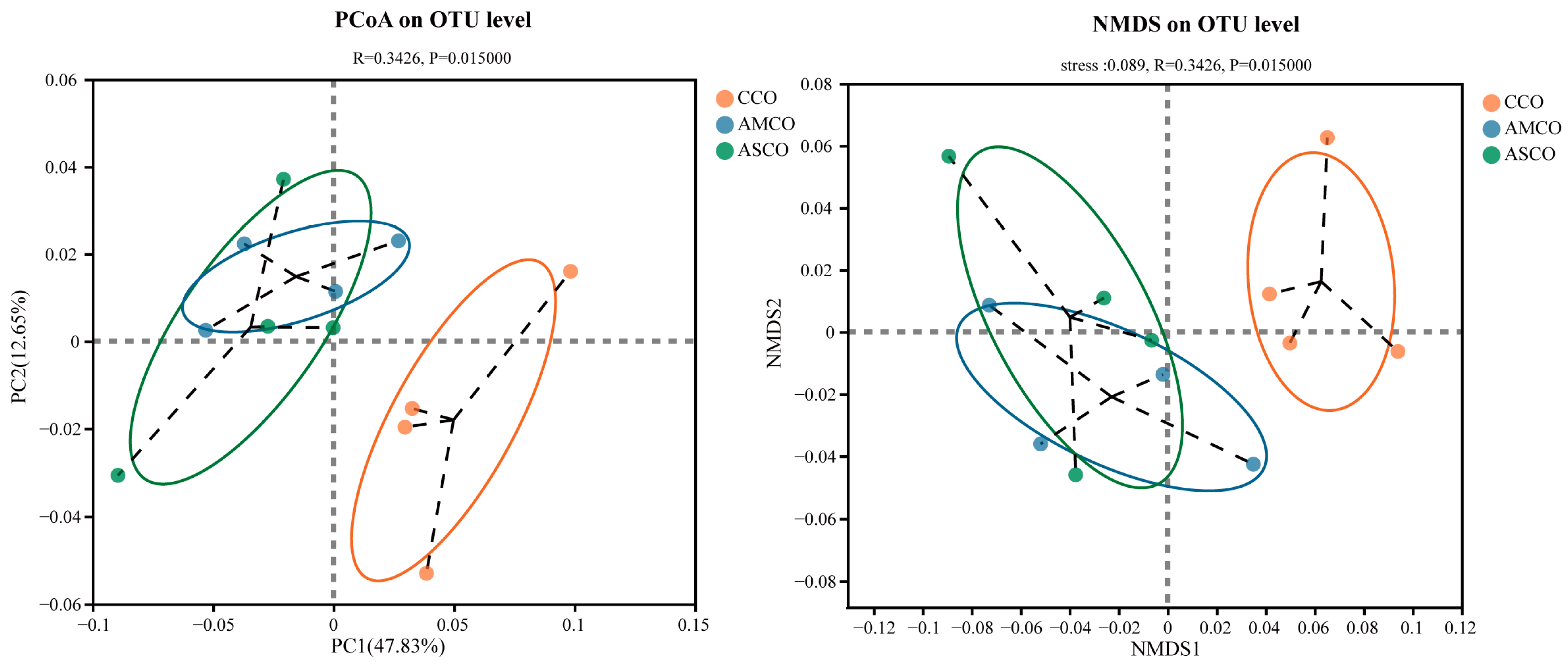
3.5. Distinct Cecum Microbial Compositions
3.6. Screening of Biomarkers for Microbes in in the Cecum
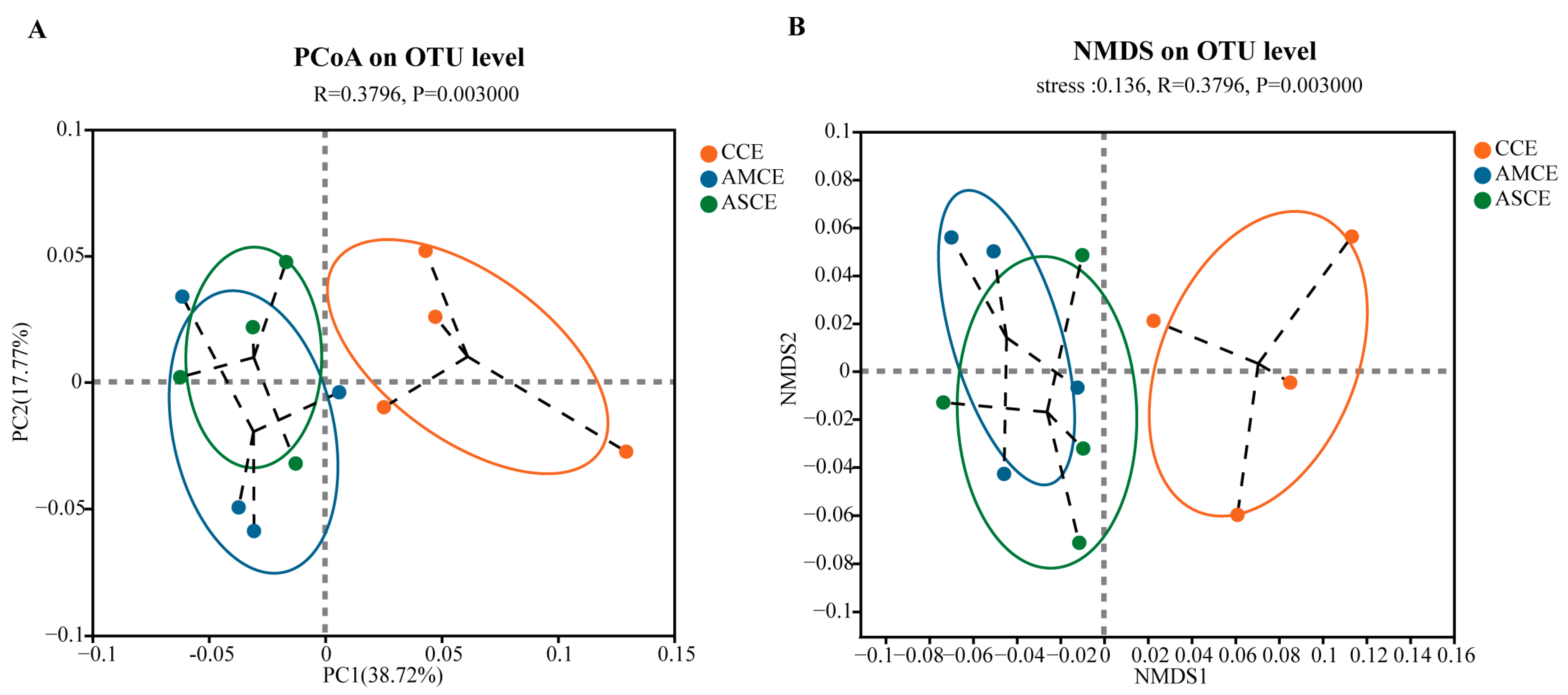
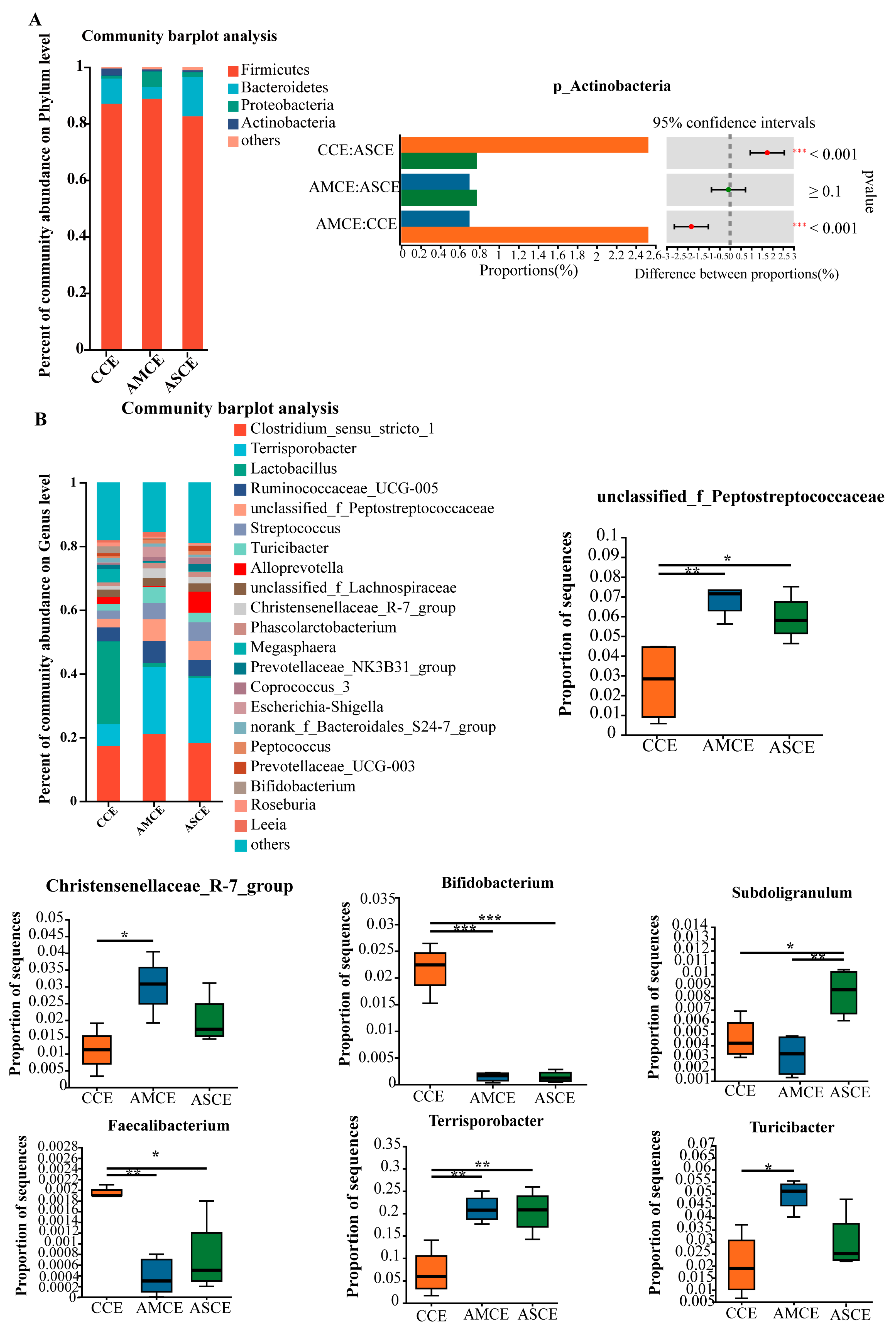
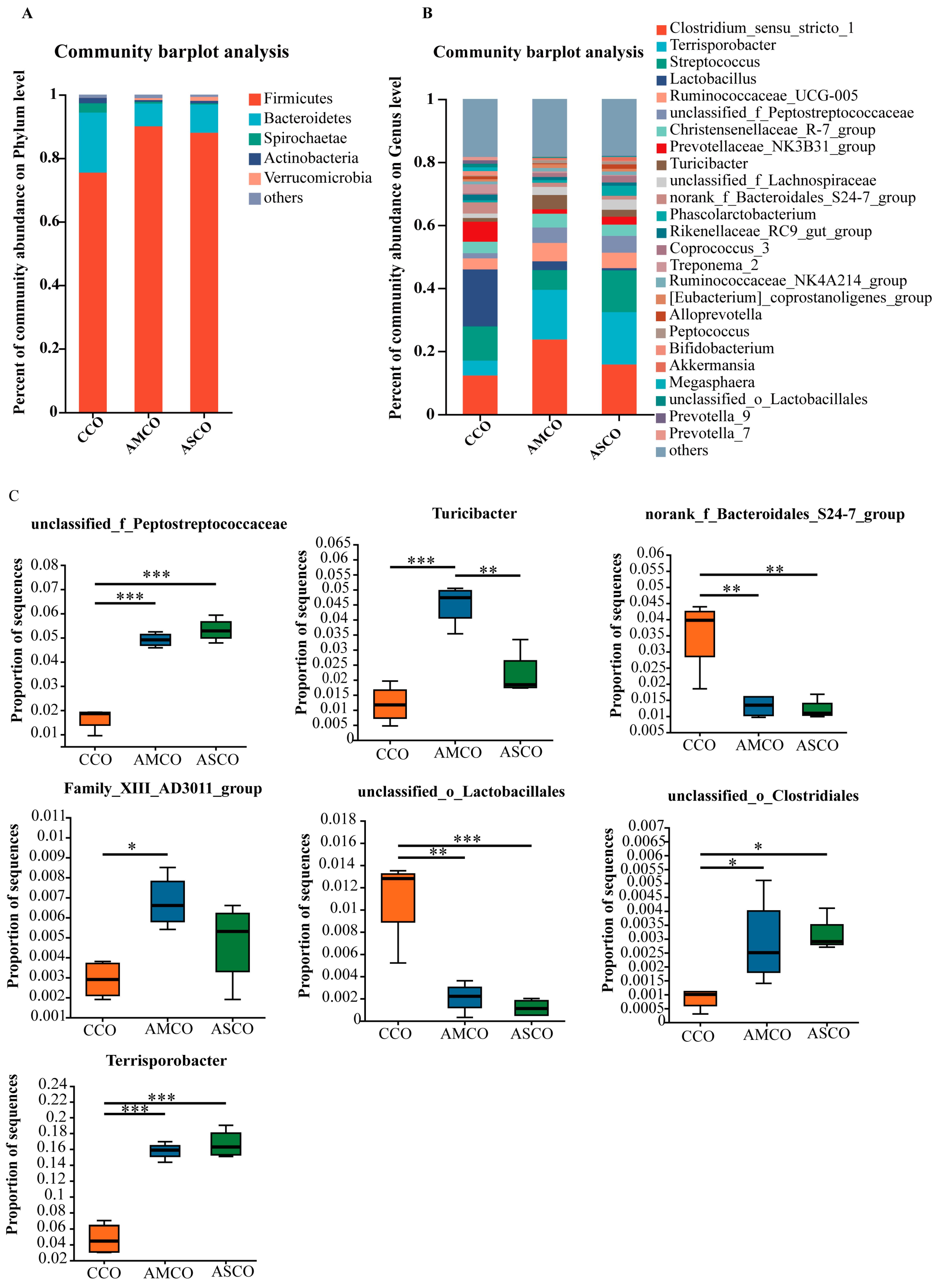
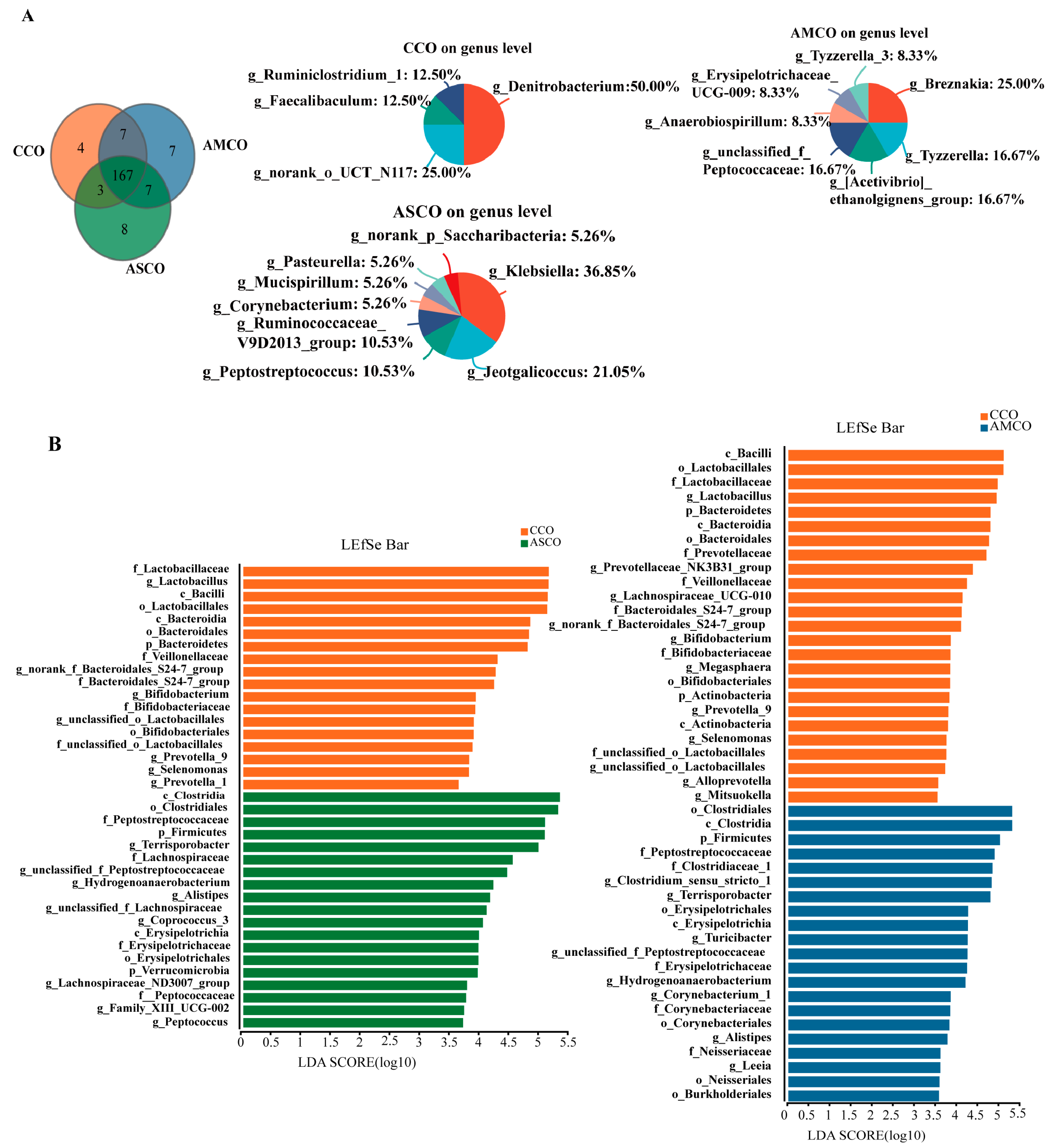
3.7. Distinct Colon Microbial Compositions
3.8. Screening of Biomarkers for Microbes in the Colon
3.9. Colon SCFAs
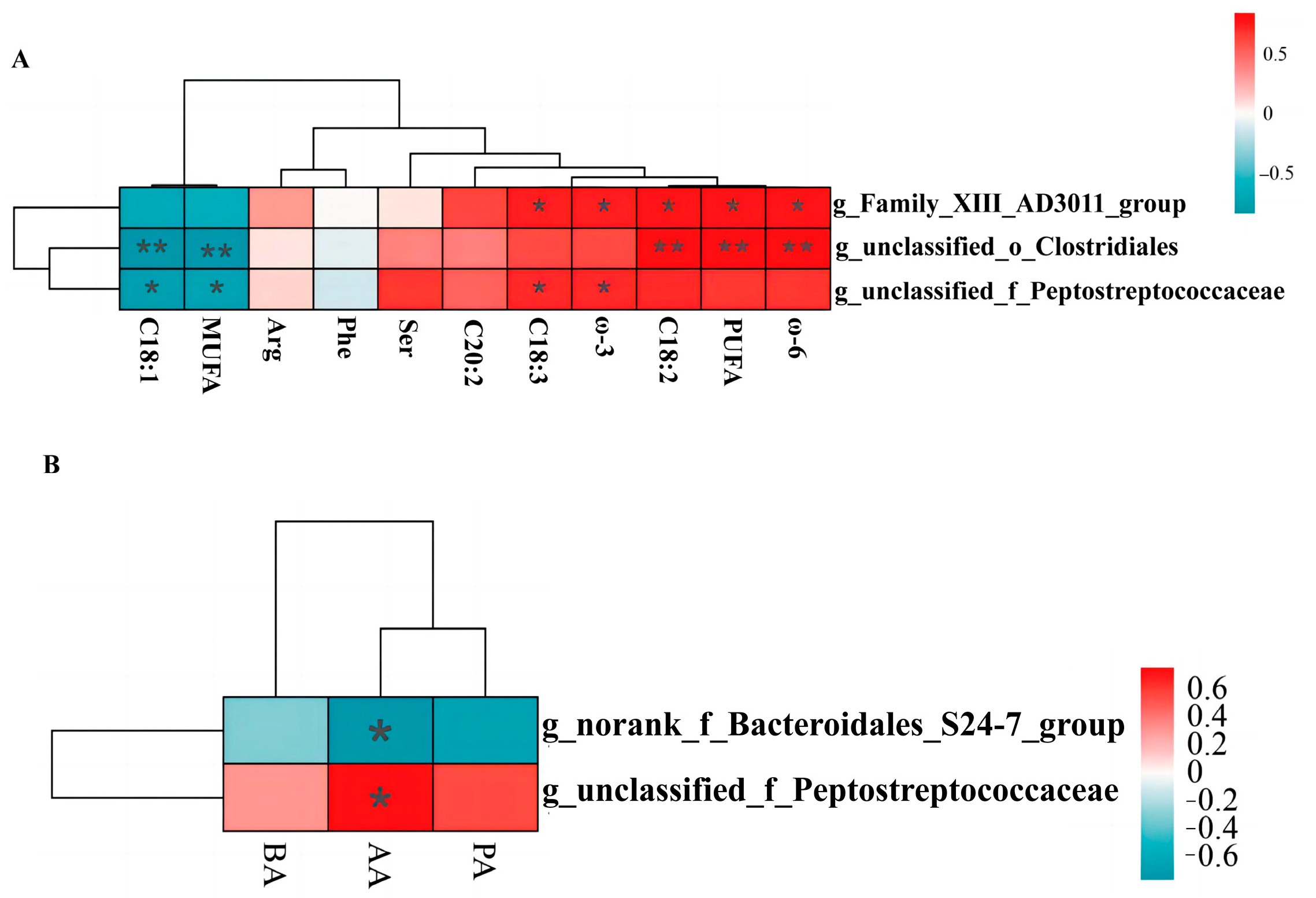
3.10. Correlation Analyses
4. Discussion
4.1. Effects of Different Diets on the Growth Performance of Fattening Pigs
4.2. Effects of Different Diets on the Meat Quality of Fattening Pigs
4.3. Effects of Different Diets on Gut Microbes and Their Metabolites in Fattening Pigs
4.4. Potential Mechanism to Improve Meat Quality
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zhang, H.; van der Wielen, N.; van der Hee, B.; Wang, J.; Hendriks, W.; Gilbert, M. Impact of Fermentable Protein, by Feeding High Protein Diets, on Microbial Composition, Microbial Catabolic Activity, Gut Health and beyond in Pigs. Microorganisms 2020, 8, 1735. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.; Cai, H.J. China Food Safety, Meeting the Challenges of Clenbuterol. Acta Aliment. 2015, 45, 331–337. [Google Scholar] [CrossRef]
- Xu, X.; Chen, X.; Chen, D.; Yu, B.; Yin, J.; Huang, Z. Effects of Dietary Apple Polyphenol Supplementation on Carcass Traits, Meat Quality, Muscle Amino Acid and Fatty Acid Composition in Finishing Pigs. Food Funct. 2019, 10, 7426–7434. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.F.; Song, X.M.; Huang, X.; Wu, J.L.; Zhou, W.D.; Zheng, H.C.; Jiang, Y.Q. Effects of Alfalfa Meal on Carcase Quality and Fat Metabolism of Muscovy Ducks. Br. Poult. Sci. 2012, 53, 681–688. [Google Scholar] [CrossRef] [PubMed]
- Zheng, M.; Niu, D.; Zuo, S.; Mao, P.; Meng, L.; Xu, C. The Effect of Cultivar, Wilting and Storage Period on Fermentation and the Clostridial Community of Alfalfa Silage. Ital. J. Anim. Sci. 2017, 17, 336–346. [Google Scholar] [CrossRef]
- Cui, Y.; Liu, B.; Sun, X.; Li, Z.; Chen, Y.; Guo, Z.; Liu, H.; Li, D.; Wang, C.; Zhu, X.; et al. Protective Effects of Alfalfa Saponins on Oxidative Stress-Induced Apoptotic Cells. Food Funct. 2020, 11, 8133–8140. [Google Scholar] [CrossRef]
- Lemmens, K.J.A.; van de Wier, B.; Vaes, N.; Ghosh, M.; van Zandvoort, M.A.M.J.; van der Vijgh, W.J.F.; Bast, A.; Haenen, G.R.M.M. The Flavonoid 7-Mono-O-(β-Hydroxyethyl)-Rutoside Is Able to Protect Endothelial Cells by a Direct Antioxidant Effect. Toxicol. In Vitro 2014, 28, 538–543. [Google Scholar] [CrossRef]
- Han, K.-H.; Kobayashi, Y.; Nakamura, Y.; Shimada, K.-I.; Aritsuka, T.; Ohba, K.; Morita, T.; Fukushima, M. Comparison of the Effects of Longer Chain Inulins with Different Degrees of Polymerization on Colonic Fermentation in a Mixed Culture of Swine Fecal Bacteria. J. Nutr. Sci. Vitaminol. 2014, 60, 206–212. [Google Scholar] [CrossRef]
- Su, Y.; Sun, X.; Zhao, S.; Hu, M.; Li, D.; Qi, S.; Jiao, X.; Sun, Y.; Wang, C.; Zhu, X.; et al. Dietary Alfalfa Powder Supplementation Improves Growth and Development, Body Health, and Meat Quality of Tibetan Sheep. Food Chem. 2022, 396, 133709. [Google Scholar] [CrossRef]
- Dal Bosco, A.; Mugnai, C.; Roscini, V.; Mattioli, S.; Ruggeri, S.; Castellini, C. Effect of Dietary Alfalfa on the Fatty Acid Composition and Indexes of Lipid Metabolism of Rabbit Meat. Meat Sci. 2014, 96, 606–609. [Google Scholar] [CrossRef]
- Liu, B.; Zhu, X.; Cui, Y.; Wang, W.; Liu, H.; Li, Z.; Guo, Z.; Ma, S.; Li, D.; Wang, C.; et al. Consumption of Dietary Fiber from Different Sources during Pregnancy Alters Sow Gut Microbiota and Improves Performance and Reduces Inflammation in Sows and Piglets. mSystems 2021, 6, e00591-20. [Google Scholar] [CrossRef] [PubMed]
- Wüstholz, J.; Carrasco, S.; Berger, U.; Sundrum, A.; Bellof, G. Fattening and Slaughtering Performance of Growing Pigs Consuming High level of Alfalfa Silage (Medicago sativa) in Organic Pig Production. Livest. Sci. 2017, 200, 46–52. [Google Scholar] [CrossRef]
- Szumacher-Strabel, M.; Stochmal, A.; Cieslak, A.; Kozłowska, M.; Kuznicki, D.; Kowalczyk, M.; Oleszek, W. Structural and Quantitative Changes of Saponins in Fresh Alfalfa Compared to Alfalfa Silage. J. Sci. Food Agric. 2019, 99, 2243–2250. [Google Scholar] [CrossRef] [PubMed]
- Carrasco, S.; Wüstholz, J.; Hahn, G.; Bellof, G. How Does Feeding Organic Broilers High Levels of Alfalfa Silage Affect the Meat Quality? Org. Agr. 2018, 8, 185–193. [Google Scholar] [CrossRef]
- Luo, Y.; Liu, Y.; Shen, Y.; He, J.; Li, H.; Lan, C.; Li, J.; Chen, H.; Chen, D.; Ren, Z.; et al. Fermented Alfalfa Meal Instead of “Grain-Type” Feedstuffs in the Diet Improves Intestinal Health Related Indexes in Weaned Pigs. Front. Microbiol. 2021, 12, 797875. [Google Scholar] [CrossRef]
- Niu, J.; Liu, X.; Xu, J.; Li, F.; Wang, J.; Zhang, X.; Yang, X.; Wang, L.; Ma, S.; Li, D.; et al. Effects of Silage Diet on Meat Quality through Shaping Gut Microbiota in Finishing Pigs. Microbiol. Spectr. 2023, 11, e0241622. [Google Scholar] [CrossRef]
- Mason, F.; Pascotto, E.; Zanfi, C.; Spanghero, M. Effect of Dietary Inclusion of Whole Ear Corn Silage on Stomach Development and Gastric Mucosa Integrity of Heavy Pigs at Slaughter. Vet. J. 2013, 198, 717–719. [Google Scholar] [CrossRef]
- Millet, S.; Meyns, T.; Aluwé, M.; De Brabander, D.; Ducatelle, R. Effect of Grinding Intensity and Crude Fibre Content of the Feed on Growth Performance and Gastric Mucosa Integrity of Growing–Finishing Pigs. Livest. Sci. 2010, 134, 152–154. [Google Scholar] [CrossRef]
- Millet, S.; Kumar, S.; De Boever, J.; Meyns, T.; Aluwé, M.; De Brabander, D.; Ducatelle, R. Effect of Particle Size Distribution and Dietary Crude Fibre Content on Growth Performance and Gastric Mucosa Integrity of Growing-Finishing Pigs. Vet. J. 2012, 192, 316–321. [Google Scholar] [CrossRef]
- Millet, S.; Kumar, S.; De Boever, J.; Ducatelle, R.; De Brabander, D. Effect of Feed Processing on Growth Performance and Gastric Mucosa Integrity in Pigs from Weaning until Slaughter. Anim. Feed. Sci. Technol. 2012, 175, 175–181. [Google Scholar] [CrossRef]
- Sousa, D.O.; Hansen, H.H.; Hallin, O.; Nussio, L.G.; Nadeau, E. A Two-Year Comparison on Nutritive Value and Yield of Eight Lucerne Cultivars and One Red Clover Cultivar. Grass Forage Sci. 2020, 75, 76–85. [Google Scholar] [CrossRef]
- Tucak, M.; Ravlić, M.; Horvat, D.; Čupić, T. Improvement of Forage Nutritive Quality of Alfalfa and Red Clover through Plant Breeding. Agronomy 2021, 11, 2176. [Google Scholar] [CrossRef]
- Kass, M.L.; Soest, P.J.V.; Pond, W.G. Utilization of Dietary Fiber from Alfalfa by Growing Swine. 2. Volatile Fatty Acid Concentrations in and Disappearance from the Gastrointestinal Tract. J. Anim. Sci. 1980, 50, 192–197. [Google Scholar] [CrossRef]
- Kung, L.; Shaver, R.D.; Grant, R.J.; Schmidt, R.J. Silage Review, Interpretation of Chemical, Microbial, and Organoleptic Components of Silages. J. Dairy Sci. 2018, 101, 4020–4033. [Google Scholar] [CrossRef] [PubMed]
- Banaszak, M.; Kuźniacka, J.; Biesek, J.; Maiorano, G.; Adamski, M. Meat Quality Traits and Fatty Acid Composition of Breast Muscles from Ducks Fed with Yellow Lupin. Animal 2020, 14, 1969–1975. [Google Scholar] [CrossRef]
- Kuzniacka, J.; Adamski, M.; Czarnecki, R.; Banaszak, M. Results of Rearing Broiler Chickens Under Various Systems. J. Agric. Sci. 2014, 6, 19. [Google Scholar] [CrossRef]
- Wu, Y.; Zhao, J.; Xu, C.; Ma, N.; He, T.; Zhao, J.; Ma, X.; Thacker, P.A. Progress towards Pig Nutrition in the Last 27 Years. J. Sci. Food Agric. 2020, 100, 5102–5110. [Google Scholar] [CrossRef]
- Qiu, Y.; Li, K.; Zhao, X.; Liu, S.; Wang, L.; Yang, X.; Jiang, Z. Fermented Feed Modulates Meat Quality and Promotes the Growth of Longissimus Thoracis of Late-Finishing Pigs. Animals 2020, 10, 1682. [Google Scholar] [CrossRef]
- Xu, B.; Zhu, L.; Fu, J.; Li, Z.; Wang, Y.; Jin, M. Overall Assessment of Fermented Feed for Pigs, A Series of Meta-Analyses. J. Anim. Sci. 2019, 97, 4810–4821. [Google Scholar] [CrossRef]
- Huff-Lonergan, E.; Lonergan, S.M. New Frontiers in Understanding Drip Loss in Pork, Recent Insights on the Role of Postmortem Muscle Biochemistry. J. Anim. Breed. Genet. 2007, 124, 19–26. [Google Scholar] [CrossRef]
- Warner, R.D. Chapter 14—The Eating Quality of Meat—IV Water-Holding Capacity and Juiciness. In Lawrie’s Meat Science, 8th ed.; Toldrá, F., Ed.; Woodhead Publishing Series in Food Science, Technology and Nutrition; Woodhead Publishing: Sawston, UK, 2017; pp. 419–459. ISBN 978-0-08-100694-8. [Google Scholar]
- Cheng, W.; Cheng, J.-H.; Sun, D.-W.; Pu, H. Marbling Analysis for Evaluating Meat Quality, Methods and Techniques. Compr. Rev. Food Sci. Food Saf. 2015, 14, 523–535. [Google Scholar] [CrossRef]
- Kim, H.; Lee, S.K.; Hong, M.W.; Park, S.R.; Lee, Y.S.; Kim, J.W.; Lee, H.K.; Jeong, D.K.; Song, Y.H.; Lee, S.J. Association of a Single Nucleotide Polymorphism in the Akirin 2 Gene with Economically Important Traits in Korean Native Cattle. Anim. Genet. 2013, 44, 750–753. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Wu, P.; Wang, K.; Wang, S.; Ji, X.; Shen, Q.; Yu, Y.; Qiu, X.; Xu, X.; Liu, Y.; et al. Combining Computer Vision Score and Conventional Meat Quality Traits to Estimate the Intramuscular Fat Content Using Machine Learning in Pigs. Meat Sci. 2022, 185, 108727. [Google Scholar] [CrossRef] [PubMed]
- Seong, J.; Yoon, H.; Kong, H.S. Identification of MicroRNA and Target Gene Associated with Marbling Score in Korean Cattle (Hanwoo). Genes Genom. 2016, 38, 529–538. [Google Scholar] [CrossRef]
- Russo, G.L. Dietary N-6 and n-3 Polyunsaturated Fatty Acids, From Biochemistry to Clinical Implications in Cardiovascular Prevention. Biochem. Pharmacol. 2009, 77, 937–946. [Google Scholar] [CrossRef]
- Calder, P.C. Functional Roles of Fatty Acids and Their Effects on Human Health. J. Parenter. Enteral Nutr. 2015, 39, 18S–32S. [Google Scholar] [CrossRef]
- Jasani, B.; Simmer, K.; Patole, S.K.; Rao, S.C. Long Chain Polyunsaturated Fatty Acid Supplementation in Infants Born at Term. Cochrane Database Syst. Rev. 2017, 3, CD000376. [Google Scholar] [CrossRef]
- Oppedisano, F.; Macrì, R.; Gliozzi, M.; Musolino, V.; Carresi, C.; Maiuolo, J.; Bosco, F.; Nucera, S.; Caterina Zito, M.; Guarnieri, L.; et al. The Anti-Inflammatory and Antioxidant Properties of n-3 PUFAs: Their Role in Cardiovascular Protection. Biomedicines 2020, 8, 306. [Google Scholar] [CrossRef]
- Calder, P.C. N-3 PUFA and Inflammation, From Membrane to Nucleus and from Bench to Bedside. Proc. Nutr. Soc. 2020, 79, 404–416. [Google Scholar] [CrossRef]
- Ji, X.-J.; Huang, H. Engineering Microbes to Produce Polyunsaturated Fatty Acids. Trends Biotechnol. 2019, 37, 344–346. [Google Scholar] [CrossRef]
- Motohiro, A.; Furukawa, T.; Yasumoto, K.; Inokuchi, K. Mechanisms Involved in Acute Lung Edema Induced in Dogs by Oleic Acid. Eur. Surg. Res. 1986, 18, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves-de-Albuquerque, C.F.; Silva, A.R.; Burth, P.; Castro-Faria, M.V.; Castro-Faria-Neto, H.C. Acute Respiratory Distress Syndrome, Role of Oleic Acid-Triggered Lung Injury and Inflammation. Mediat. Inflamm. 2015, 2015, 260465. [Google Scholar] [CrossRef] [PubMed]
- Benz, J.M.; Tokach, M.D.; Dritz, S.S.; Nelssen, J.L.; DeRouchey, J.M.; Sulabo, R.C.; Goodband, R.D. Effects of Choice White Grease and Soybean Oil on Growth Performance, Carcass Characteristics, and Carcass Fat Quality of Growing-Finishing Pigs. J. Anim. Sci. 2011, 89, 404–413. [Google Scholar] [CrossRef] [PubMed]
- Alencar, S.A.D.S.; Kiefer, C.; Nascimento, K.M.R.D.S.; Viana, L.H.; Corassa, A.; Gomes, M.D.N.B.; Marçal, D.A.; Farias, T.V.A. Dietary Soybean Oil Modulates Fatty Acid Composition of Pork. Trop. Anim. Health Prod. 2021, 53, 357. [Google Scholar] [CrossRef]
- Sabagh, E.; Taha, N.; Saleh, E.; ElFar, A.; Sadek, K. Effect of Freezing and Frozen Storage on Amino Acid Profile and Fatty Acid Pattern in Imported and Local Meat. Alex. J. Vet. Sci. 2016, 49, 113. [Google Scholar] [CrossRef]
- Chen, G.S.; Liu, M.Z. Analysis of Nutritive Characteristics of Muscles in Wild Boar Hybrids. Swine Prod. 2004, 1, 24–27. [Google Scholar]
- Dibner, J.J.; Richards, J.D.; Knight, C.D. Microbial Imprinting in Gut Development and Health1. J. Appl. Poult. Res. 2008, 17, 174–188. [Google Scholar] [CrossRef]
- Satora, M.; Magdziarz, M.; Rząsa, A.; Rypuła, K.; Płoneczka-Janeczko, K. Insight into the Intestinal Microbiome of Farrowing Sows Following the Administration of Garlic (Allium sativum) Extract and Probiotic Bacteria Cultures under Farming Conditions. BMC Vet. Res. 2020, 16, 442. [Google Scholar] [CrossRef]
- den Hengst, C.D.; Buttner, M.J. Redox Control in Actinobacteria. Biochim. Biophys. Acta 2008, 1780, 1201–1216. [Google Scholar] [CrossRef]
- Tracy, B.P.; Jones, S.W.; Fast, A.G.; Indurthi, D.C.; Papoutsakis, E.T. Clostridia, The Importance of Their Exceptional Substrate and Metabolite Diversity for Biofuel and Biorefinery Applications. Curr. Opin. Biotechnol. 2012, 23, 364–381. [Google Scholar] [CrossRef]
- Gerritsen, J.; Fuentes, S.; Grievink, W.; van Niftrik, L.; Tindall, B.J.; Timmerman, H.M.; Rijkers, G.T.; Smidt, H. Characterization of Romboutsia Ilealis Gen. Nov., Sp. Nov., Isolated from the Gastro-Intestinal Tract of a Rat, and Proposal for the Reclassification of Five Closely Related Members of the Genus Clostridium into the Genera Romboutsia Gen. Nov., Intestinibacter Gen. Nov., Terrisporobacter Gen. Nov. and Asaccharospora Gen. Nov. Int. J. Syst. Evol. Microbiol. 2014, 64, 1600–1616. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Liang, Z.; Zhang, Y. Evolution of Physicochemical Properties and Bacterial Community in Aerobic Composting of Swine Manure Based on a Patent Compost Tray. Bioresour. Technol. 2022, 343, 126136. [Google Scholar] [CrossRef] [PubMed]
- Verbarg, S.; Göker, M.; Scheuner, C.; Schumann, P.; Stackebrandt, E. The Families Erysipelotrichaceae Emend., Coprobacillaceae Fam. Nov. In The Prokaryotes, Firmicutes and Tenericutes; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 79–105. ISBN 978-3-642-30120-9. [Google Scholar]
- Rosenberg, E.; DeLong, E.F.; Lory, S.; Stackebrandt, E.; Thompson, F. The Prokaryotes, Firmicutes and Tenericutes; Springer: Berlin/Heidelberg, Germany, 2013; p. 567. ISBN 978-3-642-30119-3. [Google Scholar]
- Maki, J.J.; Looft, T. Turicibacter Bilis Sp. Nov., a Novel Bacterium Isolated from the Chicken Eggshell and Swine Ileum. Int. J. Syst. Evol. Microbiol. 2022, 72, 005153. [Google Scholar] [CrossRef]
- Waters, J.L.; Ley, R.E. The Human Gut Bacteria Christensenellaceae Are Widespread, Heritable, and Associated with Health. BMC Biol. 2019, 17, 83. [Google Scholar] [CrossRef]
- Beaumont, M.; Portune, K.J.; Steuer, N.; Lan, A.; Cerrudo, V.; Audebert, M.; Dumont, F.; Mancano, G.; Khodorova, N.; Andriamihaja, M.; et al. Quantity and Source of Dietary Protein Influence Metabolite Production by Gut Microbiota and Rectal Mucosa Gene Expression, A Randomized, Parallel, Double-Blind Trial in Overweight Humans. Am. J. Clin. Nutr. 2017, 106, 1005–1019. [Google Scholar] [CrossRef] [PubMed]
- Manor, O.; Zubair, N.; Conomos, M.P.; Xu, X.; Rohwer, J.E.; Krafft, C.E.; Lovejoy, J.C.; Magis, A.T. A Multi-Omic Association Study of Trimethylamine N-Oxide. Cell Rep. 2018, 24, 935–946. [Google Scholar] [CrossRef]
- Rosenvold, K.; Andersen, H.J. Factors of Significance for Pork Quality-a Review. Meat Sci. 2003, 64, 219–237. [Google Scholar] [CrossRef] [PubMed]
- Kwan, S.-Y.; Sabotta, C.M.; Joon, A.; Wei, P.; Petty, L.E.; Below, J.E.; Wu, X.; Zhang, J.; Jenq, R.R.; Hawk, E.T.; et al. Gut Microbiome Alterations Associated with Diabetes in Mexican Americans in South Texas. mSystems 2022, 7, e0003322. [Google Scholar] [CrossRef]
- Castonguay-Paradis, S.; Lacroix, S.; Rochefort, G.; Parent, L.; Perron, J.; Martin, C.; Lamarche, B.; Raymond, F.; Flamand, N.; Di Marzo, V.; et al. Dietary Fatty Acid Intake and Gut Microbiota Determine Circulating Endocannabinoidome Signaling beyond the Effect of Body Fat. Sci. Rep. 2020, 10, 15975. [Google Scholar] [CrossRef]
- Xie, J.; Li, L.-F.; Dai, T.-Y.; Qi, X.; Wang, Y.; Zheng, T.-Z.; Gao, X.-Y.; Zhang, Y.-J.; Ai, Y.; Ma, L.; et al. Short-Chain Fatty Acids Produced by Ruminococcaceae Mediate α-Linolenic Acid Promote Intestinal Stem Cells Proliferation. Mol. Nutr. Food Res. 2022, 66, e2100408. [Google Scholar] [CrossRef]
- Kennedy, A.; Martinez, K.; Chuang, C.-C.; LaPoint, K.; McIntosh, M. Saturated Fatty Acid-Mediated Inflammation and Insulin Resistance in Adipose Tissue, Mechanisms of Action and Implications. J. Nutr. 2009, 139, 1–4. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
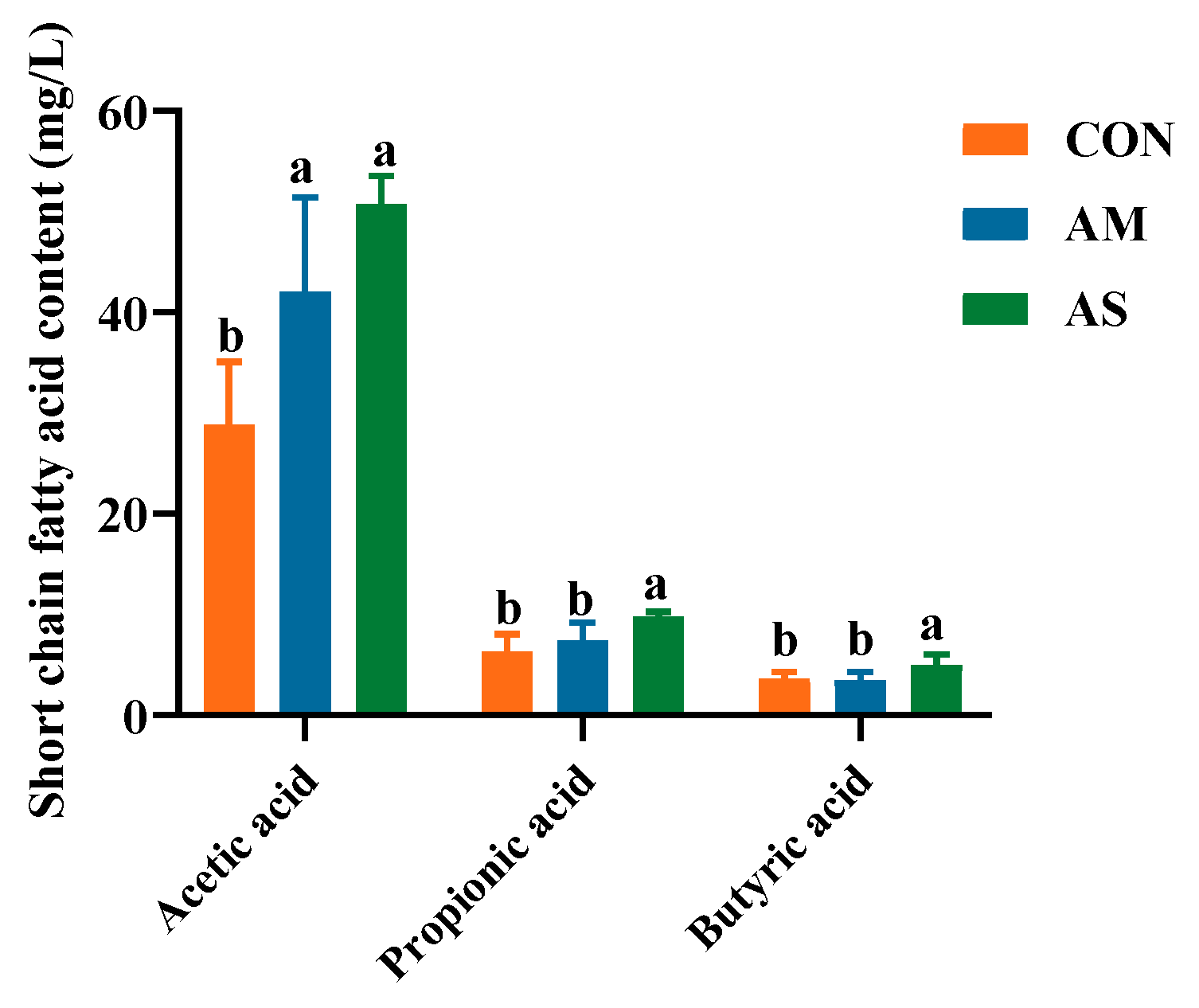{kind=link}
{kind=link}
| DM (%) | CP (%) | EE (%) | NDF (%) | ADF (%) | |
|---|---|---|---|---|---|
| Alfalfa silage | 23.21 | 17.38 | 2.54 | 36.87 | 27.55 |
| Alfalfa meal | 92.43 | 15.81 | 1.79 | 43.93 | 30.99 |
| Items | CON | AM | AS | Items | CON | AM | AS |
|---|---|---|---|---|---|---|---|
| Ingredient | Nutrient composition | ||||||
| Corn (%) | 72.41 | 65.99 | 67.18 | DE (MJ/kg) | 13.67 | 13.67 | 13.63 |
| Soya bean meal (%) | 18.12 | 18.03 | 16.88 | CP (%) | 15.19 | 15.77 | 15.77 |
| Soybean oil (%) | 0.00 | 2.00 | 2.18 | EE (%) | 4.47 | 6.11 | 6.39 |
| Bran (%) | 7.20 | 2.00 | 1.72 | NDF (%) | 12.24 | 13.15 | 13.44 |
| Calcium hydrogen phosphate (%) | 0.70 | 0.71 | 0.71 | ADF (%) | 5.13 | 6.78 | 7.24 |
| Limestone powder (%) | 0.36 | 0.10 | 0.13 | Ca (%) | 0.51 | 0.51 | 0.54 |
| Alfalfa Meal (%) | 0.00 | 10.00 | 0.00 | P (%) | 0.49 | 0.45 | 0.45 |
| Alfalfa Silage (%) | 0.00 | 0.00 | 10.00 | AP (%) | 0.24 | 0.25 | 0.25 |
| 1% premix (%) | 1.00 | 1.00 | 1.00 | Lys (%) | 0.95 | 0.95 | 0.96 |
| Lysine, 98% (%) | 0.21 | 0.17 | 0.20 | ||||
| Total (%) | 100.00 | 100.00 | 100.00 |
| Groups | ADG (kg) | ADFI (kg) | FCR |
|---|---|---|---|
| CON | 0.85 ± 0.07 | 2.70 ± 0.02 | 3.19 ± 0.24 |
| AM | 0.88 ± 0.04 | 2.77 ± 0.16 | 3.15 ± 0.21 |
| AS | 0.92 ± 0.03 | 2.71 ± 0.15 | 2.94 ± 0.09 |
| Items | CON | AM | AS | Items | CON | AM | AS |
|---|---|---|---|---|---|---|---|
| Asp | 2.02 ± 0.04 | 2.05 ± 0.03 | 2.02 ± 0.06 | leu | 1.77 ± 0.03 | 1.79 ± 0.02 | 1.77 ± 0.05 |
| Thr | 0.94 ± 0.01 | 0.94 ± 0.02 | 0.94 ± 0.04 | Tyr | 0.76 ± 0.02 | 0.77 ± 0.03 | 0.75 ± 0.02 |
| Ser | 0.74 ± 0.01 b | 0.76 ± 0.02 ab | 0.79 ± 0.03 a | Phe | 1.13 ± 0.03 b | 1.17 ± 0.01 a | 0.91 ± 0.01 c |
| Glu | 3.38 ± 0.07 | 3.43 ± 0.04 | 3.40 ± 0.07 | Lys | 2.08 ± 0.03 | 2.11 ± 0.03 | 2.12 ± 0.06 |
| Gly | 0.95 ± 0.05 | 0.98 ± 0.02 | 0.95 ± 0.02 | His | 1.11 ± 0.06 | 1.16 ± 0.04 | 1.13 ± 0.06 |
| Ala | 1.23 ± 0.03 | 1.25 ± 0.02 | 1.23 ± 0.03 | Arg | 1.50 ± 0.03 | 1.52 ± 0.01 | 1.48 ± 0.04 |
| Val | 1.14 ± 0.02 | 1.15 ± 0.02 | 1.13 ± 0.02 | Pro | 1.50 ± 0.03 | 0.82 ± 0.02 | 0.83 ± 0.02 |
| Met | 0.64 ± 0.02 | 0.63 ± 0.01 | 0.65 ± 0.04 | Trp | 0.22 ± 0.01 | 0.23 ± 0.01 | 0.24 ± 0.03 |
| ILe | 1.05 ± 0.02 | 1.08 ± 0.01 | 1.06 ± 0.03 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, J.; Liu, X.; Geng, H.; Liu, R.; Li, F.; Ma, J.; Liu, M.; Liu, B.; Sun, H.; Ma, S.; et al. Alfalfa Silage Diet Improves Meat Quality by Remodeling the Intestinal Microbes of Fattening Pigs. Foods 2023, 12, 3209. https://doi.org/10.3390/foods12173209
Xu J, Liu X, Geng H, Liu R, Li F, Ma J, Liu M, Liu B, Sun H, Ma S, et al. Alfalfa Silage Diet Improves Meat Quality by Remodeling the Intestinal Microbes of Fattening Pigs. Foods. 2023; 12(17):3209. https://doi.org/10.3390/foods12173209
Chicago/Turabian StyleXu, Junying, Xiao Liu, Hongmin Geng, Rui Liu, Fang Li, Jixiang Ma, Mengqi Liu, Boshuai Liu, Hao Sun, Sen Ma, and et al. 2023. "Alfalfa Silage Diet Improves Meat Quality by Remodeling the Intestinal Microbes of Fattening Pigs" Foods 12, no. 17: 3209. https://doi.org/10.3390/foods12173209