
Bioactive Compounds and Antioxidant Activity in Seeds of Bred Lines of Common Bean Developed from Interspecific Crosses
 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Preparation of Methanolic Extracts
2.3. Phenolic Compounds
2.3.1. Total Phenolic Content (TPC)
2.3.2. Total Flavonoid Content (TFC)
2.3.3. Total Condensed Tannin Content (TCTC)
2.4. Pigments
2.4.1. Total Monomeric Anthocyanin Content (MAC)
2.4.2. Total Carotenoid Content (TCC)
2.5. Antioxidant Activity
2.5.1. DPPH Radical Scavenging Activity (DRSA)
2.5.2. ABTS-+ Radical Scavenging Activity (ARSA)
2.5.3. Reducing Capacity FRAP (Ferric Reducing Antioxidant Power)
2.6. Sugar Content
2.6.1. Total Sugar Content (TSC)
2.6.2. Total Reducing Sugar Content (TRSC)
2.7. Data Analysis
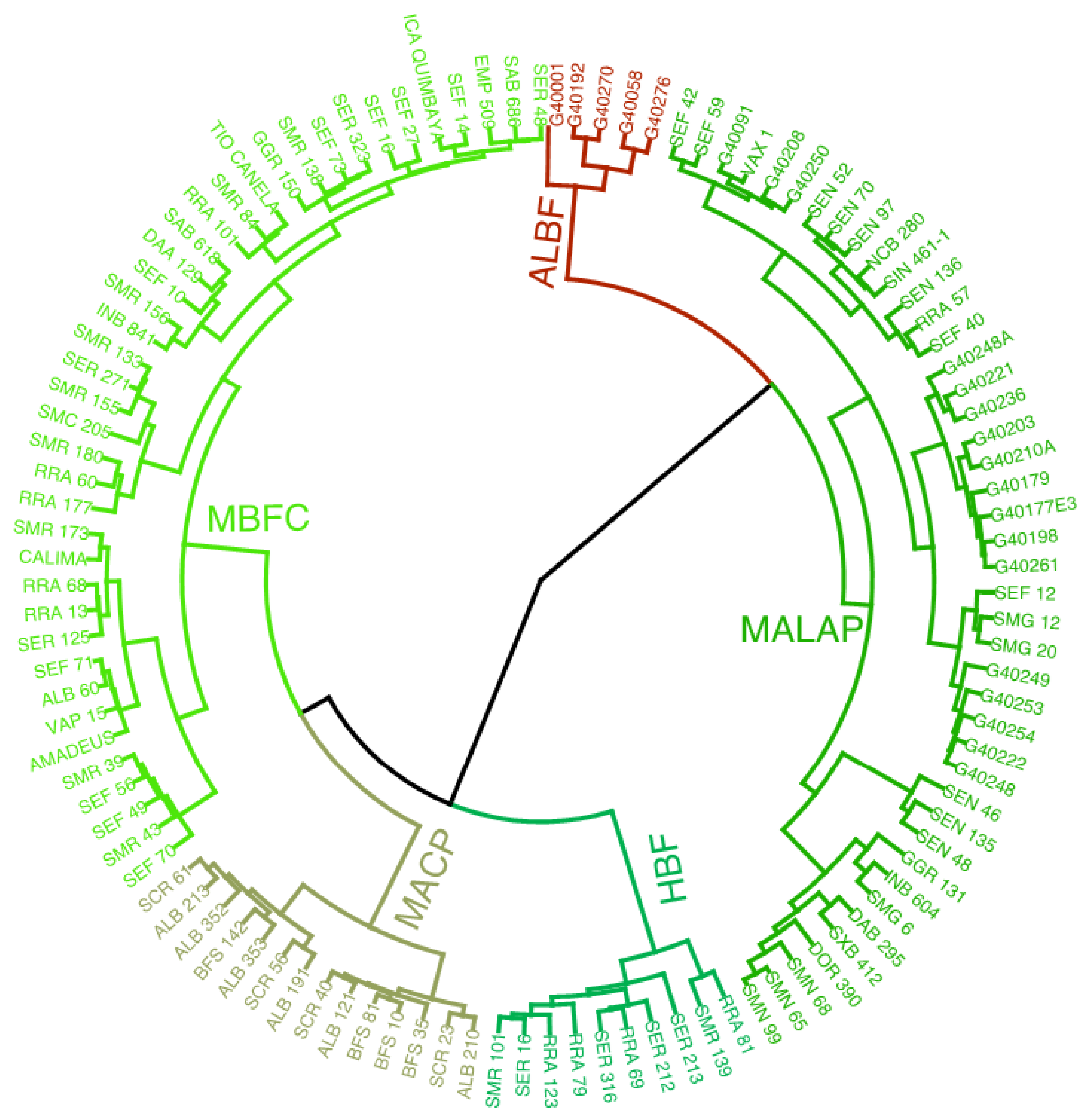
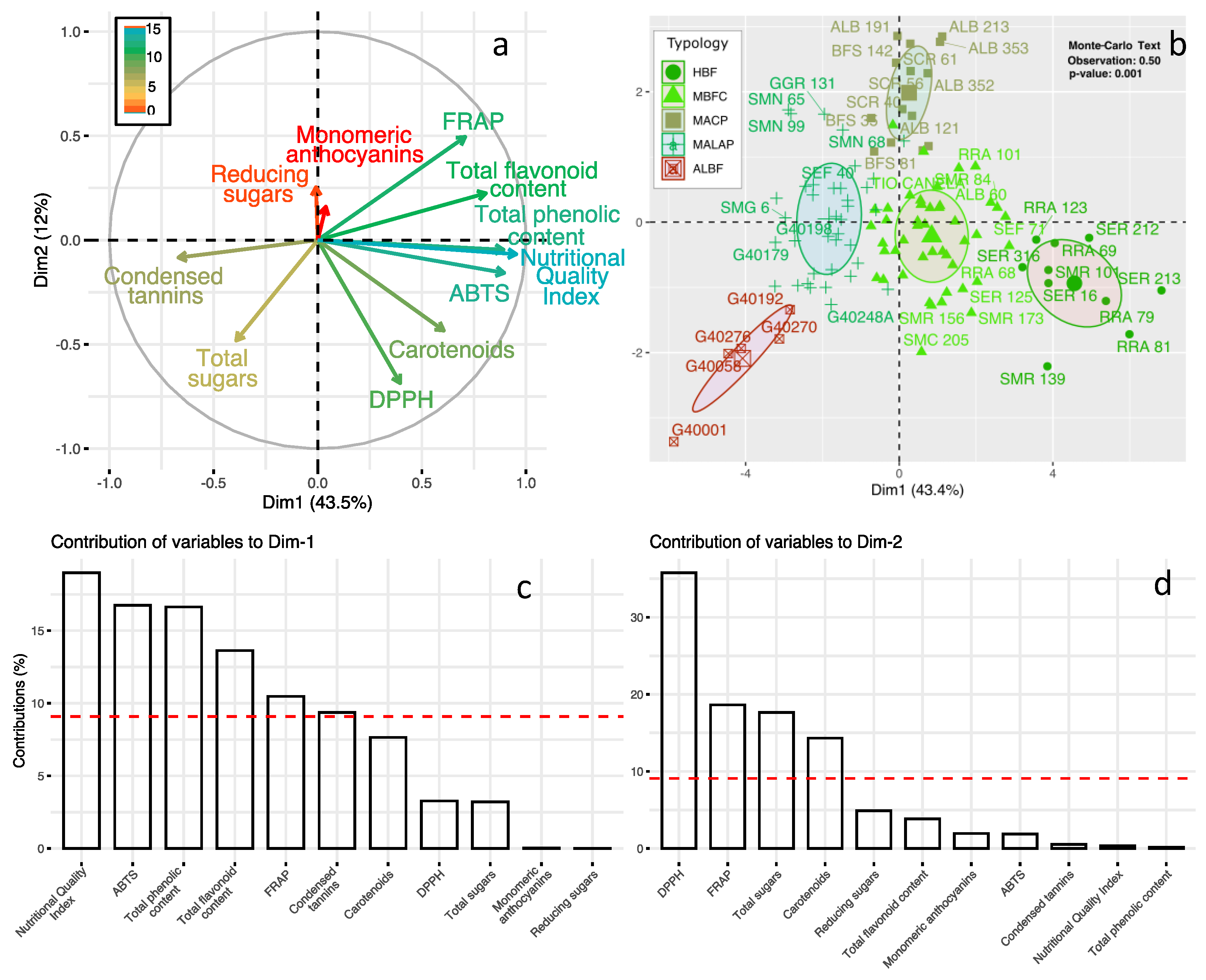
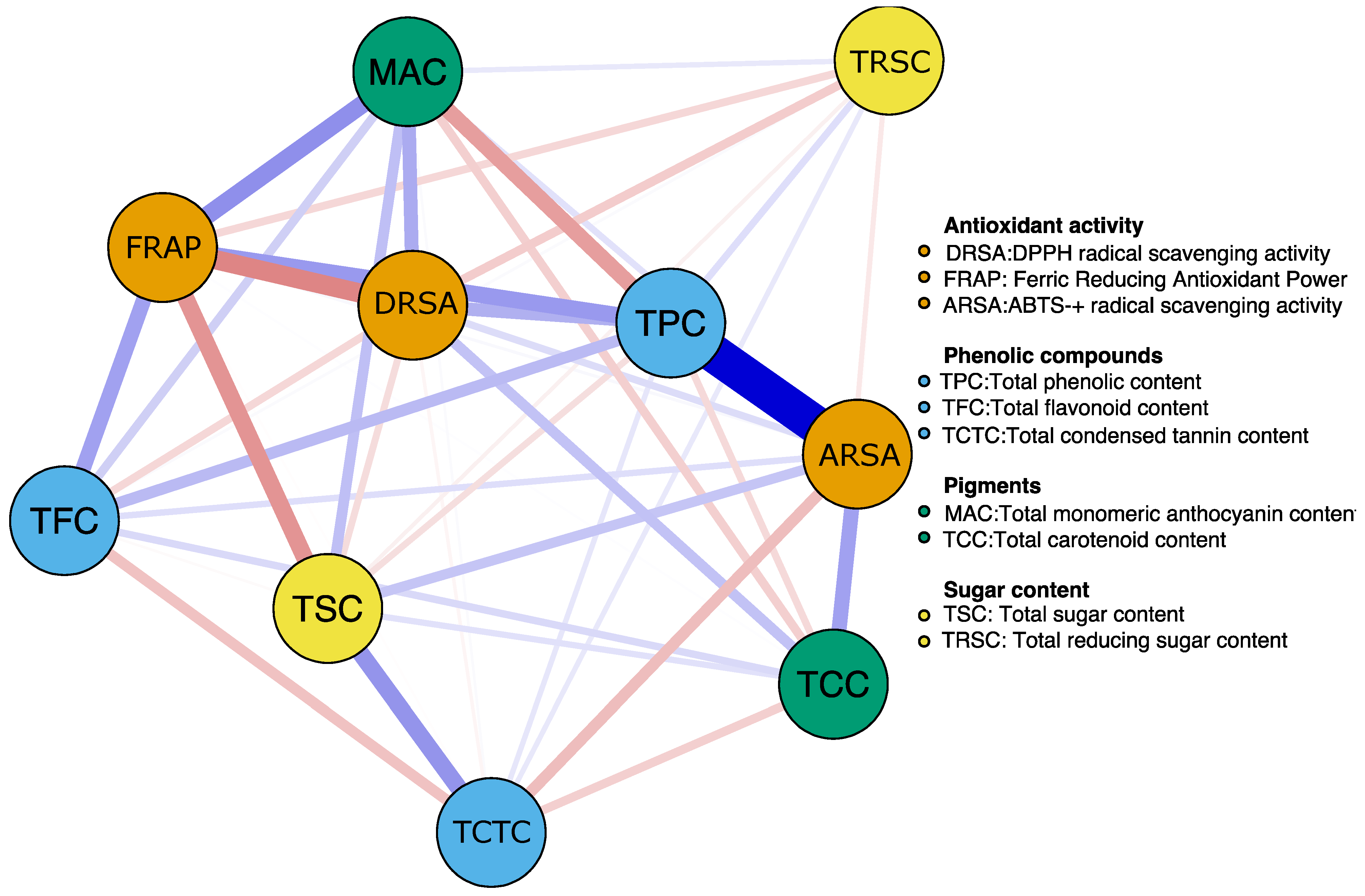
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Heredia-Rodríguez, L.; Gaytán-Martínez, M.; Morales-Sánchez, E.; Garza-Juárez, A.D.J.; Urias-Orona, V.; González-Martínez, B.E.; Lomelí, M.L.-C.; Vázquez-Rodríguez, J.A. Nutritional and technological properties of Tepary bean (Phaseolus acutifolius) cultivated in Mexican Northeast. Czech J. Food Sci. 2019, 37, 62–68. [Google Scholar] [CrossRef] [Green Version]
- Mudryj, A.N.; Yu, N.; Aukema, H.M. Nutritional and health benefits of pulses. In Applied Physiology, Nutrition and Metabolism; NRC Research Press: Ottawa, ON, Canada, 2014; Volume 39, pp. 1197–1204. [Google Scholar]
- Capistrán-Carabarin, A.; Aquino-Bolaños, E.N.; García-Díaz, Y.D.; Chávez-Servia, J.L.; Vera-Guzmán, A.M.; Carrillo-Rodríguez, J.C. Complementarity in phenolic compounds and the antioxidant activities of Phaseolus coccineus L. and P. vulgaris L. Landraces Foods 2019, 8, 295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delgado-Salinas, A.; Bibler, R.; Lavin, M. Phylogeny of the genus Phaseolus (Leguminosae): A recent diversification in an ancient landscape. Syst. Bot. 2006, 31, 779–791. [Google Scholar] [CrossRef]
- Ye, T.T.; Yamaguchi, H. Phylogenetic relationship of wild and cultivated Vigna (subgenus Ceratotropis, fabaceae) from Myanmar based on sequence variations in non-coding regions of trnT-F. Breed Sci. 2007, 57, 271–280. [Google Scholar]
- Alcázar-Valle, M.; Lugo-Cervantes, E.; Mojica, L.; Morales-Hernández, N.; Reyes-Ramírez, H.; Enríquez-Vara, J.N.; García-Morales, S. Bioactive compounds, antioxidant activity, and antinutritional content of legumes: A comparison between four Phaseolus species. Molecules 2020, 25, 3528. [Google Scholar]
- Quintana-Blanco, W.A.; Pinzón-Sandoval, E.H.; Torres, D.F. Evaluación del crecimiento de fríjol (Phaseolus vulgaris L.) cv Ica Cerinza, bajo estrés salino. Rev. UDCA Actual Divulg Científica 2016, 19, 87–95. [Google Scholar]
- Puertas-Mejía, M.A.; Mosquera-Mosquera, N.; Rojano, B. Estudio de la capacidad antioxidante in vitro de Phaseolus vulgaris L.(frijol) mediante extracción asistida por microondas. Rev. Cuba Plantas Med. 2016, 21, 42–50. [Google Scholar]
- Alcázar-Valle, M.; García-Morales, S.; Mojica, L.; Morales-Hernández, N.; Sánchez-Osorio, E.; Flores-López, L.; Enríquez-Vara, J.N.; Lugo-Cervantes, E. Nutritional, antinutritional compounds and nutraceutical significance of native bean species (Phaseolus spp.) of mexican cultivars. Agriculture 2021, 11, 1031. [Google Scholar] [CrossRef]
- Murthy, H.N.; Paek, K.-Y. Bioactive Compounds in Underutilized Vegetables and Legumes; Reference Series in Phytochemistry Series Editors. 2021, 681p. Available online: http://www.springer.com/series/13872 (accessed on 4 June 2023).
- Mina-Vargas, A.M.; McKeown, P.C.; Flanagan, N.S.; Debouck, D.G.; Kilian, A.; Hodkinson, T.R.; Spillane, C. Origin of year-long bean (Phaseolus dumosus Macfady, Fabaceae) from reticulated hybridization events between multiple Phaseolus species. Ann. Bot. 2016, 118, 957–969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marsh, L.E.; Davis, D.W. Influence of high temperature on the performance of some Phaseolus species at different developmental stages. Euphytica 1985, 34, 431–439. [Google Scholar] [CrossRef]
- Rodríguez, R.; Campa, A.; Suárez, B.; Ferreira, J.J. Phenolic content and antioxidant activity in seeds of common bean (Phaseolus vulgaris L.). Foods 2021, 10, 864. [Google Scholar] [CrossRef] [PubMed]
- Kan, L.; Nie, S.; Hu, J.; Liu, Z.; Xie, M. Antioxidant activities and anthocyanins composition of seed coats from twenty-six kidney bean cultivars. J. Funct. Foods 2016, 26, 622–631. [Google Scholar]
- Gálvez-Ranilla, L.; Genovese, M.I.; Lajolo, F.M. Polyphenols and antioxidant capacity of seed coat and cotyledon from Brazilian and Peruvian bean cultivars (Phaseolus vulgaris L.). J. Agric. Food Chem. 2007, 55, 90–98. [Google Scholar] [CrossRef]
- Aquino-Bolaños, E.N.; Garzón-García, A.K.; Alba-Jiménez, J.E.; Chávez-Servia, J.L.; Vera-Guzmán, A.M.; Carrillo-Rodríguez, J.C.; Santos-Basurto, M.A. Physicochemical characterization and functional potential of Phaseolus vulgaris l. and Phaseolus coccineus L. landrace green beans. Agronomy 2021, 11, 803. [Google Scholar] [CrossRef]
- de Mejía, E.G.; Guzmán-Maldonado, S.H.; Acosta-Gallegos, J.A.; Reynoso-Camacho, R.; Ramírez-Rodríguez, E.; Pons-Hernández, J.L.; González-Chavira, M.M.; Castellanos, J.Z.; Kelly, J.D. Effect of cultivar and growing location on the trypsin inhibitors, tannins, and lectins of common beans (Phaseolus vulgaris L.) Grown in the semiarid highlands of Mexico. J. Agric. Food Chem. 2003, 51, 5962–5966. [Google Scholar] [CrossRef]
- Teniente-Martínez, G.; Valadez-Vega, M.C.; González Cruz, L.; Cariño-Cortés, R.; Juárez-Gois, J.M.S.; Bernardino-Nicanor, A. Actividad antioxidante y contenido de compuestos fenólicos de péptidos de dos variedades de frijol ayocote (Phaseolus coccineus). Investig. Desarro Cienc Tecnol. Aliment. 2019, 4, 498–502. [Google Scholar]
- Ganesan, K.; Xu, B. Polyphenol-rich dry common beans (Phaseolus vulgaris L.) and their health benefits. Int. J. Mol. Sci. 2017, 18, 2331. [Google Scholar] [CrossRef] [Green Version]
- Chávez-Mendoza, C.; Sánchez, E. Bioactive compounds from mexican varieties of the common bean (Phaseolus vulgaris): Implications for health. Molecules 2017, 22, 1360. [Google Scholar]
- Huber, K. Phenolic Acid, Flavonoids and antioxidant activity of common brown beans (Phaseolus vulgaris L.) before and after cooking. J. Nutr. Food Sci. 2016, 6, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Lin, L.Z.; Harnly, J.M.; Pastor-Corrales, M.S.; Luthria, D.L. The polyphenolic profiles of common bean (Phaseolus vulgaris L.). Food Chem. 2008, 107, 399–410. [Google Scholar] [CrossRef] [Green Version]
- Suárez, J.C.; Urban, M.O.; Contreras, A.T.; Noriega, J.E.; Deva, C.; Beebe, S.E.; Polanía, J.A.; Casanoves, F.; Rao, I.M. Water use, leaf cooling and carbon assimilation efficiency of heat resistant common beans evaluated in Western Amazonia. Front. Plant Sci. 2021, 12, 644010. [Google Scholar] [CrossRef]
- Suárez, J.C.; Polanía, J.A.; Contreras, A.T.; Rodríguez, L.; Beebe, S.; Rao, I. Agronomical, phenological and physiological performance of common bean lines in the Amazon region of Colombia. Theor. Exp. Plant Physiol. 2018, 30, 303–320. [Google Scholar]
- Rosas-Jiménez, C.; Tercan, E.; Horstick, O.; Igboegwu, E.; Dambach, P.; Louis, V.R.; Winkler, V.; Deckert, A. Prevalence of anemia among Indigenous children in Latin America: A systematic review. Rev. Saude Publica. 2022, 56, 99. [Google Scholar] [CrossRef] [PubMed]
- del Carmen Segoviano-Lorenzo, M.; Trigo-Esteban, E.; Gyorkos, T.W.; St-Denis, K.; Guzmán FMDe Casapía-Morales, M. Prevalence of malnutrition, anemia, and soil-transmitted helminthiasis in preschool-age children living in peri-urban populations in the Peruvian Amazon. Cad Saude Publica. 2022, 38, e00248221. [Google Scholar] [CrossRef] [PubMed]
- Quizhpe, E.; San Sebastián, M.; Hurtig, A.K.; Llamas, A. Prevalencia de anemia en escolares de la zona Amazónica de Ecuador. Rev. Panam Salud Publica/Pan Am. J. Public Health 2003, 13, 355–361. [Google Scholar] [CrossRef] [Green Version]
- Suárez, J.C.; Contreras, A.T.; Anzola, J.A.; Vanegas, J.I.; Rao, I.M. Physiological characteristics of cultivated tepary bean (Phaseolus acutifolius a. gray) and its wild relatives grown at high temperature and acid soil stress conditions in the amazon region of colombia. Plants 2022, 11, 116. [Google Scholar] [CrossRef] [PubMed]
- Suárez, J.C.; Urban, M.O.; Contreras, A.T.; Grajales, M.; Cajiao, C.; Beebe, S.E.; Rao, I.M. Adaptation of interspecific Mesoamerican common bean lines to acid soils and high temperature in the Amazon region of Colombia. Plants 2021, 10, 2412. [Google Scholar] [CrossRef]
- Harkat, H.; Bousba, R.; Benincasa, C.; Atrouz, K.; Gültekin-Özgüven, M.; Altuntaş, Ü.; Demircan, E.; Zahran, H.A.; Özçelik, B. Assessment of biochemical composition and antioxidant properties of Algerian date palm (Phoenix dactylifera L.) seed oil. Plants 2022, 11, 381. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A., Jr. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar] [CrossRef]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Butler, L.G.; Price, M.L.; Brotherton, J.E. Vanillin assay for proanthocyanidins (condensed tannins): Modification of the solvent for estimation of the degree of polymerization. J. Agric. Food Chem. 1982, 30, 1087–1089. [Google Scholar] [CrossRef]
- Giusti, M.M.; Wrolstad, R.E. Characterization and measurement of anthocyanins by UV-visible spectroscopy. Curr. Protoc. Food Anal. Chem. 2001, 1, F1.2.1–F1.2.13. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophyls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzimol. 1987, 148, 350–382. [Google Scholar]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT Food Sci. Technol. Acad. Press 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Ramón, V.; Hernández, H.E.; Polania, P.; Suárez, J.C. Spatial distribution of cocoa quality: Relationship between physicochemical, functional and sensory attributes of clones from Southern Colombia. Agronomy 2023, 13, 15. [Google Scholar] [CrossRef]
- R Development Core Team. R Version 4.2.0 (2022-04-22)—“Vigorous Calisthenics” Copyright (C) 2024 The R Foundation for Statistical Computing Platform: x86_64-apple-darwin17.0 (64-bit). 2023. Available online: https://www.r-project.org/ (accessed on 4 June 2023).
- Brilhante, M.; Varela, E.; Essoh, A.P.; Fortes, A.; Duarte, M.C.; Monteiro, F.; Ferreira, V.; Correia, A.M.; Duarte, M.P.; Romeiras, M.M. Tackling food insecurity in cabo verde islands: The nutritional, agricultural and environmental values of the legume species. Foods 2021, 10, 206. [Google Scholar] [CrossRef]
- Yang, Q.Q.; Farha, A.K.; Cheng, L.; Kim, G.; Zhang, T.; Corke, H. Phenolic content and in vitro antioxidant activity in common beans (Phaseolus vulgaris L.) are not directly related to anti-proliferative activity. Food Biosci. 2020, 36, 100662. [Google Scholar] [CrossRef]
- Perez-Hernandez, L.M.; Hernández-Álvarez, A.J.; Morgan, M.; Boesch, C.; Orfila, C. Polyphenol bioaccessibility and anti-inflammatory activity of Mexican common beans (Phaseolus vulgaris L.) with diverse seed colour. CYTA J. Food 2021, 19, 682–690. [Google Scholar] [CrossRef]
- Onyilagha, J.C.; Islam, S. Flavonoids and other polyphenols of the cultivated species of the genus Phaseolus organic substrate view project Bio-active compounds & Physiological Functions of herbs and vegetables View project. Int. J. Agric. Biol. 2009, 11, 231–234. [Google Scholar]
- Murube, E.; Beleggia, R.; Pacetti, D.; Nartea, A.; Frascarelli, G.; Lanzavecchia, G.; Bellucci, E.; Nanni, L.; Gioia, T.; Marciello, U.; et al. Characterization of nutritional quality traits of a common bean germplasm collection. Foods 2021, 10, 1572. [Google Scholar] [CrossRef]
- Blair, M.W.; González, L.F.; Kimani, P.M.; Butare, L. Genetic diversity, inter-gene pool introgression and nutritional quality of common beans (Phaseolus vulgaris L.) from Central Africa. Theor. Appl. Genet. 2010, 121, 237–248. [Google Scholar] [CrossRef] [Green Version]
- Carbas, B.; Machado, N.; Oppolzer, D.; Ferreira, L.; Queiroz, M.; Brites, C.; Rosa, E.A.; Barros, A.I. Nutrients, antinutrients, phenolic composition, and antioxidant activity of common bean cultivars and their potential for food applications. Antioxidants 2020, 9, 186. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.Q.; Gan, R.Y.; Ge, Y.Y.; Zhang, D.; Corke, H. Polyphenols in Common Beans (Phaseolus vulgaris L.): Chemistry, Analysis, and Factors Affecting Composition. In Comprehensive Reviews in Food Science and Food Safety; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2018; Volume 17, pp. 1518–1539. [Google Scholar]
- Aknod, A.S.M.G.M.; Khandaker, L.; Berthold, J.; Gates, L.; Peters, K.; Delong, H.; Hossain, K. Anthocyanin, total polyphenols and antioxidant activity of common bean. Am. J. Food Technol. 2011, 6, 385–394. [Google Scholar]
- Ombra, M.N.; D’Acierno, A.; Nazzaro, F.; Riccardi, R.; Spigno, P.; Zaccardelli, M.; Pane, C.; Maione, M.; Fratianni, F. Phenolic Composition and Antioxidant and Antiproliferative Activities of the Extracts of Twelve Common Bean (Phaseolus vulgaris L.) Endemic Ecotypes of Southern Italy before and after Cooking. Oxid Med. Cell Longev. 2016, 2016, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Orak, H.H.; Karamać, M.; Orak, A.; Amarowicz, R. Antioxidant potential and phenolic compounds of some widely consumed Turkish white bean (Phaseolus vulgaris L.) varieties. Pol. J. Food Nutr. Sci. 2016, 66, 253–260. [Google Scholar] [CrossRef]
- Iniesta-González, J.J.; Ibarra-Pérez, F.J.; Gallegos-Infante, J.A.; Rocha-Guzmán, N.E.; González-Laredo, R.F. Factores antinutricios y actividad antioxidante en variedades mejoradas de frijol común (Phaseolus vulgaris). Agrociencia 2005, 39, 603–610. [Google Scholar]
- Abbas, M.; Saeed, F.; Anjum, F.M.; Afzaal, M.; Tufail, T.; Bashir, M.S.; Ishtiaq, A.; Hussain, S.; Suleria, H.A.R. Natural polyphenols: An overview. Int. J. Food Prop. Taylor Fr. 2017, 20, 1689–1699. [Google Scholar] [CrossRef] [Green Version]
- Sivaci, A.; Duman, S. Evaluation of seasonal antioxidant activity and total phenolic compounds in stems and leaves of some almond (Prunus amygdalus L.) varieties. Biol. Res. 2014, 47, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ávalos, A.; Elena, G. Metabolismo secundario de plantas. Reduca Biol. Ser. Fisiol Veg. 2009, 2, 119–145. [Google Scholar]
- Espinosa-Alonso, L.G.; Lygin, A.; Widholm, J.M.; Valverde, M.E.; Paredes-Lopez, O. Polyphenols in wild and weedy Mexican common beans (Phaseolus vulgaris L.). J. Agric. Food Chem. 2006, 54, 4436–4444. [Google Scholar] [CrossRef]
- Chávez-Mendoza, C.; Hernández-Figueroa, K.I.; Sánchez, E. Antioxidant capacity and phytonutrient content in the seed coat and cotyledon of common beans (Phaseolus vulgaris L.) from various regions in Mexico. Antioxidants 2019, 8, 5. [Google Scholar]
- Quiroz-Sodi, M.; Mendoza-Díaz, S.; Hernández-Sandoval, L.; Carrillo-Angeles, I. Characterization of the secondary metabolites in the seeds of nine native bean varieties (Phaseolus vulgaris and P. coccineus) from Querétaro, Mexico. Bot. Sci. 2018, 96, 650–661. [Google Scholar] [CrossRef] [Green Version]
- Malbos, D.; Buxeraud, J.; Faure, S.; Desport, J.C.; Vitamin, A. Actualites Pharmaceutiques; Elsevier Masson: Ile de France, France, 2021; Volume 60, pp. 23–26. [Google Scholar]
- Llerena, W.; Samaniego, I.; Angós, I.; Brito, B.; Ortiz, B.; Carrillo, W. Biocompounds content prediction in ecuadorian fruits using a mathematical model. Foods 2019, 8, 284. [Google Scholar] [CrossRef] [Green Version]
- Fratianni, A.; Irano, M.; Panfili, G.; Acquistucci, R. Estimation of color of durum wheat. Comparison of WSB, HPLC, and reflectance colorimeter measurements. J. Agric. Food Chem. 2005, 53, 2373–2378. [Google Scholar]
- Young, A.J.; Lowe, G.M. Antioxidant and prooxidant properties of carotenoids. Arch. Biochem. Biophys. 2001, 385, 20–27. [Google Scholar] [CrossRef]
- Ruiz, E.; Varela-Moreiras, G. Adequacy of the dietary intake of total and added sugars in the Spanish diet to the recommendations: ANIBES study. Nutr. Hosp. 2017, 34, 45–52. [Google Scholar]
- Plaza-Díaz, J.; Martínez Augustín, O.; Gil Hernández, Á. Los alimentos como fuente de mono y disacáridos: Aspectos bioquímicos y metabólicos. Nutr. Hosp. 2013, 28, 5–16. [Google Scholar] [PubMed]
- Jacinto-Hernández, C.; Coria-Peña, M.; Contreras-Santos, G.; Martínez-López, L.; Zapata-Martelo, E.; del Ayala-Carrillo, M.R. Azúcares totales y proteína en frijol nativo de la región Triqui Alta, Oaxaca. Rev. Mex. Cienc. Agrícolas 2019, 10, 1667–1674. [Google Scholar]
- Lee, A.A.; Owyang, C. Sugars, sweet taste receptors, and brain responses. In Molecular Nutrition Carbohydrates; Academic Press: Cambridge, MA, USA, 2019; pp. 265–283. [Google Scholar]
- Ponder, A.; Hallmann, E. The nutritional value and vitamin C content of different raspberry cultivars from organic and conventional production. J. Food Compos. Anal. 2020, 87, 103429. [Google Scholar] [CrossRef]
- Hallmann, E. The influence of organic and conventional cultivation systems on the nutritional value and content of bioactive compounds in selected tomato types. J. Sci. Food Agric. 2012, 92, 2840–2848. [Google Scholar] [CrossRef] [PubMed]
- Kapoulas, N.; Ilic, Z.S.; Durovka, M.; Trajkovic, R.; Milenkovic, L. Effect of organic and conventional production practices on nutritional value and antioxidant activity of tomatoes. Afr. J. Biotechnol. 2011, 10, 15938–15945. [Google Scholar] [CrossRef]
- Dladla, S.S.; Workneh, T.S. Evaluation of the effects of different packaging materials on the quality attributes of the tomato fruit. Appl. Sci. 2023, 13, 2100. [Google Scholar] [CrossRef]
- Karavidas, I.; Ntatsi, G.; Marka, S.; Ntanasi, T.; Consentino, B.B.; Sabatino, L.; Iannetta, P.P.M.; Savvas, D. Fresh pod yield, physical and nutritional quality attributes of common bean as influenced by conventional or organic farming practices. Plants 2023, 12, 32. [Google Scholar] [CrossRef]
- Bhuyan, D.J.; Basu, A. Bioactive Compounds in Agricultural and Food Production Waste. In Utilisation of Bioactive Compounds from Agricultural and Food Waste; CRC Press: Boca Raton, FL, USA, 2017; pp. 1–26. [Google Scholar]
- Bento, J.A.C.; Gomes, M.J.C.; Bassinello, P.Z.; Martino, H.S.D.; de Souza Neto, M.A.; Oomah, B.D. Benefits of carioca beans (Phaseolus vulgaris): Molecular mechanisms to human health and nutrition. In Molecular Mechanisms of Functional Food; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2022; pp. 112–141. [Google Scholar]
- Didinger, C.; Foster, M.T.; Bunning, M.; Thompson, H.J. Nutrition and Human Health Benefits of Dry Beans and Other Pulses. In Dry Beans and Pulses; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2022; pp. 481–504. [Google Scholar]
- Rodríguez, L.; Mendez, D.; Montecino, H.; Carrasco, B.; Arevalo, B.; Palomo, I.; Fuentes, E. Role of Phaseolus vulgaris L. in the prevention of cardiovascular diseases-cardioprotective potential of bioactive compounds. Plants 2022, 11, 186. [Google Scholar] [CrossRef]
- Vasilopoulou, E.; Georga, K.; Joergensen, M.; Naska, A.; Trichopoulou, A. The antioxidant properties of Greek foods and the flavonoid content of the Mediterranean menu. Curr. Med. Chem. Immunol. Endocr. Metab. Agents. 2005, 5, 33–45. [Google Scholar] [CrossRef]
- Saura-Calixto, F.; Goni, I. Definition of the mediterranean diet based on bioactive compounds. Crit. Rev. Food Sci. Nutr. 2009, 49, 145–152. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | HBF | MBFC | MACP | MALAP | ALBF | Overall Average | p-Value |
|---|---|---|---|---|---|---|---|
| Total sugars | 126.05 ± 13.81 b | 147.61 ± 10.49 b | 137.29 ± 9.23 b | 146.31 ± 10.08 b | 505.13 ± 63.99 a | 159.93 ± 9.43 | <0.0001 |
| Reducing sugars | 0.3 ± 0.02 a | 0.28 ± 0.01 a | 0.3 ± 0.02 a | 0.32 ± 0.01 a | 0.24 ± 0.02 b | 0.3 ± 0.01 | 0.0002 |
| Total phenolic content | 15.77 ± 0.86 a | 10.05 ± 0.25 d | 9.35 ± 0.3 c | 7.06 ± 0.22 b | 5.14 ± 0.53 e | 9.26 ± 0.28 | <0.0001 |
| Total flavonoid content | 15.15 ± 0.53 a | 12.85 ± 0.34 c | 12.61 ± 0.43 b | 8.57 ± 0.31 b | 6.83 ± 0.25 d | 11.34 ± 0.3 | <0.0001 |
| DPPH | 3068.68 ± 104.56 a | 2501.95 ± 37.61 c | 1126.2 ± 32.84 d | 2250.97 ± 80.3 b | 2167.89 ± 78.07 c | 2282.76 ± 56.75 | <0.0001 |
| FRAP | 7.19 ± 0.35 a | 5.15 ± 0.17 c | 6.93 ± 0.37 a | 3.74 ± 0.24 b | 1.85 ± 0.42 d | 4.94 ± 0.18 | <0.0001 |
| ABTS | 1791.85 ± 133.61 a | 945.56 ± 38.26 c | 827.89 ± 36.2 c | 546.25 ± 29.49 b | 329.23 ± 38.2 e | 846.98 ± 40.24 | <0.0001 |
| Carotenoids | 5.49 ± 0.96 a | 2.85 ± 0.22 c | 2.04 ± 0.2 c | 1.53 ± 0.17 b | 1.5 ± 0.23 c | 2.49 ± 0.17 | <0.0001 |
| Condensed tannins | 1.1 ± 0.15 c | 1.48 ± 0.08 b | 1.54 ± 0.1 c | 3.49 ± 0.27 c | 5.59 ± 1.61 a | 2.29 ± 0.16 | <0.0001 |
| Monomeric anthocyanins | 0.01 ± 0 | 8.8 ± 0.56 | 10.34 ± 0.79 | 0.01 ± 0 | 10.58 ± 1.02 | 8.82 ± 0.28 | |
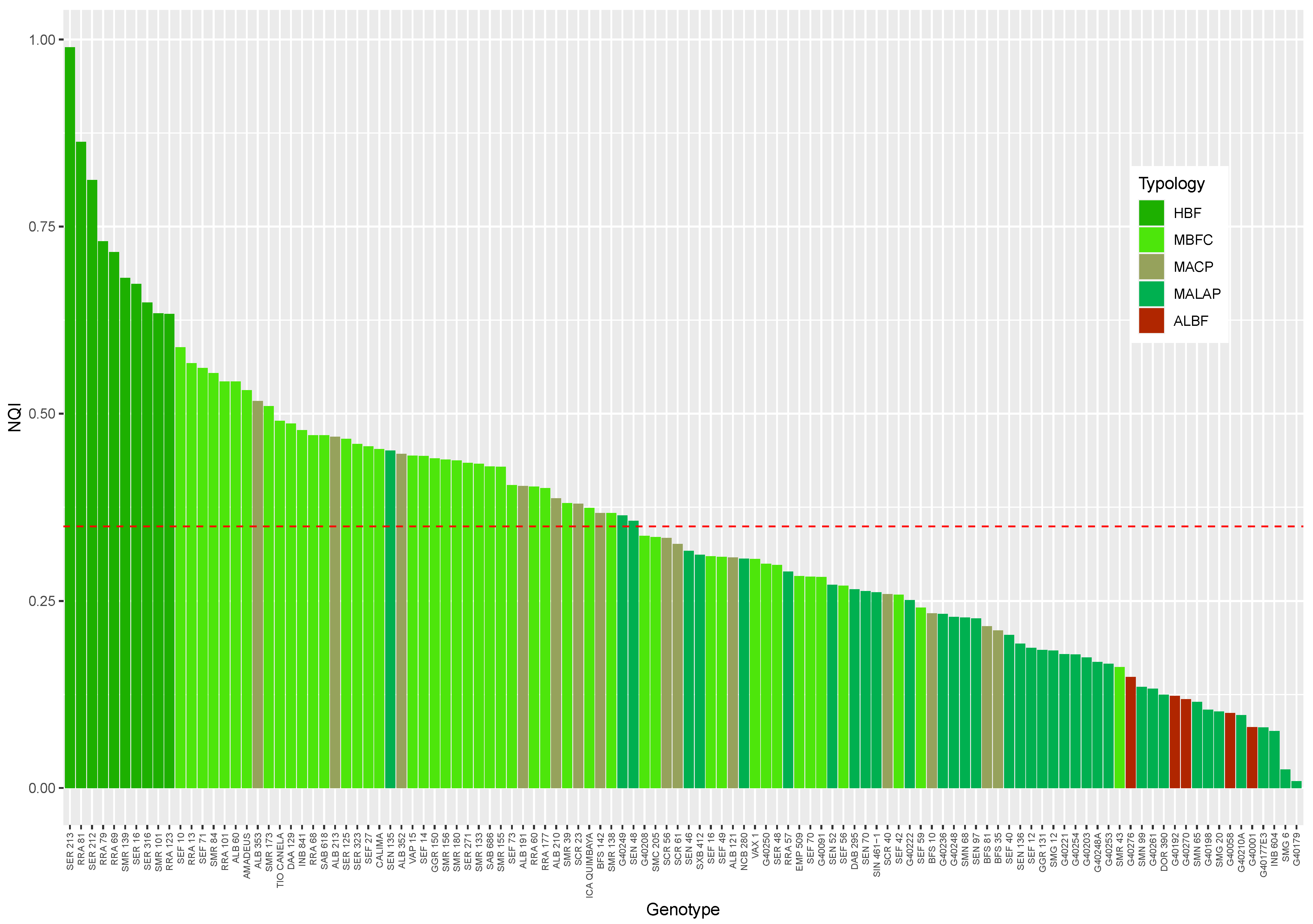
| NQI | 0.74 ± 0.04 a | 0.41 ± 0.01 d | 0.35 ± 0.03 c | 0.2 ± 0.02 b | 0.11 ± 0.01 c | 0.35 ± 0.02 | <0.0001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suárez, J.C.; Polanía-Hincapié, P.A.; Saldarriaga, S.; Ramón-Triana, V.Y.; Urban, M.O.; Beebe, S.E.; Rao, I.M. Bioactive Compounds and Antioxidant Activity in Seeds of Bred Lines of Common Bean Developed from Interspecific Crosses. Foods 2023, 12, 2849. https://doi.org/10.3390/foods12152849
Suárez JC, Polanía-Hincapié PA, Saldarriaga S, Ramón-Triana VY, Urban MO, Beebe SE, Rao IM. Bioactive Compounds and Antioxidant Activity in Seeds of Bred Lines of Common Bean Developed from Interspecific Crosses. Foods. 2023; 12(15):2849. https://doi.org/10.3390/foods12152849
Chicago/Turabian StyleSuárez, Juan Carlos, Paola Andrea Polanía-Hincapié, Sebastian Saldarriaga, Vivian Yorlady Ramón-Triana, Milan O. Urban, Stephen E. Beebe, and Idupulapati M. Rao. 2023. "Bioactive Compounds and Antioxidant Activity in Seeds of Bred Lines of Common Bean Developed from Interspecific Crosses" Foods 12, no. 15: 2849. https://doi.org/10.3390/foods12152849