
Sweeteners’ Influence on In Vitro α-Glucosidase Inhibitory Activity, Cytotoxicity, Stability and In Vivo Bioavailability of the Anthocyanins from Lingonberry Jams
 , , ,
, , ,
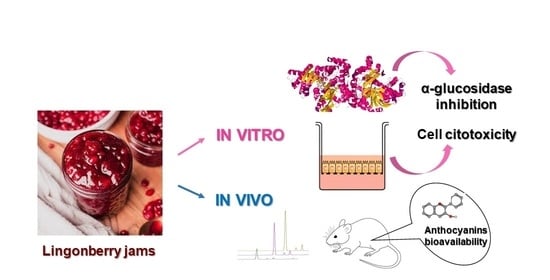
Abstract
:
1. Introduction
2. Results and Discussion
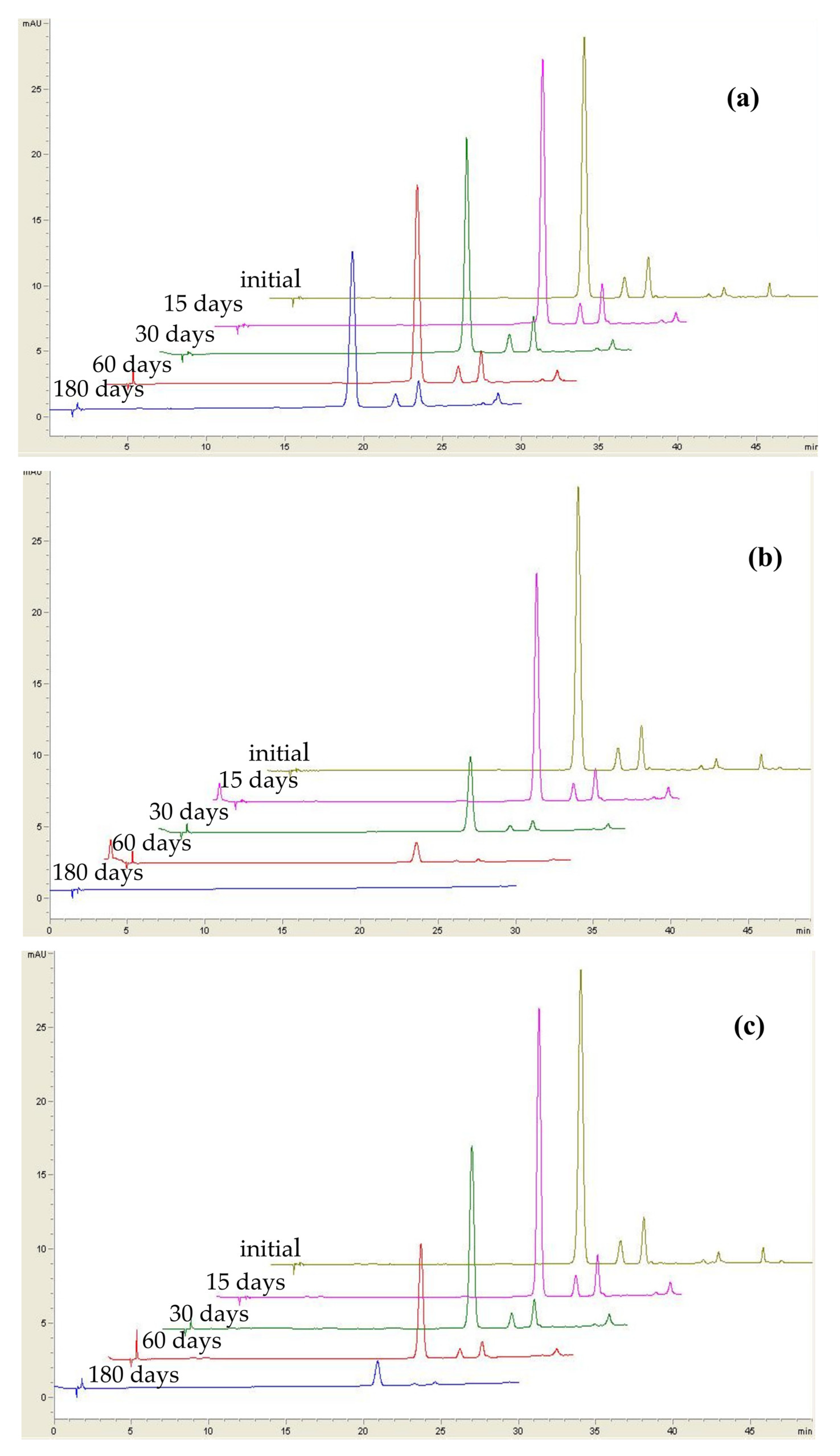
2.1. Effect of Sweeteners on the Stability of Anthocyanins from Lingonberry Jams during Storage
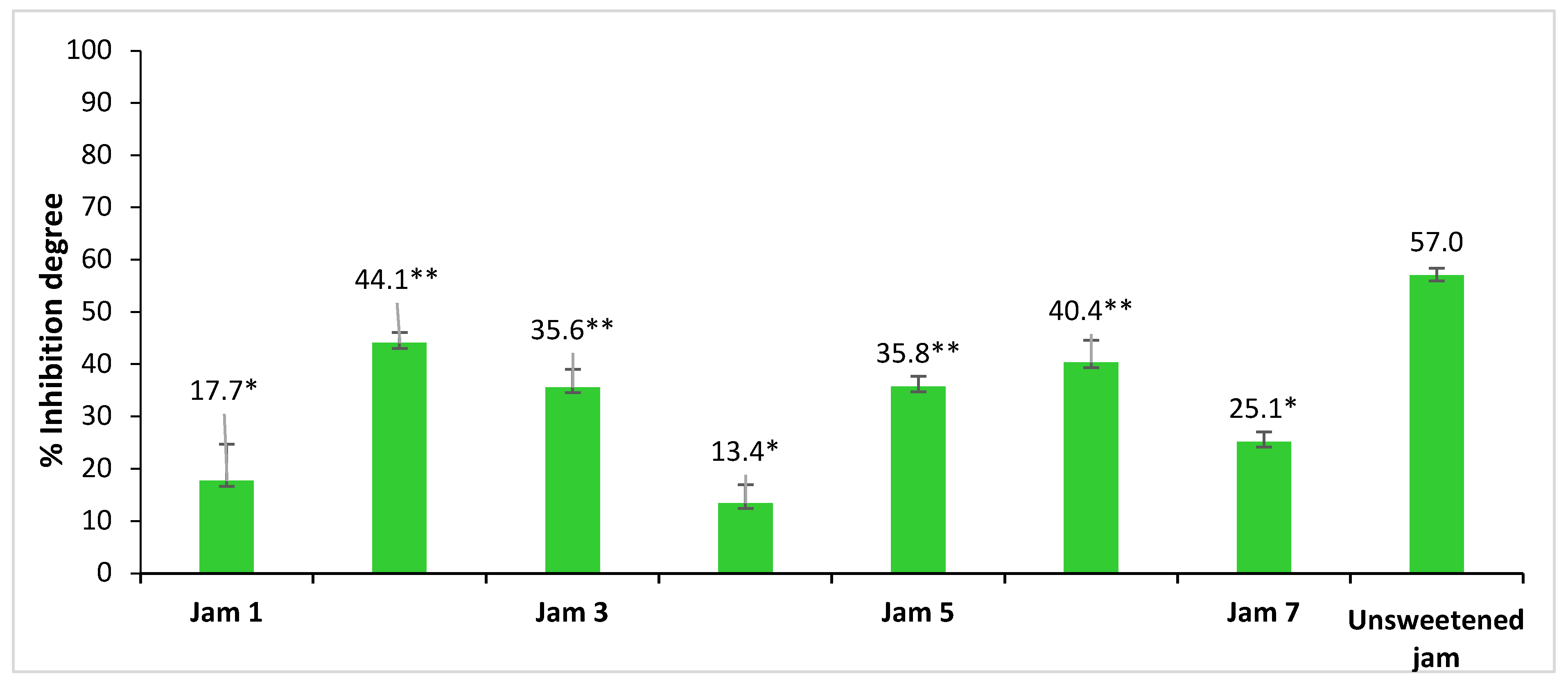
2.2. Inhibitory Activity of Lingonberry Jams against α-Glucosidase
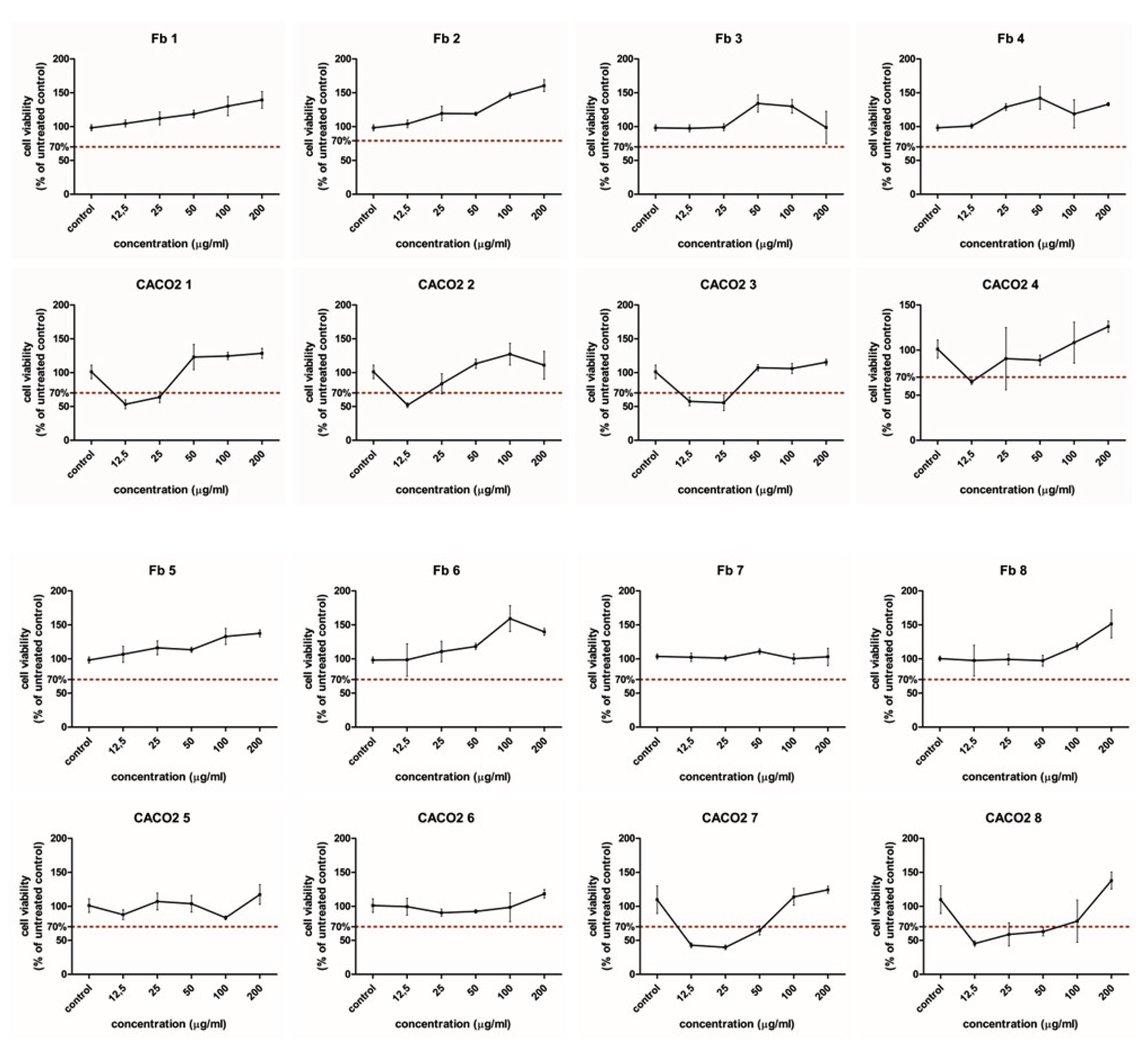
2.3. Cytotoxicity and Cell Viability in Normal and CaCo-2 Cancer Cells
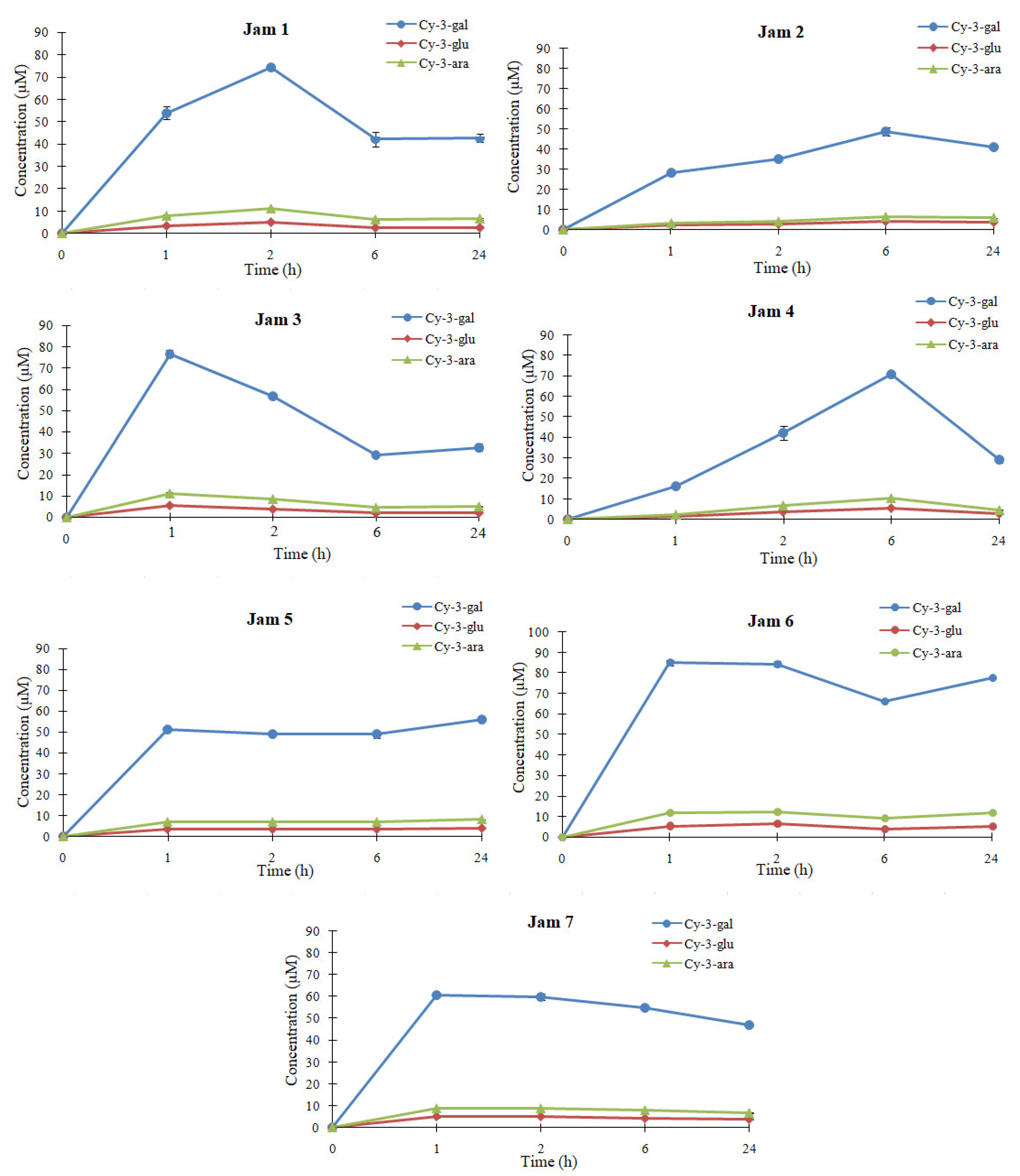
2.4. In Vivo Bioavailabiliy of Anthocyanins from Lingonberry Jams
3. Materials and Methods
3.1. Chemicals
3.2. Lingonberry Jams and Extracts Preparation
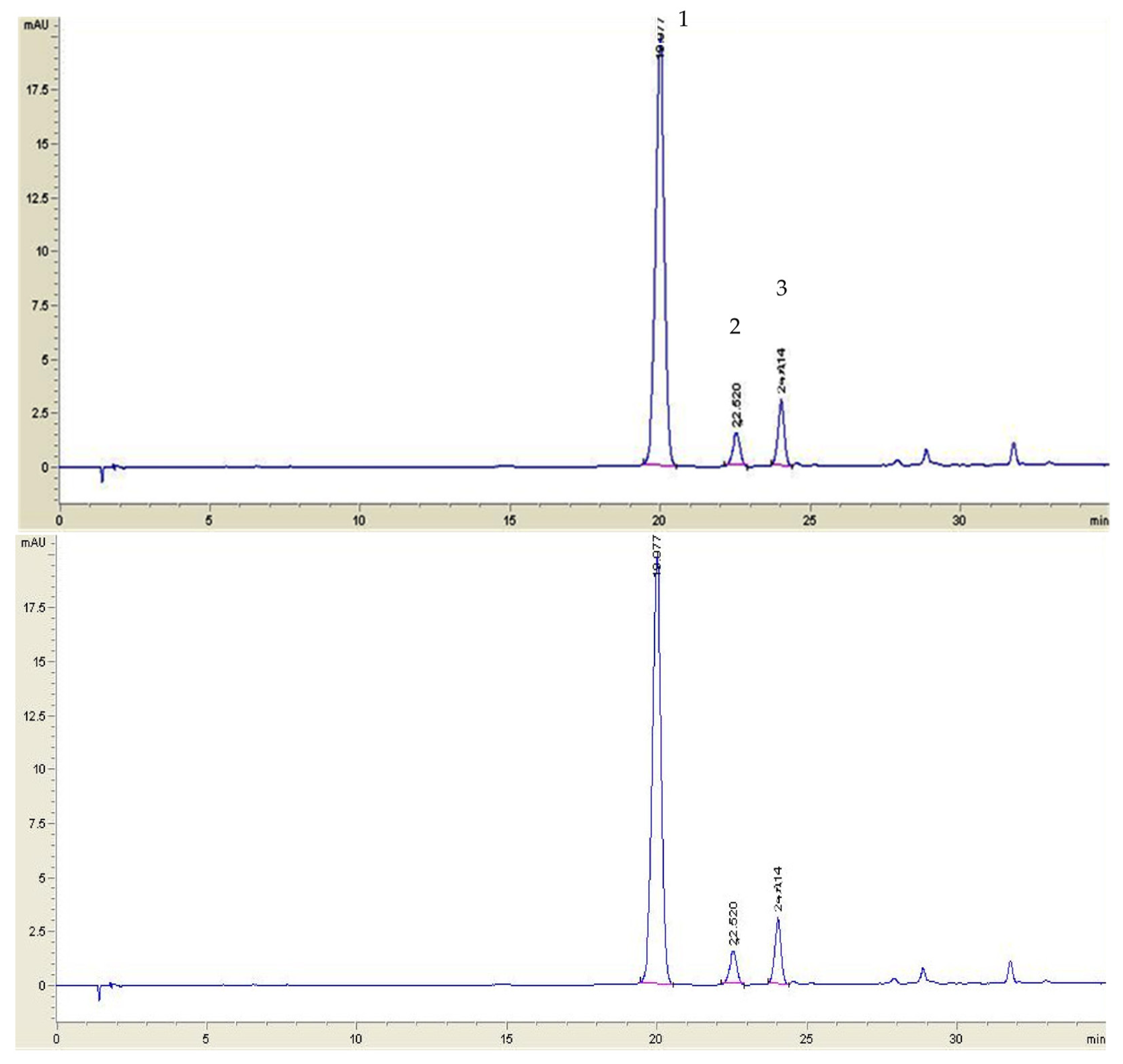
3.3. High-Performance Liquid Chromatography (HPLC) Analysis of Anthocyanins
3.3.1. Lingonberry Jams Extracts Analysis
3.3.2. HPLC Quantification of Anthocyanins in Plasma
3.4. α-Glucosidase Inhibition Assay
3.5. Cytotoxicity Assay
3.5.1. Cell Cultures
3.5.2. Cytotoxicity Assay
3.6. Animal Study Design and Plasma Analysis
3.7. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Han, P.; Yang, P.; Wang, H.; Fernandes, I.; Mateus, N.; Liu, Y. Digestion and absorption of red grape and wine anthocyanins through the gastrointestinal tract. Trends Food Sci. Technol. 2019, 83, 211–224. [Google Scholar] [CrossRef]
- Fernandes, I.; Faria, A.; Calhau, C.; de Freitas, V.; Mateus, N. Bioavailability of anthocyanins and derivatives. J. Funct. Foods 2014, 7, 54–66. [Google Scholar] [CrossRef]
- Cavalcante Braga, A.R.; Murador, D.C.; Mendes de Souza Mesquita, L.; de Rosso, V.V. Bioavailability of anthocyanins: Gaps in knowledge, challenges and future research. J. Food Compos. Anal. 2018, 68, 31–40. [Google Scholar] [CrossRef]
- Francis, F.J. Food colorants: Anthocyanins. Crit. Rev. Food Sci. Nutr. 1989, 28, 273–314. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.Z.; Shea, E.; Daneshtalab, M.; Weber, J.T. Chemical analysis of extracts from newfoundland berries and potential neuroprotective effects. Antioxidants 2016, 5, 36. [Google Scholar] [CrossRef] [Green Version]
- Crozier, A.; Jaganath, I.B.; Clifford, M.N. Dietary phenolics: Chemistry, bioavailability and effects on health. Nat. Prod. Rep. 2009, 26, 1001–1043. [Google Scholar] [CrossRef]
- Mortensen, A.; Sorensen, I.K.; Wilde, C.; Dragoni, S.; Mullerova, D.; Toussaint, O.; Zloch, Z.; Sgaragli, G.; Ovesna, J. Biological models for phytochemical research: From cell to human organism. Br. J. Nutr. 2008, 99, ES118–ES126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Sun, L.; Fang, Z.; Nisar, T.; Zou, L.; Li, D.; Guo, Y. Lyciumruthenicum Murray anthocyanins effectively inhibit α-glucosidase activity and alleviate insulin resistance. Food Biosci. 2021, 41, 100949. [Google Scholar] [CrossRef]
- Mohamed Sham Shihabudeen, H.; Hansi Priscilla, D.; Thirumurugan, K. Cinnamon extract inhibits α-glucosidase activity and dampens postprandial glucose excursion in diabetic rats. Nutr. Metab. 2011, 8, 46–56. [Google Scholar] [CrossRef] [Green Version]
- Reddivari, L.; Vanamala, J.; Chintharlapalli, S.; Safe, S.H.; Miller, J.C., Jr. Anthocyanin fraction from potato extracts is cytotoxic to prostate cancer cells through activation of caspase-dependent and caspase-independent pathways. Carcinogenesis 2007, 28, 2227–2235. [Google Scholar] [CrossRef] [Green Version]
- Ortega, N.; Reguant, J.; Romero, M.P.; Macia, A.; Motilva, M.J. Effect of fat content on the digestibility and bioaccessibility of cocoa polyphenol by an in vitro digestion model. J. Agric. Food Chem. 2009, 57, 5743–5749. [Google Scholar] [CrossRef] [PubMed]
- Bujor, O.-C.; Ginies, C.; Popa, V.I.; Dufour, C. Phenolic compounds and antioxidant activity of lingonberry (Vacciniumvitisidaea L.) leaf, stem and fruit at different harvest periods. Food Chem. 2018, 252, 356–365. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Wang, S.Y. Oxygen radical absorbing capacity of phenolics in blueberries, cranberries, chokeberries, and lingonberries. J. Agric. Food Chem. 2003, 51, 502–509. [Google Scholar] [CrossRef]
- Kostka, T.; Ostberg-Potthoff, J.J.; Stärke, J.; Guigas, C.; Matsugo, S.; Mirceski, V.; Stojanov, L.; KostadinovicVelickovska, S.; Winterhalter, P.; Esatbeyoglu, T. Bioactive phenolic compounds from lingonberry (Vaccinium vitis-idaea L.): Extraction, chemical characterization, fractionation and cellular antioxidant activity. Antioxidants 2022, 11, 467. [Google Scholar] [CrossRef] [PubMed]
- Kamiloglu, S.; Pasli, A.A.; Ozcelik, B.; Van Camp, J.; Capanoglu, E. Colour retention, anthocyanin stability and antioxidant capacity in black carrot (Daucus carota) jams and marmalades: Effect of processing, storage conditions and in vitro gastrointestinal digestion. J. Funct. Foods 2015, 13, 1–10. [Google Scholar] [CrossRef]
- Scrob, T.; Varodi, S.M.; Vintilă, G.A. The influence of formulation and storage conditions on titratable acidity, total soluble solids and sensorial profile of several lingonberry jams. Stud. Univ. Babes-Bolyai Chem. 2021, 66, 97–106. [Google Scholar] [CrossRef]
- Pittman, J.M.; Schalliol, L.A.; Ray, S.D. Insulin and other hypoglycemic drugs. In Side Effects of Drugs Annual, 1st ed.; Ray, S.D., Ed.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 435–446. [Google Scholar]
- Jan, A.; Habib, N.; Shinwari, Z.K.; Ali, M.; Ali, N. The anti-diabetic activities of natural sweetener plant Stevia: An updated review. SN Appl. Sci. 2021, 3, 517–522. [Google Scholar] [CrossRef]
- Adisakwattana, S.; Charoenlertkul, P.; Yibchokanun, S. α-Glucosidase inhibitory activity of cyanidin-3-galactoside and synergistic effect with acarbose. J. Enzyme Inhib. Med. Chem. 2009, 24, 65–69. [Google Scholar] [CrossRef]
- Lazzè, M.C.; Savio, M.; Pizzala, R.; Cazzalini, O.; Perucca, P.; Scovassi, A.I.; Stivala, L.A.; Bianchi, L. Anthocyanins induce cell cycle perturbations and apoptosis in different human cell lines. Carcinogenesis 2004, 25, 1427–1433. [Google Scholar] [CrossRef]
- Shi, N.; Chen, X.; Chen, T. Anthocyanins in Colorectal Cancer Prevention Review. Antioxidants 2021, 10, 1600. [Google Scholar] [CrossRef]
- Chen, T.; Shi, N.; Afzali, A. Chemopreventive Effects of Strawberry and Black Raspberry on Colorectal Cancer in Inflammatory Bowel Disease. Nutrients 2019, 11, 1261. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Ortiz, J.M.; Alguacil, L.F.; Salas, E.; Hermosín-Gutiérrez, I.; Gómez-Alonso, S.; González-Martín, C. Antiproliferative and cytotoxic effects of grape pomace and grape seed extracts on colorectal cancer cell lines. Food Sci. Nutr. 2019, 7, 2948–2957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- AT Bioquest, Inc. QuestGraph™IC50 Calculator. Available online: https://www.aatbio.com/tools/ic50-calculator (accessed on 14 May 2023).
- Ferguson, P.J.; Kurowska, E.M.; Freeman, D.J.; Chambers, A.F.; Koropatnick, J. In vivo inhibition of growth of human tumor lines by flavonoid fractions from cranberry extract. Nutr. Cancer 2006, 56, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Costa, J.R.; Amorim, M.; Vilas-Boas, A.; Tonon, R.V.; Cabral, L.M.C.; Pastranac, L.; Pintado, M. Impact of in vitro gastrointestinal digestion on the chemical composition, bioactive properties, and cytotoxicity of Vitis vinifera L. cv. Syrah grape pomace extract. Food Funct. 2019, 10, 1856–1859. [Google Scholar] [CrossRef]
- Rugina, D.; Scontxa, Z.; Leopold, L.; Pintea, A.; Bunea, A.; Socaciu, C. Antioxidant activities of chokeberry extracts and the cytotoxic action of their anthocyanin fraction on hela human cervical tumor cells. J. Med. Food 2012, 15, 700–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, C.; Giusti, M.M.; Malik, M.; Moyer, M.P.; Magnuson, B.A. Effects of commercial anthocyanin-rich on colonic cancer and non-tumorigenic colonic cell growth. J. Agric. Food Chem. 2004, 52, 6122–6128. [Google Scholar] [CrossRef]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carrière, F.; Boutrou, R.; Corredig, M.; Dupont, D.; et al. A standardised static in vitro digestion method suitable for food—An international consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scrob, T.; Hosu, A.; Cimpoiu, C. Sweeteners from different lingonberry jams influence on bioaccessibility of vitamin C, anthocyanins and antioxidant capacity under in vitro gastrointestinal digestion. Antioxidants 2022, 11, 442. [Google Scholar] [CrossRef]
- Talavera, S.; Felgines, C.; Texier, O.; Besson, C.; Lamaison, J.L.; Remesy, C. Anthocyanins are efficiently absorbed from the stomach in anesthetized rats. J. Nutr. 2003, 133, 4178–4182. [Google Scholar] [CrossRef] [Green Version]
- Kalt, W.; Blumberg, J.B.; McDonald, J.E.; Vinqvist-Tymchuk, M.R.; Fillmore, S.A.; Graf, B.A.; O’Leary, J.M.; Milbury, P.E. Identification of anthocyanins in the liver, eye, and brain of blueberry-fed pigs. J. Agric. Food Chem. 2008, 56, 705–712. [Google Scholar] [CrossRef]
- Walle, T.; Browning, A.M.; Steed, L.L.; Reed, S.G.; Walle, U.K. Flavonoid glucosides are hydrolyzed and thus activated in the oral cavity in humans. J. Nutr. 2005, 135, 48–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hidalgo, M.; Oruna-Concha, M.J.; Kolida, S.; Walton, G.E.; Kallithraka, S.; Spencer, J.P.E.; Gibson, G.R.; de Pascual-Teresa, S. Metabolism of Anthocyanins by Human Gut Microflora and Their Influence on Gut Bacterial Growth. J. Agric. Food Chem. 2012, 60, 3882–3890. [Google Scholar] [CrossRef] [PubMed]
- Ichiyanagi, T.; Shida, Y.; Rahman, M.M.; Hatano, Y.; Konishi, T. Bioavailability and tissue distribution of anthocyanins in bilberry (Vaccinium myrtillus L.) extract in rats. J. Agric. Food Chem. 2006, 54, 6578–6587. [Google Scholar] [CrossRef] [PubMed]
- Lehtonen, H.M.; Rantala, M.; Suomela, J.P.; Viitanen, M.; Kallio, H. Protective effects of bilberry (Vaccinium myrtillus L.) extract on restraint stress-induced liver damage in mice. J. Agric. Food Chem. 2008, 56, 7803–7808. [Google Scholar] [CrossRef]
- Agulló, V.; Domínguez-Perles, R.; García-Viguera, C. Sweetener influences plasma concentration of flavonoids in humans after an acute intake of a new (poly)phenol-rich beverage. Nutr. Metab. Cardiovasc. Dis. 2021, 31, 930–938. [Google Scholar] [CrossRef]
- Agulló, V.; García-Viguera, C.; Domínguez-Perles, R. The use of alternative sweeteners (sucralose and stevia) in healthy soft-drink beverages, enhances the bioavailability of polyphenols relative to the classical caloric sucrose. Food Chem. 2022, 370, 131051. [Google Scholar] [CrossRef]
- Filip, G.A.; Florea, A.; Olteanu, D.; Clichici, S.; David, L.; Moldovan, B.; Cenariu, M.; Scrobota, I.; Potara, M.; Baldea, I. Biosynthesis of silver nanoparticles using Sambucus nigra L. fruit extract for targeting cell death in oral dysplastic cells. Mater. Sci. Eng. C Mater. Biol. Appl. 2021, 123, 111974. [Google Scholar] [CrossRef]
- Talavera, S.; Felgines, C.; Texier, O.; Besson, C.; Mazur, A.; Lamaison, J.; Remesy, C. Bioavailability of a bilberry anthocyanin extract and its impact on plasma antioxidant capacity in rats. J. Sci. Food Agric. 2006, 86, 90–97. [Google Scholar] [CrossRef]
- Felgines, C.; Texier, O.; Besson, C.; Fraisse, D.; Lamaison, J.L.; Remesy, C. Blackberry anthocyanins are slightly bioavailable in rats. J. Nutr. 2002, 132, 1249–1253. [Google Scholar] [CrossRef] [Green Version]
- Seraglio, S.K.T.; Valese, A.C.; Daguerb, H.; Bergamo, G.; Azevedo, M.S.; Nehring, P.; Gonzaga, V.; Fett, R.; Costa, A.C.O. Effect of in vitro gastrointestinal digestion of phenolic compounds, minerals, and antioxidant capacity of Mimosa scabrella Bentham honeydew honeys. Food Res. Int. 2017, 99, 670–678. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Anthocyanin | 4 °C | 25 °C (Light Conditions) | 25 °C (Dark Conditions) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Day 15 | Day 30 | Day 60 | Day 180 | Day 15 | Day 30 | Day 60 | Day 180 | Day 15 | Day 30 | Day 60 | Day 180 | ||
| Jam 1 (white sugar) | Cyd-3-gal | 91.1 ± 3.2 | 88.7 ± 4.1 | 77.9 ± 2.2 | 61.9 ± 2.8 | 71.2 ± 3.4 | 26.2 ± 1.5 | 7.10 ± 0.41 | 0.10 ± 0.01 | 83.9 ± 4.4 | 61.2 ± 3.3 | 37.7 ± 1.1 | 8.01 ± 0.44 |
| Cyd-3-glu | 94.5 ± 2.9 | 89.1 ± 3.9 | 86.7 ± 3.1 | 59.3 ± 3.4 | 72.6 ± 4.2 | 25.9 ± 1.8 | 6.31 ± 0.22 | 0.51 ± 0.03 | 94.1 ± 5.8 | 64.4± | 33.8 ± 2.3 | 5.90 ± 0.21 | |
| Cyd-3-ara | 96.9 ± 4.1 | 88.6 ± 3.5 | 81.2 ± 2.8 | 63.1 ± 2.6 | 72.8 ± 2.5 | 19.9 ± 0.9 | 5.90 ± 0.19 | 0.32 ± 0.01 | 89.7 ± 4.7 | 59.1± | 32.5 ± 1.9 | 4.81 ± 0.14 | |
| Jam 2 (fructose) | Cyd-3-gal | 95.1 ± 3.3 | 94.2 ± 5.2 | 86.7 ± 2.6 | 67.6 ± 1.9 | 60.4 ± 2.9 | 26.3 ± 1.6 | 10.3 ± 0.8 | 0.11 ± 0.01 | 90.7 ± 6.1 | 54.0± | 26.2 ± 1.0 | 1.42 ± 0.08 |
| Cyd-3-glu | 99.2 ± 5.0 | 90.9 ± 4.6 | 87.6 ± 3.2 | 62.9 ± 2.2 | 60.4 ± 1.8 | 24.2 ± 1.0 | 10.8 ± 0.5 | n.d. | 97.2 ± 5.9 | 50.8± | 24.2 ± 1.5 | 0.66 ± 0.00 | |
| Cyd-3-ara | 95.2 ± 1.9 | 90.4 ± 3.9 | 82.5 ± 3.3 | 68.6 ± 3.4 | 55.5 ± 1.2 | 18.9 ± 1.0 | 10.7 ± 0.3 | 0.30 ± 0.01 | 96.7 ± 5.1 | 50.3± | 25.8 ± 1.1 | 0.72 ± 0.02 | |
| Jam 3 (erythritol) | Cyd-3-gal | 93.5 ± 2.1 | 80.9 ± 2.1 | 78.2 ± 2.9 | 64.5 ± 3.8 | 38.9 ± 3.3 | 14.7 ± 0.8 | 4.70 ± 0.14 | n.d. | 73.9 ± 4.0 | 61.9± | 38.2 ± 2.3 | n.d. |
| Cyd-3-glu | 96.3 ± 4.2 | 87.5 ± 1.7 | 83.3 ± 3.6 | 60.8 ± 2.8 | 46.6 ± 1.8 | 14.2 ± 0.7 | 5.83 ± 0.25 | n.d. | 74.4 ± 4.4 | 69.2± | 34.1 ± 2.0 | n.d. | |
| Cyd-3-ara | 92.8 ± 2.3 | 81.3 ± 3.0 | 79.9 ± 4.1 | 66.9 ± 1.9 | 35.6 ± 2.1 | 9.4 ± 0.62 | 4.54 ± 0.33 | n.d. | 74.9 ± 3.8 | 61.2± | 32.6 ± 1.9 | n.d. | |
| Jam 4 (brown sugar) | Cyd-3-gal | 91.3 ± 4.9 | 89.7 ± 2.8 | 88.3 ± 2.2 | 61.3 ± 2.4 | 42.2 ± 2.0 | 18.5 ± 1.0 | 8.01 ± 0.41 | 0.32 ± 0.02 | 68.5 ± 2.3 | 56.9± | 31.7 ± 2.4 | 4.02 ± 0.11 |
| Cyd-3-glu | 96.7 ± 5.3 | 93.1 ± 4.0 | 83.6 ± 3.7 | 60.9 ± 3.3 | 38.7 ± 1.7 | 18.7 ± 1.3 | 8.83 ± 0.28 | n.d. | 59.9 ± 1.9 | 51.1± | 29.2 ± 1.8 | 1.11 ± 0.08 | |
| Cyd-3-ara | 85.7 ± 1.8 | 82.9 ± 3.5 | 80.4 ± 4.5 | 61.8 ± 2.9 | 30.9 ± 2.1 | 15.9 ± 1.7 | 6.71 ± 0.33 | n.d. | 62.2 ± 3.0 | 55.4± | 31.4 ± 2.0 | 2.23 ± 0.06 | |
| Jam 5 (coconut sugar) | Cyd-3-gal | 97.7 ± 3.9 | 67.9 ± 2.8 | 66.4 ± 2.9 | 20.5 ± 1.0 | 31.4 ± 1.6 | 7.80 ± 0.61 | 1.92 ± 0.09 | 0.10 ± 0.01 | 40.5 ± 1.5 | 16.4± | 5.21 ± 0.23 | 0.33 ± 0.00 |
| Cyd-3-glu | 75.6 ± 1.8 | 64.9 ± 1.9 | 51.6 ± 1.6 | 15.3 ± 1.1 | 27.1 ± 1.9 | 5.40 ± 0.31 | 7.72 ± 0.37 | n.d. | 29.8 ± 1.1 | 16.1± | 4.23 ± 0.15 | 0.52 ± 0.01 | |
| Cyd-3-ara | 99.1 ± 6.1 | 72.1 ± 2.4 | 71.3 ± 2.7 | 19.6 ± 1.6 | 28.8 ± 1.1 | 10.4 ± 0.82 | 2.52 ± 0.11 | n.d. | 40.4 ± 3.6 | 20.8± | 3.80 ± 0.12 | 0.43 ± 0.02 | |
| Jam 6 (stevia) | Cyd-3-gal | 98.1 ± 4.8 | 97.3 ± 5.3 | 90.4 ± 1.1 | 77.5 ± 2.8 | 46.1 ± 3.1 | 12.8 ± 0.64 | 2.72 ± 0.15 | n.d. | 82.9 ± 5.8 | 52.7± | 52.1 ± 3.6 | n.d. |
| Cyd-3-glu | 97.2 ± 3.3 | 98.2 ± 6.0 | 97.5 ± 5.6 | 51.6 ± 1.7 | 46.5 ± 3.0 | 12.1 ± 0.53 | 6.22 ± 0.39 | n.d. | 80.4 ± 4.7 | 54.6± | 38.6 ± 1.0 | n.d. | |
| Cyd-3-ara | 99.2 ± 6.0 | 93.6 ± 4.8 | 87.5 ± 4.0 | 55.8 ± 1.0 | 37.8 ± 2.8 | 9.60 ± 0.22 | 4.42 ± 0.30 | n.d. | 85.7 ± 3.0 | 51.9± | 42.6 ± 3.1 | n.d. | |
| Jam 7 (saccharine) | Cyd-3-gal | 93.1 ± 4.4 | 91.8 ± 2.7 | 83.1 ± 3.8 | 66.9 ± 2.3 | 31.8 ± 2.4 | 7.40 ± 0.41 | 1.72 ± 0.10 | n.d. | 76.1 ± 2.1 | 46.2± | 40.7 ± 2.7 | n.d. |
| Cyd-3-glu | 92.7 ± 2.2 | 91.3 ± 4.1 | 72.6 ± 2.7 | 62.8 ± 2.8 | 32.4 ± 2.2 | 6.81 ± 0.43 | 4.30 ± 0.23 | n.d. | 73.8 ± 0.2.2 | 41.5± | 25.8 ± 1.1 | n.d. | |
| Cyd-3-ara | 95.7 ± 3.7 | 94.2 ± 3.6 | 85.9 ± 2.2 | 68.5 ± 3.7 | 20.5 ± 1.1 | 5.31 ± 0.35 | 3.51 ± 0.09 | n.d. | 78.5 ± 3.4 | 44.4± | 33.1 ± 2.1 | n.d. | |
| Jam | Sweetener | IC50 (µg/mL) |
|---|---|---|
| 1 | White sugar | 35.51 |
| 2 | Fructose | 35.52 |
| 3 | Erythritol | 44.60 |
| 4 | Brown sugar | 202.0 |
| 5 | Coconut sugar | 230.9 |
| 6 | Stevia | 92.29 |
| 7 | Saccharine | 90.87 |
| 8 | Unsweetened | 117.3 |
| Anthocyanins | Bioavailability (%) | ||||||
|---|---|---|---|---|---|---|---|
| Jam 1 (White Sugar) | Jam 2 (Fructose) | Jam 3 (Erythritol) | Jam 4 (Brown Sugar) | Jam 5 (Coconut Sugar) | Jam 6 (Stevia) | Jam 7 (Saccharine) | |
| 1 h | |||||||
| Cy-3-gal | 15.2 ± 0.9 a | 8.10 ± 0.3 a,b | 28.3 ±1.0 a,b,c | 4.91 ± 0.3 a,b,c,d | 28.1 ± 0.2 a,b,d,e | 28.0 ± 0.7 a,b,f | 20.3 ± 0.4 a,b,c,d,e,f |
| Cy-3-glu | 12.4 ± 1.9 a | 8.21 ± 0.9 b | 26.1 ±0.6 a,b,c | 5.30 ± 0.1 c,d | 29.1 ± 1.5 a,b,d,e | 23.8 ± 0.1 a,b,c,d, | 22.6 ± 1.4 a,b,d,e |
| Cy-3-ara | 15.5 ± 0.3 a | 7.11 ± 0.2 a,b | 27.9 ±1.3 a,b,c | 5.11 ± 0.7 a,c,d | 28.5 ± 0.3 a,b,d,e | 29.1 ± 1.1 a,b,d,f | 20.6 ± 0.1 a,b,c,d,e,f |
| 2 h | |||||||
| Cy-3-gal | 20.9 ± 0.1 a | 10.0 ± 0.1 a,b | 20.9 ±0.1 b,c | 12.7 ± 1.2 a,c,d | 27.0 ± 0.7 a,b,c,d,e | 27.8 ± 0.7 a,b,c,d,f | 20.0 ± 0.6 b,d,e,f |
| Cy-3-glu | 18.0 ± 1.3 a | 10.2 ± 0.0 a,b | 17.8 ±1.4 b,c | 14.9 ± 0.5 b,d | 28.0 ± 1.4 a,b,c,d | 28.7 ± 1.2 a,b,c,d,e | 22.8 ± 1.4 b,d,e |
| Cy-3-ara | 21.6 ± 0.5 a | 9.01 ± 0.4 a,b | 21.7 ±0.5 b,c | 14.4 ± 0.6 a,b,c,d | 28.0 ± 1.4 a,b,c,d,e | 29.7 ± 0.4 a,b,c,d,f | 20.3 ± 0.6 b,d,e,f |
| 6 h | |||||||
| Cy-3-gal | 11.9 ± 1.0 a | 13.8 ± 0.8 b | 10.7 ±0.3 b,c | 21.4 ± 0.7 a,b,c,d | 26.9 ± 1.6 a,b,c,d,e | 21.8 ± 0.4 a,b,c,e,f | 18.3 ± 0.1 a,b,c,d,e,f |
| Cy-3-glu | 9.11 ± 1.7 a | 15.3 ± 0.2 a,b | 9.28 ±0.4 b,c | 23.0 ± 0.6 a,b,d | 27.3 ± 1.8 a,b,c,e | 16.9 ± 0.1 a,b,c,d,e,f | 20.3 ± 0.5 a,b,d,e,f |
| Cy-3-ara | 12.1 ± 0.1 a | 13.1 ± 0.1 a,b | 11.5 ±0.8 c | 22.3 ± 0.5 a,b,c,d | 28.0 ± 1.4 a,b,c,d,e | 22.4 ± 0.7 a,b,c,e,f | 18.5 ± 0.1 a,b,c,d,e,f |
| 24 h | |||||||
| Cy-3-gal | 12.1 ± 0.6 a | 11.6 ± 0.4 b | 12.1 ±0.8 b,c | 8.81 ± 0.6 a,b,c,d | 30.8 ± 0.9 a,b,c,d,e | 25.6 ± 0.3 a,b,c,d,e,f | 15.7 ± 0.1 a,b,d,e,f |
| Cy-3-glu | 9.80 ± 1.6 a | 14.6 ± 0.1 a,b | 9.71 ±0.3 b,c | 10.9 ± 0.2 b,d | 32.0 ± 0.6 a,b,c,d,e | 22.8 ± 0.3 a,b,c,d,e,f | 17.1 ± 0.7 a,b,d,e,f |
| Cy-3-ara | 12.9 ± 0.6 a | 12.0 ± 0.3 b | 12.8 ±0.9 c | 9.90 ± 0.7 d | 32.7 ± 0.4 a,b,c,d,e | 28.8 ± 0.4 a,b,c,d,e,f | 15.7 ± 0.1 a,b,c,d,e,f |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scrob, T.; Filip, G.A.; Baldea, I.; Varodi, S.M.; Cimpoiu, C. Sweeteners’ Influence on In Vitro α-Glucosidase Inhibitory Activity, Cytotoxicity, Stability and In Vivo Bioavailability of the Anthocyanins from Lingonberry Jams. Foods 2023, 12, 2569. https://doi.org/10.3390/foods12132569
Scrob T, Filip GA, Baldea I, Varodi SM, Cimpoiu C. Sweeteners’ Influence on In Vitro α-Glucosidase Inhibitory Activity, Cytotoxicity, Stability and In Vivo Bioavailability of the Anthocyanins from Lingonberry Jams. Foods. 2023; 12(13):2569. https://doi.org/10.3390/foods12132569
Chicago/Turabian StyleScrob, Teodora, Gabriela Adriana Filip, Ioana Baldea, Sânziana Maria Varodi, and Claudia Cimpoiu. 2023. "Sweeteners’ Influence on In Vitro α-Glucosidase Inhibitory Activity, Cytotoxicity, Stability and In Vivo Bioavailability of the Anthocyanins from Lingonberry Jams" Foods 12, no. 13: 2569. https://doi.org/10.3390/foods12132569