
Characterization of Lactic Acid Bacteria Strains Isolated from Algerian Honeybee and Honey and Exploration of Their Potential Probiotic and Functional Features for Human Use

Abstract
:1. Introduction
2. Materials and Methods
2.1. Honeybees and Honey Sampling
2.2. Isolation of Acid Tolerant LAB and Characterization of Fermentation Pattern
2.3. Molecular Identification and Phylogenetic Analysis
2.4. Assessment of Proteolytic, Lipolytic Activities and EPS Production
2.5. Resistance to Simulated Gastro-Intestinal Conditions
2.6. Autoaggregation and Cell Surface Hydrophobicity
2.7. Antimicrobial Activity
2.8. In Vitro Cholesterol Lowering Ability
2.9. Antibiotic Susceptibility
2.10. Hemolytic Activity
2.11. Biogenic Amines Production
2.12. Statistical Analysis
3. Results and Discussion
3.1. Physico-Chemical Characterization of Honey Samples
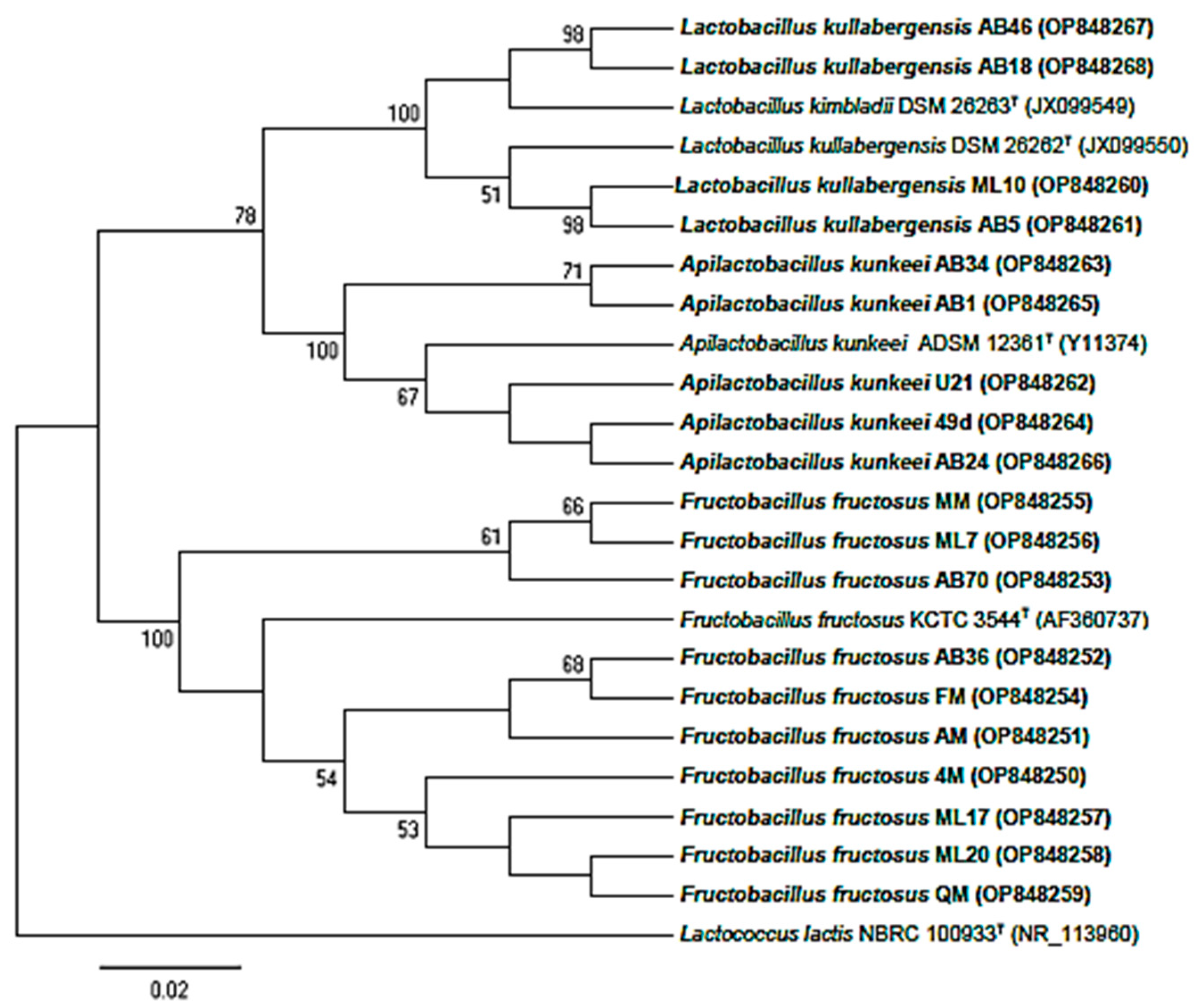
3.2. Molecular Identification and Phylogenetic Analysis
3.3. Biochemical Characterization
3.4. Assessment of Proteolytic, Lipolytic Activities and EPS Production
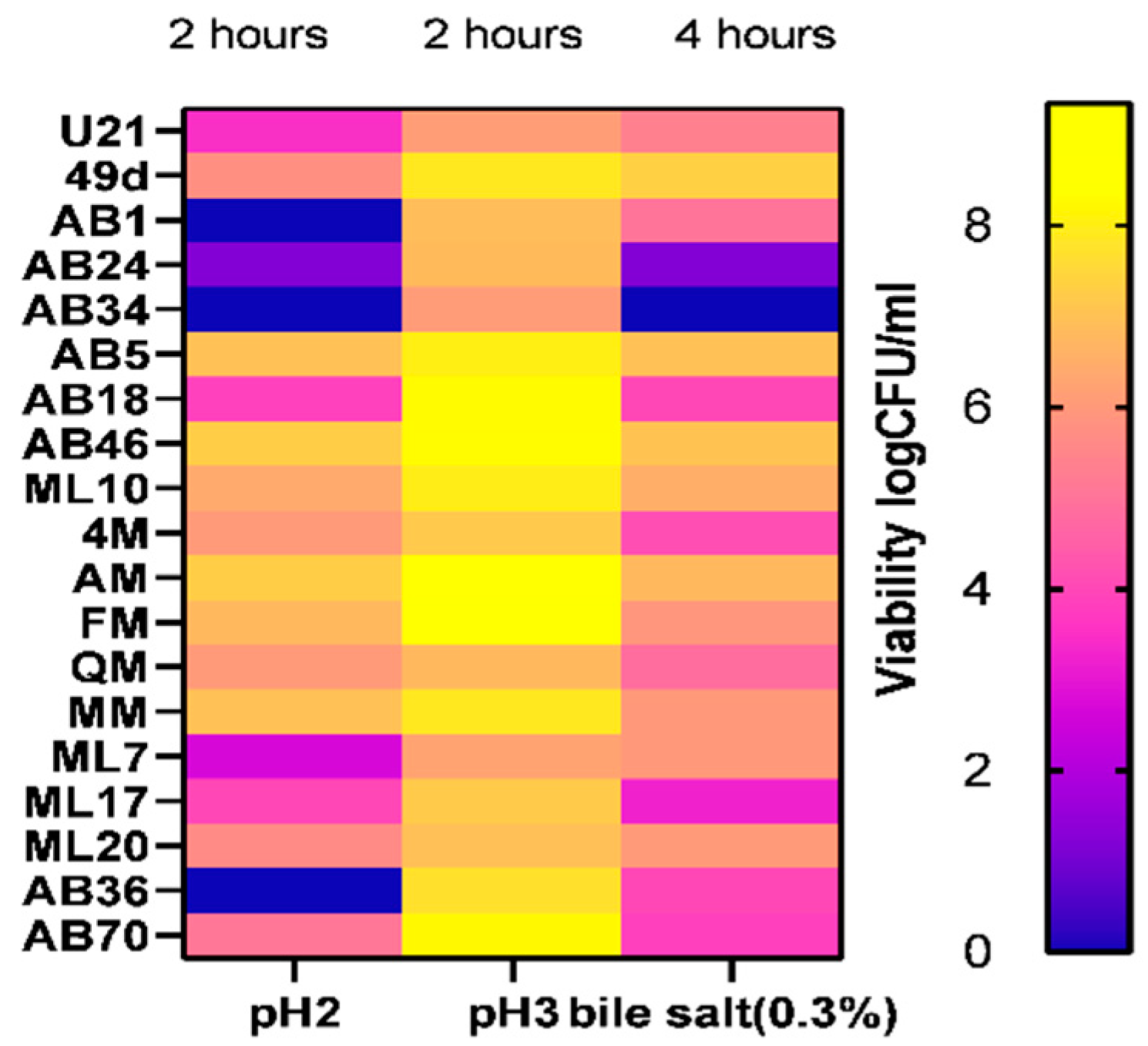
3.5. Resistance to Simulated Gastro-Intestinal Conditions
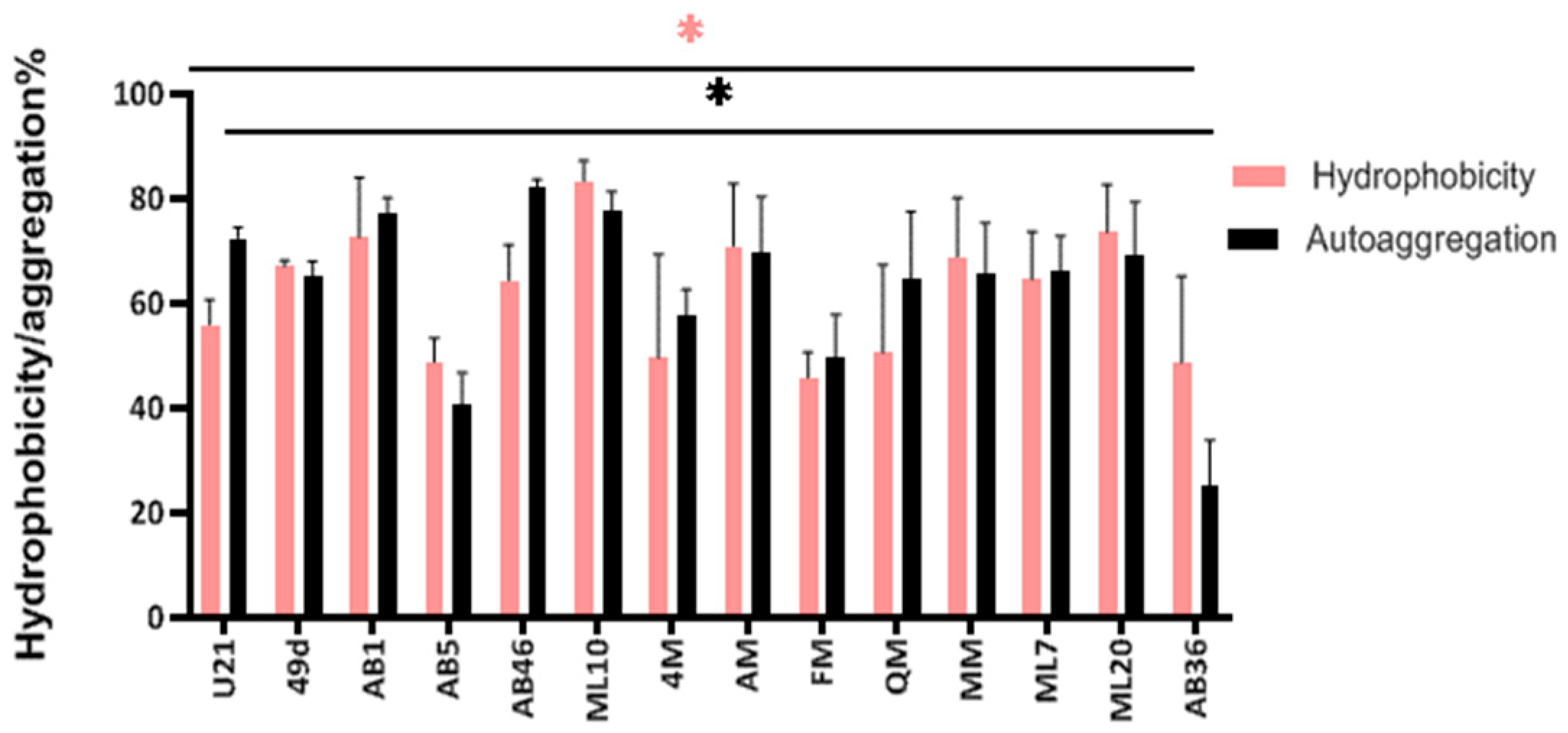
3.6. Autoaggregation and Cell Surface Hydrophobicity
3.7. Antimicrobial Activity
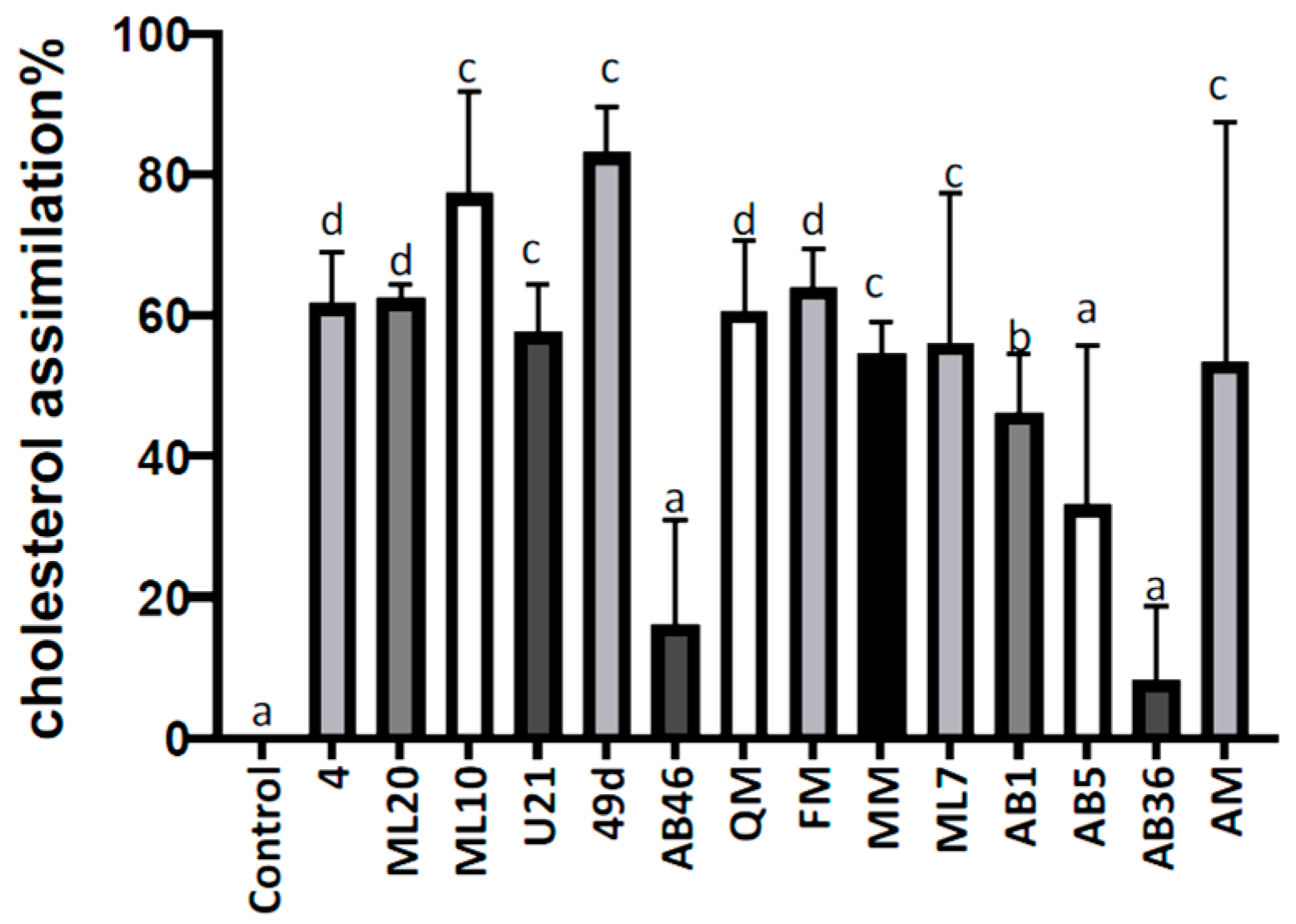
3.8. In Vitro Cholesterol Lowering Ability
3.9. Antibiotic Susceptibility
3.10. Hemolysis
3.11. Biogenic Amines Production
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oak, S.J.; Jha, R. The Effects of Probiotics in Lactose Intolerance: A Systematic Review. Crit. Rev. Food Sci. Nutr. 2019, 59, 1675–1683. [Google Scholar] [CrossRef]
- Pimentel, T.C.; Gomes de Oliveira, L.I.; de Lourdes Chaves Macedo, E.; Costa, G.N.; Dias, D.R.; Schwan, R.F.; Magnani, M. Understanding the Potential of Fruits, Flowers, and Ethnic Beverages as Valuable Sources of Techno-Functional and Probiotics Strains: Current Scenario and Main Challenges. Trends Food Sci. Technol. 2021, 114, 25–59. [Google Scholar] [CrossRef]
- Gupta, S.; Abu-Ghannam, N. Probiotic Fermentation of Plant Based Products: Possibilities and Opportunities. Crit. Rev. Food Sci. Nutr. 2012, 52, 183–199. [Google Scholar] [CrossRef]
- Xiao, J.; Zhang, Y.; Yang, Z. Lactic Acid Bacteria in Health and Disease. In Lactic Acid Bacteria: Fundamentals and Practice; Springer: Dordrecht, The Netherlands, 2014; pp. 303–374. ISBN 978-94-017-8841-0. [Google Scholar] [CrossRef]
- Chagas Barros, R.G.; Santos de Oliveira, C.; Santos Oliveira, L.T.; Pereira, U.C.; Matos Silva, T.O.; Denadai, M.; Narain, N. Enhancement of Phenolic Antioxidants Production in Submerged Cultures of Endophytic Microorganisms Isolated from Achachairu (Garcinia humilis), Araçá-Boi (Eugenia stipitata) and Bacaba (Oenocarpus bacaba) Fruits. LWT 2019, 111, 370–377. [Google Scholar] [CrossRef]
- Linares-Morales, J.R.; Cuellar-Nevárez, G.E.; Rivera-Chavira, B.E.; Gutiérrez-Méndez, N.; Pérez-Vega, S.B.; Nevárez-Moorillón, G.V. Selection of Lactic Acid Bacteria Isolated from Fresh Fruits and Vegetables Based on Their Antimicrobial and Enzymatic Activities. Foods 2020, 9, 1399. [Google Scholar] [CrossRef] [PubMed]
- Vásquez, A.; Forsgren, E.; Fries, I.; Paxton, R.J.; Flaberg, E.; Szekely, L.; Olofsson, T.C. Symbionts as Major Modulators of Insect Health: Lactic Acid Bacteria and Honeybees. PLoS ONE 2012, 7, e33188. [Google Scholar] [CrossRef]
- Olofsson, T.C.; Vásquez, A. Detection and Identification of a Novel Lactic Acid Bacterial Flora within the Honey Stomach of the Honeybee Apis mellifera. Curr. Microbiol. 2008, 57, 356–363. [Google Scholar] [CrossRef]
- Takatani, N.; Endo, A. Viable Fructophilic Lactic Acid Bacteria Present in Honeybee-Based Food Products. FEMS Microbiol. Lett. 2021, 368, fnab150. [Google Scholar] [CrossRef]
- Li, T.T.; Gu, C.T. Apilactobacillus zhangqiuensis Sp. Nov. and Apilactobacillus xinyiensis Sp. Nov., Isolated from the Gut of Honeybee (Apis mellifera). Int. J. Syst. Evol. Microbiol. 2022, 72, 005402. [Google Scholar] [CrossRef]
- Olofsson, T.; Vasquez, A. Lactobacillus Apinorum and Lactobacillus Mellifer from Honeybees in Medical, Food and Feed Applications. 2019. Available online: https://patents.google.com/patent/WO2016024910A1/en (accessed on 23 May 2023).
- Butler, È.; Alsterfjord, M.; Olofsson, T.C.; Karlsson, C.; Malmström, J.; Vásquez, A. Proteins of Novel Lactic Acid Bacteria from Apis mellifera mellifera: An Insight into the Production of Known Extra-Cellular Proteins during Microbial Stress. BMC Microbiol. 2013, 13, 235. [Google Scholar] [CrossRef] [Green Version]
- Iorizzo, M.; Testa, B.; Lombardi, S.J.; Ganassi, S.; Ianiro, M.; Letizia, F.; Succi, M.; Tremonte, P.; Vergalito, F.; Cozzolino, A.; et al. Antimicrobial Activity against Paenibacillus larvae and Functional Properties of Lactiplantibacillus plantarum Strains: Potential Benefits for Honeybee Health. Antibiotics 2020, 9, 442. [Google Scholar] [CrossRef] [PubMed]
- Westfall, S.; Lomis, N.; Prakash, S. Longevity Extension in Drosophila through Gut-Brain Communication. Sci. Rep. 2018, 8, 8362. [Google Scholar] [CrossRef]
- Harris, J.W.; Woodring, J. Effects of Stress, Age, Season, and Source Colony on Levels of Octopamine, Dopamine and Serotonin in the Honey Bee (Apis mellifera L.) Brain. J. Insect Physiol. 1992, 38, 29–35. [Google Scholar] [CrossRef]
- Asama, T.; Arima, T.-H.; Gomi, T.; Keishi, T.; Tani, H.; Kimura, Y.; Tatefuji, T.; Hashimoto, K. Lactobacillus Kunkeei YB38 from Honeybee Products Enhances IgA Production in Healthy Adults. J. Appl. Microbiol. 2015, 119, 818–826. [Google Scholar] [CrossRef]
- Butler, É.; Oien, R.F.; Lindholm, C.; Olofsson, T.C.; Nilson, B.; Vásquez, A. A Pilot Study Investigating Lactic Acid Bacterial Symbionts from the Honeybee in Inhibiting Human Chronic Wound Pathogens. Int. Wound J. 2016, 13, 729–737. [Google Scholar] [CrossRef] [PubMed]
- Olofsson, T.C.; Butler, È.; Markowicz, P.; Lindholm, C.; Larsson, L.; Vásquez, A. Lactic Acid Bacterial Symbionts in Honeybees–an Unknown Key to Honey’s Antimicrobial and Therapeutic Activities. Int. Wound J. 2016, 13, 668–679. [Google Scholar] [CrossRef]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.; Harris, H.M.; Mattarelli, P.; O’Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J. A Taxonomic Note on the Genus Lactobacillus: Description of 23 Novel Genera, Emended Description of the Genus Lactobacillus Beijerinck 1901, and Union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef] [PubMed]
- Endo, A. Fructophilic Lactic Acid Bacteria Inhabit Fructose-Rich Niches in Nature. Microb. Ecol. Health Dis. 2012, 23, 18563. [Google Scholar] [CrossRef]
- Maeno, S.; Nishimura, H.; Tanizawa, Y.; Dicks, L.; Arita, M.; Endo, A. Unique Niche-Specific Adaptation of Fructophilic Lactic Acid Bacteria and Proposal of Three Apilactobacillus Species as Novel Members of the Group. BMC Microbiol. 2021, 21, 41. [Google Scholar] [CrossRef]
- Endo, A.; Salminen, S. Honeybees and Beehives Are Rich Sources for Fructophilic Lactic Acid Bacteria. Syst. Appl. Microbiol. 2013, 36, 444–448. [Google Scholar] [CrossRef]
- Endo, A.; Maeno, S.; Tanizawa, Y.; Kneifel, W.; Arita, M.; Dicks, L.; Salminen, S. Fructophilic Lactic Acid Bacteria, a Unique Group of Fructose-Fermenting Microbes. Appl. Environ. Microbiol. 2018, 84, e01290-18. [Google Scholar] [CrossRef] [Green Version]
- Berríos, P.; Fuentes, J.A.; Salas, D.; Carreño, A.; Aldea, P.; Fernández, F.; Trombert, A.N. Inhibitory Effect of Biofilm-Forming Lactobacillus kunkeei Strains against Virulent Pseudomonas aeruginosa in Vitro and in Honeycomb Moth (Galleria mellonella) Infection Model. Benef. Microbes 2018, 9, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Kleerebezem, M.; Hols, P.; Bernard, E.; Rolain, T.; Zhou, M.; Siezen, R.J.; Bron, P.A. The Extracellular Biology of the Lactobacilli. FEMS Microbiol. Rev. 2010, 34, 199–230. [Google Scholar] [CrossRef] [PubMed]
- Sakandar, H.A.; Kubow, S.; Sadiq, F.A. Isolation and In-Vitro Probiotic Characterization of Fructophilic Lactic Acid Bacteria from Chinese Fruits and Flowers. LWT 2019, 104, 70–75. [Google Scholar] [CrossRef]
- Acín Albiac, M.; Di Cagno, R.; Filannino, P.; Cantatore, V.; Gobbetti, M. How Fructophilic Lactic Acid Bacteria May Reduce the FODMAPs Content in Wheat-Derived Baked Goods: A Proof of Concept. Microb. Cell Factories 2020, 19, 182. [Google Scholar] [CrossRef] [PubMed]
- Bivolarski, V.; Iliev, I.; Ivanova, I.; Nikolova, M.; Salim, A.; Mihaylova, G.; Vasileva, T. Characterization of Structure/Prebiotic Potential Correlation of Glucans and Oligosaccharides Synthetized by Glucansucrases from Fructophilic Lactic Acid Bacteria from Honey Bee Apis mellifera. Biotechnol. Biotechnol. Equip. 2021, 35, 674–687. [Google Scholar] [CrossRef]
- Sugiyama, K.; Iijima, K.; Yoshino, M.; Dohra, H.; Tokimoto, Y.; Nishikawa, K.; Idogaki, H.; Yoshida, N. Nicotinamide Mononucleotide Production by Fructophilic Lactic Acid Bacteria. Sci. Rep. 2021, 11, 7662. [Google Scholar] [CrossRef]
- Belhadj, H.; Harzallah, D.; Khennouf, S.; Dahamna, S.; Bouharati, S.; Baghiani, A. Isolation, Identification and Antimicrobial Activity of Lactic Acid Bacteria from Algerian Honeybee Collected Pollen. Acta Hortic. 2010, 854, 51–58. [Google Scholar] [CrossRef]
- Belhadj, H.; Harzallah, D.; Bouamra, D.; Khennouf, S.; Dahamna, S.; Ghadbane, M. Phenotypic and Genotypic Characterization of Some Lactic Acid Bacteria Isolated from Bee Pollen: A Preliminary Study. Biosci. Microbiota Food Health 2014, 33, 11–23. [Google Scholar] [CrossRef] [Green Version]
- Chahbar, N.; Mahamed, A.L. Contribution to Identification of the Microflora of the Digestive Tract and Pollen of Algerian Honeybees: Apis mellifera intermissa and Apis mellifera sahariensis. Int. J. Curr. Microbiol. Appl. Sci. 2014, 3, 7. [Google Scholar]
- Homrani, M.; Dalache, F.; Bouzouina, M.; Nemmiche, S.; Homrani, A. Antibacterial Activity of Lactobacilli Detected in Algerian Raw Honeys against Gram-Negative Bacteria. South Asian J. Exp. Biol. 2018, 8, 83–90. [Google Scholar]
- FAOLEX. Codex Alimentarius Directive 2014/63/EU of the European Parliament and of the Council Amending Council Directive 2001/110/EC Relating to Honey. Off. J. Eur. Union L 2014, 164, 1–5. Available online: https://www.fao.org/faolex/results/details/en/c/LEX-FAOC134436 (accessed on 21 December 2022).
- Mathialagan, M.; Thangaraj Edward, Y.S.J.; David, P.M.M.; Senthilkumar, M.; Srinivasan, M.R.; Mohankumar, S. Isolation, Characterization and Identification of Probiotic Lactic Acid Bacteria (LAB) from Honey Bees. Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 894–906. [Google Scholar] [CrossRef]
- Hammes, W.P.; Hertel, C. The Genera Lactobacillus and Carnobacterium. In The Prokaryotes; Springer: New York, NY, USA, 1992; pp. 320–403. [Google Scholar] [CrossRef]
- Klayraung, S.; Viernstein, H.; Sirithunyalug, J.; Okonogi, S. Probiotic Properties of Lactobacilli Isolated from Thai Traditional Food. Sci. Pharm. 2008, 76, 485–504. [Google Scholar] [CrossRef] [Green Version]
- Mora, D.; Maguin, E.; Masiero, M.; Parini, C.; Ricci, G.; Manachini, P.L.; Daffonchio, D. Characterization of Urease Genes Cluster of Streptococcus thermophilus. J. Appl. Microbiol. 2004, 96, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.-H.; Ha, S.-M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A Taxonomically United Database of 16S RRNA Gene Sequences and Whole-Genome Assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef] [PubMed]
- Franciosi, E.; Settanni, L.; Cavazza, A.; Poznanski, E. Biodiversity and Technological Potential of Wild Lactic Acid Bacteria from Raw Cows’ Milk. Int. Dairy J. 2009, 19, 3–11. [Google Scholar] [CrossRef]
- Sierra, G. A Simple Method for the Detection of Lipolytic Activity of Micro-Organisms and Some Observations on the Influence of the Contact between Cells and Fatty Substrates. Antonie Van Leeuwenhoek 1957, 23, 15–22. [Google Scholar] [CrossRef]
- Leveau, J.Y.; Bouix, M.; De Roissart, H. La Flore Lactique. In Techniques d’analyse et de Contrôle Dans Les Industries Agro Alimentaires; Lavoisier: Paris, France, 1991; pp. 152–186. [Google Scholar]
- Balzaretti, S.; Taverniti, V.; Rondini, G.; Minuzzo, M.; Remagni, M.; Fiore, W.; Arioli, S.; Guglielmetti, S. The Vaginal Isolate Lactobacillus Paracasei LPC-S01 (DSM 26760) Is Suitable for Oral Administration. Front. Microbiol. 2015, 6, 952. [Google Scholar] [CrossRef] [Green Version]
- Vergalito, F.; Testa, B.; Cozzolino, A.; Letizia, F.; Succi, M.; Lombardi, S.J.; Tremonte, P.; Pannella, G.; Di Marco, R.; Sorrentino, E. Potential Application of Apilactobacillus kunkeei for Human Use: Evaluation of Probiotic and Functional Properties. Foods 2020, 9, 1535. [Google Scholar] [CrossRef]
- Cozzolino, A.; Vergalito, F.; Tremonte, P.; Iorizzo, M.; Lombardi, S.J.; Sorrentino, E.; Luongo, D.; Coppola, R.; Di Marco, R.; Succi, M. Preliminary Evaluation of the Safety and Probiotic Potential of Akkermansia muciniphila DSM 22959 in Comparison with Lactobacillus rhamnosus GG. Microorganisms 2020, 8, 189. [Google Scholar] [CrossRef] [Green Version]
- Ebrahimi, M.; Sadeghi, A.; Rahimi, D.; Purabdolah, H.; Shahryari, S. Postbiotic and Anti-Aflatoxigenic Capabilities of Lactobacillus kunkeei as the Potential Probiotic LAB Isolated from the Natural Honey. Probiotics Antimicrob. Proteins 2020, 13, 343–355. [Google Scholar] [CrossRef] [PubMed]
- Testa, B.; Lombardi, S.J.; Macciola, E.; Succi, M.; Tremonte, P.; Iorizzo, M. Efficacy of Olive Leaf Extract (Olea europaea L. cv Gentile Di Larino) in Marinated Anchovies (Engraulis encrasicolus, L.) Process. Heliyon 2019, 5, e01727. [Google Scholar] [CrossRef] [Green Version]
- Tremonte, P.; Pannella, G.; Succi, M.; Tipaldi, L.; Sturchio, M.; Coppola, R.; Luongo, D.; Sorrentino, E. Antimicrobial Activity of Lactobacillus plantarum Strains Isolated from Different Environments: A Preliminary Study. Int. Food Res. J. 2017, 24, 852–859. [Google Scholar]
- Rudel, L.L.; Morris, M.D. Determination of Cholesterol Using O-Phthalaldehyde. J. Lipid Res. 1973, 14, 364–366. [Google Scholar] [CrossRef] [PubMed]
- Temmerman, R.; Pot, B.; Huys, G.; Swings, J. Identification and Antibiotic Susceptibility of Bacterial Isolates from Probiotic Products. Int. J. Food Microbiol. 2003, 81, 1–10. [Google Scholar] [CrossRef]
- Mårtensson, A.; Abolhalaj, M.; Lindstedt, M.; Mårtensson, A.; Olofsson, T.C.; Vásquez, A.; Greiff, L.; Cervin, A. Clinical Efficacy of a Topical Lactic Acid Bacterial Microbiome in Chronic Rhinosinusitis: A Randomized Controlled Trial. Laryngoscope Investig. Otolaryngol. 2017, 2, 410–416. [Google Scholar] [CrossRef]
- Ouchemoukh, S.; Louaileche, H.; Schweitzer, P. Physicochemical Characteristics and Pollen Spectrum of Some Algerian Honeys. Food Control 2007, 18, 52–58. [Google Scholar] [CrossRef]
- Adjlane, N.; Haddad, N.; Ameur, K.L.; Kesraoui, S.; Moussaoui, D. Physicochemical and Microbiological Characteristics of Some Samples of Honey Produced by Beekeepers in Algeria. Acta Technol. Agric. 2014, 17, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Abdellah, F.; Makhloufi, C.; Boukraa, L.; Hammoudi, S.M.; Amel, S.; Dellel, N.; Benamara, A.; Benhadiri, M.; Marouf, N.; Benaraba, R. Physico-Chemical Properties and Antibacterial and Antioxidant Activity of Two Varieties of Honey from Algerian Steppe. J. Apitherapy Nat. 2020, 3, 59–74. [Google Scholar] [CrossRef]
- Gonnet, M. Le Miel. Composition, Proprietes, Conservation; OPIDA: Avignon, France, 1982; p. 30. [Google Scholar]
- Ellegaard, K.M.; Engel, P. New Reference Genome Sequences for 17 Bacterial Strains of the Honey Bee Gut Microbiota. Microbiol. Resour. Announc. 2018, 7, e00834-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papa, G.; Di Prisco, G.; Spini, G.; Puglisi, E.; Negri, I. Acute and Chronic Effects of Titanium Dioxide (TiO2) PM1 on Honey Bee Gut Microbiota under Laboratory Conditions. Sci. Rep. 2021, 11, 5946. [Google Scholar] [CrossRef]
- Duong, B.; Lien, N.; Thu, H.; Hoa, N.; Pham, L.; Yun, B.-R.; Yoo, M.-S.; Cho, Y.S.; Quyen, D. Investigation of the Gut Microbiome of Apis Cerana Honeybees from Vietnam. Biotechnol. Lett. 2020, 42, 2309–2317. [Google Scholar] [CrossRef]
- Maddaloni, M.; Hoffman, C.; Pascual, D.W. Paratransgenesis Feasibility in the Honeybee (Apis mellifera) Using Fructobacillus Fructosus Commensal. J. Appl. Microbiol. 2014, 117, 1572–1584. [Google Scholar] [CrossRef] [PubMed]
- Vásquez, A.; Olofsson, T.C. The Lactic Acid Bacteria Involved in the Production of Bee Pollen and Bee Bread. J. Apic. Res. 2009, 48, 189–195. [Google Scholar] [CrossRef]
- Erban, T.; Ledvinka, O.; Kamler, M.; Hortova, B.; Nesvorna, M.; Tyl, J.; Titera, D.; Markovic, M.; Hubert, J. Bacterial Community Associated with Worker Honeybees (Apis mellifera) Affected by European Foulbrood. PeerJ 2017, 5, e3816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filannino, P.; Di Cagno, R.; Addante, R.; Pontonio, E.; Gobbetti, M. Metabolism of Fructophilic Lactic Acid Bacteria Isolated from the Apis mellifera L. Bee Gut: Phenolic Acids as External Electron Acceptors. Appl. Environ. Microbiol. 2016, 82, 6899–6911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olofsson, T.C.; Alsterfjord, M.; Nilson, B.; Butler, È.; Vásquez, A. Lactobacillus apinorum Sp. Nov., Lactobacillus mellifer Sp. Nov., Lactobacillus mellis Sp. Nov., Lactobacillus melliventris Sp. Nov., Lactobacillus kimbladii Sp. Nov., Lactobacillus helsingborgensis Sp. Nov. and Lactobacillus kullabergensis Sp. Nov., Isolated from the Honey Stomach of the Honeybee Apis mellifera. Int. J. Syst. Evol. Microbiol. 2014, 64, 3109. [Google Scholar] [CrossRef]
- Aziz, G.; Tariq, M.; Zaidi, A.H. Mining Indigenous Honeybee Gut Microbiota for Lactobacillus with Probiotic Potential. Microbiology 2021, 167, 001032. [Google Scholar] [CrossRef]
- Leska, A.; Nowak, A.; Motyl, I. Isolation and Some Basic Characteristics of Lactic Acid Bacteria from Honeybee (Apis mellifera L.) Environment—A Preliminary Study. Agriculture 2022, 12, 1562. [Google Scholar] [CrossRef]
- Li, T.T.; Gu, C.T. Lactobacillus huangpiensis Sp. Nov. and Lactobacillus laiwuensis Sp. Nov., Isolated from the Gut of Honeybee (Apis mellifera). Int. J. Syst. Evol. Microbiol. 2022, 72, 005237. [Google Scholar] [CrossRef]
- Liu, D.D.; Li, Y.Q.; Zhang, L.P.; Ding, W.; Tian, W.L.; Gu, C.T. Apilactobacillus nanyangensis Sp. Nov., Secundilactobacillus hailunensis Sp. Nov., Secundilactobacillus yichangensis Sp. Nov., Levilactobacillus andaensis Sp. Nov., Levilactobacillus wangkuiensis Sp. Nov., Levilactobacillus lanxiensis Sp. Nov., Lacticaseibacillus mingshuiensis Sp. Nov. and Lacticaseibacillus suilingensis Sp. Nov., Isolated from Traditional Chinese Pickle and the Gut of Honeybee (Apis mellifera). Int. J. Syst. Evol. Microbiol. 2021, 71, 004898. [Google Scholar] [CrossRef]
- Corby-Harris, V.; Maes, P.; Anderson, K.E. The Bacterial Communities Associated with Honey Bee (Apis mellifera) Foragers. PLoS ONE 2014, 9, e95056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwong, W.K.; Mancenido, A.L.; Moran, N.A. Genome Sequences of Lactobacillus Sp. Strains WkB8 and WkB10, Members of the Firm-5 Clade, from Honey Bee Guts. Genome Announc. 2014, 2, e01176-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellegaard, K.M.; Tamarit, D.; Javelind, E.; Olofsson, T.C.; Andersson, S.G.; Vásquez, A. Extensive Intra-Phylotype Diversity in Lactobacilli and Bifidobacteria from the Honeybee Gut. BMC Genom. 2015, 16, 284. [Google Scholar] [CrossRef] [Green Version]
- Endo, A.; Irisawa, T.; Futagawa-Endo, Y.; Takano, K.; du Toit, M.; Okada, S.; Dicks, L.M.T. Characterization and Emended Description of Lactobacillus Kunkeei as a Fructophilic Lactic Acid Bacterium. Int. J. Syst. Evol. Microbiol. 2012, 62, 500–504. [Google Scholar] [CrossRef] [Green Version]
- Bello, S.; Rudra, B.; Gupta, R.S. Phylogenomic and Comparative Genomic Analyses of Leuconostocaceae Species: Identification of Molecular Signatures Specific for the Genera Leuconostoc, Fructobacillus and Oenococcus and Proposal for a Novel Genus Periweissella Gen. Nov. Int. J. Syst. Evol. Microbiol. 2022, 72, 005284. [Google Scholar] [CrossRef]
- Iorizzo, M.; Pannella, G.; Lombardi, S.J.; Ganassi, S.; Testa, B.; Succi, M.; Sorrentino, E.; Petrarca, S.; De Cristofaro, A.; Coppola, R. Inter-and Intra-Species Diversity of Lactic Acid Bacteria in Apis Mellifera Ligustica Colonies. Microorganisms 2020, 8, 1578. [Google Scholar] [CrossRef]
- Taylor, M.A.; Robertson, A.W.; Biggs, P.J.; Richards, K.K.; Jones, D.F.; Parkar, S.G. The Effect of Carbohydrate Sources: Sucrose, Invert Sugar and Components of Mānuka Honey, on Core Bacteria in the Digestive Tract of Adult Honey Bees (Apis mellifera). PLoS ONE 2019, 14, e0225845. [Google Scholar] [CrossRef] [Green Version]
- Zheng, H.; Perreau, J.; Powell, J.E.; Han, B.; Zhang, Z.; Kwong, W.K.; Tringe, S.G.; Moran, N.A. Division of Labor in Honey Bee Gut Microbiota for Plant Polysaccharide Digestion. Proc. Natl. Acad. Sci. USA 2019, 116, 25909–25916. [Google Scholar] [CrossRef]
- Endo, A.; Dicks, L.M.T. The Genus Fructobacillus. In Lactic Acid Bacteria: Biodiversity and Taxonomy; John Wiley & Sons: New York, NY, USA, 2014; pp. 381–389. [Google Scholar] [CrossRef]
- Sulistiani. Selection of Potential Probiotic Lactic Acid Bacteria Isolated from Palm Sap (Borassus Flabellifer Linn.) Origin Kupang, East Nusa Tenggara. AIP Conf. Proc. 2018, 2002, 2002–020059. [Google Scholar] [CrossRef]
- Di Cagno, R.; Filannino, P.; Cantatore, V.; Gobbetti, M. Novel Solid-State Fermentation of Bee-Collected Pollen Emulating the Natural Fermentation Process of Bee Bread. Food Microbiol. 2019, 82, 218–230. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, S.D.; Araújo, C.M.; Borges, G.d.S.C.; Lima, M.d.S.; Viera, V.B.; Garcia, E.F.; de Souza, E.L.; de Oliveira, M.E.G. Improvement in Physicochemical Characteristics, Bioactive Compounds and Antioxidant Activity of Acerola (Malpighia Emarginata D.C.) and Guava (Psidium guajava L.) Fruit by-Products Fermented with Potentially Probiotic Lactobacilli. LWT 2020, 134, 110200. [Google Scholar] [CrossRef]
- Flemming, H.C.; Wingender, J. The Biofilm Matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Ispirli, H.; Dertli, E. Detection of Fructophilic Lactic Acid Bacteria (FLAB) in Bee Bread and Bee Pollen Samples and Determination of Their Functional Roles. J. Food Process. Preserv. 2021, 45, e15414. [Google Scholar] [CrossRef]
- Meng, X.; Gangoiti, J.; Wang, X.; Grijpstra, P.; van Leeuwen, S.S.; Pijning, T.; Dijkhuizen, L. Biochemical Characterization of a GH70 Protein from Lactobacillus kunkeei DSM 12361 with Two Catalytic Domains Involving Branching Sucrase Activity. Appl. Microbiol. Biotechnol. 2018, 102, 7935–7950. [Google Scholar] [CrossRef] [PubMed]
- Asenjo, F.; Olmos, A.; Henríquez-Piskulich, P.; Polanco, V.; Aldea, P.; Ugalde, J.A.; Trombert, A.N. Genome Sequencing and Analysis of the First Complete Genome of Lactobacillus kunkeei Strain MP2, an Apis mellifera Gut Isolate. PeerJ 2016, 4, e1950. [Google Scholar] [CrossRef] [Green Version]
- Djukic, M.; Poehlein, A.; Strauß, J.; Tann, F.J.; Leimbach, A.; Hoppert, M.; Daniel, R. High Quality Draft Genome of Lactobacillus kunkeei EFB6, Isolated from a German European Foulbrood Outbreak of Honeybees. Stand. Genom. Sci. 2015, 10, 16. [Google Scholar] [CrossRef] [Green Version]
- Endo, A.; Tanizawa, Y.; Tanaka, N.; Maeno, S.; Kumar, H.; Shiwa, Y.; Okada, S.; Yoshikawa, H.; Dicks, L.; Nakagawa, J. Comparative Genomics of Fructobacillus Spp. and Leuconostoc Spp. Reveals Niche-Specific Evolution of Fructobacillus Spp. BMC Genom. 2015, 16, 1117. [Google Scholar] [CrossRef] [Green Version]
- Endo, A.; Okada, S. Reclassification of the Genus Leuconostoc and Proposals of Fructobacillus fructosus Gen. Nov., Comb. Nov., Fructobacillus durionis Comb. Nov., Fructobacillus ficulneus Comb. Nov. and Fructobacillus pseudoficulneus Comb. Nov. Int. J. Syst. Evol. Microbiol. 2008, 58, 2195–2205. [Google Scholar] [CrossRef] [Green Version]
- Verón, H.E.; Di Risio, H.D.; Isla, M.I.; Torres, S. Isolation and Selection of Potential Probiotic Lactic Acid Bacteria from Opuntia Ficus-Indica Fruits That Grow in Northwest Argentina. LWT 2017, 84, 231–240. [Google Scholar] [CrossRef]
- Succi, M.; Tremonte, P.; Pannella, G.; Tipaldi, L.; Cozzolino, A.; Romaniello, R.; Sorrentino, E.; Coppola, R. Pre-Cultivation with Selected Prebiotics Enhances the Survival and the Stress Response of Lactobacillus rhamnosus Strains in Simulated Gastrointestinal Transit. Front. Microbiol. 2017, 8, 1067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Cui, Y.; Qu, X. Mechanisms and Improvement of Acid Resistance in Lactic Acid Bacteria. Arch. Microbiol. 2018, 200, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Guan, N.; Liu, L. Microbial Response to Acid Stress: Mechanisms and Applications. Appl. Microbiol. Biotechnol. 2020, 104, 51–65. [Google Scholar] [CrossRef] [Green Version]
- Bibi, A.; Xiong, Y.; Rajoka, M.S.R.; Mehwish, H.M.; Radicetti, E.; Umair, M.; Shoukat, M.; Khan, M.K.I.; Aadil, R.M. Recent Advances in the Production of Exopolysaccharide (EPS) from Lactobacillus Spp. and Its Application in the Food Industry: A Review. Sustainability 2021, 13, 12429. [Google Scholar] [CrossRef]
- Cotter, P.D.; Hill, C. Surviving the Acid Test: Responses of Gram-Positive Bacteria to Low PH. Microbiol. Mol. Biol. Rev. 2003, 67, 429–453. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, L.; Margolles, A.; Sánchez, B. Bile Resistance Mechanisms in Lactobacillus and Bifidobacterium. Front. Microbiol. 2013, 4, 396. [Google Scholar] [CrossRef] [Green Version]
- Iorizzo, M.; Lombardi, S.J.; Ganassi, S.; Testa, B.; Ianiro, M.; Letizia, F.; Succi, M.; Tremonte, P.; Vergalito, F.; Cozzolino, A. Antagonistic Activity against Ascosphaera apis and Functional Properties of Lactobacillus kunkeei Strains. Antibiotics 2020, 9, 262. [Google Scholar] [CrossRef]
- Trunk, T.; Khalil, H.S.; Leo, J.C.; Trunk, T.; Khalil, H.S.; Leo, J.C. Bacterial Autoaggregation. AIMS Microbiol. 2018, 4, 140–164. [Google Scholar] [CrossRef]
- Serna-Cock, L.; Rojas-Dorado, M.; Ordoñez-Artunduaga, D.; García-Salazar, A.; García-González, E.; Aguilar, C.N. Crude Extracts of Metabolites from Co-Cultures of Lactic Acid Bacteria Are Highly Antagonists of Listeria Monocytogenes. Heliyon 2019, 5, e02448. [Google Scholar] [CrossRef] [Green Version]
- Zendo, T.; Ohashi, C.; Maeno, S.; Piao, X.; Salminen, S.; Sonomoto, K.; Endo, A. Kunkecin A, a New Nisin Variant Bacteriocin Produced by the Fructophilic Lactic Acid Bacterium, Apilactobacillus kunkeei FF30-6 Isolated From Honey Bees. Front. Microbiol. 2020, 11, 2130. [Google Scholar] [CrossRef]
- Dixon, A.; Robertson, K.; Yung, A.; Que, M.; Randall, H.; Wellalagodage, D.; Cox, T.; Robertson, D.; Chi, C.; Sun, J. Efficacy of Probiotics in Patients of Cardiovascular Disease Risk: A Systematic Review and Meta-Analysis. Curr. Hypertens. Rep. 2020, 22, 74. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Guo, M.-J.; Gao, Q.; Yang, J.-F.; Yang, L.; Pang, X.-L.; Jiang, X.-J. The Effects of Probiotics on Total Cholesterol: A Meta-Analysis of Randomized Controlled Trials. Medicine 2018, 97, e9679. [Google Scholar] [CrossRef]
- Bhat, B.; Bajaj, B.K. Multifarious Cholesterol Lowering Potential of Lactic Acid Bacteria Equipped with Desired Probiotic Functional Attributes. 3 Biotech 2020, 10, 200. [Google Scholar] [CrossRef]
- Lye, H.-S.; Rahmat-Ali, G.R.; Liong, M.-T. Mechanisms of Cholesterol Removal by Lactobacilli under Conditions That Mimic the Human Gastrointestinal Tract. Int. Dairy J. 2010, 20, 169–175. [Google Scholar] [CrossRef]
- Tomaro-Duchesneau, C.; Jones, M.L.; Shah, D.; Jain, P.; Saha, S.; Prakash, S. Cholesterol Assimilation by Lactobacillus Probiotic Bacteria: An in Vitro Investigation. BioMed Res. Int. 2014, 2014, 380316. [Google Scholar] [CrossRef] [Green Version]
- Olmos, A.; Henríquez-Piskulich, P.; Sanchez, C.; Rojas-Herrera, M.; Moreno-Pino, M.; Gómez, M.; Rodríguez Da Silva, R.; Maracaja-Coutinho, V.; Aldea, P.; Trombert, A.N. Draft Genome of Chilean Honeybee (Apis mellifera) Gut Strain Lactobacillus kunkeei MP2. Genome Announc. 2014, 2, e01013-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patil, M.; Jadhav, A.; Patil, U. Functional Characterization and in Vitro Screening of Fructobacillus Fructosus MCC 3996 Isolated from Butea Monosperma Flower for Probiotic Potential. Lett. Appl. Microbiol. 2020, 70, 331–339. [Google Scholar] [CrossRef]
- Niu, K.-M.; Kothari, D.; Cho, S.-B.; Han, S.-G.; Song, I.-G.; Kim, S.-C.; Kim, S.-K. Exploring the Probiotic and Compound Feed Fermentative Applications of Lactobacillus plantarum SK1305 Isolated from Korean Green Chili Pickled Pepper. Probiotics Antimicrob. Proteins 2019, 11, 801–812. [Google Scholar] [CrossRef]
- Gueimonde, M.; Sánchez, B.; de Los Reyes-Gavilán, C.G.; Margolles, A. Antibiotic Resistance in Probiotic Bacteria. Front. Microbiol. 2013, 4, 202. [Google Scholar] [CrossRef] [Green Version]
- Fraqueza, M.J. Antibiotic Resistance of Lactic Acid Bacteria Isolated from Dry-Fermented Sausages. Int. J. Food Microbiol. 2015, 212, 76–88. [Google Scholar] [CrossRef]
- Wolupeck, H.L.; Morete, C.A.; DallaSanta, O.R.; Luciano, F.B.; Madeira, H.M.F.; de Macedo, R.E.F. Methods for the Evaluation of Antibiotic Resistance in Lactobacillus Isolated from Fermented Sausages. Ciênc. Rural 2017, 47, 08. [Google Scholar] [CrossRef] [Green Version]
- Bin Masalam, M.S.; Bahieldin, A.; Alharbi, M.G.; Al-Masaudi, S.; Al-Jaouni, S.K.; Harakeh, S.M.; Al-Hindi, R.R. Isolation, Molecular Characterization and Probiotic Potential of Lactic Acid Bacteria in Saudi Raw and Fermented Milk. Evid. Based Complement. Alternat. Med. 2018, 2018, e797046. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Wang, Y.; Cui, H.; Li, Y.; Sun, Y.; Qiu, H.-J. Characterization of Lactic Acid Bacteria Isolated From the Gastrointestinal Tract of a Wild Boar as Potential Probiotics. Front. Vet. Sci. 2020, 7, 49. [Google Scholar] [CrossRef] [Green Version]
- Jha, V.; Sarang, C.; Sawant, D.; Nalawade, K.; Dhamapurkar, V.; Kaur, N.; Thakur, K.; Amin, S.; Mane, P.; Marath, A. Exploration of Probiotic Potential of Lactic Acid Bacteria Isolated from Different Food Sources. Am. J. Biosci. 2022, 10, 118–130. [Google Scholar]
- Ruiz-Capillas, C.; Herrero, A.M. Impact of Biogenic Amines on Food Quality and Safety. Foods 2019, 8, 62. [Google Scholar] [CrossRef] [Green Version]
- Santos, M.S. Biogenic Amines: Their Importance in Foods. Int. J. Food Microbiol. 1996, 29, 213–231. [Google Scholar] [CrossRef]
- Spano, G.; Russo, P.; Lonvaud-Funel, A.; Lucas, P.; Alexandre, H.; Grandvalet, C.; Coton, E.; Coton, M.; Barnavon, L.; Bach, B. Biogenic Amines in Fermented Foods. Eur. J. Clin. Nutr. 2010, 64, S95–S100. [Google Scholar] [CrossRef] [Green Version]
- Papageorgiou, M.; Lambropoulou, D.; Morrison, C.; Kłodzińska, E.; Namieśnik, J.; Płotka-Wasylka, J. Literature Update of Analytical Methods for Biogenic Amines Determination in Food and Beverages. TrAC Trends Anal. Chem. 2018, 98, 128–142. [Google Scholar] [CrossRef] [Green Version]
- Landete, J.M.; Ferrer, S.; Polo, L.; Pardo, I. Biogenic Amines in Wines from Three Spanish Regions. J. Agric. Food Chem. 2005, 53, 1119–1124. [Google Scholar] [CrossRef]
- Jaguey-Hernández, Y.; Aguilar-Arteaga, K.; Ojeda-Ramirez, D.; Añorve-Morga, J.; González-Olivares, L.G.; Castañeda-Ovando, A. Biogenic Amines Levels in Food Processing: Efforts for Their Control in Foodstuffs. Food Res. Int. 2021, 144, 110341. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| U21 | AB1 | 49d | AB24 | AB34 | ML10 | AB5 | AB18 | AB46 | 4M | AM | FM | QM | MM | ML7 | ML17 | ML20 | AB36 | AB70 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| L-arabinose | - | - | - | - | - | - | + | - | - | - | w | - | - | - | - | - | - | - | - |
| D-ribose | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| D-galactose | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| D-glucose | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| D-fructose | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| D-monose | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| L-rhamnose | - | - | - | - | - | - | w | - | - | - | - | - | - | - | - | - | - | - | - |
| D-mannitol | w | w | - | w | w | - | + | - | - | + | + | + | + | + | - | - | - | + | + |
| N-acetylglucosamine | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| Amygdaline | + | + | + | + | + | + | + | w | w | + | + | + | + | + | + | + | + | + | + |
| Arbutine | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| Esculine citrate de fer | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| Salicine | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| D-celobiose | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| D-maltose | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| D-lactose(bovine) | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| D-melibiose | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| D-saccharose | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| D-trehalose | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| D-raffinose | w | w | w | w | w | - | w | w | w | + | + | + | + | + | + | + | + | + | + |
| Amidon | - | - | - | - | - | - | - | w | w | - | - | - | - | - | - | - | - | - | - |
| Gentiobiose | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| D-tagatose | - | - | - | - | - | - | - | + | + | - | w | - | - | - | - | - | w | - | - |
| Potassium gluconate | w | w | - | w | - | - | - | - | - | w | w | - | - | - | - | - | w | - | - |
| U21 | AB1 | 49d | ML10 | AB5 | AB46 | 4M | AM | FM | QM | MM | ML7 | AB36 | ML20 | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| K. pneumonia ATCC 70060 | BC | 20.0 ± 0.1 | 23.00 ± 0.06 | 19.0 ± 0.06 | 18.50 ± 0.03 | 18.50 ± 0.03 | 15.50 ± 0.03 | 23.0 ± 0.2 | 10.0 ± 0.0 | 12.0 ± 0.2 | 11.50 ± 0.03 | 14.6 ± 0.3 | 23.0 ± 0.1 | 23.0 ± 0.2 | 20.0 ± 0.1 |
| CFS | - | - | - | - | - | - | - | - | - | - | - | - | - | - | |
| B. cereus ATCC 10876 | BC | 16.0 ± 0.1 | 19.5 ± 0.03 | 15.0 ± 0.2 | 22.0 ± 0.1 | 19.30 ± 0.04 | 13.5 ± 0.1 | 20.30 ± 0.04 | 16.0 ± 0.2 | 17.50 ± 0.03 | 14.0 ± 0.1 | 12.30 ± 0.08 | 13.0 ± 0.1 | 17.0 ± 0.3 | 23.0 ± 0.4 |
| CFS | - | - | - | - | - | - | - | - | - | - | - | - | - | 22.5 ± 0.5 | |
| S. aureus ATCC 43300 | BC | 23.0 ± 0.4 | 20.0 ± 0.1 | 15.0 ± 0.2 | 22.3 ± 0.1 | 17.10 ± 0.05 | 16.0 ± 0.2 | 21.0 ± 0.2 | 14.0 ± 0.4 | 16.5 ± 0.1 | 19.0 ± 0.3 | 19.0 ± 0.2 | 19.0 ± 0.2 | 19.0 ± 0.3 | 18.0 ± 0.1 |
| CFS | 15.0 ± 0.6 | - | - | - | - | - | - | - | - | - | - | - | - | - | |
| E. coli ATCC 25522 | BC | 15.0 ± 0.3 | 12.5 ± 0.1 | - | 27.6 ± 0.4 | 14.0 ± 0.2 | - | 21.0 ± 0.1 | - | 16.0 ± 0.2 | 14.6 ± 0.1 | 14.0 ± 0.06 | 13.0 ± 0.2 | 19.0 ± 0.6 | 18.0 ± 0.1 |
| CFS | - | - | - | - | - | - | - | - | - | - | - | - | - | 10.0 ± 0.2 | |
| P. aeruginosa ATCC 27853 | BC | 19.0 ± 0.3 | 14.0 ± 0.2 | - | 11.0 ± 0.2 | 13.3 ± 0.2 | - | 13.0 ± 0.2 | 10.6 ± 0.1 | 14.0 ± 0.2 | 10.00 ± 0.06 | 12.6 ± 0.2 | 08.00 ± 0.06 | 15.0 ± 0.2 | 14.0 ± 0.2 |
| CFS | - | - | - | - | - | - | - | - | - | - | - | - | - | - | |
| K. pneumonia ATCC 700603 | B. cereus ATCC 10876 | S. aureus ATCC 43300 | E. coli ATCC 25522 | P. aeruginosa ATCC 27853 | |
|---|---|---|---|---|---|
| Ampicillin | - | - | - | - | - |
| Cefoxitin | 12.0 ± 0.8 | - | 10.0 ± 0.1 | - | - |
| Chloranohenicol | 12.0 ± 0.3 | - | 18.0 ± 1.1 | - | - |
| Clindamicyn | - | - | - | - | - |
| Oxacillin | - | - | - | - | - |
| Penicillin | - | - | - | - | - |
| Tetracycline | - | 28.0 ± 0.6 | 23.00 ± 0.06 | - | - |
| Trimethoprim | - | - | 16.0 ± 0.5 | - | - |
| Vancomycin | - | - | 14.0 ± 1.0 | - | - |
| Erytromicyn | - | - | - | - | - |
| Naladixic acid | - | - | 13.0 ± 0.2 | - | 14.00 ± 0.08 |
| Streptomycin | 19.0 ± 0.9 | 28.00 ± 0.03 | 20.00 ± 0.04 | 20.0 ± 0.3 | 13.0 ± 0.4 |
| Gentamycin | 14.00 ± 0.07 | 14.0 ± 1.0 | 23.0 ± 0.3 | 13.0 ± 0.4 | 16.0 ± 0.3 |
| U21 | AB1 | 49d | ML10 | AB5 | AB46 | 4M | AM | FM | QM | MM | ML7 | AB36 | ML20 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ampicillin | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| Cefoxitin | S | S | S | I | S | S | I | S | S | S | I | I | I | S |
| Chloranohenicol | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| Clindamicyn | S | S | S | S | S | S | S | S | S | S | S | S | S | S |
| Oxacillin | R | R | R | R | R | R | R | R | R | R | R | R | R | R |
| Penicillin | S | R | R | R | R | S | R | S | R | R | I | I | R | I |
| Tetracycline | R | I | I | R | R | I | I | R | R | R | R | R | I | I |
| Trimethoprim | S | R | R | R | R | R | I | R | I | I | I | I | I | I |
| Vancomycin | R | R | R | R | R | R | R | R | R | R | R | R | R | R |
| Erythromicyn | S | S | S | S | S | S | I | S | S | S | S | S | I | S |
| Naladixic acid | R | R | R | R | R | R | R | R | R | R | R | R | R | R |
| Streptomycin | R | I | R | R | I | I | S | S | R | I | R | R | I | S |
| Gentamycin | R | R | R | R | R | I | R | R | R | R | R | R | R | I |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meradji, M.; Bachtarzi, N.; Mora, D.; Kharroub, K. Characterization of Lactic Acid Bacteria Strains Isolated from Algerian Honeybee and Honey and Exploration of Their Potential Probiotic and Functional Features for Human Use. Foods 2023, 12, 2312. https://doi.org/10.3390/foods12122312
Meradji M, Bachtarzi N, Mora D, Kharroub K. Characterization of Lactic Acid Bacteria Strains Isolated from Algerian Honeybee and Honey and Exploration of Their Potential Probiotic and Functional Features for Human Use. Foods. 2023; 12(12):2312. https://doi.org/10.3390/foods12122312
Chicago/Turabian StyleMeradji, Meriem, Nadia Bachtarzi, Diego Mora, and Karima Kharroub. 2023. "Characterization of Lactic Acid Bacteria Strains Isolated from Algerian Honeybee and Honey and Exploration of Their Potential Probiotic and Functional Features for Human Use" Foods 12, no. 12: 2312. https://doi.org/10.3390/foods12122312