
A Comprehensive Study on Antibiotic Resistance among Coagulase-Negative Staphylococci (CoNS) Strains Isolated from Ready-to-Eat Food Served in Bars and Restaurants

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Identification of CoNS by MALDI-TOF
2.2. Phenotypic Antibiotic Resistance
2.3. Molecular Mechanisms of Antibiotic Resistance
2.4. Statistical Analysis
3. Results
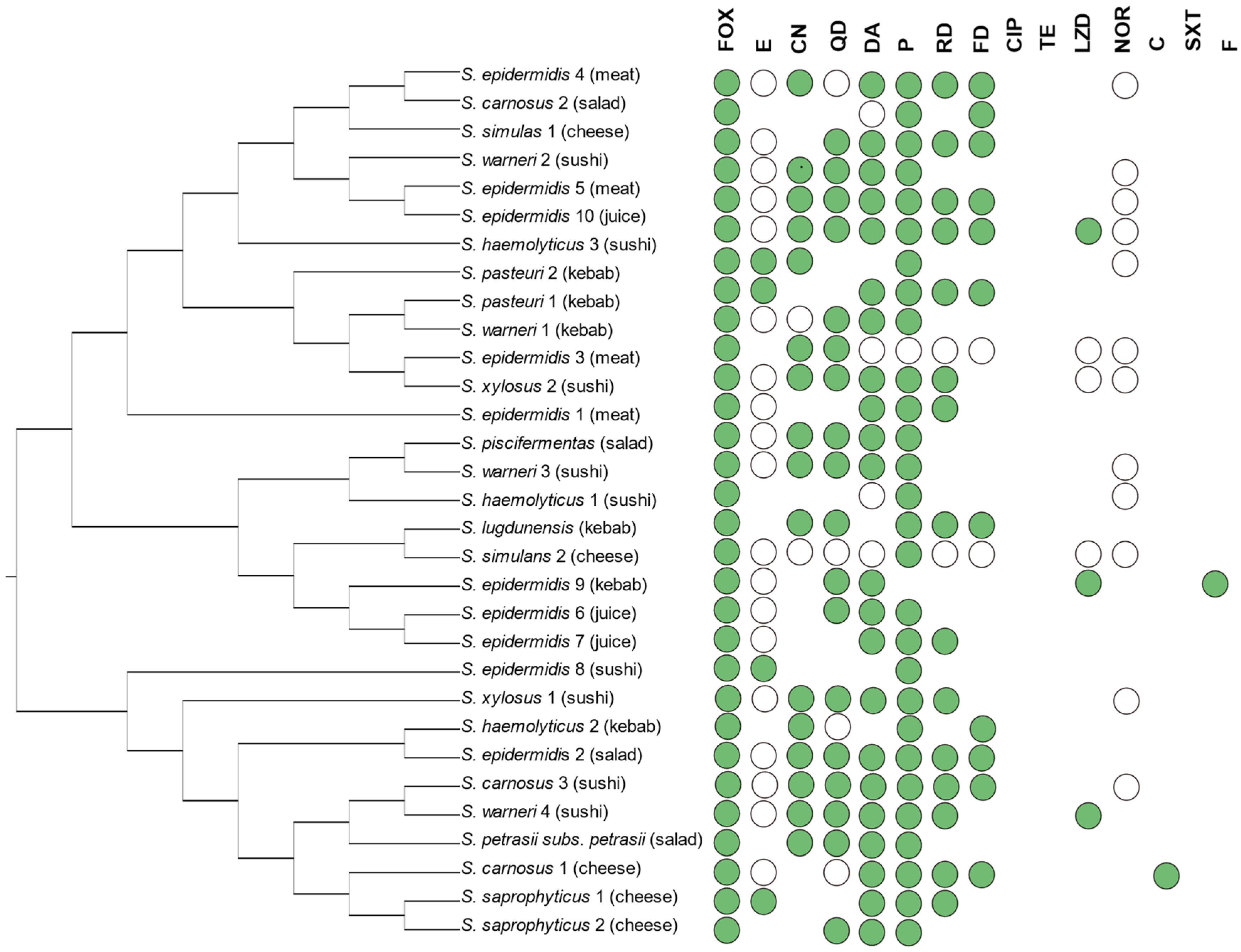
3.1. Occurrence and Phenotypic Resistance in CoNS from Ready-to-Eat
3.2. Genotypic Antimicrobial Resistance in CoNS
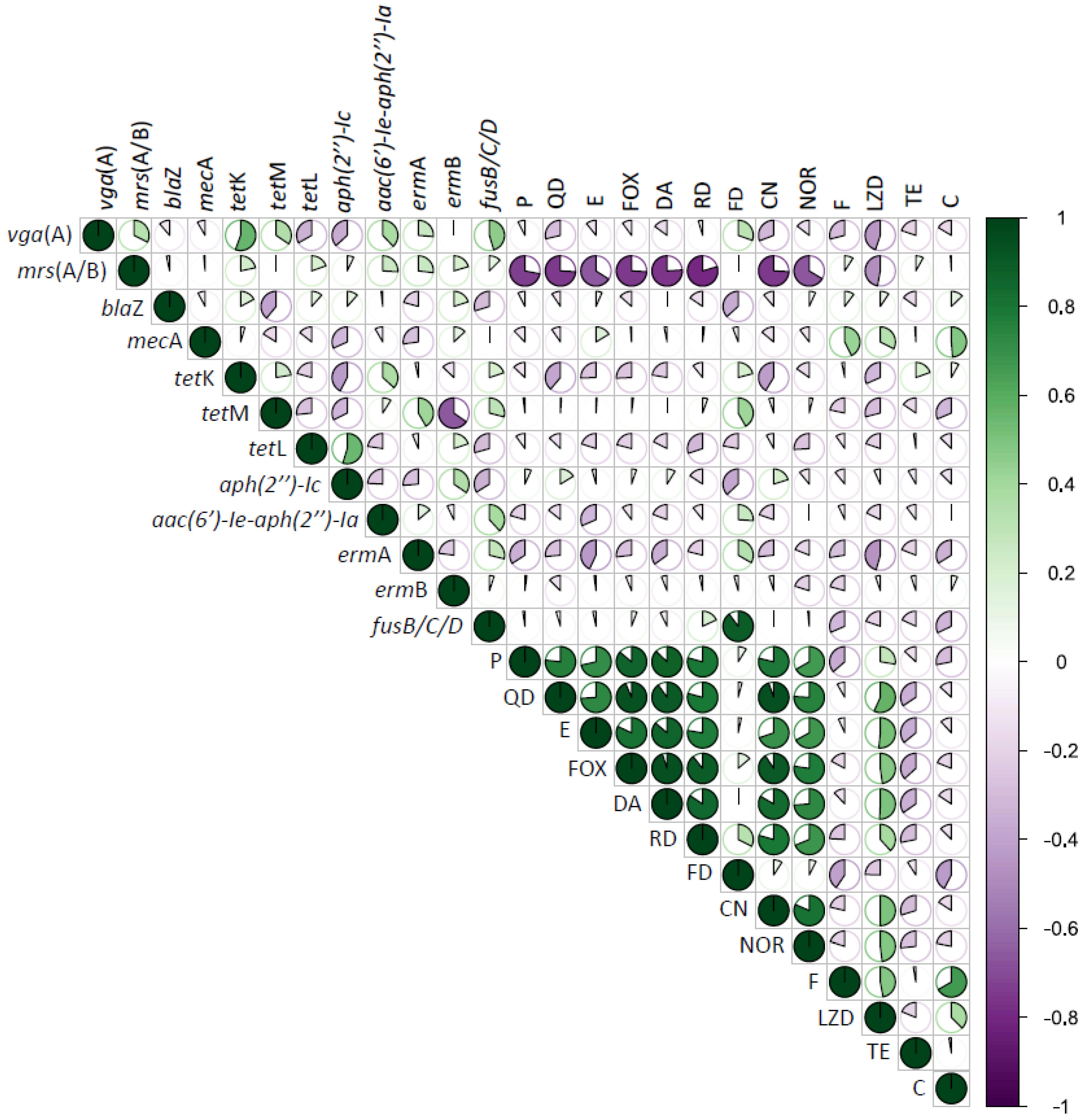
3.3. Correlation between Phenotype and Genotype Antimicrobial Resistance in CoNS from RTE Food
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rehman, M.; Ahmed, S.; Ahmed, U.; Tamanna, K.; Sabir, M.S.; Niaz, Z. An overview of self-medication: A major cause of antibiotic resistance and a threat to global public health. J. Pak. Med. Assoc. 2021, 71, 3–943. [Google Scholar] [CrossRef]
- John, J., Jr. The treatment of resistant staphylococcal infections. F1000Res 2020, 26, 9. [Google Scholar] [CrossRef] [PubMed]
- França, A.; Gaio, V.; Lopes, N.; Melo, L.D.R. Virulence Factors in Coagulase-Negative Staphylococci. Pathogens 2021, 4, 170. [Google Scholar] [CrossRef]
- Pedroso, S.H.; Sandes, S.H.; Filho, R.A.; Nunes, A.C.; Serufo, J.C.; Farias, L.M.; Carvalho, A.R.; Bomfim, R.Q.M.; Santos, S.G. Coagulase-negative Staphylococci isolated from human bloodstream infections showed multidrug resistance profile. Microb. Drug Resist. 2018, 24, 5–635. [Google Scholar] [CrossRef]
- Collignon, P.J.; McEwen, S.A. One Health-Its Importance in Helping to Better Control Antimicrobial Resistance. Trop. Med. Infect. Dis. 2019, 29, 22. [Google Scholar] [CrossRef] [Green Version]
- Sundin, G.W.; Wang, N. Antibiotic Resistance in Plant-Pathogenic Bacteria. Annu. Rev. Phytopathol. 2018, 56, 161–180. [Google Scholar] [CrossRef]
- Pepi, M.; Focardi, S. Antibiotic-Resistant Bacteria in Aquaculture and Climate Change: A Challenge for Health in the Mediterranean Area. Int. J. Environ. Res. Public Health 2021, 26, 5723. [Google Scholar] [CrossRef] [PubMed]
- Aslantaş, Ö.; Yıldırım, N. Isolation of Methicillin Resistant (MR) Staphylococci from Chicken Meat Samples. Harran Üniversitesi Vet. Fakültesi Derg. 2021, 10, 126–131. [Google Scholar] [CrossRef]
- Silva, V.; Caniça, M.; Ferreira, E.; Vieira-Pinto, M.; Saraiva, C.; Pereira, J.E.; Capelo, J.L.; Igrejas, G.; Poeta, P. Multidrug-Resistant Methicillin-Resistant Coagulase-Negative Staphylococci in Healthy Poultry Slaughtered for Human Consumption. Antibiotics 2022, 11, 365. [Google Scholar] [CrossRef]
- Chajęcka-Wierzchowska, W.; Zadernowska, A.; Nalepa, B.; Sierpińska, M.; Łaniewska-Trokenheim, Ł. Retail ready-to-eat food as a potential vehicle for Staphylococcus spp. harboring antibiotic resistance genes. J. Food Prot. 2014, 77, 993–998. [Google Scholar] [CrossRef]
- Chajęcka-Wierzchowska, W.; Zadernowska, A.; Nalepa, B.; Sierpińska, M.; Łaniewska-Trokenheim, Ł. Coagulase-negative staphylococci (CoNS) isolated from ready-to-eat food of animal origin-phenotypic and genotypic antibiotic resistance. Food Microbiol. 2015, 46, 222–226. [Google Scholar] [CrossRef]
- Chajęcka-Wierzchowska, W.; Gajewska, J.; Wiśniewski, P.; Zadernowska, A. Enterotoxigenic Potential of Coagulase-Negative Staphylococci from Ready-to-Eat Food. Pathogens 2020, 9, 734. [Google Scholar] [CrossRef] [PubMed]
- Zakrzewski, A.J.; Zarzecka, U.; Chajęcka-Wierzchowska, W.; Zadernowska, A. A Comparison of Methods for Identifying Enterobacterales Isolates from Fish and Prawns. Pathogens 2022, 11, 410. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Xing, J.; Li, B.; Wang, P.; Liu, J. Use of tuf as a target for sequence-based identification of Gram-positive cocci of the genus Enterococcus, Streptococcus, coagulase-negative Staphylococcus, and Lactococcus. Ann. Clin. Microbiol. Antimicrob. 2012, 27, 11–31. [Google Scholar] [CrossRef] [Green Version]
- Chajęcka-Wierzchowska, W.; Zadernowska, A.; Gajewska, J. S. epidermidis strains from artisanal cheese made from unpasteurized milk in Poland-Genetic characterization of antimicrobial resistance and virulence determinants. Int. J. Food Microbiol. 2019, 294, 55–59. [Google Scholar] [CrossRef]
- Kaase, M.; Lenga, S.; Friedrich, S.; Szabados, F.; Sakinc, T.; Kleine, B.; Gatermann, S.G. Comparison of phenotypic methods for penicillinase detection in Staphylococcus aureus. Clin. Microbiol. Infect. 2008, 14, 614–616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelli, A.; Voidarou, C.; Venardou, B.; Fotou, K.; Tsinas, A.; Bonos, E.; Fthenakis, G.C.; Skoufos, I.; Tzora, A. Antimicrobial and Methicillin Resistance Pattern of Potential Mastitis-Inducing Staphylococcus aureus and Coagulase-Negative Staphylococci Isolates from the Mammary Secretion of Dairy Goats. Biology 2022, 11, 1591. [Google Scholar] [CrossRef] [PubMed]
- Hritcu, O.M.; Schmidt, V.M.; Salem, S.E.; Maciuca, I.E.; Moraru, R.F.; Lipovan, I.; Mareş, M.; Solcan, G.; Timofte, D. Geographical Variations in Virulence Factors and Antimicrobial Resistance Amongst Staphylococci Isolated From Dogs From the United Kingdom and Romania. Front. Vet. Sci. 2020, 7, 1–10. [Google Scholar] [CrossRef]
- Padmasini, E.; Padmaraj, R.; Ramesh, S.S. High level aminoglycoside resistance and distribution of aminoglycoside resistant genes among clinical isolates of Enterococcus species in Chennai, India. Sci. World J. 2014, 4, 329157. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, N.; Mahbub Alam, M.; Nishimoto, Y.; Urasawa, S.; Uehara, N.; Watanabe, N. Distribution of aminoglycoside resistance genes in recent clinical isolates of Enterococcus faecalis, Enterococcus faecium and Enterococcus avium. Epidemiol. Infect. 2001, 126, 197–204. [Google Scholar] [CrossRef]
- Kehrenberg, C.; Cuny, C.; Strommenger, B.; Schwarz, S.; Witte, W. Methicillin-resistant and -susceptible Staphylococcus aureus strains of clonal lineages ST398 and ST9 from swine carry the multidrug resistance gene cfr. Antimicrob. Agents Chemother. 2009, 53, 779–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mališová, L.; Jakubů, V.; Pomorská, K.; Musílek, M.; Žemličková, H. Spread of Linezolid-Resistant Enterococcus spp. in Human Clinical Isolates in the Czech Republic. Antibiotics 2021, 10, 219. [Google Scholar] [CrossRef] [PubMed]
- Brenciani, A.; Morroni, G.; Vincenzi, C.; Manso, E.; Mingoia, M.; Giovanetti, E.; Varaldo, P.E. Detection in Italy of two clinical Enterococcus faecium isolates carrying both the oxazolidinone and phenicol resistance gene optrA and a silent multiresistance gene cfr. J. Antimicrob. Chemother. 2016, 71, 1118–1119. [Google Scholar] [CrossRef] [Green Version]
- Pillai, S.K.; Sakoulas, G.; Wennersten, C.; Eliopoulos, G.M.; Moellering, R.C., Jr.; Ferraro, M.J.; Gold, H.S. Linezolid resistance in Staphylococcus aureus: Characterization and stability of resistant phenotype. J. Infect. Dis. 2002, 1, 1603–1607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Commission Regulation (EC) No 2073/2005 of 15 November 2005 on Microbiological Criteria for Foodstuffs. Available online: https://eur-lex.europa.eu/eli/reg/2005/2073/oj (accessed on 24 November 2022).
- Podkowik, M.; Bystroń, J.; Bania, J. Genotypes, antibiotic resistance and virulence factors of staphylococci from ready-to-eat food. Foodborne Pathog. Dis. 2012, 9, 91–93. [Google Scholar] [CrossRef]
- Al-Khafaji, M.H.M.; Flayyih, M.T. Relationship between methicillin resistance and the presence of femA and mecA genes in coagulase positive and negative staphylococci isolated from milk and cheese. World J. Exp. Biosci. 2015, 3, 50–56. [Google Scholar]
- Hammad, A.M.; Watanabe, W.; Fujii, T.; Shimamoto, T. Occurrence and characteristics of methicillin-resistant and -susceptible Staphylococcus aureus and methicillin-resistant coagulase-negative staphylococci from Japanese retail ready-to-eat raw fish. Int. J. Food Microbiol. 2012, 156, 286–289. [Google Scholar] [CrossRef]
- Stamm, J.; Weißelberg, S.; Both, A.; Failla, A.V.; Nordholt, G.; Büttner, H.; Linder, S.; Aepfelbacher, M.; Rohde, H. Development of an artificial synovial fluid useful for studying Staphylococcus epidermidis joint infections. Front. Cell Infect. Microbiol. 2022, 29, 948151. [Google Scholar] [CrossRef]
- Berends, M.S.; Luz, C.F.; Ott, A.; Andriesse, G.I.; Becker, K.; Glasner, C.; Friedrich, A.W. Trends in Occurrence and Phenotypic Resistance of Coagulase-Negative Staphylococci (CoNS) Found in Human Blood in the Northern Netherlands between 2013 and 2019. Microorganisms 2022, 7, 1801. [Google Scholar] [CrossRef]
- Lee, J.Y.H.; Monk, I.R.; Goncalves Da Silva, A.; Seemann, T.; Chua, K.Y.L.; Kearns, A.; Hill, R.; Woodford, N.; Bartels, M.D.; Strommenger, B.; et al. Global spread of three multidrug-resistant lineages of Staphylococcus epidermidis. Nat. Microbiol. 2018, 3, 1175–1185. [Google Scholar] [CrossRef]
- Seng, R.; Kitti, T.; Thummeepak, R.; Kongthai, P.; Leungtongkam, U.; Wannalerdsakun, S.; Sitthisak, S. Biofilm formation of methicillin-resistant coagulase negative staphylococci (MR-CoNS) isolated from community and hospital environments. PLoS ONE 2017, 31, e0184172. [Google Scholar] [CrossRef] [PubMed]
- Osman, K.; Alvarez-Ordóñez, A.; Ruiz, L.; Badr, J.; ElHofy, F.; Al-Maary, K.S.; Moussa, I.M.I.; Hessain, A.M.; Orabi, A.; Saad, A.; et al. Antimicrobial resistance and virulence characterization of Staphylococcus aureus and coagulase-negative staphylococci from imported beef meat. Ann. Clin. Microbiol. Antimicrob. 2017, 16, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Adawy, H.; Ahmed, M.; Hotzel, H.; Monecke, S.; Schulz, J.; Hartung, J.; Ehricht, R.; Neubauer, H.; Hafez, H.M. Characterization of methicillin-resistant Staphylococcus aureus isolated from healthy turkeys and broilers using DNA microarrays. Front. Microbiol. 2016, 19, 2019. [Google Scholar] [CrossRef]
- Kumar, L.R.G.; Kasim, A.K.; Lekshmi, M.; Nayak, B.B.; Kumar, S. Incidence of methicillin-resistant staphylococci in fresh seafood. Adv. Microbiol. 2016, 06, 399–406. [Google Scholar] [CrossRef] [Green Version]
- Chenouf, N.S.; Mama, O.M.; Messaï, C.R.; Ruiz-Ripa, L.; Fernández-Fernández, R.; Carvalho, I.; Zitouni, A.; Hakem, A.; Torres, C. Detection of methicillin-resistant coagulase-negative staphylococci and PVL/mecA genes in cefoxitin-susceptible Staphylococcus aureus (t044/ST80) from unpasteurized milk sold in stores in Djelfa, Algeria. J. Dairy Sci. 2021, 104, 2684–2692. [Google Scholar] [CrossRef]
- Sergelidis, D.; Abrahim, A.; Papadopoulos, T.; Soultos, N.; Martziou, E.; Koulourida, V.; Govaris, A.; Pexara, A.; Zdragas, A.; Papa, A. Isolation of methicillin-resistant Staphylococcus spp. from ready-to-eat fish products. Lett. Appl. Microbiol. 2014, 59, 500–506. [Google Scholar] [CrossRef]
- Pyzik, E.; Marek, A.; Stȩpień-Pyśniak, D.; Urban-Chmiel, R.; Jarosz, L.S.; Jagiełło-Podȩbska, I. Detection of antibiotic resistance and classical enterotoxin genes in coagulase-negative staphylococci isolated from poultry in Poland. J. Vet. Res. 2019, 63, 183–190. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Lin, J.; Li, L.; Cai, W.; Ye, J.; He, S.; Zhang, W.; Liu, N.; Gong, Z.; Ye, X.; et al. Methicillin-Resistant Coagulase-Negative Staphylococci Carriage is a Protective Factor of Methicillin-Resistant Staphylococcus Aureus Nasal Colonization in HIV-Infected Patients: A Cross-Sectional Study. Can. J. Infect. Dis. Med. Microbiol. 2021, 12, 5717413. [Google Scholar] [CrossRef]
- Teeraputon, S.; Santanirand, P.; Wongchai, T.; Songjang, W.; Lapsomthob, N.; Jaikrasun, D.; Toonkaew, S.; Tophon, P. Prevalence of methicillin resistance and macrolide-lincosamide-streptogramin B resistance in Staphylococcus haemolyticus among clinical strains at a tertiary-care hospital in Thailand. New Microbes New Infect. 2017, 24, 28–33. [Google Scholar] [CrossRef]
- Bishr, A.; Abdelaziz, S.; Yahia, I.; Yassien, M.; Hassouna, N.; Aboshanab, K. Association of macrolide resistance genotypes and synergistic antibiotic combinations for combating macrolide-resistant MRSA recovered from hospitalized patients. Biology 2021, 10, 624. [Google Scholar] [CrossRef]
- Zmantar, T.; Kouidhi, B.; Miladi, H.; Bakhrouf, A. Detection of macrolide and disinfectant resistance genes in clinical Staphylococcus aureus and coagulase-negative staphylococci. BMC Res. Notes 2011, 4, 453. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Iwasaki, T.; Ohtani, S.; Matsui, H.; Nejima, R.; Mori, Y.; Kagaya, F.; Yagi, A.; Yoshimura, A.; Hanaki, H.; et al. Benzalkonium chloride resistance in Staphylococcus epidermidis on the ocular surface of glaucoma patients under long-term administration of eye drops. Transl. Vis. Sci. Technol. 2020, 7, 9. [Google Scholar] [CrossRef] [PubMed]
- Hwang, D.; Kim, S.M.; Kim, H.J. Modelling of tetracycline resistance gene transfer by commensal Escherichia coli food isolates that survived in gastric fluid conditions. Int. J. Antimicrob. Agents 2017, 49, 81–87. [Google Scholar] [CrossRef]
- Li, J.; Liu, D.; Tian, X.; Koseki, S.; Chen, S.; Ye, X.; Ding, T. Novel antibacterial modalities against methicillin 449 resistant Staphylococcus aureus derived from plants. Crit. Rev. Food Sci. Nutr. 2019, 59, S153–S161. [Google Scholar] [CrossRef] [PubMed]
- Soonthornchaikul, N.; Garelick, H. Antimicrobial resistance of Campylobacter species isolated from edible bivalve 447 molluscs purchased from Bangkok markets, Thailand. Foodborne Pathog. Dis. 2009, 6, 947–951. [Google Scholar] [CrossRef] [PubMed]
- Michalik, M.; Kosecka-Strojek, M.; Wolska, M.; Samet, A. First case of staphylococci carrying linezolid resistance genes from laryngological infections in Poland. Pathogens 2021, 13, 335. [Google Scholar] [CrossRef]
- Mittal, G.; Bhandari, V.; Gaind, R.; Rani, V.; Chopra, S.; Dawar, R.; Sardana, R.; Verma, P.K. Linezolid resistant coagulase negative staphylococci (LRCoNS) with novel mutations causing blood stream infections (BSI) in India. BMC Infect. Dis. 2019, 14, 717. [Google Scholar] [CrossRef] [Green Version]
- Musumeci, R.; Calaresu, E.; Gerosa, J.; Oggioni, D.; Bramati, S.; Morelli, P.; Mura, I.; Piana, A.; Are, B.M.; Cocuzza, C.E. Resistance to linezolid in Staphylococcus spp. clinical isolates associated with ribosomal binding site modifications: Novel mutation in domain V of 23S rRNA. New Microbiol. 2016, 39, 269–273. [Google Scholar]
- Barski, P.; Piechowicz, L.; Galiński, J.; Kur, J. Rapid assay for detection of methicillin-resistant staphylococcus aureus using multiplex PCR. Mol. Cell. Probes 1996, 10, 471–475. [Google Scholar] [CrossRef]
- Gevers, D.; Danielsen, M.; Huys, G.; Swings, J. Molecular characterization of tet(M) genes in Lactobacillus isolates from different types of fermented dry sausage. Appl. Environ. Microbiol. 2003, 69, 1270–1275. [Google Scholar] [CrossRef] [Green Version]
- Roberts, M.C. Epidemiology of tetracycline-resistance determinants. Trends Microbiol. 1994, 2, 353–357. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Species | No. of Isolates (%) | Antimicrobial Resistance | |
|---|---|---|---|
| Phenotype | Genotype | ||
| S. epidermidis | 21 (24.7%) | CN(4), DA(9), E(16), FOX(10), FD(6), LZD(2), NOR(3), P(13), RD(6), QD(5) | aac(6′)-Ie-aph(2″)-Ia(11), blaZ(17), ermA(8), ermB(11), fusB/C/D(5), mecA(11), mph(C)(3), msr(A/B)(15), tetK(7), tetL(2), tetM(3), vga(A)(7) |
| S. warneri | 14 (16.5%) | C(1), CN(4), DA(5), E(4), FOX(4), FD(2), LZD(1), NOR(1), P(8),TE(1), RD(2), QD(4) | aac(6′)-Ie-aph(2″)-Ia(7), blaZ(13), ermB(3), fusB/C/D(2), mecA(1), msr(A/B)(6), tetK(6), tetM(1), vga(A)(4) |
| S. carnosus | 9 (10.6%) | CN(1), DA(3), E(3), FOX(3), FD(4), NOR(1), P(3), RD(2), QD(3) | aac(6′)-Ie-aph(2″)-Ia(5), aph(2″)-Ic(1), blaZ(9), ermA(3), ermB(6), fusB/C/D(2), mecA(3), mph(C)(1), msr(A/B)(8), tetK(5), tetM(2), vga(A)(3) |
| S. simulans | 9 (10.6%) | CN(1), DA(1), E(1), FOX(2), FD(3), P(4), RD(2), TE(1), QD(1) | aac(6′)-Ie-aph(2″)-Ia(3), blaZ(6), ermA(5), ermB(1), fusB/C/D(3), msr(A/B)(6), tetK(1), tetM(2), vga(A)(5) |
| S. xylosus | 8 (9.4%) | CN(2), DA(3), E(2), FOX(2), FD(1), LZD(1), NOR(2), P(3), RD(1), QD(2) | aac(6′)-Ie-aph(2″)-Ia(5), blaZ(8), ermA(4), ermB(3), fusB/C/D(1), mecA(1), msr(A/B)(5), vga(A)(2) |
| S. saprophyticus | 6 (7.1%) | DA(3), E(3), FOX(2), FD(2), P(4), RD(1), QD(1) | aac(6′)-Ie-aph(2″)-Ia(2), blaZ(4), ermA(3), mecA(1), msr(A/B)(5), tetK(3), tetM(5), vga(A)(2) |
| S. pasteuri | 5 (5.9%) | CN(1), DA(2), E(3), FOX(2), FD(2), P(5),, RD(1), QD(1) | blaZ(5), ermA(2), ermB(2), mecA(2), msr(A/B)(2), tetK(2), tetM(1) |
| S. heamolyticus | 4 (4.7%) | CN(2), DA(1), E(1), FOX(3), FD(2), NOR(2), P(4), TE(1), QD(1) | aac(6′)-Ie-aph(2″)-Ia(2), blaZ(3), ermA(2), ermB(2), mecA(3), msr(A/B)(4), tetK(1), |
| S. petrasii subsp. petrasii | 4 (4.7%) | C(2), CN(1), DA(1), F(1), FOX(1), P(1), QD(1) | aac(6′)-Ie-aph(2″)-Ia(2), aph(2″)-Ic(1), blaZ(4), ermB(3), mecA(2), msr(A/B)(4), tetK(2) |
| S. lentus | 2 (2.4%) | P(1) | aac(6′)-Ie-aph(2″)-Ia(1), blaZ(2), ermA(1), ermB(1), mecA(1), msr(A/B)(1) |
| S. piscifermentas | 2 (2.4%) | CN(1), DA(1), E(1), FOX(1), FD(1), P(1), QD(1) | aph(2″)-Ic(1), blaZ(1), ermB(2), fusB/C/D(1), msr(A/B)(2) |
| S. lugdenensis | 1 (1.2%) | CN(1), FOX(1), FD(1), P(1), QD(1), RD(1), | ermA(1) |
| Total | 85 | C(3), CN(19), DA(29), E(34), F(1), FD(24), FOX(31), LZD(4), NOR(9), P(48), QD(21), RD(16), TE(3) | aac(6′)-Ie-aph(2″)-Ia(39), aph(2″)-Ic(3), blaZ(72), ermA(29), ermB(34), fusB/C/D(15), mecA(25), mph(C)(4), msr(A/B)(58), tetK(27), tetL(2), tetM(14), vga(A)(26) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chajęcka-Wierzchowska, W.; Gajewska, J.; Zadernowska, A.; Randazzo, C.L.; Caggia, C. A Comprehensive Study on Antibiotic Resistance among Coagulase-Negative Staphylococci (CoNS) Strains Isolated from Ready-to-Eat Food Served in Bars and Restaurants. Foods 2023, 12, 514. https://doi.org/10.3390/foods12030514
Chajęcka-Wierzchowska W, Gajewska J, Zadernowska A, Randazzo CL, Caggia C. A Comprehensive Study on Antibiotic Resistance among Coagulase-Negative Staphylococci (CoNS) Strains Isolated from Ready-to-Eat Food Served in Bars and Restaurants. Foods. 2023; 12(3):514. https://doi.org/10.3390/foods12030514
Chicago/Turabian StyleChajęcka-Wierzchowska, Wioleta, Joanna Gajewska, Anna Zadernowska, Cinzia Lucia Randazzo, and Cinzia Caggia. 2023. "A Comprehensive Study on Antibiotic Resistance among Coagulase-Negative Staphylococci (CoNS) Strains Isolated from Ready-to-Eat Food Served in Bars and Restaurants" Foods 12, no. 3: 514. https://doi.org/10.3390/foods12030514