
Visualizing the Distribution of Lipids in Peanut Seeds by MALDI Mass Spectrometric Imaging
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
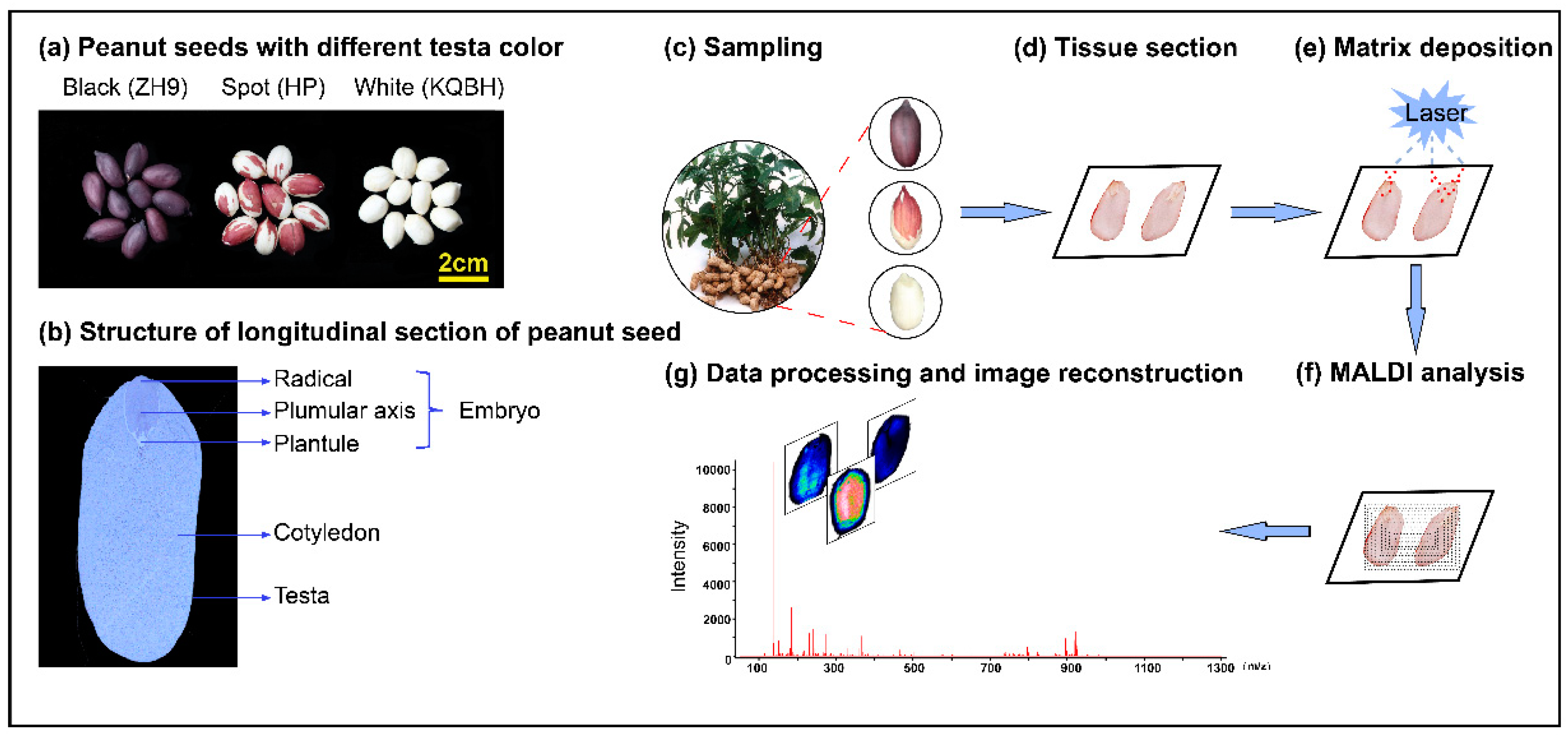
2.1. Reagents and Samples
2.2. Peanut Sample Section and Spray
2.3. MALDI-MSI Parameters
2.4. Data Analysis
3. Results and Discussion
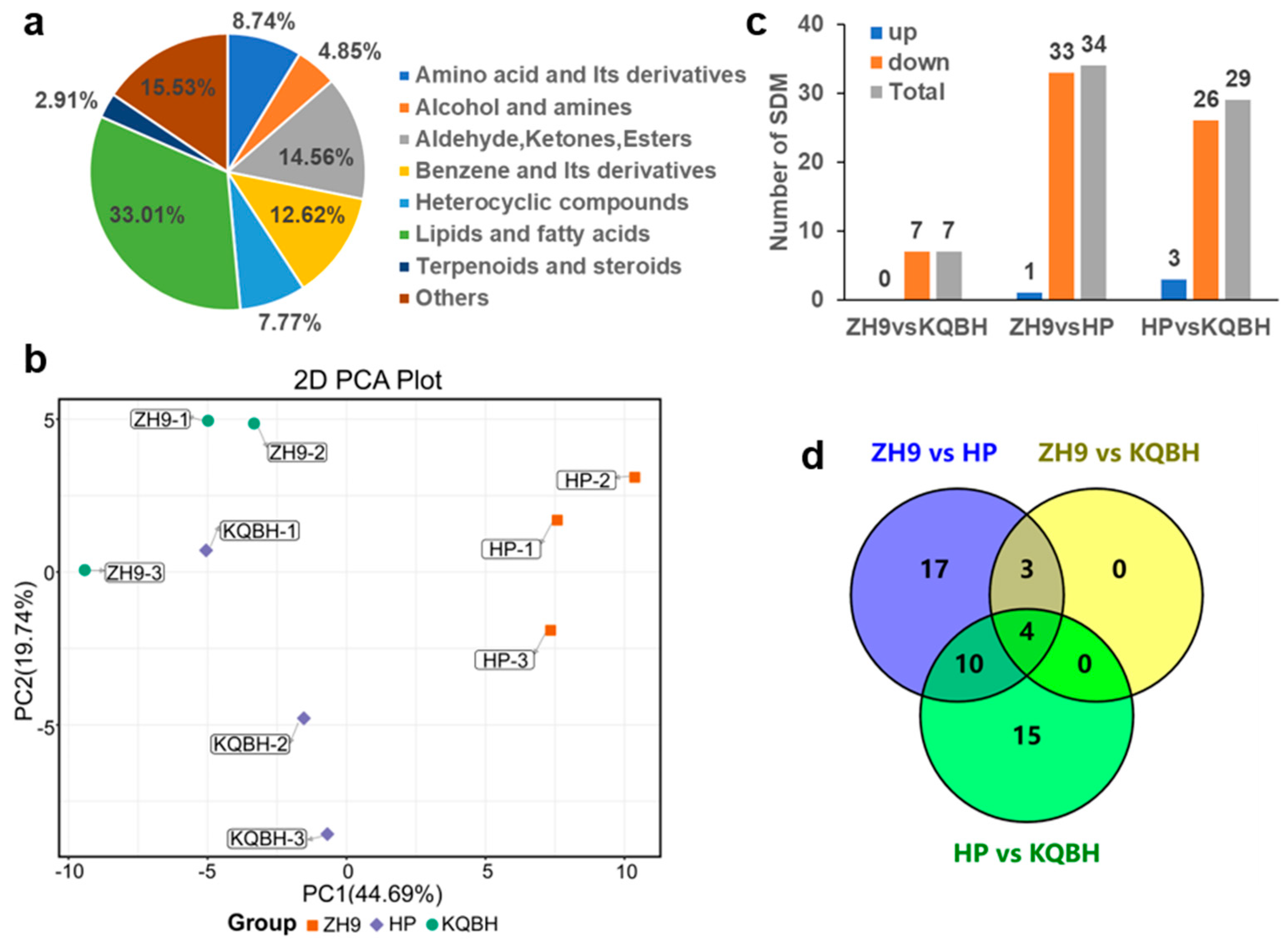
3.1. Identification of Differential Metabolites in Peanut Seeds by MALDI-MSI
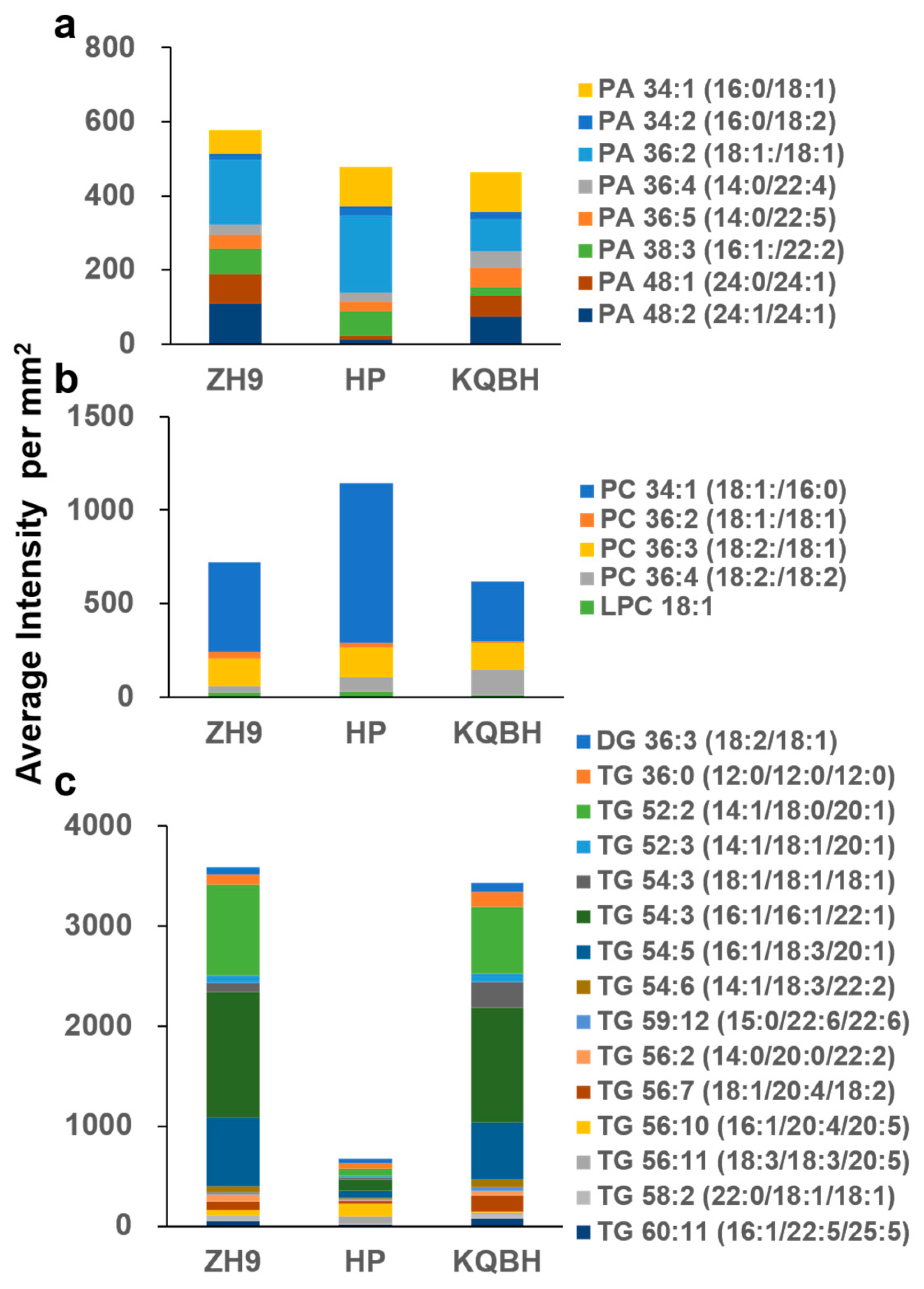
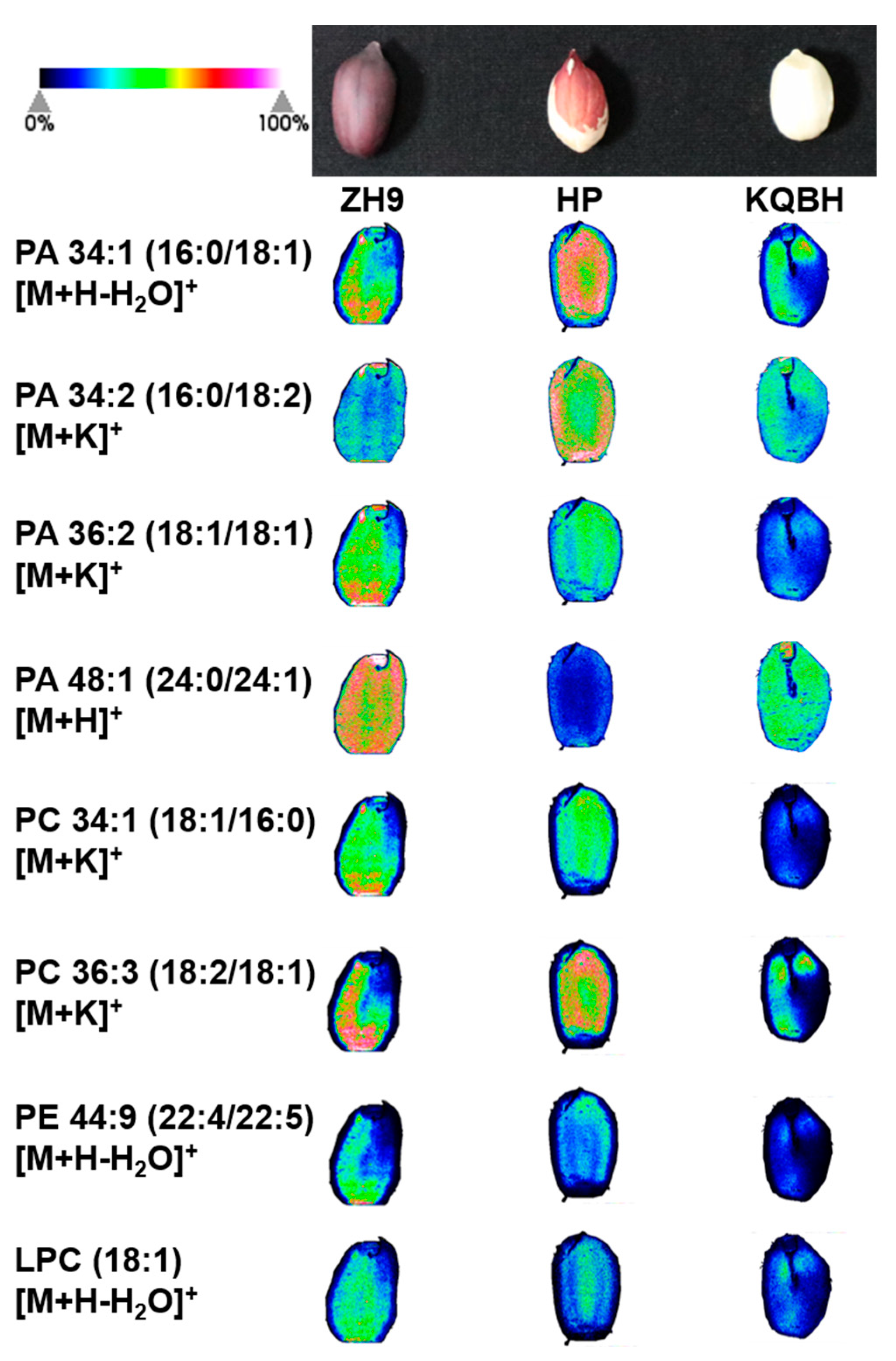
3.2. Spatial Distribution of Glycerophospholipids
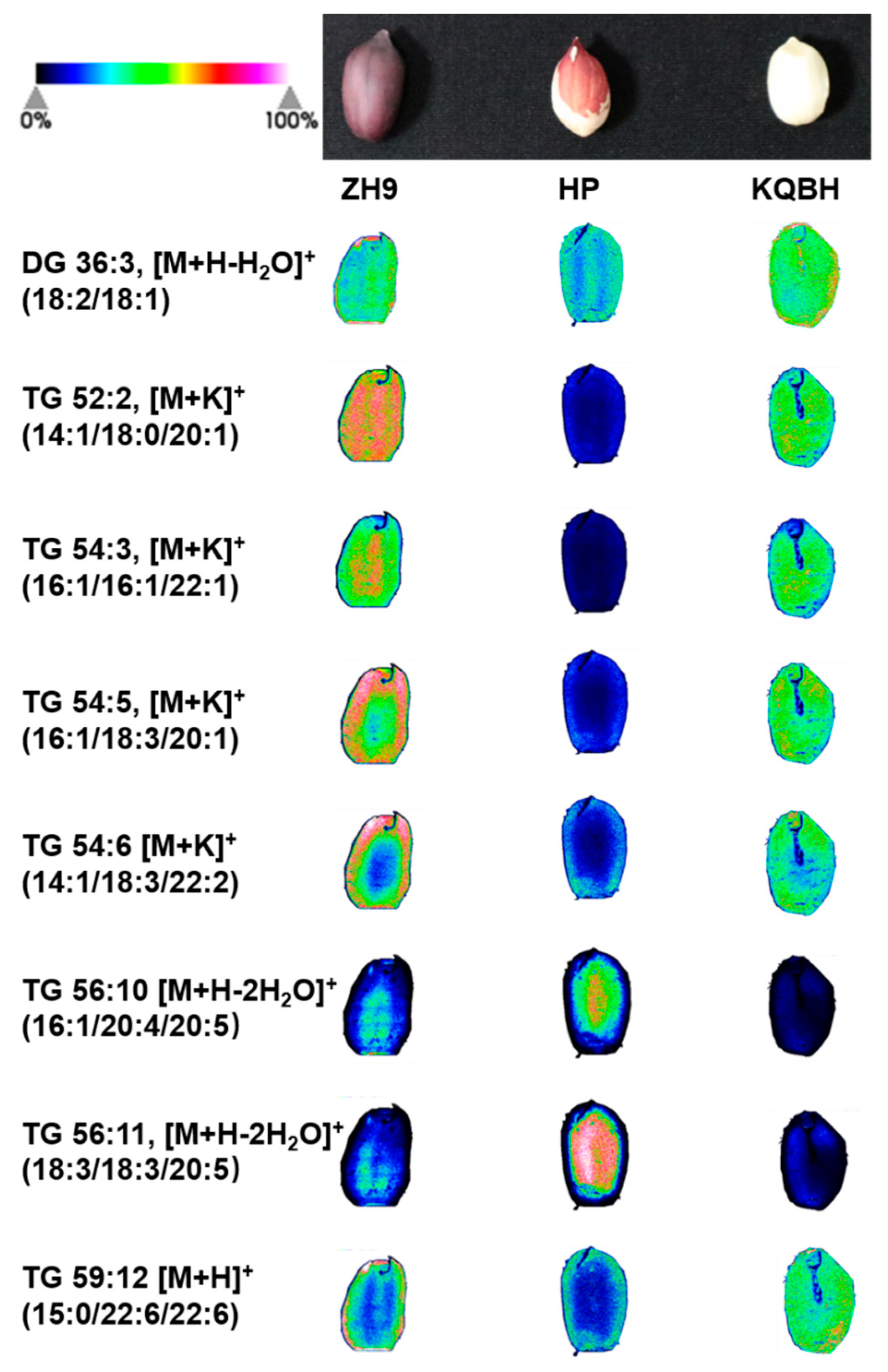
3.3. Spatial Distribution of Glycerolipids
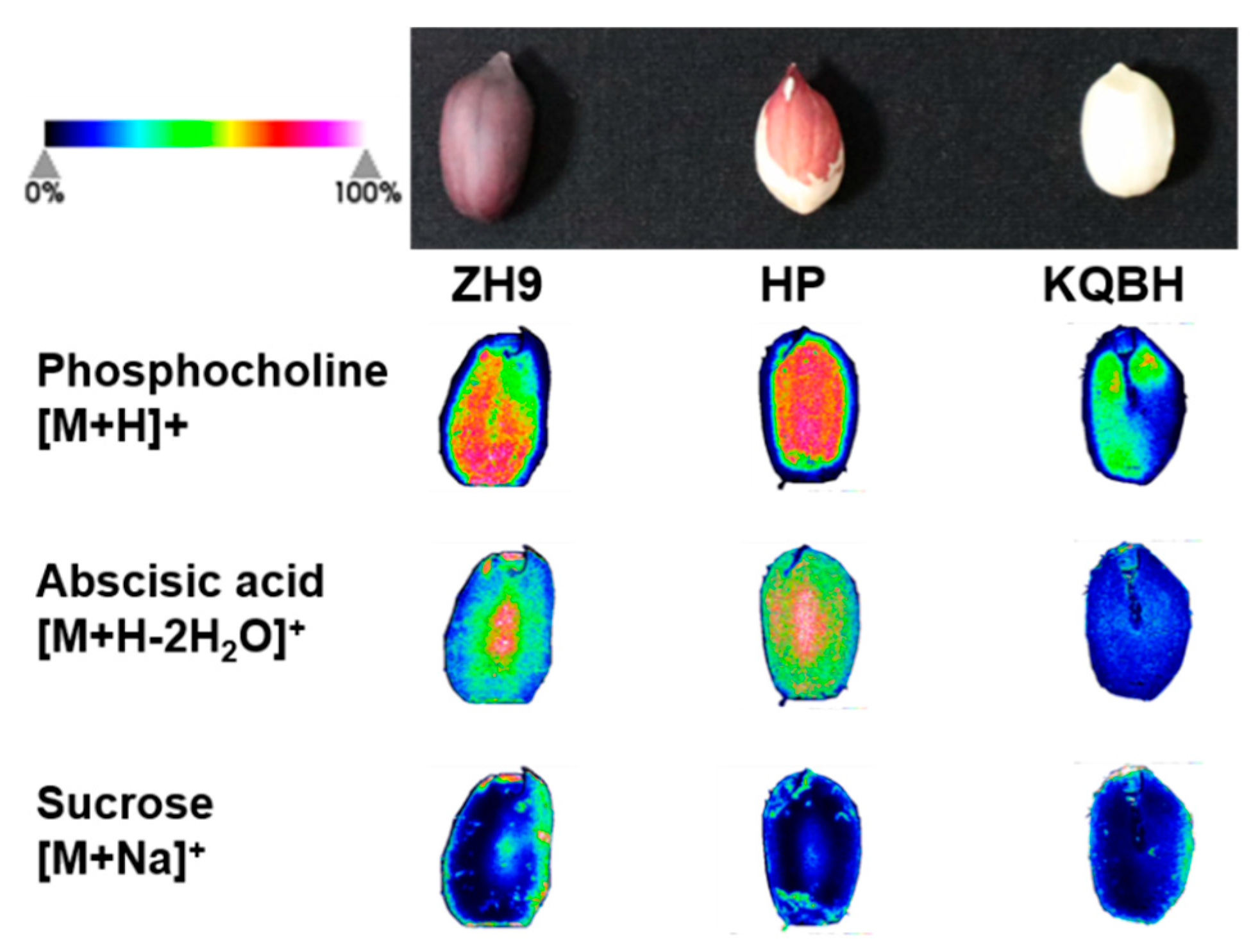
3.4. Spatial Distribution of Other Key Metabolites
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Arya, S.S.; Salve, A.R.; Chauhan, S. Peanuts as functional food: A review. J. Food Sci. Technol. 2016, 53, 31–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Sen, G.; Ho, C.-T.; Bai, N. Review on chemical compositions and biological activities of peanut (Arachis hypogeae L.). J. Food Biochem. 2022, 46, e14119. [Google Scholar] [CrossRef]
- Nawade, B.; Mishra, G.P.; Radhakrishnan, T.; Dodia, S.M.; Ahmad, S.; Kumar, A.; Kumar, A.; Kundu, R. High oleic peanut breeding: Achievements, perspectives, and prospects. Trends Food Sci. Technol. 2018, 78, 107–119. [Google Scholar] [CrossRef]
- Peng, J.; Jia, Y.; Du, X.; Wang, Y.; Yang, Z.; Li, K. Study of physicochemical stability of anthocyanin extracts from black peanut skin and their digestion enzyme and adipogenesis inhibitory activities. LWT-Food Sci. Technol. 2019, 107, 107–116. [Google Scholar] [CrossRef]
- Li, K.; Zhang, M.; Chen, H.; Peng, J.; Jiang, F.; Shi, X.; Bai, Y.; Jian, M.; Jia, Y. Anthocyanins from black peanut skin protect against UV-B induced keratinocyte cell and skin oxidative damage through activating Nrf 2 signaling. Food Funct. 2019, 10, 6815–6828. [Google Scholar] [CrossRef] [PubMed]
- Kuang, Q.; Yu, Y.; Attree, R.; Xu, B. A comparative study on anthocyanin, saponin, and oil profiles of black and red seed coat peanut (Arachis hypogacea) grown in China. Int. J. Food Prop. 2017, 20, S131–S140. [Google Scholar] [CrossRef] [Green Version]
- Attree, R.; Du, B.; Xu, B. Distribution of phenolic compounds in seed coat and cotyledon, and their contribution to antioxidant capacities of red and black seed coat peanuts (Arachis hypogaea L.). Ind. Crops Prod. 2015, 67, 448–456. [Google Scholar] [CrossRef]
- Shem-Tov, Y.; Badani, H.; Segev, A.; Hedvat, I.; Galili, S.; Hovav, R. Determination of total polyphenol, flavonoid and anthocyanin contents and antioxidant capacities of skins from peanut (Arachis hypogaea) lines with different skin colors. J. Food Biochem. 2012, 36, 301–308. [Google Scholar] [CrossRef]
- Liu, H.; Hong, Y.B.; Lu, Q.; Li, H.F.; Gu, J.Z.; Ren, L.; Deng, L.; Zhou, B.J.; Chen, X.P.; Liang, X.Q. Integrated analysis of comparative lipidomics and proteomics reveals the dynamic changes of lipid molecular species in high-oleic acid peanut seed. J. Agric. Food Chem. 2020, 68, 426–438. [Google Scholar] [CrossRef] [PubMed]
- Heyman, H.M.; Dubery, I.A. The potential of mass spectrometry imaging in plant metabolomics: A review. Phytochem. Rev. 2016, 15, 297–316. [Google Scholar] [CrossRef]
- Boughton, B.A.; Thinagaran, D.; Sarabia, D.; Bacic, A.; Roessner, U. Mass spectrometry imaging for plant biology: A review. Phytochem. Rev. 2016, 15, 445–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshimura, Y.; Zaima, N. Application of Mass Spectrometry Imaging for Visualizing Food Components. Foods 2020, 9, 575. [Google Scholar] [CrossRef]
- Hou, J.; Zhang, Z.; Zhang, L.; Wu, W.; Huang, Y.; Jia, Z.; Zhou, L.; Gao, L.; Long, H.; Lei, M.; et al. Spatial lipidomics of eight edible nuts by desorption electrospray ionization with ion mobility mass spectrometry imaging. Food Chem. 2022, 371, 130893. [Google Scholar] [CrossRef] [PubMed]
- Sagara, T.; Bhandari, D.R.; Spengler, B.; Vollmann, J. Spermidine and other functional phytochemicals in soybean seeds: Spatial distribution as visualized by mass spectrometry imaging. Food Sci. Nutr. 2020, 8, 675–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, S.; Sturtevant, D.; Aziz, M.; Jin, C.; Li, Q.; Chapman, K.D.; Guo, L. Spatial analysis of lipid metabolites and expressed genes reveals tissue-specific heterogeneity of lipid metabolism in high- and low-oil Brassica napus L. seeds. Plant J. 2018, 94, 915–932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woodfield, H.K.; Sturtevant, D.; Borisjuk, L.; Munz, E.; Guschina, I.A.; Chapman, K.; Harwood, J.L. Spatial and temporal mapping of key lipid species in brassica napus seeds. Plant Physiol. 2017, 173, 1998–2009. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Yang, E.; Chaurand, P.; Raghavan, V. Visualizing the distribution of strawberry plant metabolites at different maturity stages by MALDI-TOF imaging mass spectrometry. Food Chem. 2021, 345, 128838. [Google Scholar] [CrossRef]
- Yoshimura, Y.; Goto-Inoue, N.; Moriyama, T.; Zaima, N. Significant advancement of mass spectrometry imaging for food chemistry. Food Chem. 2016, 210, 200–211. [Google Scholar] [CrossRef]
- Huang, Y.T.; Ma, R.; Xu, Y.J.; Zhong, K.; Bu, Q.; Gao, H. A Comparison of lipid contents in different types of peanut cultivars using UPLC-Q-TOF-MS-based lipidomic study. Foods 2022, 11, 4. [Google Scholar] [CrossRef]
- Sarabia, L.D.; Boughton, B.A.; Rupasinghe, T.; van de Meene, A.M.L.; Callahan, D.L.; Hill, C.B.; Roessner, U. High-mass-resolution MALDI mass spectrometry imaging reveals detailed spatial distribution of metabolites and lipids in roots of barley seedlings in response to salinity stress. Metabolomics 2018, 14, 63. [Google Scholar] [CrossRef]
- Reszczynska, E.; Hanaka, A. Lipids composition in plant membranes. Cell Biochem. Biophys. 2020, 78, 401–414. [Google Scholar] [CrossRef] [PubMed]
- Colin, L.A.; Jaillais, Y. Phospholipids across scales: Lipid patterns and plant development. Curr. Opin. Plant Biol. 2020, 53, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.U. Lipid Metabolism in Plants. Plants 2020, 9, 871. [Google Scholar] [CrossRef] [PubMed]
- Bates, P.D.; Stymne, S.; Ohlrogge, J. Biochemical pathways in seed oil synthesis. Curr. Opin. Plant Biol. 2013, 16, 358–364. [Google Scholar] [CrossRef] [Green Version]
- Horn, P.J.; Korte, A.R.; Neogi, P.B.; Love, E.; Fuchs, J.; Strupat, K.; Borisjuk, L.; Shulaev, V.; Lee, Y.J.; Chapman, K.D. Spatial mapping of lipids at cellular resolution in embryos of cotton. Plant Cell 2012, 24, 622–636. [Google Scholar] [CrossRef] [Green Version]
- Horn, P.J.; Silva, J.E.; Anderson, D.; Fuchs, J.; Borisjuk, L.; Nazarenus, T.J.; Shulaev, V.; Cahoon, E.B.; Chapman, K.D. Imaging heterogeneity of membrane and storage lipids in transgenic Camelina sativa seeds with altered fatty acid profiles. Plant J. 2013, 76, 138–150. [Google Scholar] [CrossRef]
- Sturtevant, D.; Dueñas, M.E.; Lee, Y.J.; Chapman, K.D. Three-dimensional visualization of membrane phospholipid distributions in Arabidopsis thaliana seeds: A spatial perspective of molecular heterogeneity. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2017, 1862, 268–281. [Google Scholar] [CrossRef] [Green Version]
- Cox, K.L., Jr.; Gurazada, S.G.R.; Duncan, K.E.; Czymmek, K.J.; Topp, C.N.; Meyers, B.C. Organizing your space: The potential for integrating spatial transcriptomics and 3D imaging data in plants. Plant Physiol. 2022, 188, 703–712. [Google Scholar] [CrossRef]
- Rao, A.; Barkley, D.; Franca, G.S.; Yanai, I. Exploring tissue architecture using spatial transcriptomics. Nature 2021, 596, 211–220. [Google Scholar] [CrossRef]
- Lin, Y.C.; Liu, Y.C.; Nakamura, Y. The choline/ethanolamine kinase family in Arabidopsis: Essential role of CEK4 in phospholipid biosynthesis and embryo development. Plant Cell 2015, 27, 1497–1511. [Google Scholar] [CrossRef]
- Huo, K.S.; Shui, L.Y.; Mai, Y.T.; Zhou, N.; Liu, Y.; Zhang, C.X.; Niu, J. Effects of exogenous abscisic acid on oil content, fatty acid composition, biodiesel properties and lipid components in developing Siberian apricot (Prunus sibirica) seeds. Plant Physiol. Biochem. 2020, 154, 260–267. [Google Scholar] [CrossRef]
- Penfield, S.; Li, Y.; Gilday, A.D.; Graham, S.; Graham, I.A. Arabidopsis ABA INSENSITIVE4 regulates lipid mobilization in the embryo and reveals repression of seed germination by the endosperm. Plant Cell 2006, 18, 1887–1899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bishi, S.K.; Kumar, L.; Dagla, M.C.; Mahatma, M.K.; Rathnakumar, A.L.; Lalwani, H.B.; Misra, J.B. Characterization of Spanish peanut germplasm (Arachis hypogaea L.) for sugar profiling and oil quality. Ind. Crops Prod. 2013, 51, 46–50. [Google Scholar] [CrossRef]
- Garrett, R.; Rezende, C.M.; Ifa, D.R. Revealing the spatial distribution of chlorogenic acids and sucrose across coffee bean endosperm by desorption electrospray ionization-mass spectrometry imaging. LWT-Food Sci. Technol. 2016, 65, 711–717. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Types | m/za | Score b | Adduct | Formula | Putative Compound c |
|---|---|---|---|---|---|
| DG | 618.5223 | 0.98 | M+H-H2O | C39H70O5 | DG 36:3 (18:2/18:1) |
| TG | 638.5485 | 0.97 | M+H-2H2O | C39H74O6 | TG 36:0 (12:0/12:0/12:0) |
| 896.6894 | 0.73 | M+H-2H2O | C59H92O6 | TG 56:11 (18:3/18:3/20:5) | |
| 898.7050 | 0.68 | M+H-2H2O | C59H94O6 | TG 56:10 (16:1/20:4/20:5) | |
| 856.7520 | 0.98 | M+Na | C55H100O6 | TG 52:3 (14:1/18:1/20:1) | |
| 858.7676 | 0.87 | M+K | C55H102O6 | TG 52:2 (14:1/18:0/20:1) | |
| 952.7520 | 0.52 | M+Na+HCOOH | C63H100O6 | TG 60:11 (16:1/22:5/25:5) | |
| 904.7520 | 0.58 | M+K+HCOOH | C59H100O6 | TG 56:7 (18:1/20:4/18:2) | |
| 884.7833 | 0.79 | M+Na | C57H104O6 | TG 54:3 (18:1/18:1/18:1) | |
| 878.7363 | 0.89 | M+K | C57H98O6 | TG 54:6 (14:1/18:3/22:2) | |
| 880.7520 | 0.96 | M+K | C57H100O6 | TG 54:5 (16:1/18:3/20:1) | |
| 884.7833 | 0.98 | M+K | C57H104O6 | TG 54:3 (16:1/16:1/22:1) | |
| 936.7207 | 1.00 | M+H | C62H96O6 | TG 59:12 (15:0/22:6/22:6) | |
| 914.8302 | 0.99 | M+K | C59H110O6 | TG 56:2 (14:0/20:0/22:2) | |
| 942.8615 | 0.98 | M+K | C61H114O6 | TG 58:2 (22:0/18:1/18:1) | |
| PA | 672.4730 | 0.97 | M+K | C37H69O8P | PA 34:2 (16:0/18:2) |
| 694.4574 | 0.92 | M+K | C39H67O8P | PA 36:5 (14:0/22:5) | |
| 696.4730 | 0.95 | M+K | C39H69O8P | PA 36:4 (14:0/22:4) | |
| 754.4550 | 0.98 | M+H-H2O | C37H72O11P2 | PA 34:1 (16:0/18:1) | |
| 700.5043 | 0.89 | M+K | C39H73O8P | PA 36:2 (18:1:/18:1) | |
| 726.5200 | 0.98 | M+K | C41H75O8P | PA 38:3 (16:1:/22:2) | |
| 868.6921 | 0.99 | M+H | C51H97O8P | PA 48:2 (24:1/24:1) | |
| 870.7078 | 0.99 | M+H | C51H99O8P | PA 48:1 (24:0/24:1) | |
| PC | 759.5778 | 0.94 | M+K | C42H82NO8P | PC 34:1 (18:1:/16:0) |
| 785.5935 | 0.98 | M+Na | C44H84NO8P | PC 36:2 (18:1:/18:1) | |
| 781.5622 | 0.99 | M+K | C44H80NO8P | PC 36:4 (18:2:/18:2) | |
| 783.5778 | 0.96 | M+K | C44H82NO8P | PC 36:3 (18:2:/18:1) | |
| PE | 841.5622 | 0.95 | M+H-H2O | C49H80NO8P | PE 44:9 (22:4/22:5) |
| LPC | 521.3481 | 0.86 | M+H-H2O | C26H52NO7P | LPC 18:1 |
| Other | 183.0660 | 0.91 | M+H | C5H14NO4P | Phosphocholine |
| 264.1362 | 0.80 | M+H-2H2O | C15H20O4 | Abscisic acid | |
| 342.1162 | 0.92 | M+Na | C12H22O11 | Sucrose |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Chen, Y.; Liu, Y.; Ouyang, L.; Yao, R.; Wang, Z.; Kang, Y.; Yan, L.; Huai, D.; Jiang, H.; et al. Visualizing the Distribution of Lipids in Peanut Seeds by MALDI Mass Spectrometric Imaging. Foods 2022, 11, 3888. https://doi.org/10.3390/foods11233888
Wang X, Chen Y, Liu Y, Ouyang L, Yao R, Wang Z, Kang Y, Yan L, Huai D, Jiang H, et al. Visualizing the Distribution of Lipids in Peanut Seeds by MALDI Mass Spectrometric Imaging. Foods. 2022; 11(23):3888. https://doi.org/10.3390/foods11233888
Chicago/Turabian StyleWang, Xin, Yuning Chen, Yue Liu, Lei Ouyang, Ruonan Yao, Zhihui Wang, Yanping Kang, Liying Yan, Dongxin Huai, Huifang Jiang, and et al. 2022. "Visualizing the Distribution of Lipids in Peanut Seeds by MALDI Mass Spectrometric Imaging" Foods 11, no. 23: 3888. https://doi.org/10.3390/foods11233888