
The Influence of Flower Head Order and Gibberellic Acid Treatment on the Hydroxycinnamic Acid and Luteolin Derivatives Content in Globe Artichoke Cultivars


 , and
, and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Design
2.2. Average Weight of Artichoke Flower Head Orders
2.3. Extraction of Phenolic Compounds
2.4. Identification and Quantification of Hydroxycinnamic Acids and Luteolin Derivatives
2.5. Statistical Analysis
3. Results and Discussion
3.1. Effect of Flower Head Order and Gibberellic Acid Treatment on Average Flower Head Weight
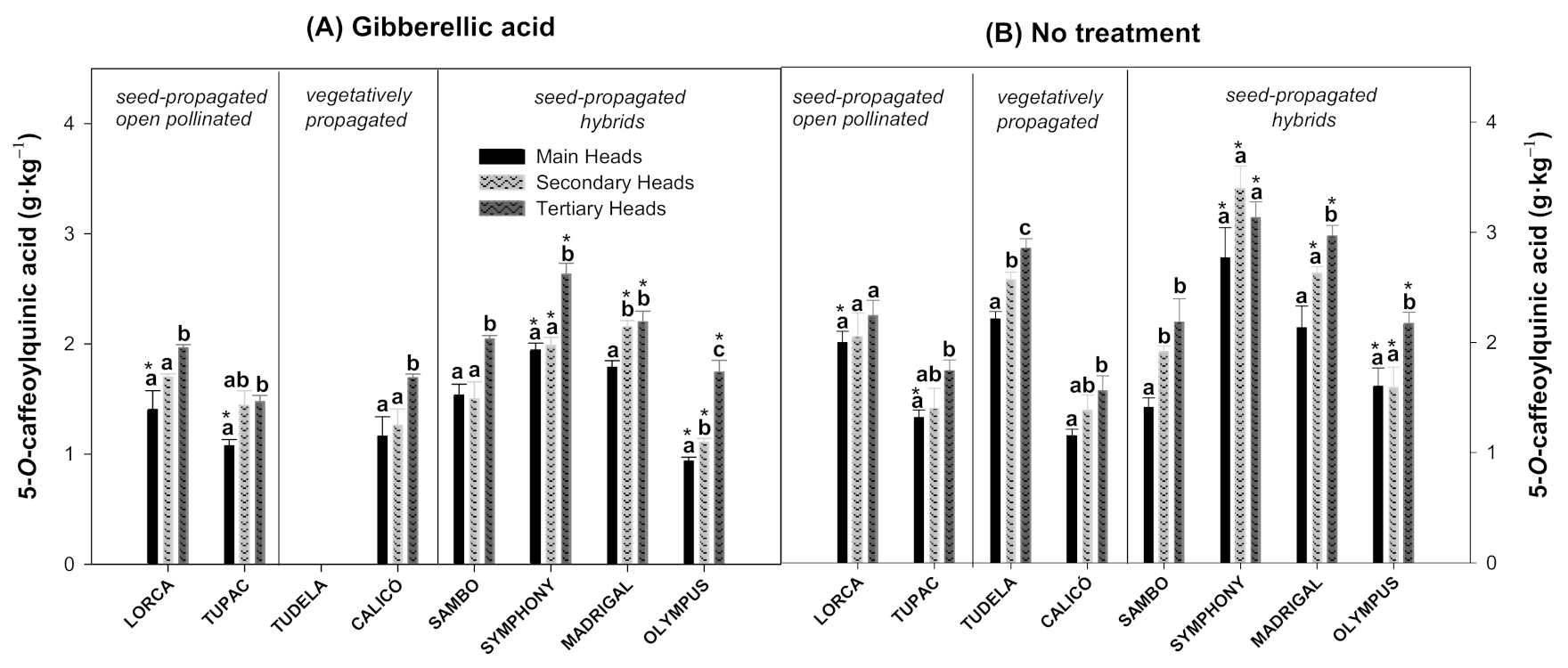
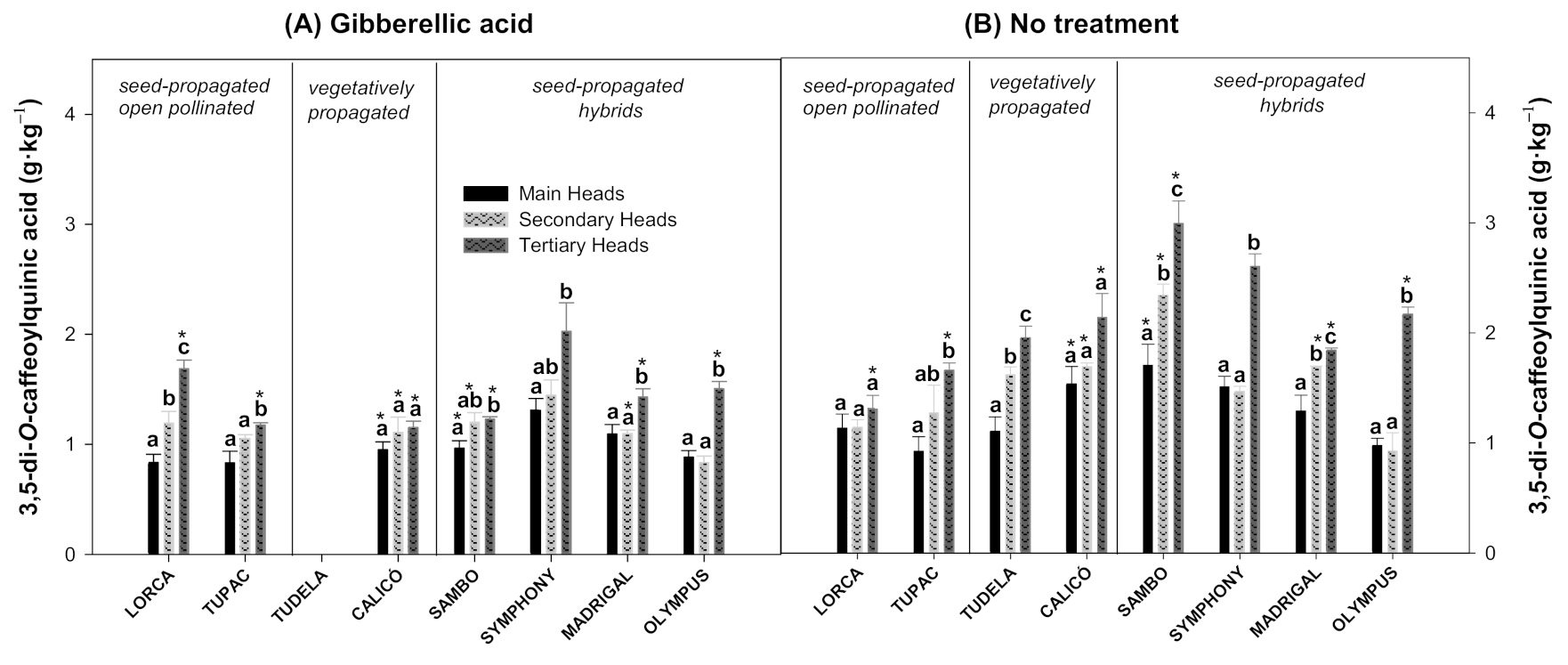
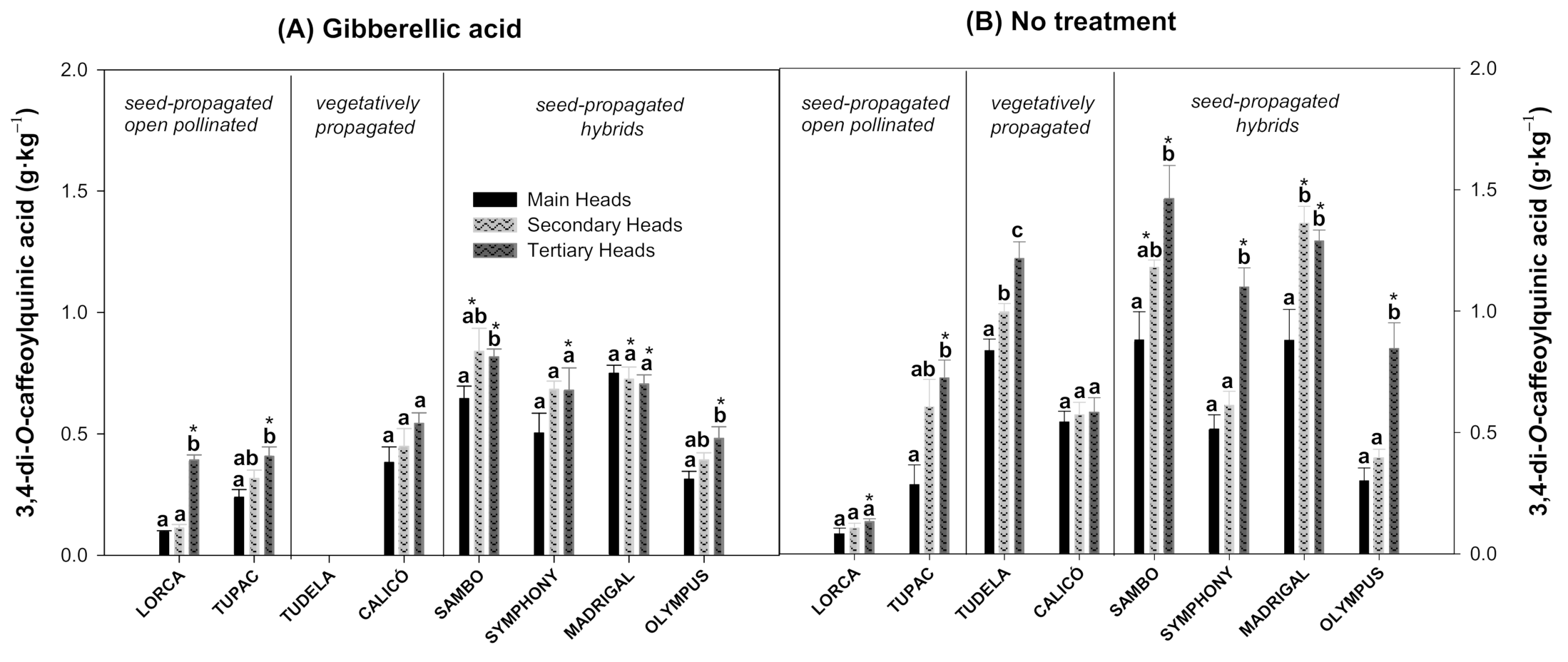
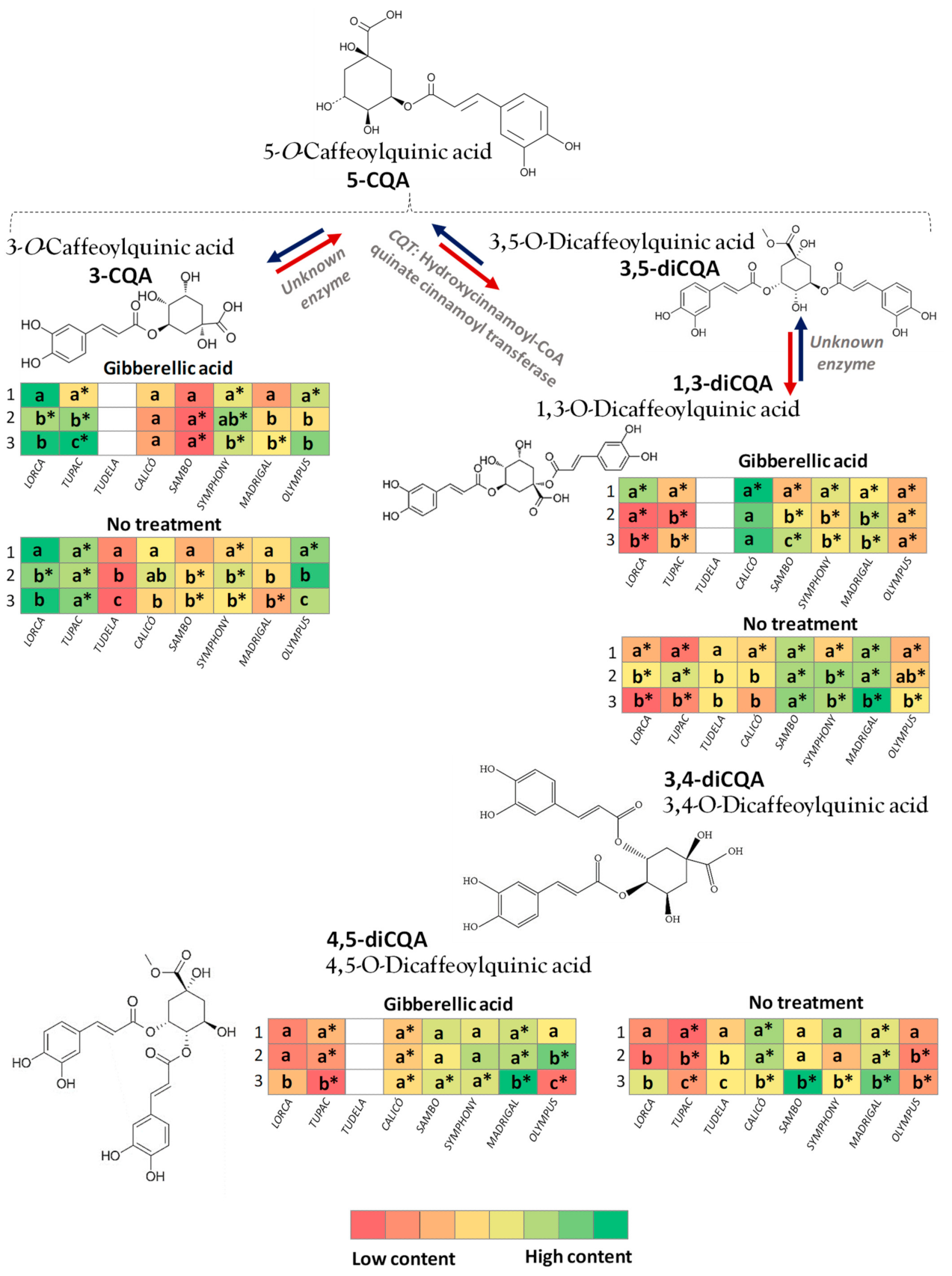
3.2. Effect of Flower Head Order and Gibberellic Acid Treatment on Hydroxycinnamic Acid Content
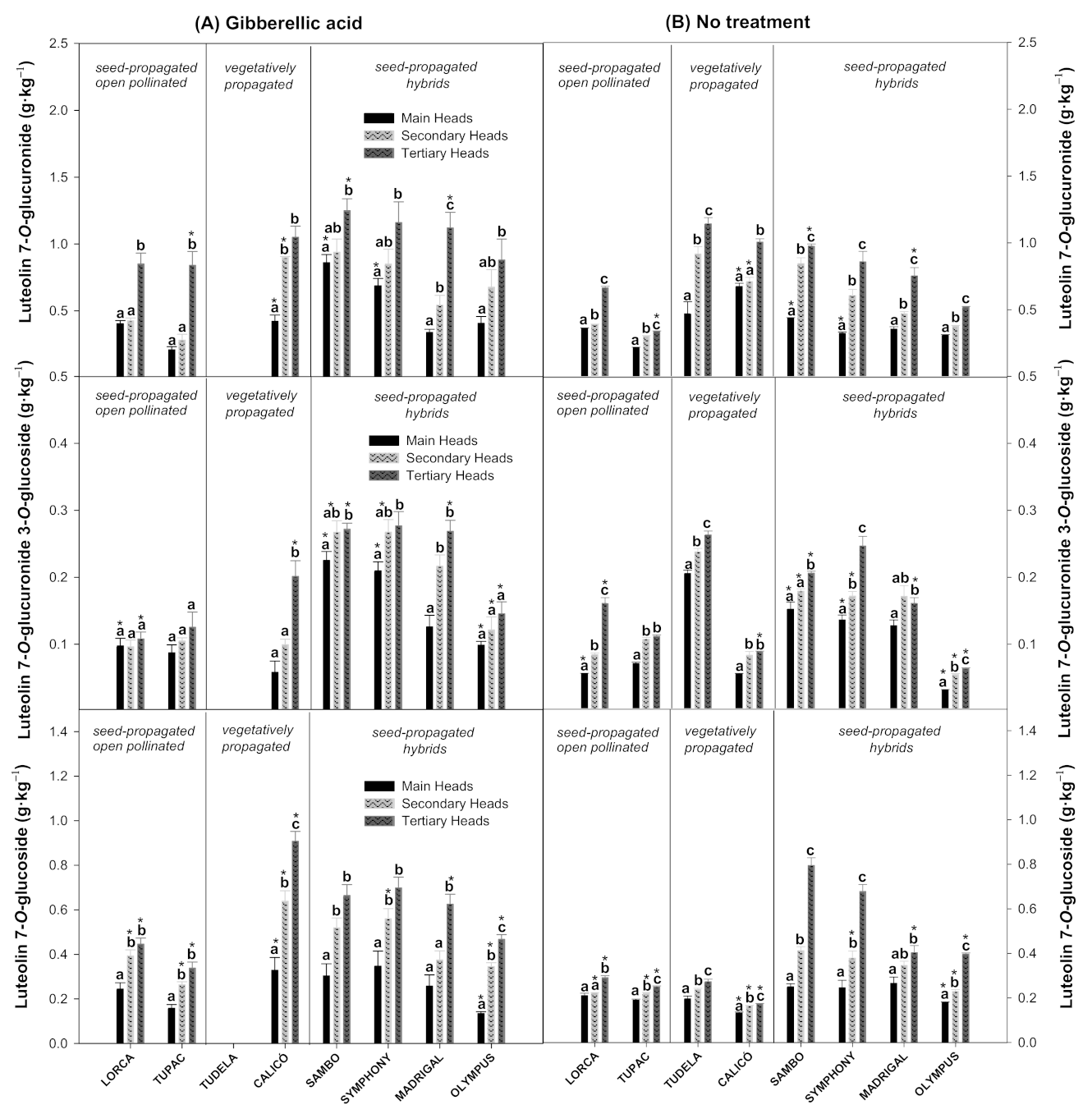
3.3. Effect of Flower Head Order and Gibberellic Acid Treatment on the Luteolin Derivatives Content
3.4. Effect of Flower Head Order and Gibberellic Acid Treatment on Total Phenolic Content
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brat, P.; Georgè, S.; Bellamy, A.; Du Chaffaut, L.; Scalbert, A.; Mennen, L.; Arnault, N.; Amiot, M.J. Daily polyphenol intake in France from fruit and vegetables. J. Nut. 2006, 136, 2368–2373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jun, N.J.; Jang, K.C.; Kim, S.C.; Moon, D.Y.; Seong, K.C.; Kang, K.H. Radical scavenging activity and content of cynarin (1,3-dicaffeoylquinic acid) in Artichoke (Cynara scolymus L.). J. Appl. Biol. Chem. 2007, 50, 244–248. [Google Scholar]
- Pinelli, P.; Agostini, F.; Comino, C.; Lanteri, S.; Portis, E.; Romani, A. Simultaneous quantification of caffeoyl esters and flavonoids in wild and cultivated cardoon leaves. Food Chem. 2007, 105, 1695–1701. [Google Scholar] [CrossRef]
- Häusler, M.; Ganzera, M.; Abel, G.; Popp, M.; Stuppner, H. Determination of caffeoylquinic acids and flavonoids in Cynara scolymus L. by high performance liquid chromatography. Chromatographia 2002, 56, 407–411. [Google Scholar] [CrossRef]
- Vázquez-Olivo, G.; Antunes-Ricardo, M.; Gutiérrez-Uribe, J.A.; Osuna-Enciso, T.; León-Félix, J.; Basilio-Heredia, J. Cellular antioxidant activity and in vitro intestinal permeability of phenolic compounds from four varieties of mango bark (Magnifera indica L.). J. Sci. Food Agric. 2019, 99, 3481–3489. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, S.; Gascón, S.; Luquin, A.; Laguna, M.; Ancín-Azpilicueta, C.; Rodríguez-Yoldi, M.J. Rosa canina extracts have antiproliferative and antioxidant effects on Caco-2 human colon cancer. PLoS ONE 2016, 11, e0159136. [Google Scholar] [CrossRef] [Green Version]
- Lombardo, S.; Pandino, G.; Mauromicale, G.; Carle, R.; Knodler, M.; Schieber, A. New seed propagated cultivars of globe artichoke suitable for processing uses. Acta Hortic. 2012, 942, 139–146. [Google Scholar] [CrossRef]
- Pandino, G.; Lombardo, S.; Mauromicale, G.; Williamson, G. Profile of polyphenols and phenolic acids in bracts and receptacles of globe artichoke (Cynara cardunculus var. scolymus) germplasm. J. Food Compos. Anal. 2011, 24, 148–153. [Google Scholar] [CrossRef]
- Mauromicale, G.; Portis, E.; Acquardo, A.; Lo Monaco, A.; Pesce, G.R.; Lanteri, S. An integrated model to accelerate the development of seed-propagated varieties of globe artichoke. Crop Breed. Appl. Biot. 2018, 18, 72–80. [Google Scholar] [CrossRef] [Green Version]
- Mauromicale, G.; Ierna, A. Characteristics of heads of seed-grown globe artichoke (Cynara cardunculus L. var. scolymus (L.) Fiori) as affected by harvest period: Sowing date and gibberellic acid. Agronomie 2000, 20, 197–204. [Google Scholar] [CrossRef]
- Baixauli, C.; Giner, A.; Aguilar, J.M.; Najera, I.; Miguel, A.; Lopez Galarza, A.; Pascual, B.; San Bautista, A.; Maroto, J.V. Agronomic behaviour of seed propagated artichoke cultivars in the Spanish Mediterranean area. Acta Hortic. 2012, 942, 361–367. [Google Scholar] [CrossRef]
- Lombardo, S.; Pandino, G.; Ierna, A.; Mauromicale, G. Variation of polyphenols in a germplasm collection of globe artichoke. Food Res. Int. 2012, 46, 544–551. [Google Scholar] [CrossRef]
- Saleh, S.A.; Ezzo, M.I.; Glala, A.A. Comparison between water requirements of globe artichoke grown from seeds and from vegetative organs under new reclaimed lands. Egypt. J. Appl. Sci. 2007, 22, 671–681. [Google Scholar]
- Saleh, S.A.; GH, A.E.L.-R.; El-Bassyouni, M.S.; Abdalla, A.M. Effect of gibberellic acid and micronutrients foliar applications on different seed-propagated artichokes under Egyptian condition. Egypt. J. Appl. Sci. 2017, 32, 615–634. [Google Scholar]
- Macua, J.I.; Lahoz, I.; Bozal, J.M. Evolution in seed propagated artichoke growing in cold zones of Spain. Acta Hortic. 2011, 942, 331–336. [Google Scholar] [CrossRef]
- Sałata, A.; Gruszecki, R.; Dyduch, J. Morphological and qualitative characterization of Globe artichoke (Cynara scolymus L.) cultivars ‘Symphony’ and ‘Madrigal’ on depending of the heads growth. Acta Sci. Pol. Hort. Cultus. 2012, 11, 67–80. [Google Scholar] [CrossRef]
- Foury, C. Trials on gibberellic acid application to a spring crop of artichoke (Cynara scolymus) cv. Blanc Hyerois. Ann. L’Amelior. Plantes 1977, 27, 411–426. [Google Scholar]
- Mauromicale, G.; Ierna, A. Effects of gibberellic acid and sowing date on harvest time and yields of seed-grown globe artichoke (Cynara scolymus L.). Agronomie 1995, 15, 527–538. [Google Scholar] [CrossRef] [Green Version]
- Maroto, J.V. Effects of gibberellic acid (GA3) applications on globe artichoke production. Acta Hortic. 2007, 730, 137–142. [Google Scholar] [CrossRef]
- Foury, C. Le deuxiéme congrés internacional de l’Artichaut. P.H.M. Rev. Hortic. 1974, 145, 31–36. [Google Scholar]
- Elia, A.; Calabrese, N.; Bianco, V.V. Sowing time, gibberellic acid treatment and cultivars of “seed” propagated artichoke. Acta Hortic. 1994, 371, 347–354. [Google Scholar] [CrossRef]
- Calabrese, N.; Bianco, V.V. Effect of gibberellic acid on yield and quality of seed grown artichoke (Cynara cardunculus L. var. scolymus L.). Acta Hortic. 2000, 514, 25–32. [Google Scholar] [CrossRef]
- Schrader, W.L. Growth regulator effects on earliness and yield in artichokes grown as annual from seed. HortScience 1992, 27, 643. [Google Scholar] [CrossRef] [Green Version]
- Sharaf-Eldin, M.A.; Schnitzler, W.H.; Nitz, G.; Razin, A.M.; El-Oksh, I.I. The effect of gibberellic acid (GA3) on some phenolic substances in globe artichoke (Cynara cardunculus var. scolymus (L) Fiori). Sci. Hortic. 2007, 111, 326–329. [Google Scholar] [CrossRef]
- Giménez, M.J.; Abadía, R.; Valero, D.; Serrano, M.; García-Pastor, M.E.; Medina-Santamarina, J.; Martínez-Esplá, A.; Zapata, P.J. Variation in polyphenolic composition and physiological characteristics of ’Blanca de Tudela’ cultivar affected by water stress. Acta Hortic. 2020, 1284, 235–240. [Google Scholar] [CrossRef]
- Martínez-Esplá, A.; García-Pastor, M.E.; Zapata, P.J.; Guillén, F.; Serrano, M.; Valero, D.; Gironés-Vilaplana, A. Preharvest application of oxalic acid improves quality and phytochemical content of artichoke (Cynara scolymus L.) at harvest and during storage. Food Chem. 2017, 230, 343–349. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Esplá, A.; Valero, D.; Martínez-Romero, D.; Castillo, S.; Giménez, M.J.; García-Pastor, M.E.; Serrano, M.; Zapata, P.J. Preharvest Application of Methyl Jasmonate as an Elicitor Improves the Yield and Phenolic Content of Artichoke. J. Agric. Food Chem. 2017, 65, 9247–9254. [Google Scholar] [CrossRef]
- Grabowska, A.; Caruso, G.; Mehrafarin, A.; Kalisz, A.; Gruszecki, R.; Kunicki, E.; Sękara, A. Application of modern agronomic and biotechnological strategies to valorise worldwide globe artichoke (Cynara cardunculus L.) potential—An analytical overview. Ital. J. Agron. 2018, 13, 279–289. [Google Scholar] [CrossRef]
- Macua, J.I. New horizons for artichoke cultivation. Acta Hortic. 2007, 730, 39–48. [Google Scholar] [CrossRef]
- Bonasia, A.; Conversa, G.; Lazzizera, C.; Gambacorta, G.; Elia, A. Morphological and qualitative characterization of globe artichoke head from new seed-propagated cultivars. J. Sci. Food Agric. 2010, 90, 2689–2693. [Google Scholar] [CrossRef]
- Dumičić, G.; Goreta Ban, S.; Bućan, L.; Borošić, J.; Poljak, M. Effect of gibberellic acid plication on growth and yield of artichokes under summer conditions. J. Food Agric. Environ. 2009, 7, 620–626. [Google Scholar] [CrossRef]
- Goreta, S.; Bućan, L.; Dumičić, G. Effect of environment and gibberellic acid (GA3) on earliness and yield of globe artichokes. Acta Hortic. 2004, 660, 155–159. [Google Scholar] [CrossRef]
- Basnizki, J.; Goldschmidt, E.E. Further examination of gibberellin A3 effects on flowering of globe artichokes (Cynara scolymus L.) under controlled environment and field conditions. Isr. J. Plant Sci. 1994, 42, 159–166. [Google Scholar] [CrossRef]
- Schütz, K.; Kammerer, D.; Carle, R.; Schieber, A. Identification and quantification of caffeoylquinic acids and flavonoide from artichoke (Cynara scolymus L.) heads, juice, and pomace by HPLC-DAD-ESI/MSn. J. Agric. Food Chem. 2004, 52, 4090–4096. [Google Scholar] [CrossRef]
- Lattanzio, V.; Kroon, P.A.; Linsalata, V.; Cardinali, A. Globe artichoke: A functional food and source of nutraceutical ingredients. J. Funct. Foods. 2009, 1, 131–144. [Google Scholar] [CrossRef]
- Lombardo, S.; Pandino, G.; Mauromicale, G. The nutraceutical response of two globe artichoke cultivars to contrasting NPK fertilizer regimes. Food Res. Int. 2015, 76, 852–859. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G.; Graziani, G.; Ritieni, A.; Cardarelli, M.; De Pascale, S. Phenolic composition, antioxidant activity and mineral profile in two seed-propagated artichoke cultivars as affected by microbial inoculants and planting time. Food Chem. 2017, 234, 10–19. [Google Scholar] [CrossRef]
- Gagliardi, A.; Giuliani, M.M.; Carucci, F.; Francavilla, M.; Gatta, G. Effects of the Irrigation with Treated Wastewaters on the Proximate Composition, Mineral, and Polyphenolic Profile of the Globe Artichoke Heads [Cynara cardunculus (L.)]. Agronomy 2020, 10, 53. [Google Scholar] [CrossRef] [Green Version]
- Ferro, A.M.; Ramos, P.; Guerreiro, O.; Jerónimo, E.; Pires, I.; Capel, C.; Capel, J.; Lozano, R.; Duarte, M.F.; Oliveira, M.M.; et al. Impact of novel SNPs identified in Cynara cardunculus genes on functionality of proteins regulating phenylpropanoid pathway and their association with biological activities. BMC Genom. 2017, 18, 183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turkiewicz, I.P.; Wojdyło, A.; Tkacz, K.; Nowicka, P.; Hernández, F. Antidiabetic, Anticholinesterase and Antioxidant Activity vs. Terpenoids and Phenolic Compounds in Selected New Cultivars and Hybrids of Artichoke Cynara scolymus L. Molecules 2019, 24, 1222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sałata, A. Wpływ Zabiegów Agrotechnicznych na Plon i Wartość Biologiczna˛ Karczocha Zwyczajnego (Cynara Scolymus L.); Wydawnictwo Uniwersytetu Przyrodniczego w Lublinie: Lublinie, Poland, 2010. [Google Scholar]
- Lombardo, S.; Pandino, G.; Mauromicale, G.; Knödler, M.; Reinhold, C.; Schieber, A. Influence of genotype, harvest and plant part on polyphenolic composition of globe artichoke (Cynara cardunculus L. var. scolymus L. (Fiori)). Food Chem. 2010, 119, 1175–1181. [Google Scholar] [CrossRef]
- Pandino, G.; Courts, F.L.; Lombardo, S.; Mauromicale, G.; Williamson, G. Caffeylquinic acids and flavonoids in the immature inflorescence of globe artichoke, wild and cultivated cardoon (Cynara cardunculus L.). J. Agric. Food Chem. 2010, 58, 1026–1031. [Google Scholar] [CrossRef] [PubMed]
- Pavlica, S.; Gebhardt, R. Protective effects of ellagic and chlorogenic acids against oxidative stress in PC12 cells. Free Radic. Res. 2005, 39, 1377–1390. [Google Scholar] [CrossRef]
- Roberfroid, M.B. A European consensus of scientific concepts of functional foods. Nutrition 2000, 16, 689–691. [Google Scholar] [CrossRef]
- Wang, M.; Simon, J.E.; Aviles, I.F.; He, K.; Zheng, Q.Y.; Tadmor, Y. Analysis of antioxidative phenolic compounds in artichoke (Cynara scolymus L.). J. Agric. Food Chem. 2003, 51, 601–608. [Google Scholar] [CrossRef] [PubMed]
- Schütz, K.; Persike, M.; Carle, R.; Schieber, A. Characterization and quantification of anthocyanins in selected artichoke (Cynara scolymus L.) cultivars by HPLC–DAD–ESI–MSn. Anal. Bioanal. Chem. 2006, 384, 1511–1517. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gibberellic Acid | No Treatment | |||||
|---|---|---|---|---|---|---|
| Main Heads | Secondary Heads | Tertiary Heads | Main Heads | Secondary Heads | Tertiary Heads | |
| LORCA | 240.37 ± 10.40c | 175.76 ± 12.84b | 122.57 ± 4.08a | 273.82 ± 19.68c | 160.48 ± 3.20b | 123.17 ± 8.50a |
| TUPAC | 280.99 ± 7.01b | 178.65 ± 9.86a | 167.15 ± 7.91a | 306.38 ± 14.57c | 179.52 ± 5.38b | 148.61 ± 4.66a |
| TUDELA | - | - | - | 209.39 ± 8.81c | 180.34 ± 5.20b | 156.33 ± 6.99a |
| CALICÓ | 472.22 ± 32.75c | 361.82 ± 23.25b | 178.93 ± 13.25a * | 496.62 ± 18.50c | 333.26 ± 13.86b | 261.69 ± 22.49a * |
| SAMBO | 373.90 ± 20.96c * | 300.25 ±18.86b * | 189.75 ± 9.94a * | 540.20 ± 13.58c * | 393.21 ± 11.93b * | 266.39 ± 18.76a * |
| SYMPHONY | 243.13 ± 15.58c * | 158.69 ± 8.24b | 106.55 ± 6.83a | 391.55 ± 21.64c * | 196.34 ± 12.69b | 121.33 ± 6.40a |
| MADRIGAL | 297.83 ± 10.70c * | 222.88 ± 10.17b * | 155.40 ± 5.45a * | 455.75 ± 32.62c * | 279.19 ± 15.92b * | 224.87 ± 7.44a * |
| OLYMPUS | 254.83 ± 12.25c | 206.98 ± 13.64b | 138.59 ± 8.58a * | 281.98 ± 11.76c | 225.26 ± 7.04b | 182.26 ± 12.33a * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giménez, M.J.; Giménez-Berenguer, M.; García-Pastor, M.E.; Parra, J.; Zapata, P.J.; Castillo, S. The Influence of Flower Head Order and Gibberellic Acid Treatment on the Hydroxycinnamic Acid and Luteolin Derivatives Content in Globe Artichoke Cultivars. Foods 2021, 10, 1813. https://doi.org/10.3390/foods10081813
Giménez MJ, Giménez-Berenguer M, García-Pastor ME, Parra J, Zapata PJ, Castillo S. The Influence of Flower Head Order and Gibberellic Acid Treatment on the Hydroxycinnamic Acid and Luteolin Derivatives Content in Globe Artichoke Cultivars. Foods. 2021; 10(8):1813. https://doi.org/10.3390/foods10081813
Chicago/Turabian StyleGiménez, María José, Marina Giménez-Berenguer, María Emma García-Pastor, Joaquín Parra, Pedro Javier Zapata, and Salvador Castillo. 2021. "The Influence of Flower Head Order and Gibberellic Acid Treatment on the Hydroxycinnamic Acid and Luteolin Derivatives Content in Globe Artichoke Cultivars" Foods 10, no. 8: 1813. https://doi.org/10.3390/foods10081813