
Synergistic Effects of Climate Change and Alien Fish Invasions in Freshwater Ecosystems: A Review



Department of Chemistry, Biology and Biotechnologies, University of Perugia, 06123 Perugia, Italy
*
Authors to whom correspondence should be addressed.
Fishes 2023, 8(10), 486; https://doi.org/10.3390/fishes8100486
Submission received: 31 August 2023
/
Revised: 21 September 2023
/
Accepted: 26 September 2023
/
Published: 28 September 2023
(This article belongs to the Special Issue Interactive Effects of Climate Change, Invasive Species, and Native Fish in Freshwater Ecosystems)
Abstract
:The interaction between climate warming and alien fish invasions could have severe consequences in inland waters, which represent isolated environments with high biodiversity richness. Due to the current climatic trend, increasing water temperatures and salinization, shorter ice cover period, and altered flow regimes are predicted for these environments. The aim of this review is to offer a comprehensive analysis at a global scale of the recent studies available in the literature, focused on the responses of invasive freshwater fish to the effects of climate change in both lotic and lentic environments. To achieve this purpose, we selected 34 relevant papers, using the Scopus and Google Scholar database. New alien species establishment, distribution range expansion of species already introduced, and exacerbating impacts on freshwater ecosystems and the native fish communities they host were included in the topics covered by the analyzed papers. Despite the uncertainties related to the unpredictability with which the effects of global warming will occur, the findings of the selected studies served as a support to evaluate conservation and management implications, with particular reference to possible mitigation strategies.
Key Contribution: The interactions between climate warming and alien fish invasions in inland waters is a topic of major concern for the biodiversity loss in different ecoregions of the planet. The analysis of the current knowledge on this issue, besides offering a comprehensive picture at a global scale, also allowed us to identify some gaps and future research directions and to discuss possible mitigation strategies.
1. Introduction
Freshwaters acquire particular importance from a conservation point of view, being among the most threatened environments in the world [1]. In inland waters, geographical isolation has favored speciation processes, responsible for the high biodiversity richness that these environments host [2]: 10% of all described species and 30% of vertebrates are supported by freshwaters, which make up only 0.8% of the planet’s surface [3]. At the same time, inland waters could be considered hotspots of anthropogenic degradation [4], where species become extinct faster than in other environments [5,6], and 83% of inhabiting animal populations showed a marked decreasing abundance trend between 1970 and 2018 [7]. Also, the International Union for Conservation of Nature (IUCN) Red List assessment reported that a substantial part of freshwater vertebrates is threatened [2,8]. Due to their position at the lowest level in the surrounding landscape, and their high level of hydrographic interconnections, freshwater ecosystems are subject to being the receptors of multiple anthropogenic stressors [9]. Major threats to freshwater biodiversity include overexploitation, water pollution, habitat alteration, and biological invasions; the direct effects of climate changes, in terms of altered precipitations pattern and increasing temperatures, overlap and could intensify all these interacting threats [5,10].
The impacts of human activities on inland waters have a negative effect on freshwater fish, which must face growing and increasingly intense threats, endangering the survival of many species [11]. As reported in the Eschmeyer’s Catalog of Fishes [12], over 18,000 fish species inhabit freshwaters, accounting for about 51% of known world fish diversity. According to the freshwater ecoregion world maps and respective belonging species reported in Abell et al. [13], fish biodiversity hotspots, in terms of species richness and endemism percentage, are predominantly concentrated in central-western Africa (including large portions of the Congo basin; the southern Gulf of Guinea watersheds; and Malawi, Tanganyika, and Victoria Lakes); Southeast Asia (Pearl River basin and adjacent hydrographic systems); and South America (most of the Amazon and Orinoco basins). Also, the Mediterranean area represents a biodiversity hotspot in terms of freshwater fish species richness [14]. Just as the distribution of global fish diversity is not homogeneous, but varies according to the geographical area, also the intensity and arrangement of threats show marked differences at the regional scale [11]. Of a total of 8389 freshwater fish species, 29% assessed for the IUCN Red List by 2017 were globally threatened with extinction, of which 5% were listed as Critically Endangered [15]. The highest numbers of threatened species referred to the Arabian Peninsula (50%), New Zealand (49%), Madagascar (43%), the eastern Mediterranean (41%), Europe (40%) [14,15], and South Africa (36%) [16]. Despite the limited information availability, high potential risks for the native fish diversity in the Amazon Region, in South America, are also expected in the near future [17]. The main threats to native fish fauna, with different intensity, depending on the geographic region considered, are habitat deterioration, water pollution (including lake eutrophication), river damming, over-water abstractions, alien species invasions, poaching, overfishing, changes in land use (i.e., deforestation and conversion of wetland habitats for farming), climate-induced decreasing precipitations and increasing water temperatures, and collections for aquarist trade [11].
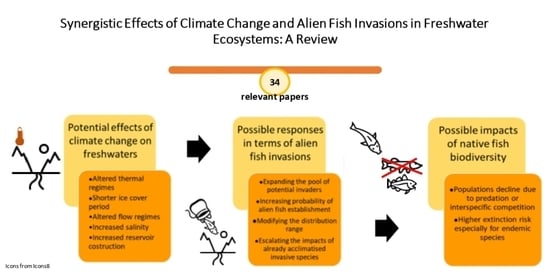
Invasive species and climate change are listed among the global drivers causing the decline of freshwater vertebrates [18], mainly represented by fish. Many fish species have limited dispersal ability to be able to respond to novel environmental conditions, and water temperature and habitat availability, which play a key role in the species survival, are closely linked to climate conditions [19]. The main effects of climate change on inland waters, which can influence the dispersal of alien species and adversely affect native fish biodiversity, are illustrated in Figure 1. Many studies from different countries provided evidence that global climatic variations involve increasing temperatures in running and standing waters [20,21], shorter duration of ice cover in Northern Hemisphere lakes [22], altered streamflow patterns [23,24], rising salinization [25], and increasing construction of artificial basins in arid areas [26].
According to the invasion triangle concept [27], the connections between climate change and species invasions need to be analyzed as a whole. In fact, biological invasions are multifactorial processes, in which the invader attributes, the biotic components, and the environmental characteristics of a potentially invasible site interact with each other in determining the invasion potential (Figure 2); within this conceptual framework are incorporated the external influences, including global environmental changes: as a result of novel environmental conditions, the susceptibility of an environment to being invaded can increase, and a novel set of species can exert its invasiveness and rapidly conquer new spaces, as well as the introduction pathways and dispersal patterns can change, and the alien species already acclimatized can expand their range, generating various impacts on indigenous species [27]. Accordingly to this concept, possible responses to the effects of global warming in terms of alien fish invasions include increasing the likelihood of new species entering and establishing themselves [28,29]. In this perspective, reservoirs could represent hot spots for the establishment and subsequent spread of invasive fish species, such as many limnophilous cyprinids [30,31]. The construction of new aquaculture plants for tropical fish could also encourage the spread of alien species [28]. For “casual” alien species, sensu Zenetos et al. [32], whose population survival is climate-limited, higher establishment rates under climate change conditions could be expected, on the condition that the introduction effort (i.e., the number of alien propagules repeatedly introduced to a location [33]) remains constant over time [28]. Many alien species can benefit from water warming, as they are often thermophilic species with wider habitat preferences [34]. Furthermore, climate-related changes in hydrologic conditions in lotic systems could result in the increased success of invasive species [35]; extreme climatic events have the potential to promote new arrivals from distant areas, favoring the spread of non-native species through intense floods [28]. As also reported for other systematic groups, another response to climate changes effects consists of alien fish range expansions, in terms of distribution shifts poleward and upward in elevation and, therefore, in the colonization of new environments with suitable thermal conditions [36]. In addition to increasing the invasibility of some environments, climate change can confer an invasive character or, in any case, accentuate the invasiveness of alien species that otherwise would have less impact on native biodiversity; in particular, as regards physiological aspects, rising temperatures could favor the performance improvement of alien fish, in terms of higher fecundity and faster growth [37,38]. Figure 3 reports the visual representation of the possible climate change influences on all the four phases of a successful alien freshwater fish invasion process: introduction, colonization, establishment, and spread.
The enhancement of interspecific competition for food and habitat, the exacerbation of predatory activity on native species, and invasive hybridization are among the possible cause of native populations decline and biodiversity loss, deriving from the above-mentioned synergistic effects between climate changes and alien species invasions [40,41]. These effects are of particular concern for the conservation of endemic species, which, having a limited distribution range, are particularly prone to extinction.
Despite the importance of this topic, there are relatively few studies that analyze the synergy between climate changes and alien species invasion, although in recent years, the focus on the subject is growing.
The aims of this review were to (i) explore the current state of knowledge on the possible synergistic effects between climate change and alien fish invasions in freshwater ecosystems, which are threatening the survival of many fish species; (ii) highlight what could be the knowledge gaps and the possible future research directions; and (iii) discuss management implications and possible mitigation strategies.
2. Materials and Methods
In order to select the scientific peer-reviewed literature, we conducted a search, considering papers including a combination of the following terms: ”climate + change” and “fish + invasions” and “native + fish” and ”freshwaters”. These searches were performed in Scopus and Google Scholar and were not restricted by year of publication. A total of 34 peer-reviewed papers were selected. Only papers focusing mainly on freshwater fish were included in the analysis.
3. Results
Among the 34 selected papers, 15 report on studies conducted in European freshwaters, while 4 papers refer to North America, 8 refer to Asian countries, 2 were conducted in South Africa, 1 refers to Central America, 1 is focused on subarctic regions, while 3 studies were carried out at global scale (Table 1). The geographical areas cover various ecoregions of the planet, testifying that the effects of climate change, combined with the impacts of biological invasions, is a topic of global concern for the biodiversity loss in many aquatic ecosystems belonging to different areas.
The target species reveal an extreme ecological diversification, including both rheophilous and limnophilous fish species. Although the articles were selected without setting time limits, most of them were published in recent years, falling in the period between 2008 and 2022, indicating that the subject is very topical. As far as the typology is concerned, thirty papers are original articles, while the remaining four papers are reviews based on literature data.
In the following paragraphs, for each of the main topics covered in the selected articles (altered ecological connectivity, altered distribution of species due to changes in the thermal regime, altered food webs) were analyzed and discussed the aspects relating to the impacts on inland waters, the implications for climate change, alien fish invasions and native biodiversity, possible mitigation measures and management implications, knowledge gaps, and future directions.
3.1. Altered Ecological Connectivity in Freshwaters
The alterations of the ecological connectivity in freshwaters, under climate change conditions, could be the consequence of the increase in the construction of dams to create new artificial reservoirs as a response to the rise in water demands, but they can also be due to the total drying up of river stretches during particularly dry periods, because of decreasing rainfall and increasing water withdrawals for human purposes. As a consequence of the flow rates reduction, the interruptions in river continuity can also be due to the presence of pollution, due to the decreasing dilution of pollutants.
Especially in developing areas, such as the Yangtse river, lower Mekong basin, Thailand, Laos, Cambodia, an increasing river damming is reported, while worldwide, about 50% of freshwater ecoregions are affected by large and medium-sized dams [70], which represent insurmountable barriers for aquatic organisms and determine river facies alterations both upstream and downstream of the dam. In areas characterized by Mediterranean climates, droughts of increasing intensity and frequency may also lead to strong alterations of river connectivity, with strong repercussions on water chemistry [46]. In this context, identifying the main drivers and ecological consequences of altered connectivity is a crucial aspect for the conservation of fish biodiversity and ecosystem functions.
3.1.1. Impacts of Altered Ecological Connectivity in Inland Waters
The often dendritic shape of hydrographic networks strongly influences the ecological consequences of river connectivity alterations, for example, by amplifying the effects of rivers damming and river flow regulation on the movements of fish, which are able to swim actively in the river, but in any case, they are bound to the aquatic habitat for their survival [70]. Also, the alteration of the chemical–physical characteristics of the water following dry periods could exert a heavy impact on the river ecosystem, in terms of increasing water temperature, turbidity, and salinity and reduction of flow and oxygen levels, causing negative impacts on the physiology of native fish, which, within certain limits, are adapted to face dry seasons, beyond which local extinction phenomena can occur [46]. The release of hypolimnic waters from reservoirs (i.e., colder and anoxic waters) can drastically change the thermal and chemical characteristics of the watercourses downstream of the dam, with profound repercussions on the fish community composition [76].
The impossibility for the fish to move longitudinally along the rivers due to the presence of weirs prevents them from performing ample migrations for breeding purposes and short movements to look for feeding habitats and covers or to escape from competitors and predators [70]. Thus, the extinction of many diadromous fish, which need to migrate between fresh and marine waters to complete their life cycle, has occurred [46,77]. The drastic decline of diadromous fishes (i.e., sturgeons, lampreys, eels, Atlantic salmon) has been observed especially in Europe over the last century, with the size and numerosity of weirs as main drivers of the extinction rate [78].
3.1.2. Implications for Climate Changes, Alien Fish Invasions, and Native Biodiversity
The possible interactive effects between climate change and biological invasions in inland waters depend on the ecological features of the species involved and on the characteristics of the ecosystem abiotic component [46,65]. Many invasive species, such as the common carp Cyprinus carpio, can take advantage under increasing drought frequency conditions, given their broad adaptability and wide ecological tolerance, as well as their high ability to colonize, and often dominate, strongly modified ecosystems [70].
Intense and long-lasting dry periods, with the consequent transition from lotic to lentic conditions, can promote the range expansion of alien species, producing strong impacts on native fish biodiversity. This is the case of the sandfish Labeo seeberi, an endangered cyprinid fish endemic to a single river basin in South Africa’s Cape Fold Ecoregion, which is a global biodiversity hotspot. Cerrilla et al. [46] reported a drastic decline for its remaining populations, due to the combined action of climate change, in terms of droughts, and non-native centrarchid (e.g., bluegill sunfish) predatory impacts.
3.1.3. Possible Mitigation Measures and Management Implications
To mitigate the impacts of altered ecological connectivity, sound management strategies aimed at the maintenance of the natural variability in flows and water levels should be implemented. The compliance with the “environmental flows” (i.e., the quantity and timing of water flows required to maintain the components, functions, processes, and resilience of aquatic ecosystems) rules; it is essential to allow the movement of fish and ensure the reproductive success, in terms of spawning and recruitment, of native fish populations [70,79]. The impacts due to the presence of dams and weirs insurmountable by the fish fauna can be mitigated by fish passages; if they are structured and sized to be efficient and effective for the target species, these facilities allow the fish movement along the longitudinal gradient of the rivers [70]. However, in some cases, the restoration of ecological connectivity may result in the further expansion of the alien fish species coming from downstream reaches. Therefore, management actions should be graduated according to the preservation of endemic freshwater fish, considering both the risks deriving from possible invasions and those deriving from the demographic and genetic isolation of native populations [70,80]. From the perspective of biodiversity conservation, a proper water resources management and concrete actions to contrast the further expansion of alien species (i.e., alien species eradication programs and native species reintroduction actions) should be implemented [46].
3.1.4. Knowledge Gaps and Future Directions
To fill knowledge gaps, more in-depth knowledge regarding the ecological preferences of fish species, and the demographic characteristics of the populations, with particular reference to the dynamics of their movements, are required, as they are currently limited for many species [70].
As future directions, the collection and systemization of experimental data collected in a multidisciplinary way using different methodologies (molecular analyses, telemetry studies, stable isotope techniques) will provide an overall picture of the altered ecological connectivity implications at the spatial–temporal level [70].
3.2. Altered Species Distribution Due to Changes in Thermal Regime
The freshwater fish species richness and the fish assemblage composition vary according to the latitude, and it is the result of the dispersal processes that have characterized the fish fauna in the course of past geological eras, with special reference to glaciations [13,21]. However, the current distribution of fish species is also affected by global warming, which results in freshwaters increasing temperatures [67]. Being ectothermic organisms, fish are particularly sensitive to changes in temperature [81]; they exclusively rely on behavioral strategies for thermoregulation, by moving in the water according to their thermal preferences [21]. Therefore, fish move in search of their thermal optimum, as long as suitable temperatures are available; otherwise, they can face local extinction phenomena [21,43]. Thermal-mediated range shift distribution is one of the most studied climate change effects on freshwater fish. In particular, in the subarctic area, has been observed the distribution range contraction of cold-water species, which do not have the possibility to move further upstream due to dispersal inability; the same area could be subject to invasion by warm-water fish, which could take advantage of increasing temperatures and expand their range [67]. A similar prediction can be referred to temperate regions, where changes in the thermal regime can increase the invasibility of freshwater ecosystems by thermophilic species. Also, the Mediterranean rivers, due to the particular climatic and geographic conditions and their high species richness, are particularly vulnerable to habitat variations and alien species introductions, experiencing a drastic decline in native fish biodiversity [82]. Carosi et al. [57] provided evidence, in the Tiber River basin (Central Italy), for synergistic effects between climate changes and alien fish invasions, in terms of species range shifts towards upper stream reaches mediated by rising water temperatures. Albeit to a lesser extent, also for tropical freshwater are predicted water warming and prolonged drought conditions in the next decades, together with salinization resulting from sea level rise, with negative repercussions on native biodiversity [83].
3.2.1. Ecological Impacts of Altered Thermal Regime in Freshwaters
In general, increasing water temperatures in freshwater systems results in water chemistry alterations, in terms of decreasing dissolved oxygen concentration and exacerbation of contaminants toxicity [35]. In lakes, eutrophication phenomena may be amplified, and stratification could become more pronounced and longer. These effects strongly affect the availability of a suitable habitat for fish. The composition of fish assemblages may be altered as a response to species range shifts. In freshwaters, climate-induced rising temperatures could also exert ecological community-level impacts on food webs, altering predator–prey interactions and enhancing the growth and feeding rate of warm-waters invaders to the detriment of native fish species [44].
The effects of alterations in the thermal regime in freshwaters can be differentiated according to the different climatic areas. This because higher latitudes are predicted to experience higher temperature change than tropical and subtropical latitudes [35].
According to Rolls et al. [67], sub-arctic freshwater could be considered as sentinel ecosystems of the interactive effects between climate change and alien species invasions, due to the pronounced changes in terms of rising temperatures compared to the global average, and due to the high susceptibility to being invaded by species coming from temperate zones. As ecological responses to global warming, subarctic freshwater is facing a shortening of the ice cover duration, while intensification of summer dry periods will lead to reductions in summer discharge and habitat availability for fish [67]. Other potential effects of warming temperatures will likely be the disappearance or seasonal drying up of small rivers and shallow lakes due to evaporation processes [67].
Long-term studies on ice phenology conducted in many rivers and lakes in Eurasia and North America provided evidence for later freezing, earlier ice break-up, and thus shorter ice duration due to increasing winter/early spring air temperatures [84]. In stratified lakes, the heating of the hypolimnetic layer can favor anoxic conditions. The mixing of the water can be influenced by a diversification of the wind patterns and modify the balance in the distribution of heat within the water mass. The surface waters could experience an alteration of the chemical–physical characteristics as a function of the action of the wind and the temperature [84].
Jeppesen et al. [85] investigated climate-induced changes in several Danish freshwaters using long-term data series. According to the climatic models for the continental zones, for Denmark is predicted a warmer and wetter climate, with high temperatures in late summer and winter, more precipitation in winter, and lower precipitation in summer. The study provided evidence that these climatic changes are already ongoing, causing increasing differences in temperatures between surface and bottom water in both medium-deep and deep lakes, reflecting a more stable and longer stratification of the lakes, even if this effect could be buffered by the action of the wind.
The Mediterranean climate regions are among the most vulnerable ecosystems to the effects of climate change. The water warming and the increasingly intense and prolonged summer drought periods can make the Mediterranean freshwaters increasingly inhospitable, especially for cold-water species, due to high temperatures and low oxygen levels. In Mediterranean inland waters, to face increasing drought and the progressive worsening of water quality, freshwater fish could shift their distributional range, adapt to the new environmental conditions through life-history traits plasticity, or could face extinction, resulting in decreasing freshwater [86,87].
In subtropical areas, increased storm intensity and frequency results in extreme and rapid flow events, while baseflows remain low and drought periods are increasingly extended over time. Mulholland et al. [88] reported for southern Florida and eastern Mexico several ecological effects of rising temperatures in freshwaters, including an overall increase in primary production rates, organic matter decomposition, and nutrient cycling; reduction in water quality and suitable habitat for cool-water species; expansion of subtropical species northwards, including alien species, whose range was originally limited to southern Florida; and increasing eutrophication of Florida lakes.
3.2.2. Implications for Climate Changes, Alien Fish Invasions, and Native Biodiversity
In response to climate variation, many invasive freshwater species are extending their distribution, and this raises a big concern for the native biodiversity conservation [34]. Some studies provided evidence for synergistic effects between climate changes and alien fish species invasions, mediated by rising water temperatures; for warm- or cool-water species, distribution shifts toward higher altitudes and latitudes where showed, while cold-water species could experience range contractions [57,67].
In subarctic regions, increasing temperatures are associated with widespread poleward invasions of warm-water species as cyprinids (e.g., Leuciscus idus (L., 1758), Rutilus rutilus (L., 1758)), percids (Gymnocephalus cernua, Perca fluviatilis L., 1758), esocids (Esox lucius L., 1758), and centrarchids (Micropterus dolomieu, Micropterus salmoides (Lacepède, 1802)). Rolls et al. [67] reported individual-, population-, and community-level effects for fish inhabiting these areas. At the individual level, temperature increases can promote the reproductive success of spring-spawning invasive species (e.g., R. rutilus and P. fluviatilis), potentially increasing their invasive power. Moreover, the increasing water turbidity resulting from increased nutrient input can negatively affect the feeding behavior of subarctic freshwater fish, as many of them are visual hunting predators needing transparent waters to sight their prey; on the other hand, invasive species are not affected by this phenomenon, as they are adapted to feeding in water turbidity conditions. Another effect linked to the increase in nutrient inputs and ecosystem productivity concerns the anoxia conditions that can affect the hypolimnic zone of stratified lakes; these areas in the summer season can represent thermal refuge for cold-water species, which can undergo local extinctions in case of protracted oxygen depletion. At the population level, increasing winter temperature could promote the decrease of the spawning period duration and of the recruitment rate of cold-water species, and at the same time, promote earlier and protracted spawning of cool- and warm-climate species. Climate change also alters subarctic freshwater community interactions by influencing the habitat use, abundance, and structure of species populations and increasing interspecific competition for thermal refugia and prey sources.
In temperate regions, increasing water temperatures are predicted to promote the establishment and invasiveness of non-native warm-water fishes. Britton et al. [73] tested the possible responses of alien species introduced in England and Wales to warming temperatures, using climate predicting models, and spatial distribution and propagule pressure data. Their results suggested that climate change effects, combined with wide distribution and high propagule pressure, could favor the establishment with self-sustaining populations and invasiveness of C. carpio and Silurus glanis; both species were expected to enhance their growth performance and their reproductive success, with potential ecological impacts on native fishes, through interspecific competition or predation, and increasing water turbidity due to their benthic feeding and resulting sediments movement. Also, Souza et al. [47], on the basis of observational data, have demonstrated the distribution range expansion of C. carpio in the European lakes where it has been introduced, favored by the rising temperatures, which promote the reproductive success of the species and the establishment of self-sustaining populations; also, in this case, the species benefited from restocking activities. C. carpio is also the most abundant invasive species in the Azorean lakes, where the species produces detrimental ecological effects in terms of rising water turbidity and releasing of nutrients from sediments, due to the above-mentioned bottom-feeding behavior; all these effects will likely be enhanced by climate changes, thanks to the high ecological traits plasticity, which renders the species able to cope with changing environmental conditions [56].
The external boundaries of species ranges acquire particular importance in the analysis of the projected distribution dynamics of invasive species, providing insight into the mechanisms limiting or promoting range expansion in changing environments; moreover, it is precisely in these sites that it is advisable to concentrate management efforts to limit their dispersal. In the Columbia River basin (North America), through the combination of environmental DNA technology with species distribution models, Rubenson and Olden [59] analyzed the distribution range of the alien smallmouth bass M. dolomieu, paying special attention to the upstream boundary regions and estimating its overlap with that of native salmonids. The study provided evidence that warmer water temperature is a major driver in the spread of M. dolomieu and predicted an upstream invasion extension of 69% by 2080, under a moderate climate change scenario. The spatial overlap, ranging from 3 to 62%, with critical spawning and rearing habitats for native fish, is of concern due to predator and competitive impacts of M. dolomieu on juvenile salmonids. The same authors [66] analyzed in both space and time the population structure and seasonal movements patterns of M. dolomieu at the upstream extent of its range boundary in the John Day River, Oregon (USA), under climate changes conditions; their results highlighted an overall trend of upstream expansion, driven by favorable thermal conditions and by individual traits (i.e., body morphology of farther mobile individuals) that predispose fish to the exploration of new spawning habitats, with both front-edge (persistent populations) and leading-edge (not yet established populations) invasion extent. The methodological approach of the research assumes relevance from a management point of view, since understanding the movement patterns of invasive species is essential for the prediction of future range shifts and forecasting the speed of invasion.
At the global level, Jourdan et al. [52] used species distribution models to identify the climatic niche shift for two highly invasive warm-water mosquitofish, Gambusia affinis (Baird & Girard, 1853) and G. holbrooki, and evaluated their potential for further climate-related range extension. Their projections under future climatic scenarios showed high potential for many areas in Central Europe to be invaded by mosquitofish, mainly due to rising winter temperatures that will favor the establishment of the species. The potential for G. holbrooki invasion of Central Europe was also suggested by Benejam et al. [74], who explored the latitudinal variation of life-history traits and parasite load in the species, along a gradient from Southern France to Southern Spain, under climate change conditions; their results highlighted that the key role played by latitude allowed for elucidating some aspects of the interactive mechanisms of climate change and G. holbrooki invasiveness.Research focused on the feeding ecology of nine populations of G. holbrooki introduced in Italy and Spain showed that the feeding of the species does not seem to be negatively influenced by climate-induced rising water temperatures compared to other freshwater species, supporting the hypothesis of competitive superiority of the invasive mosquitofish over the native [55].
Also in Mediterranean running waters, the combined effect of water warming and decreasing flow rates may encourage colonization by invasive cool- and warm-water fish species. Carosi et al. [57] provided evidence for the range shift towards upstream reaches of the invasive Barbus barbus and Padogobius bonelli (Bonaparte, 1846), which have significantly extended their range in the Tiber River basin (central Italy) over the past decades; according to their findings, river fragmentation, elevation, water temperature, and average current speed seem to be major drivers in colonization processes, enhancing the dispersal ability of the alien species under global warming conditions. Similar results were obtained by Radinger et al. [64] for the Ebro River catchment; the authors found a close link between alien species distribution and water temperature, suggesting a further expansion of the upper-range boundary for non-native cyprinids (as carp and bleak), thanks to climate changes-related new spawning habitat availability.
There are concerns also for native tropical fish species, which are particularly vulnerable to water heating, since tropical freshwaters experience fairly constant high temperatures and low climatic variations throughout the year [42,89]. Gomez-Maldonado and Camacho-Cervantes [42] tested the effect of temperature on the refuge use and exploratory behavior of the endangered Skiffia bilineata that cohabits with the invasive Poecilia reticulata in central Mexico; they found that non-native species, under rising temperature conditions, were not affected in their refuge usage, while increasing their exploratory behavior, thus enhancing their dispersal ability. These findings confirmed that invaders could be less affected by climate changes, as they are often more plastic and resilient.
Some of the selected studies applied the risk screening tool called Aquatic Species Invasiveness Screening Kit (AS-ISK) [90] to predict alien species establishment and their invasion success. AS-ISK is applicable to all climatic areas and comprises a set of questions on the species biogeographical and historical traits, its biological and ecological interactions, its potential socio-economic impacts, and those on ecosystem services; additional questions are aimed to predict how forecasted changes in climate are likely to influence the risks of the introduction, establishment, dispersal, and impact of the species. To et al. [45] assessed the potential effects of the predicted future climatic conditions on the invasiveness of the 25 alien species occurring in lake Taal (Philippines). Using AS-ISK, the authors found that one of the species whose invasiveness may be most favored by global warming is Clarias batrachus (Siluriformes: Claridae), a tropical species native to Southeast Asia capable of tolerating extreme environmental conditions (water hypoxia, high salinity and pH values); rising temperatures (up to 35 °C) are expected to provide more suitable habitat availability for the establishment of the species. Mumladze et al. [50], using the same tool, identified the highest-invasiveness-risk species, under current and predicted climate conditions, for the South Caucasus biodiversity hotspot, including Armenia, Azerbaijan, and Georgia. They found that in this area, characterized by a continental mesophilic climate and by high endemic freshwater fish species richness, the number of very high-risk species is expected to triple within a climate change context. Among the most potentially invasive extant species, there are the gibel carp Carassius gibelio (Bloch, 1782) and the topmouth gudgeon Pseudorasbora parva both species may exert detrimental effects on the native species and ecosystems of the South Caucasus, which can be amplified by climate warming. Also, the horizon North African catfish Clarias gariepinus (Burchell, 1822) was ranked as a very high-risk species for this biogeographic region. Similar results were found by Tarkan et al. [68] for the Mediterranean shallow Lake Marmara in West Anatolia (Turkey); also, in this case, the invasive potential assessment of alien species with AS-ISK identified C. gibelio as the species having the highest score within the climate change risk assessment section, suggesting that the species could represent a major threat for the endemic fish diversity of the lake. Still in Turkey, AS-ISK was applied in combination with molecular analysis (mtDNA barcoding) to assess the origin, distribution, and potential impact of the invasive ruffe G. cernua [51]. The molecular approach revealed that the species probably originates from the European invasive range, while the risk assessment highlighted for G. cernua a high potential to invade in the next years, under the influence of climate change, all suitable habitats in Thrace water bodies; there, the species probably spreads using the natural connections of the hydrographic network, and establishes self-sustaining populations in many Turkish freshwater ecosystems, where it will be likely able to exert strong ecological negative impacts.
In headwater streams in Southern Africa (Cape Fold Ecoregion), Shelton et al. [65] found that rising water temperatures appear to limit the density and distribution of the invasive predatory rainbow trout Oncorhynchus mykiss, and thus, could limit negative impacts on small-bodied native fish, such as species within the genera Galaxias, Sandelia, and Pseudobarbus, during the summer season. Surprisingly, in this case, climate changes could act in favor of native species threatened by O. mykiss, even if, as a result of warming water, new habitats will be available for other limnophilic species coming from downstream reaches.
Another important implication for climate changes, alien fish invasion, and native biodiversity in freshwaters concerns the reduction of the native fish growth performance, with negative repercussions on fishery yield and then on food production, one of the ecosystem services provided by freshwater ecosystems [43]. As mentioned above, since fish are ectothermic organisms, growth is strongly influenced by water temperature, as well as by the food resources availability and by intra- and interspecific competition phenomena. The climate-induced lack of synchrony between food availability and optimal growth temperature, and the diet overlap with invasive species, are at the basis of the growth and fishery yield reduction. Within certain limits, defined by the speed with which environmental changes occur, and by the cost of adaptations, fish populations have the potential to cope with such adversities, through behavioral thermoregulation and physiological traits adaptations. DeWeber et al. [43], using long-term data series, applied integrated bioenergetics and growth modeling to analyze how nutrient dynamics, Gasterosteus aculeatus L., 1758 invasion, and rising temperature could affect the growth performance in the endemic cold-water whitefish Coregonus wartmanni inhabiting lake Constance in Germany. On the basis of their results, the combined effect of rising temperatures and competition with invasive G. aculeatus is predicted to exacerbate the whitefish growth reduction and to reduce fishery yields: warming increases respiration costs, and competition with G. aculeatus reduces consumption and thereby growth during the summer. The study provided an example of how climate-induced changes in freshwater ecosystems and invasive species can interact to strongly alter fish growth. Avlijaš et al. [44] analyzed the effects of warming temperatures on the functional responses of tench Tinca tinca, an Eurasian fish invading the Great Lakes basin (Canada); the authors demonstrated that rising temperature will improve tench feeding efficiency on low-density gastropod populations, producing indirect effects on food webs and enhancing competition with native fishes having similar dietary habits, such as redhorse species (Moxostoma spp.).
In some European temperate regions (Austria, Germany, UK, France, and Poland), the alien pumpkinseed Lepomis gibbosus of North American origin is predicted to enhance its invasiveness under warming temperature conditions [60,71,73,91,92]. The species was introduced in Europe more than a century ago, but not in all countries of its invaded range is it considered invasive [48]. Zięba et al. [60], through the application of a life-history traits-based model for the risk assessment of the species in Poland, revealed that among the major traits favoring L. gibbosus invasiveness, there was the high dispersal ability, promoted by rising temperatures and increasing hydrological variability; in addition to natural range expansion, the authors suggested that the plasticity of the species life-history traits, under climate warming conditions, favors growth performance and increasing recruitment rates, to the detriment of native species. A study conducted in UK experimental ponds by Copp et al. [69] was aimed at predicting the trophic consequences of increased L. gibbosus invasiveness, using stable isotope analysis. Their results provided evidence for significant negative impacts of L. gibbosus on the isotopic ecology and growth rates of the native gudgeon Gobio gobio and roach R. rutilus; according to the niche variation hypothesis, their feeding interactions with the invader led to diet constriction and increased diet specialization. L. gibbosus is also widely distributed in Ukraine, where Tytar et al. [48], based on the application of ecological niches models, prospected the potential expansion of the species into Belarus and Baltic countries; they found climatic predictors, with special reference to temperature, to be relevant for the species habitat suitability evaluation. A long-term study carried out on the fish community of Prespa Lakes (Southeastern Europe) suggested that L. gibbosus could represent a treat for the endemic Alburnoides prespensis (Karaman, 1924), for which a declining population trend has been recorded, starting from the period in which the invasive species has become established [57]; also, in this case, warming temperatures are proved to enhance the invasiveness of L. gibbosus, in terms of a prolonged spawning period and higher recruitment rate.
Climate-related rising temperatures will probably enhance also the round goby Neogobius melanostomus expansion, since the thermal optimum reported for the species is 26 °C [72]; thanks to its wide tolerance to extreme environmental conditions (i.e., high salinity level, low oxygen content), the species became one of the most widespread invasive fish at the global level, producing negative impacts on native species through competition for food and spawning habitats, and having also ecological effects in terms of food webs alteration and contaminants bioaccumulation [72].
Alpine freshwater ecosystems, as bioclimatic and biogeographic islands, can be considered particularly vulnerable to fish invasions, and climate change is likely to increase the invasibility of these systems. With the aim to understand the invasion pattern of P. parva and Carassius auratus in the Qinghai–Tibetan Plateau (China), two recent studies focused on the analysis of life-history traits variation according to environmental factors; the investigations revealed that the life-history plasticity of both species allowed them to regulate their growth and reproduction to adapt to extreme environmental conditions [62,63]. Climate change may favor the further expansion of C. auratus and P. parva in the Tibetan Plateau, whose dispersal is promoted also by the systematic release into the wild, for religious reasons, of specimens purchased in local markets.
Investigating changes over time of the correlation between fish functional structure and environmental conditions is important to discover the mechanisms underlying the composition of fish communities. Lin et al. [54] found that in the Min River (Southeastern China), a heavily impacted subtropical river ecosystem, environmental filtering selects over time species with similar functional traits. The identification of precipitation, temperature, and species invasion as the most important drivers for fish functional diversity can help to predict fish species redistribution in an ongoing climate change context. A study conducted across China, through the application of the species distribution model, predicted an increasing invasion risk under climate change conditions, especially in the southeastern part of the country [61]; warming waters could favor the spread and establishment of many warm-water alien species that will have new suitable habitats to colonize, already facilitated by the unintentional releases in the wild linked to the practices of aquaculture and trade in ornamental fish [61].
Many aquatic ecosystems are affected by multiple anthropogenic stressors; in these cases, it is difficult to establish the cause–effect relationship between the stressor and the negative effects, knowledge of which is essential to design effective management strategies aimed at conservation. The comparison between species and functional traits shifts in two highly perturbed Lake Erie basins (Canada) revealed that the dominant drivers of long-term community changes are altered nutrient inputs, instead of climate warming and invasive species [53]. Importantly, the identification of the dominant effect of a specific stressor offers the possibility of preparing targeted concrete conservation actions for natural ecosystems.
3.2.3. Possible Mitigation Measures and Management Implications
The application of risk screening tools, such as the Aquatic Species Invasiveness Screening kit (AS-ISK) [90], has proven to be effective in the prediction of alien species establishment and their invasion success under climate warming conditions; this kind of methodology could be useful in supporting alien species management strategies for the early detection, control, or eradication of invaders. According to Rubenson and Olden [59], molecular approaches, such as environmental DNA, can be useful to integrate predictions based on species distribution models, supporting the management of invasive species.
Effective conservation policy and management measures to be taken to contrast the climate-induced expansion of alien fish species should be integrated by concrete actions aimed at mitigating the global change effects on freshwater ecosystems already affected by multiple anthropogenic stressors [57]. The establishment of protection areas, where total autochthonous assemblages are preserved and, thus, play a key role in maintaining biodiversity, could be a good conservation strategy; also, the improvement of water quality in polluted rivers is required to minimize additional anthropogenic stressors for resilient fish populations to adapt to climate change. Moreover, the restoration of riparian areas and the cooling effect of vegetation on water temperature could be considered an effective management strategy to compensate for climate-induced losses in a suitable habitat for cold-water species and, at the same time, limiting the upstream dispersal of invasive warm-water fish [59,87].
The application of bioenergy models could be effective to understand fish growth responses to climate change effects and to support informed ecosystem-based management [43]. The synergistic effects of rising temperatures and alien fish invasions could affect the growth of species of fishery interest, resulting in a drastic decline in fishery yield, as in the afore-mentioned case of whitefish C. wartmanni in Lake Constance (Germany), whose growth performance was reduced by the combined effect of climate change and G. aculeatus invasion. According to DeWeber et al. [43], an integrated approach, based on both fish stock assessment and decision analysis, is essential to support management choices in terms of smaller mesh size or quotas to limit fishing effort.
3.2.4. Knowledge Gaps and Future Research Directions
Many authors agree on the need to assess the potential impacts of emerging aquatic invaders under projected climate change scenarios [44,51,52] and to predict which species are likely to become invasive under future warmer climatic conditions [60,75]. Especially in developing regions, there is a knowledge gap concerning biological invasions, in terms of high-quality fish-monitoring data, which are often limited or lacking [50], mainly due to limited resources for research. Providing current inventory, predicting future distributions of alien species, and understanding their impacts on native biodiversity are of crucial importance to implement conservation measures for freshwater ecosystems [50,59].
In a climate warming context, considering that rising temperatures could represent a threat for cold-water fish, and at the same time can represent an added value for warm-water species, the definition of the thermal requirements for freshwater fish is necessary to develop proper management strategies. The definition of the critical limit of thermal tolerance could be useful to assess the species ability to face thermal stress and their capacity to expand their range in warmer waters, with different implications depending on whether they are native or invasive fish [49].
As suggested by Gomez-Maldonado & Camacho-Cervantes [42], the changes occurring in the competitive interactions between natives and invaders, under different thermal conditions, represent another crucial aspect to be investigated to make further progress in the research; the authors examined behavioral aspects, providing evidence that invasive guppies in central Mexico benefited from exhibiting social behavior with native species [42]. Also, DeWeber et al. [43] underlined the importance of considering biotic interactions and synergistic effects in global change studies, as well as the value of mechanistic models for predicting possible future scenarios.
4. Conclusions
The analysis of the selected publications allowed for identifying some common methodologies that have proved to be particularly effective in deepening the knowledge on the synergistic effects between climate change and fish invasions in freshwaters, and in predicting the future evolution of the fish community composition, which could be radically changed due to local extinction or range shift phenomena of species that move upstream in search of a thermal optimum. For example, among the most used methodologies, approaches of the species distribution models and the AS-ISK can be counted.
Many authors agree that preventing the introduction of alien species is certainly the best strategy to pursue, rather than tackling costly and often incomplete eradication programs, especially if no action is taken in the early stages of the invasion process. In this context, understanding the mechanisms underlying the interactions between biological invasions and climate change is essential to predict how invasions may evolve in the future and, therefore, be able to act in advance. Some of these papers also underline the need to increase the knowledge of ecological features and dispersal patterns of invasive and native species for both prediction and intervention. The adoption of mitigation measures and sound management practice proposed in this review could prevent biodiversity loss, prioritizing hot-spot areas for biodiversity, where the climate changes effect is predicted to be more severe.
Many articles are focused on the effects of thermal regime alterations, highlighting increasing temperature as a key factor for the range expansion of invasive species and a threatening factor for native species, with special reference to the analysis of the projected distribution dynamics of freshwater fish communities facing global change. Less represented are the studies aimed at understanding the effects of river continuity interruptions, suggesting that there are knowledge gaps in this field that should be filled through further research. The reduction of flows and the alteration of natural hydrological regimes are among the effects of climate change that can most benefit alien species. Native species are adapted to the natural regimes of the watercourses, in which they have evolved and have synchronized their biological cycles according to them; altering the flow regime means favoring the less specialized and more adaptable alien species.
Finally, in the context of analysis aimed at highlighting the possible effects of climate change, only a few studies used long-term data, although conducting such studies is important. Moreover, as far as the study area is concerned, studies conducted at least at the catchment scale are essential from a management point of view.
Author Contributions
Conceptualization, A.C. and M.L.; writing—original draft preparation, A.C. and F.L.; writing—review and editing, A.C., M.L. and F.L.; visualization, A.C.; supervision, M.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Cantonati, M.; Poikane, S.; Pringle, C.M.; Stevens, L.E.; Turak, E.; Heino, J.; Richardson, J.S.; Bolpagni, R.; Borrini, A.; Cid, N.; et al. Characteristics, main impacts, and stewardship of natural and artificial freshwater environments: Consequences for biodiversity conservation. Water 2020, 12, 260. [Google Scholar] [CrossRef]
- Reid, A.J.; Carlson, A.K.; Creed, I.F.; Eliason, E.J.; Gell, P.A.; Johnson, P.T.; Kidd, K.A.; MacCormack, T.J.; Olden, J.D.; Ormerod, S.J.; et al. Emerging threats and persistent conservation challenges for freshwater biodiversity. Biol. Rev. 2019, 94, 849–873. [Google Scholar] [CrossRef] [PubMed]
- Strayer, D.L.; Dudgeon, D. Freshwater biodiversity conservation: Recent progress and future challenges. J. N. Am. Benthol. Soc. 2010, 29, 344–358. [Google Scholar] [CrossRef]
- Geist, J. Integrative freshwater ecology and biodiversity conservation. Ecol. Indic. 2011, 11, 1507–1516. [Google Scholar] [CrossRef]
- Dudgeon, D.; Arthington, A.H.; Gessner, M.O.; Kawabata, Z.I.; Knowler, D.J.; Lévêque, C.; Naiman, R.J.; Prieur-Richard, A.-H.; Soto, D.; Stiassny, M.L.J.; et al. Freshwater biodiversity: Importance, threats, status and conservation challenges. Biol. Rev. 2006, 81, 163–182. [Google Scholar] [CrossRef] [PubMed]
- Dudgeon, D. Multiple threats imperil freshwater biodiversity in the Anthropocene. Curr. Biol. 2019, 29, R960–R967. [Google Scholar] [CrossRef] [PubMed]
- WWF (World Wildlife Fund). Living Planet Report 2022—Building a Nature-Positive Society; WWF: Gland, Switzerland, 2022. [Google Scholar]
- Smith, K.G.; Barrios, V.; Darwall, W.R.T.; Numa, C. The status and distribution of freshwater biodiversity in the Eastern Mediterranean; IUCN: Cambridge, UK; Malaga, Spain; Gland, Switzerland, 2014; p. 132. [Google Scholar]
- GEO BON and FWBON. Inland Waters in the Post-2020 Global Biodiversity Framework. 2022. Available online: https://geobon.org/science-briefs/ (accessed on 18 August 2023).
- Poff, N.L.; Olden, J.D.; Strayer, D.L. Climate change and freshwater fauna extinction risk. In Saving a Million Species: Extinction Risk from Climate Change; Hannah, L., Ed.; Island Press/Center for Resource Economics: Washington, DC, USA, 2012; pp. 309–336. [Google Scholar] [CrossRef]
- Darwall, W.R.; Freyhof, J. Lost fishes, who is counting? The extent of the threat to freshwater fish biodiversity. In Conservation of Freshwater Fishes, 1st ed.; Closs, G.P., Krkosek, M., Olden, J.D., Eds.; Cambridge University Press: Cambridge, UK, 2016; pp. 1–36. [Google Scholar]
- Fricke, R.; Eschmeyer, W.N.; Van der Laan, R. (Eds.) Eschmeyer’s Catalog of Fishes: Genera, Species, References. Version of 11 Jan 2023. Available online: https://www.calacademy.org/scientists/projects/eschmeyers-catalog-of-fishes (accessed on 5 February 2023).
- Abell, R.; Thieme, M.L.; Revenga, C.; Bryer, M.; Kottelat, M.; Bogutskaya, N.; Coad, B.; Mandrak, N.; Balderas, S.C.; Bussing, W.; et al. Freshwater ecoregions of the world: A new map of biogeographic units for freshwater biodiversity conservation. BioScience 2008, 58, 403–414. [Google Scholar] [CrossRef]
- Geiger, M.F.; Herder, F.; Monaghan, M.T.; Almada, V.; Barbieri, R.; Bariche, M.; Berrebi, P.; Bohlen, J.; Casal-Lopez, M.; Delmastro, G.B.; et al. Spatial heterogeneity in the Mediterranean Biodiversity Hotspot affects barcoding accuracy of its freshwater fishes. Mol. Ecol. Resour. 2014, 14, 1210–1221. [Google Scholar] [CrossRef]
- Ramsar Convention on Wetlands. Global Wetland Outlook: State of the World’s Wetlands and their Services to People; Ramsar Convention Secretariat: Gland, Switzerland, 2018; p. 84. [Google Scholar]
- Jordaan, M.S.; Chakona, A.; Van der Colff, D. Protected areas and endemic freshwater fishes of the Cape Fold Ecoregion: Missing the boat for fish conservation? Front. Environ. Sci. 2020, 8, 502042. [Google Scholar] [CrossRef]
- Doria, C.R.D.C.; Agudelo, E.; Akama, A.; Barros, B.; Bonfim, M.; Carneiro, L.; Briglia-Ferreira, S.R.; Carvalho, L.N.; Bonilla-Castillo, C.A.; Charvet, P.; et al. The Silent Threat of Non-native Fish in the Amazon: ANNF Database and Review. Front. Ecol. Evol. 2021, 9, 646702. [Google Scholar] [CrossRef]
- Collen, B.; Whitton, F.; Dyer, E.E.; Baillie, J.E.; Cumberlidge, N.; Darwall, W.R.; Pollock, C.; Richman, N.I.; Soulsby, A.-M.; Böhm, M. Global patterns of freshwater species diversity, threat and endemism. Glob. Ecol. Biogeogr. 2014, 23, 40–51. [Google Scholar] [CrossRef] [PubMed]
- Woodward, G.; Perkins, D.M.; Brown, L.E. Climate change and freshwater ecosystems: Impacts across multiple levels of organization. Phil. Trans. R. Soc. B 2010, 365, 2093–2106. [Google Scholar] [CrossRef]
- Morrongiello, J.R.; Beatty, S.J.; Bennett, J.C.; Crook, D.A.; Ikedife, D.N.; Kennard, M.J.; Kerezsy, A.; Lintermans, M.; McNeil, D.G.; Pusey, B.J.; et al. Climate change and its implications for Australia’s freshwater fish. Mar. Freshw. Res. 2011, 62, 1082–1098. [Google Scholar] [CrossRef]
- Jeppesen, E.; Meerhoff, M.; Holmgren, K.; González-Bergonzoni, I.; Teixeira-de Mello, F.; Declerck, S.A.; De Meester, L.; Søndergaard, M.; Lauridsen, T.L.; Bjerring, R.; et al. Impacts of climate warming on lake fish community structure and potential effects on ecosystem function. Hydrobiologia 2010, 646, 73–90. [Google Scholar] [CrossRef]
- Sharma, S.; Richardson, D.C.; Woolway, R.I.; Imrit, M.A.; Bouffard, D.; Blagrave, K.; Daly, J.; Filazzola, A.; Granin, N.; Korhonen, J.; et al. Loss of ice cover, shifting phenology, and more extreme events in Northern Hemisphere lakes. J. Geophys. Res. Biogeosci. 2021, 126, e2021JG006348. [Google Scholar] [CrossRef]
- Patil, R.; Wei, Y.; Pullar, D.; Shulmeister, J. Sensitivity of streamflow patterns to river regulation and climate change and its implications for ecological and environmental management. J. Environ. Manag. 2022, 319, 115680. [Google Scholar] [CrossRef] [PubMed]
- Dhungel, S.; Tarboton, D.G.; Jin, J.; Hawkins, C.P. Potential Effects of Climate Change on Ecologically Relevant Streamflow Regimes. River Res. Appl. 2016, 32, 1827–1840. [Google Scholar] [CrossRef]
- Castillo, A.M.; Sharpe, D.M.; Ghalambor, C.K.; De León, L. Exploring the effects of salinization on trophic diversity in freshwater ecosystems: A quantitative review. Hydrobiologia 2018, 807, 1–17. [Google Scholar] [CrossRef]
- Fenoglio, S.; Bo, T.; Cucco, M.; Mercalli, L.; Malacarne, G. Effects of global climate change on freshwater biota: A review with special emphasis on the Italian situation. Ital. J. Zool. 2010, 77, 374–383. [Google Scholar] [CrossRef]
- Perkins, L.B.; Leger, E.A.; Nowak, R.S. Invasion triangle: An organizational framework for species invasion. Ecol. Evol. 2011, 1, 610–625. [Google Scholar] [CrossRef]
- Hulme, P.E. Climate change and biological invasions: Evidence, expectations, and response options. Biol. Rev. 2017, 92, 1297–1313. [Google Scholar] [CrossRef] [PubMed]
- Döll, P.; Bunn, S.E. Cross-chapter box on the impact of climate change on freshwater ecosystems due to altered river flow regimes. In Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part A: Global and Sectoral Aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Field, C.B., Barros, V.R., Dokken, D.J., Mach, K.J., Mastrandrea, M.D., Bilir, T.E., Chatterjee, M., Ebi, K.L., Estrada, Y.O., Genova, R.C., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2014; pp. 143–146. [Google Scholar]
- Liew, J.H.; Tan, H.H.; Yeo, D.C. Dammed rivers: Impoundments facilitate fish invasions. Freshw. Biol. 2016, 61, 1421–1429. [Google Scholar] [CrossRef]
- Clavero, M.; Hermoso, V. Reservoirs promote the taxonomic homogenization of fish communities within river basins. Biodivers. Conserv. 2011, 20, 41–57. [Google Scholar] [CrossRef]
- Zenetos, A.; Çinar, M.E.; Pancucci-Papadopoulou, M.A.; Harmelin, J.G.; Furnari, G.; Andaloro, F.; Bellou, N.; Streftaris, N.; Zibrowius, H. Annotated list of marine alien species in the Mediterranean with records of the worst invasive species. Mediterr. Mar. Sci. 2005, 6, 63–118. [Google Scholar] [CrossRef]
- Lockwood, J.L.; Cassey, P.; Blackburn, T. The role of propagule pressure in explaining species invasions. Trends Ecol. Evol. 2005, 20, 223–228. [Google Scholar] [CrossRef]
- Di Santo, V.; Jordan, H.L.; Cooper, B.; Currie, R.J.; Beitinger, T.L.; Bennett, W.A. Thermal tolerance of the invasive red-bellied pacu and the risk of establishment in the United States. J. Therm. Biol. 2018, 74, 110–115. [Google Scholar] [CrossRef]
- Ficke, A.D.; Myrick, C.A.; Hansen, L.J. Potential impacts of global climate change on freshwater fisheries. Rev. Fish Biol. Fish. 2007, 17, 581–613. [Google Scholar] [CrossRef]
- Walther, G.R.; Roques, A.; Hulme, P.E.; Sykes, M.T.; Pyšek, P.; Kühn, I.; Zobel, M.; Bacher, S.; Botta-Dukát, Z.; Bugmann, H.; et al. Alien species in a warmer world: Risks and opportunities. Trends Ecol. Evol. 2009, 24, 686–693. [Google Scholar] [CrossRef]
- Nunn, A.D.; Cowx, I.G.; Frear, P.A.; Harvey, J.P. Is water temperature an adequate predictor of recruitment success in cyprinid fish populations in lowland rivers? Freshw. Biol. 2003, 48, 579–588. [Google Scholar] [CrossRef]
- Johnson, A.C.; Acreman, M.C.; Dunbar, M.J.; Feist, S.W.; Giacomello, A.M.; Gozlan, R.E.; Hinsley, S.A.; Ibbotson, A.T.; Jarvie, H.P.; Jones, J.I.; et al. The British river of the future: How climate change and human activity might affect two contrasting river ecosystems in England. Sci. Total Environ. 2009, 407, 4787–4798. [Google Scholar] [CrossRef]
- Invasive Species Council. Inquiry into Ecosystem Decline in Victoria by the Environment and Planning Committee of the Victorian Parliament. Submission by the Invasive Species Council November 2020. Available online: https://www.parliament.vic.gov.au/images/stories/committees/SCEP/Ecosystem_Decline/submissions/S943_-_Invasive_Species_Council_Redacted.pdf (accessed on 1 February 2023).
- Sharma, S.; Vander Zanden, M.J.; Magnuson, J.J.; Lyons, J. Comparing climate change and species invasions as drivers of coldwater fish population extirpations. PLoS ONE 2011, 6, e22906. [Google Scholar] [CrossRef] [PubMed]
- Muhlfeld, C.C.; Kovach, R.P.; Jones, L.A.; Al-Chokhachy, R.; Boyer, M.C.; Leary, R.F.; Lowe, W.H.; Luikart, G.; Allendorf, F.W. Invasive hybridization in a threatened species is accelerated by climate change. Nat. Clim. Chang. 2014, 4, 620–624. [Google Scholar] [CrossRef]
- Gomez-Maldonado, S.; Camacho-Cervantes, M. Effect of a temperature gradient on the behaviour of an endangered Mexican topminnow and an invasive freshwater fish. Sci. Rep. 2022, 12, 20584. [Google Scholar] [CrossRef] [PubMed]
- DeWeber, J.T.; Baer, J.; Rösch, R.; Brinker, A. Turning summer into winter: Nutrient dynamics, temperature, density dependence and invasive species drive bioenergetic processes and growth of a keystone coldwater fish. Oikos 2022, 2022, e09316. [Google Scholar] [CrossRef]
- Avlijaš, S.; Mandrak, N.E.; Ricciardi, A. Effects of substrate and elevated temperature on the growth and feeding efficiency of an invasive cyprinid fish, Tench (Tinca tinca). Biol. Invasions 2022, 24, 2383–2397. [Google Scholar] [CrossRef]
- To, D.A.L.; Gomez, G.M.L.; Ramos, D.R.Y.; Palillo, C.D.V.; Go, N.K.C.; Gilles, A.S.; Briones, J.C.A.; Pavia, R.T.B. Invasiveness Risk Assessment of Non-native Freshwater Fish Species (Order: Siluriformes) for Lake Taal, Philippines. Philipp. J. Sci. 2022, 151, 833–841. [Google Scholar] [CrossRef]
- Cerrilla, C.; Afrika, J.; Impson, D.; Jordaan, M.S.; Kotze, N.; Paxton, B.R.; Reed, C.; Schumann, M.; van der Walt, J.A.; Shelton, J.M. Rapid population decline in one of the last recruiting populations of the endangered Clanwilliam sandfish (Labeo seeberi): The roles of climate change and non-native fish. Aquat. Conserv. Mar. Freshw. Ecosyst. 2022, 32, 781–796. [Google Scholar] [CrossRef]
- Souza, A.T.; Argillier, C.; Blabolil, P.; Děd, V.; Jarić, I.; Monteoliva, A.P.; Reynaud, N.; Ribeiro, F.; Ritterbusch, D.; Sala, P.; et al. Empirical evidence on the effects of climate on the viability of common carp (Cyprinus carpio) populations in European lakes. Biol. Invasions 2022, 24, 1213–1227. [Google Scholar] [CrossRef]
- Tytar, V.; Nekrasova, O.; Pupins, M.; Čeirāns, A.; Skute, A. Modelling the range expansion of pumpkinseed Lepomis gibbosus across Europe, with special focus on Latvia and Ukraine. North West J. Zool. 2021, 18, 143–150. [Google Scholar]
- Radtke, G.; Wolnicki, J.; Kapusta, A.; Przybylski, M.; Kaczkowski, Z. Critical thermal maxima of three small-bodied fish species (Cypriniformes) of different origin and protection status. Eur. Zool. J. 2022, 89, 1351–1361. [Google Scholar] [CrossRef]
- Mumladze, L.; Kuljanishvili, T.; Japoshvili, B.; Epitashvili, G.; Kalous, L.; Vilizzi, L.; Piria, M. Risk of invasiveness of non-native fishes in the South Caucasus biodiversity and geopolitical hotspot. NeoBiota 2022, 76, 109–133. [Google Scholar] [CrossRef]
- Tarkan, A.S.; Emiroğlu, Ö.; Aksu, S.A.D.İ.; Başkurt, S.; Aksu, İ.; Vilizzi, L.; Yoğurtçuoğlu, B. Coupling molecular and risk analysis to investigate the origin, distribution and potential impact of non-native species: An application to ruffe Gymnocephalus cernua in Turkey. Eur. Zool. J. 2022, 89, 109–121. [Google Scholar] [CrossRef]
- Jourdan, J.; Riesch, R.; Cunze, S. Off to new shores: Climate niche expansion in invasive mosquitofish (Gambusia spp.). Ecol. Evol. 2021, 11, 18369–18400. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, J.S.; Fraker, M.E.; Hood, J.M.; Frank, K.T.; DuFour, M.R.; Gorman, A.M.; Ludsin, S.A. Functional traits reveal the dominant drivers of long-term community change across a North American Great Lake. Glob. Chang. Biol. 2021, 27, 6232–6251. [Google Scholar] [CrossRef]
- Lin, L.; Deng, W.; Huang, X.; Liu, Y.; Huang, L.; Kang, B. How fish traits and functional diversity respond to environmental changes and species invasion in the largest river in Southeastern China. PeerJ 2021, 9, e11824. [Google Scholar] [CrossRef]
- Pirroni, S.; de Pennafort Dezen, L.; Santi, F.; Riesch, R. Comparative gut content analysis of invasive mosquitofish from Italy and Spain. Ecol. Evol. 2021, 11, 4379–4398. [Google Scholar] [CrossRef]
- Costa, A.C.; Balibrea, A.; Raposeiro, P.M.; Santos, S.; Souto, M.; Gonçalves, V. Non-indigenous and invasive freshwater species on the Atlantic Islands of the Azores Archipelago. Front. Ecol. Evol. 2021, 9, 631214. [Google Scholar] [CrossRef]
- Carosi, A.; Ghetti, L.; Lorenzoni, M. The role of climate changes in the spread of freshwater fishes: Implications for alien cool and warm-water species in a Mediterranean basin. Water 2021, 13, 347. [Google Scholar] [CrossRef]
- Bounas, A.; Catsadorakis, G.; Koutseri, I.; Nikolaou, H.; Nicolas, D.; Malakou, M.; Crivelli, A.J. Temporal trends and determinants of fish biomass in two contrasting natural lake systems: Insights from a spring long-term monitoring scheme. Knowl. Manag. Aquat. Ecosyst. 2021, 422, 28. [Google Scholar] [CrossRef]
- Rubenson, E.S.; Olden, J.D. An invader in salmonid rearing habitat: Current and future distributions of smallmouth bass (Micropterus dolomieu) in the Columbia River Basin. Can. J. Fish. Aquat. Sci. 2020, 77, 314–325. [Google Scholar] [CrossRef]
- Zięba, G.; Vilizzi, L.; Copp, G.H. How likely is Lepomis gibbosus to become invasive in Poland under conditions of climate warming? Acta Ichthyol. Piscat. 2020, 50, 35–51. [Google Scholar] [CrossRef]
- Liu, C.; Comte, L.; Xian, W.; Chen, Y.; Olden, J.D. Current and projected future risks of freshwater fish invasions in China. Ecography 2019, 42, 2074–2083. [Google Scholar] [CrossRef]
- Jia, Y.; Kennard, M.J.; Liu, Y.; Sui, X.; Chen, Y.; Li, K.; Wang, G.; Chen, Y. Understanding invasion success of Pseudorasbora parva in the Qinghai-Tibetan Plateau: Insights from life-history and environmental filters. Sci. Total Environ. 2019, 694, 133739. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Liu, Y.; Chen, K.; Sun, H.; Chen, Y. Climate, habitat and human disturbance driving the variation of life-history traits of the invasive goldfish Carassius auratus (Linnaeus, 1758) in a Tibetan Plateau river. Aquat. Invasions 2019, 14, 724–737. [Google Scholar] [CrossRef]
- Radinger, J.; Alcaraz-Hernández, J.D.; García-Berthou, E. Environmental filtering governs the spatial distribution of alien fishes in a large, human-impacted Mediterranean river. Divers. Distrib. 2019, 25, 701–714. [Google Scholar] [CrossRef]
- Shelton, J.M.; Weyl, O.L.; Esler, K.J.; Paxton, B.R.; Impson, N.D.; Dallas, H.F. Temperature mediates the impact of non-native rainbow trout on native freshwater fishes in South Africa’s Cape Fold Ecoregion. Biol. Invasions 2018, 20, 2927–2944. [Google Scholar] [CrossRef]
- Rubenson, E.S.; Olden, J.D. Dynamism in the upstream invasion edge of a freshwater fish exposes range boundary constraints. Oecologia 2017, 184, 453–467. [Google Scholar] [CrossRef]
- Rolls, R.J.; Hayden, B.; Kahilainen, K.K. Conceptualising the interactive effects of climate change and biological invasions on subarctic freshwater fish. Ecol. Evol. 2017, 7, 4109–4128. [Google Scholar] [CrossRef]
- Tarkan, A.S.; Sarı, H.M.; İlhan, A.; Kurtul, I.; Vilizzi, L. Risk screening of non-native and translocated freshwater fish species in a Mediterranean-type shallow lake: Lake Marmara (West Anatolia). Zool. Middle East 2017, 63, 48–57. [Google Scholar] [CrossRef]
- Copp, G.H.; Britton, J.R.; Guo, Z.; Ronni Edmonds-Brown, V.; Pegg, J.; Vilizzi, L.; Davison, P.I. Trophic consequences of non-native pumpkinseed Lepomis gibbosus for native pond fishes. Biol. Invasions 2017, 19, 25–41. [Google Scholar] [CrossRef]
- Crook, D.A.; Lowe, W.H.; Allendorf, F.W.; Erős, T.; Finn, D.S.; Gillanders, B.M.; Hadwen, W.L.; Harrod, C.; Hermoso, V.; Jennings, S.; et al. Human effects on ecological connectivity in aquatic ecosystems: Integrating scientific approaches to support management and mitigation. Sci. Total Environ. 2015, 534, 52–64. [Google Scholar] [CrossRef] [PubMed]
- Rabitsch, W.; Milasowszky, N.; Nehring, S.; Wiesner, C.; Wolter, C.; Essl, F. The times are changing: Temporal shifts in patterns of fish invasions in central European fresh waters. J. Fish Biol. 2013, 82, 17–33. [Google Scholar] [CrossRef] [PubMed]
- Kornis, M.S.; Mercado-Silva, N.; Vander Zanden, M.J. Twenty years of invasion: A review of round goby Neogobius melanostomus biology, spread and ecological implications. J. Fish Biol. 2012, 80, 235–285. [Google Scholar] [CrossRef] [PubMed]
- Britton, J.R.; Cucherousset, J.; Davies, G.D.; Godard, M.J.; Copp, G.H. Non-native fishes and climate change: Predicting species responses to warming temperatures in a temperate region. Freshw. Biol. 2010, 55, 1130–1141. [Google Scholar] [CrossRef]
- Benejam, L.; Alcaraz, C.; Sasal, P.; Simon-Levert, G.; García-Berthou, E. Life history and parasites of the invasive mosquitofish (Gambusia holbrooki) along a latitudinal gradient. Biol. Invasions 2009, 11, 2265–2277. [Google Scholar] [CrossRef]
- Rahel, F.J.; Olden, J.D. Assessing the effects of climate change on aquatic invasive species. Conserv. Biol. 2008, 22, 521–533. [Google Scholar] [CrossRef]
- Franchi, E.; Carosi, A.; Ghetti, L.; Giannetto, D.; Pedicillo, G.; Pompei, L.; Lorenzoni, M. Changes in the fish community of the upper Tiber River after construction of a hydro-dam. J. Limnol. 2014, 73, 876. [Google Scholar] [CrossRef]
- Drouineau, H.; Carter, C.; Rambonilaza, M.; Beaufaron, G.; Bouleau, G.; Gassiat, A.; Lambert, P.; le Floch, S.; Tétard, S.; De Oliveira, E. River continuity restoration and diadromous fishes: Much more than an ecological issue. Environ. Manag. 2018, 61, 671–686. [Google Scholar] [CrossRef]
- Merg, M.L.; Dézerald, O.; Kreutzenberger, K.; Demski, S.; Reyjol, Y.; Usseglio-Polatera, P.; Belliard, J.C. Modeling diadromous fish loss from historical data: Identification of anthropogenic drivers and testing of mitigation scenarios. PLoS ONE 2020, 15, e0236575. [Google Scholar] [CrossRef]
- Carosi, A.; Ghetti, L.; La Porta, G.; Lorenzoni, M. Ecological effects of the European barbel Barbus barbus (L., 1758) (Cyprinidae) invasion on native barbel populations in the Tiber River basin (Italy). Eur. Zool. J. 2017, 84, 420–435. [Google Scholar] [CrossRef]
- Carosi, A.; Ghetti, L.; Forconi, A.; Lorenzoni, M. Fish community of the river Tiber basin (Umbria-Italy): Temporal changes and possible threats to native biodiversity. Knowl. Manag. Aquat. Ecosyst. 2015, 416, 22. [Google Scholar] [CrossRef]
- Isaak, D.J.; Rieman, B.E. Stream isotherm shifts from climate change and implications for distributions of ectothermic organisms. Glob. Chang. Biol. 2013, 19, 742–751. [Google Scholar] [CrossRef] [PubMed]
- Marr, S.M.; Olden, J.D.; Leprieur, F.; Arismendi, I.; Ćaleta, M.; Morgan, D.L.; Nocita, A.; Šanda, R.; Tarkan, A.S.; García-Berthou, E. A global assessment of freshwater fish introductions in mediterranean-climate regions. Hydrobiologia 2013, 719, 317–329. [Google Scholar] [CrossRef]
- Sauz-Sánchez, J.D.J.; Rodiles-Hernández, R.; Andrade-Velázquez, M.; Mendoza-Carranza, M. Modelling the potential distribution of two tropical freshwater fish species under climate change scenarios. Aquat. Conserv. Mar. Freshw. Ecosys. 2021, 31, 2737–2751. [Google Scholar] [CrossRef]
- Nickus, U.; Bishop, K.; Erlandsson, M.; Evans, C.D.; Forsius, M.; Laudon, H.; Livingstone, D.M.; Monteith, D.; Thies, H. Direct impacts of climate change on freshwater ecosystems. In Climate Change Impacts on Freshwater Ecosystems, 1st ed.; Kernan, M., Battarbee, R., Moss, B., Eds.; Blackwell Publishing Ltd.: Hoboken, NJ, USA, 2010; pp. 38–64. [Google Scholar] [CrossRef]
- Jeppesen, E.; Kronvang, B.; Jørgensen, T.B.; Larsen, S.E.; Andersen, H.E.; Søndergaard, M.; Liboriussen, L.; Bjerring, R.; Johansson, L.; Trolle, D.; et al. Recent climate-induced changes in freshwaters in Denmark. In Climatic Change and Global Warming of Inland Waters: Impacts and Mitigation for Ecosystems and Societies, 1st ed.; Goldman, C.R., Kumagai, M., Robarts, R.D., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2013; pp. 155–171. [Google Scholar]
- Filipe, A.F.; Lawrence, J.E.; Bonada, N. Vulnerability of stream biota to climate change in mediterranean climate regions: A synthesis of ecological responses and conservation challenges. Hydrobiologia 2013, 719, 331–351. [Google Scholar] [CrossRef]
- Carosi, A.; Padula, R.; Ghetti, L.; Lorenzoni, M. Endemic freshwater fish range shifts related to global climate changes: A long-term study provides some observational evidence for the Mediterranean area. Water 2019, 11, 2349. [Google Scholar] [CrossRef]
- Mulholland, P.J.; Best, G.R.; Coutant, C.C.; Hornberger, G.M.; Meyer, J.L.; Robinson, P.J.; Stenberg, J.R.; Turner, R.E.; Vera-Herrera, F.; Wetzel, R.G. Effects of climate change on freshwater ecosystems of the south-eastern United States and the Gulf Coast of Mexico. Hydrol. Process. 1997, 11, 949–970. [Google Scholar] [CrossRef]
- Danladi Bello, A.A.; Hashim, N.B.; Mohd Haniffah, M.R. Predicting impact of climate change on water temperature and dissolved oxygen in tropical rivers. Climate 2017, 5, 58. [Google Scholar] [CrossRef]
- Copp, G.H.; Russell, I.C.; Peeler, E.J.; Gherardi, F.; Tricarico, E.; MacLeod, A.; Cowx, I.G.; Nunn, A.D.; Occhipinti Ambrogi, A.; Savini, D.; et al. European Non-native Species in Aquaculture Risk Analysis Scheme–a summary of assessment protocols and decision support tools for use of alien species in aquaculture. Fish. Manag. Ecol. 2016, 23, 1–11. [Google Scholar] [CrossRef]
- Buisson, L.; Thuiller, W.; Lek, S.; Lim, P.U.Y.; Grenouillet, G. Climate change hastens the turnover of stream fish assemblages. Glob. Chang. Biol. 2008, 14, 2232–2248. [Google Scholar] [CrossRef]
- Fobert, E.; Fox, M.G.; Ridgway, M.; Copp, G.H. Heated competition: How climate change will affect non-native pumpkinseed Lepomis gibbosus and native perch Perca fluviatilis interactions in the UK. J. Fish Biol. 2011, 79, 1592–1607. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
A conceptual scheme of the potential effects of climate changes on inland waters, the possible responses in terms of alien fish invasions, and their relative negative impacts on native fish biodiversity.
Figure 1.
A conceptual scheme of the potential effects of climate changes on inland waters, the possible responses in terms of alien fish invasions, and their relative negative impacts on native fish biodiversity.

Figure 2.
Conceptual diagram of the alien freshwater fish invasion triangle (Adapted from Perkins et al. [27]).
Figure 2.
Conceptual diagram of the alien freshwater fish invasion triangle (Adapted from Perkins et al. [27]).

Figure 3.
Influence of climate change on all the phases of a successful alien freshwater fish invasion process. Adapted with permission from Walther et al. [36], Copyright 2023 from Elsevier, and from Invasive Species Council [39].


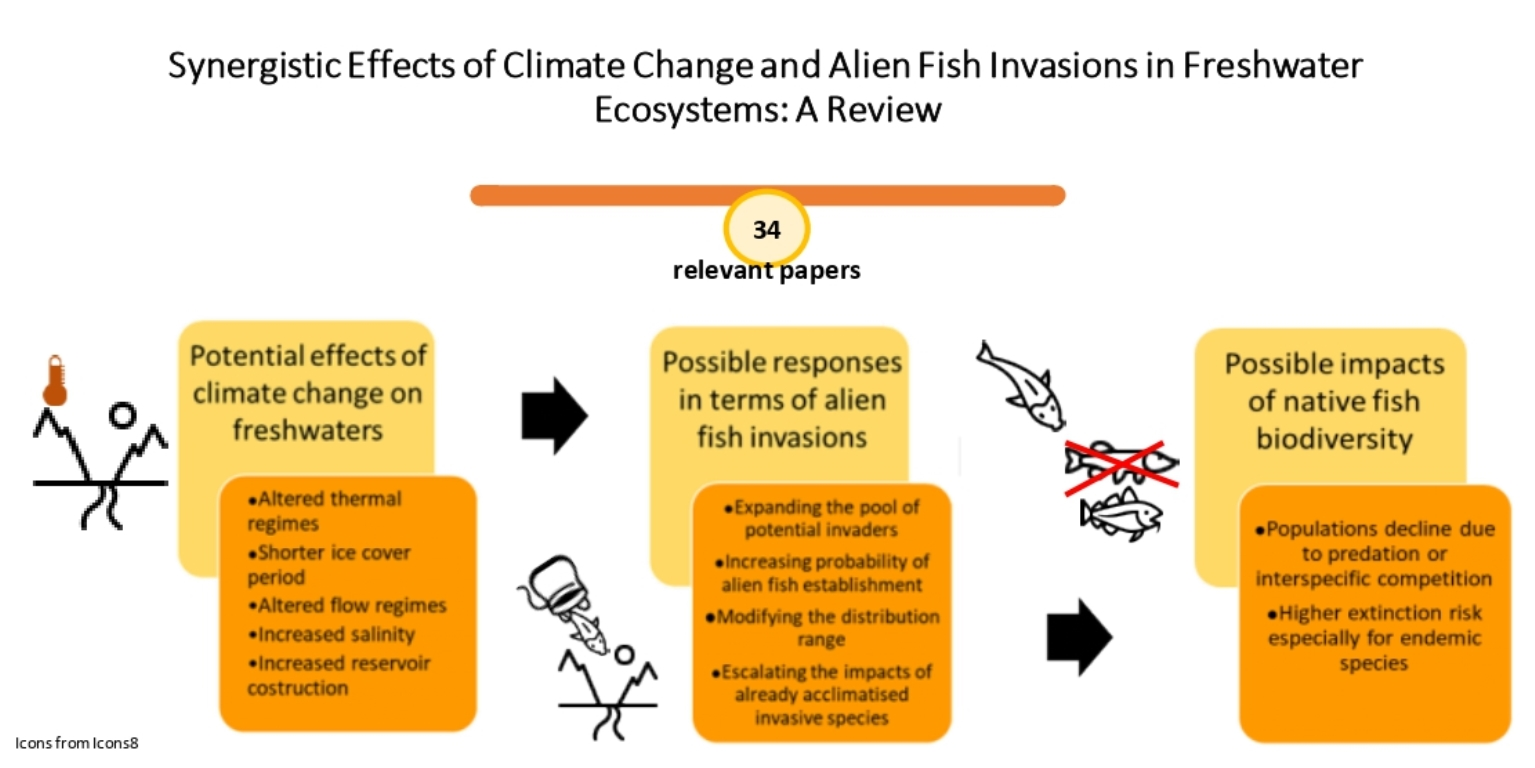{kind=link}
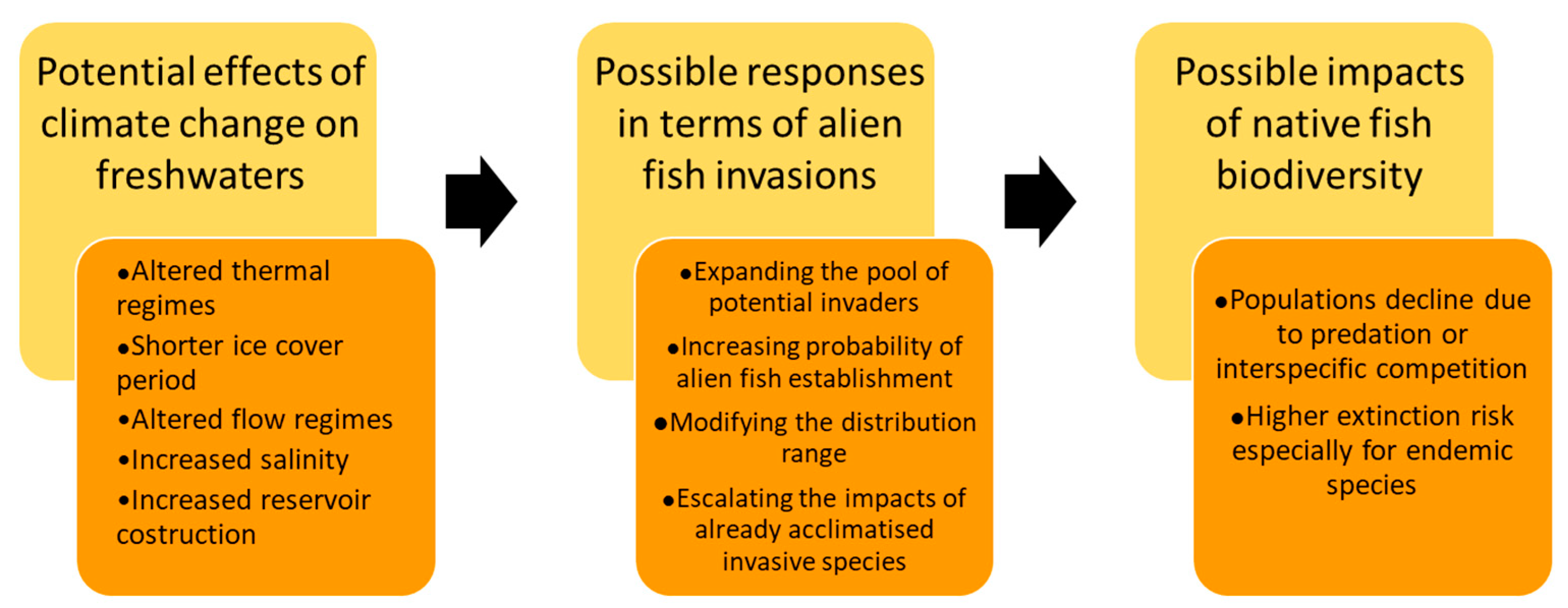{kind=link}
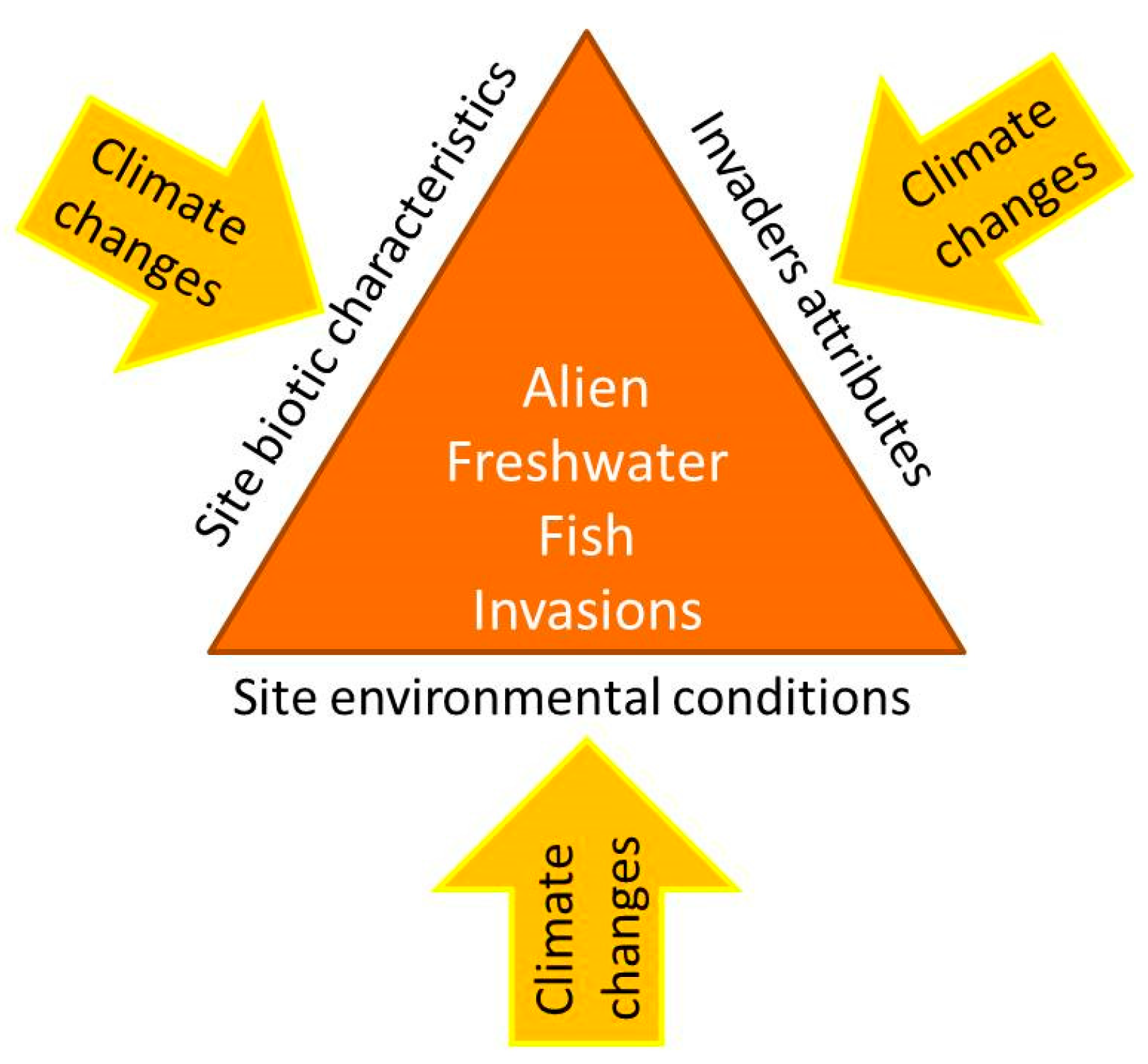{kind=link}
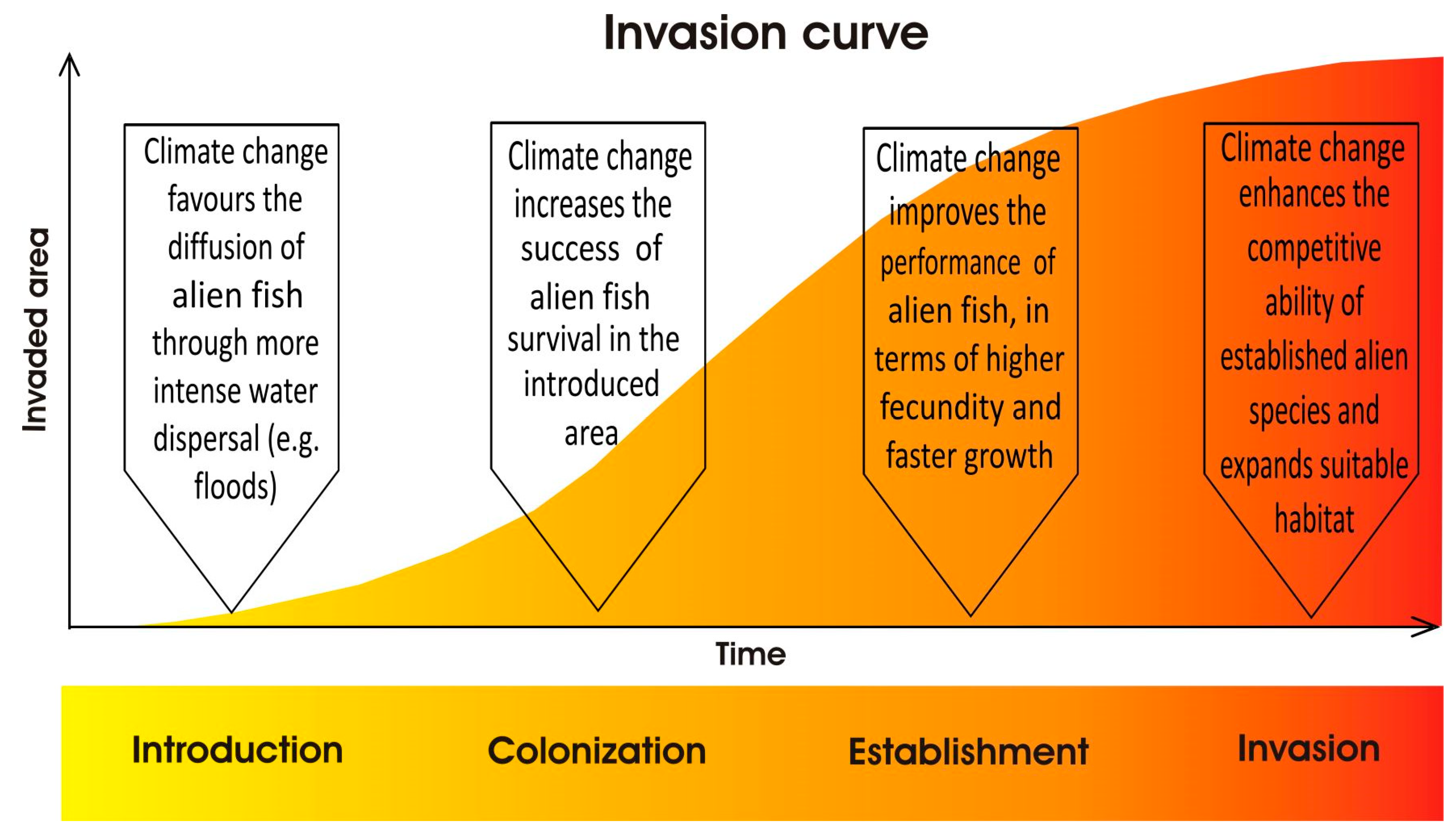{kind=link}
Table 1.
List of selected papers, broken down by climatic area, geographic region, target species, and methodological approach.
Table 1.
List of selected papers, broken down by climatic area, geographic region, target species, and methodological approach.
| Climatic Area | Geographic Area | Reference | Target Species | Methodological Approach |
|---|---|---|---|---|
| Tropical wet/semiarid | Central Mexico | Gomez-Maldonado and Camacho-Cervantes (2022) [42] | Skiffia bilineata (Bean, 1887) Poecilia reticulata Peters, 1859 | Behavioral observations in aquarium |
| Temperate | Germany | DeWeber et al. (2022) [43] | Coregonus wartmanni (Bloch, 1784) | Bioenergetics and growth modeling |
| Continental | North America | Avlijaš et al. (2022) [44] | Tinca tinca (L., 1758) | Growth and functional response experiments |
| Tropical | Philippines | To et al. (2022) [45] | Clarias batrachus (L., 1758) | Aquatic Species Invasiveness Screening kit (AS-ISK) |
| Mediterranean | South Africa | Cerrilla et al. (2022) [46] | Labeo seeberi Gilchrist & Thompson, 1911 | Observational data analysis |
| Continental, Temperate oceanic, Mediterranean | Europe | Souza et al. (2022) [47] | Cyprinus carpio L., 1758 | Observational data analysis |
| Temperate | Ukraine, Latvia | Tytar et al. (2022) [48] | Lepomis gibbosus (L., 1758) | Ecological niches models |
| Temperate | Poland | Radtke et al. (2022) [49] | Eupallasella percnurus (Pallas, 1814), Leucaspius delineates (Heckel, 1843), Pseudorasbora parva (Temminck & Schlegel, 1846) | Critical Thermal Method in aquarium |
| Continental mesophilic | South Caucasus | Mumladze et al. (2022) [50] | 32 alien species | AS-ISK |
| Temperate, Mediterranean | Turkey | Tarkan et al. (2022) [51] | Gymnocephalus cernua (L., 1758) | AS-ISK, mtDNA barcoding |
| - | Global | Jourdan et al. (2021) [52] | Gambusia spp. | Species distribution models |
| Humid continental | North America | Sinclair et al. (2021) [53] | Lake Eire fish community | Functional trait compositional changes |
| Humid subtropical, Oceanic | Southeastern China | Lin et al. (2021) [54] | Min River fish community | Functional diversity changes |
| Mediterranean | Italy, Spain | Pirroni et al. (2021) [55] | Gambusia holbrooki Girard, 1859 | Feeding ecology |
| Humid subtropical, Mediterranean | Azores Archipelago | Costa et al. (2021) [56] | 132 alien freshwater species | Literature review |
| Mediterranean | Italy | Carosi et al. (2021) [57] | Barbus barbus(L., 1758), Gobio gobio (L., 1758), Padogobius bonelli (Bonaparte, 1846), P. parva | Observational data analysis |
| Temperate, Mediterranean | Southeastern Europe | Bounas et al. (2021) [58] | Prespa Lakes fish community | Structural change models, random forest |
| Continental, Temperate oceanic | North America | Rubenson and Olden (2020) [59] | Micropterus dolomieu Lacepède, 1802 | Species distribution models, environmental DNA |
| Temperate | UK, France, Poland | Zięba et al. (2020) [60] | L. gibbosus | AS-ISK |
| Tropical, Subarctic, Alpine, Temperate, Arid | China | Liu et al. (2019) [61] | Freshwater fish | Species distribution model |
| Alpine | Tibetan Plateau | Jia et al. (2019) [62] | P. parva | Linear mixed-effects models, random forest |
| Alpine | Tibetan Plateau | Jia et al. (2019) [63] | Carassius auratus (L., 1758) | Multiple regression, hierarchical partitioning models |
| Alpine, Semiarid, Mediterranean | Iberian peninsula | Radinger et al. (2019) [64] | Freshwater fish | Boosted regression tree, species distribution model |
| Mediterranean | South Africa | Shelton et al. (2018) [65] | Oncorhynchus mykiss (Walbaum, 1792) | Distance-based Linear Models |
| Continental, Temperate oceanic | North America | Rubenson and Olden (2017) [66] | M. dolomieu | Telemetry analysis |
| Subpolar | Subarctic regions | Rolls et al. (2017) [67] | Freshwater fish | Literature review |
| Mediterranean | West Anatolia | Tarkan et al. (2017) [68] | 35 alien species in Marmara Lake | AS-ISK |
| Temperate | UK | Copp et al. (2017) [69] | L.gibbosus | Stable isotope analysis |
| - | Global | Crook et al. (2015) [70] | Freshwater animals | Literature review |
| Temperate, Continental | Germany, Austria | Rabitsch et al. (2013) [71] | 15 alien species | Species spatial distribution |
| Temperate, Continental | North America, Eurasia, Baltic Sea | Kornis et al. (2012) [72] | Neogobius melanostomus (Pallas, 1814) | Literature review |
| Temperate | UK | Britton et al. (2010) [73] | C. carpio, Silurus glanis L., 1758 | Climate models, spatial distribution models |
| Mediterranean | France, Spain | Benejam et al. (2009) [74] | G. holbrooki | Observational data analysis |
| - | Global | Rahel and Olden (2008) [75] | Alien species | Literature review |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Carosi, A.; Lorenzoni, F.; Lorenzoni, M. Synergistic Effects of Climate Change and Alien Fish Invasions in Freshwater Ecosystems: A Review. Fishes 2023, 8, 486. https://doi.org/10.3390/fishes8100486
AMA Style
Carosi A, Lorenzoni F, Lorenzoni M. Synergistic Effects of Climate Change and Alien Fish Invasions in Freshwater Ecosystems: A Review. Fishes. 2023; 8(10):486. https://doi.org/10.3390/fishes8100486
Chicago/Turabian StyleCarosi, Antonella, Francesca Lorenzoni, and Massimo Lorenzoni. 2023. "Synergistic Effects of Climate Change and Alien Fish Invasions in Freshwater Ecosystems: A Review" Fishes 8, no. 10: 486. https://doi.org/10.3390/fishes8100486