
In Case of Fire, Escape or Die: A Trait-Based Approach for Identifying Animal Species Threatened by Fire

 , , , , , ,
, , , , , ,  , , , , ,
, , , , ,  , , , , and
, , , , and
Abstract
:1. Introduction
2. Methods
2.1. Fire Exposure
2.2. Fire Sensitivity
3. Results
3.1. Sensitivity
3.1.1. During Fire
Dormancy
Escape Decision
Habitat Use
Mobility
Morphology
Nest Substrate
Reproductive Cycles
Sensory Detection of Fire Cues
Social Organization
3.1.2. Post Fire
Behavioral Plasticity
Dormancy
Endogenous Circadian Rhythms
Mobility
Morphology
3.2. Fire Vulnerability Index
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Beerling, D.J.; Osborne, C.P. The origin of the savanna biome. Glob. Chang. Biol. 2006, 12, 2023–2031. [Google Scholar] [CrossRef]
- Simon, M.F.; Grether, R.; de Queiroz, L.P.; Skema, C.; Pennington, R.T.; Hughes, C.E. Recent assembly of the Cerrado, a neotropical plant diversity hotspot, by in situ evolution of adaptations to fire. Proc. Natl. Acad. Sci. USA 2009, 106, 20359–20364. [Google Scholar] [CrossRef] [Green Version]
- Balch, J.K.; Bradley, B.A.; Abatzoglou, J.T.; Nagy, R.C.; Fusco, E.J.; Mahood, A.L. Human-started wildfires expand the fire niche across the United States. Proc. Natl. Acad. Sci. USA 2017, 114, 2946–2951. [Google Scholar] [CrossRef] [Green Version]
- Le Page, Y.; Morton, D.; Hartin, C.; Bond-Lamberty, B.; Pereira, J.M.C.; Hurtt, G.; Asrar, G. Synergy between land use and climate change increases future fire risk in Amazon forests. Earth Syst. Dyn. 2017, 8, 1237–1246. [Google Scholar] [CrossRef] [Green Version]
- Silva Junior, C.H.L.; Aragão, L.E.O.C.; Fonseca, M.G.; Almeida, C.T.; Vedovato, L.B.; Anderson, L.O. Deforestation-Induced Fragmentation Increases Forest Fire Occurrence in Central Brazilian Amazonia. Forests 2018, 9, 305. [Google Scholar] [CrossRef] [Green Version]
- Pyne, S.J. From Pleistocene to Pyrocene: Fire Replaces Ice. Earth’s Futur. 2020, 8, e2020EF001722. [Google Scholar] [CrossRef]
- Nimmo, D.G.; Carthey, A.J.R.; Jolly, C.J.; Blumstein, D.T. Welcome to the Pyrocene: Animal survival in the age of megafire. Glob. Chang. Biol. 2021, 27, 5684–5693. [Google Scholar] [CrossRef] [PubMed]
- Brennan, K.E.C.; Moir, M.L.; Wittkuhn, R.S. Fire refugia: The mechanism governing animal survivorship within a highly flammable plant. Austral Ecol. 2011, 36, 131–141. [Google Scholar] [CrossRef]
- Nimmo, D.G.; Avitabile, S.; Banks, S.C.; Bird, R.B.; Callister, K.; Clarke, M.F.; Dickman, C.R.; Doherty, T.S.; Driscoll, D.A.; Greenville, A.C.; et al. Animal movements in fire-prone landscapes. Biol. Rev. 2018, 94, 981–998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pausas, J.G. Generalized fire response strategies in plants and animals. Oikos 2018, 128, 147–153. [Google Scholar] [CrossRef] [Green Version]
- Jolly, C.J.; Dickman, C.R.; Doherty, T.S.; van Eeden, L.M.; Geary, W.L.; Legge, S.M.; Woinarski, J.C.Z.; Nimmo, D.G. Animal mortality during fire. Glob. Chang. Biol. 2022, 28, 2053–2065. [Google Scholar] [CrossRef] [PubMed]
- Pausas, J.G.; Parr, C.L. Towards an understanding of the evolutionary role of fire in animals. Evol. Ecol. 2018, 32, 113–125. [Google Scholar] [CrossRef]
- Tomas, W.M.; Berlinck, C.N.; Chiaravalloti, R.M.; Faggioni, G.P.; Strüssmann, C.; Libonati, R.; Abrahão, C.R.; Alvarenga, G.D.V.; Bacellar, A.E.D.F.; Batista, F.R.D.Q.; et al. Distance sampling surveys reveal 17 million vertebrates directly killed by the 2020′s wildfires in the Pantanal, Brazil. Sci. Rep. 2021, 11, 23547. [Google Scholar] [CrossRef] [PubMed]
- Engstrom, R.T. First-Order Fire Effects on Animals: Review and Recommendations. Fire Ecol. 2010, 6, 115–130. [Google Scholar] [CrossRef]
- Meddens, A.J.H.; A Kolden, C.; A Lutz, J.; Smith, A.M.S.; Cansler, C.A.; Abatzoglou, J.T.; Meigs, G.W.; Downing, W.M.; A Krawchuk, M. Fire Refugia: What Are They, and Why Do They Matter for Global Change? Bioscience 2018, 68, 944–954. [Google Scholar] [CrossRef] [Green Version]
- Martinez, A.; Meddens, A.; Kolden, C.; Hudak, A. An Assessment of Fire Refugia Importance Criteria Ranked by Land Managers. Fire 2019, 2, 27. [Google Scholar] [CrossRef] [Green Version]
- Steenvoorden, J.; Meddens, A.J.H.; Martinez, A.J.; Foster, L.J.; Kissling, W.D. The potential importance of unburned islands as refugia for the persistence of wildlife species in fire-prone ecosystems. Ecol. Evol. 2019, 9, 8800–8812. [Google Scholar] [CrossRef]
- Costa, B.M.; Pantoja, D.L.; Sousa, H.C.; de Queiroz, T.A.; Colli, G.R. Long-term, fire-induced changes in habitat structure and microclimate affect Cerrado lizard communities. Biodivers. Conserv. 2019, 29, 1659–1681. [Google Scholar] [CrossRef]
- DeSouza, O.; Albuquerque, L.B.; Tonello, V.M.; Pinto, L.P.; Junior, R.R. Effects of fire on termite generic richness in a savanna-like ecosystem (‘Cerrado’) of Central Brazil. Sociobiology 2003, 42, 639–649. [Google Scholar]
- Adger, W.N. Vulnerability. Glob. Environ. Chang. 2006, 16, 268–281. [Google Scholar] [CrossRef]
- Weis, S.W.M.; Agostini, V.N.; Roth, L.M.; Gilmer, B.; Schill, S.R.; Knowles, J.E.; Blyther, R. Assessing vulnerability: An integrated approach for mapping adaptive capacity, sensitivity, and exposure. Clim. Chang. 2016, 136, 615–629. [Google Scholar] [CrossRef] [Green Version]
- Schumacher, V.; Setzer, A.; Saba, M.M.; Naccarato, K.P.; Mattos, E.; Justino, F. Characteristics of lightning-caused wildfires in central Brazil in relation to cloud-ground and dry lightning. Agric. For. Meteorol. 2022, 312, 108723. [Google Scholar] [CrossRef]
- dos Santos, A.C.; Montenegro, S.D.R.; Ferreira, M.C.; Barradas, A.C.S.; Schmidt, I.B. Managing fires in a changing world: Fuel and weather determine fire behavior and safety in the neotropical savannas. J. Environ. Manag. 2021, 289, 112508. [Google Scholar] [CrossRef]
- Cochrane, M.A. Fire science for rainforests. Nature 2003, 421, 913–919. [Google Scholar] [CrossRef] [PubMed]
- Robinson, N.M.; Leonard, S.W.; Ritchie, E.; Bassett, M.; Chia, E.K.; Buckingham, S.; Gibb, H.; Bennett, A.F.; Clarke, M.F. Review: Refuges for fauna in fire-prone landscapes: Their ecological function and importance. J. Appl. Ecol. 2013, 50, 1321–1329. [Google Scholar] [CrossRef]
- Leonard, S.W.; Bennett, A.F.; Clarke, M.F. Determinants of the occurrence of unburnt forest patches: Potential biotic refuges within a large, intense wildfire in south-eastern Australia. For. Ecol. Manag. 2014, 314, 85–93. [Google Scholar] [CrossRef]
- Oliveira, S.L.J.; Campagnolo, M.; Price, O.; Edwards, A.C.; Russell-Smith, J.; Pereira, J.M.C. Ecological Implications of Fine-Scale Fire Patchiness and Severity in Tropical Savannas of Northern Australia. Fire Ecol. 2015, 11, 10–31. [Google Scholar] [CrossRef] [Green Version]
- Hantson, S.; Andela, N.; Goulden, M.L.; Randerson, J.T. Human-ignited fires result in more extreme fire behavior and ecosystem impacts. Nat. Commun. 2022, 13, 2717. [Google Scholar] [CrossRef]
- Abatzoglou, J.T.; Williams, A.P. Impact of anthropogenic climate change on wildfire across western US forests. Proc. Natl. Acad. Sci. USA 2016, 113, 11770–11775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcês, A.; Pires, I. The Hell of Wildfires: The Impact on Wildlife and Its Conservation and the Role of the Veterinarian. Conservation 2023, 3, 96–108. [Google Scholar] [CrossRef]
- Gutiérrez, J.; De Miguel, J. Fires in nature: A review of the challenges for wild animals. Eur. J. Ecol. 2021, 7, 95–117. [Google Scholar] [CrossRef]
- McLauchlan, K.K.; Higuera, P.E.; Miesel, J.; Rogers, B.M.; Schweitzer, J.; Shuman, J.K.; Tepley, A.J.; Varner, J.M.; Veblen, T.T.; Adalsteinsson, S.A.; et al. Fire as a fundamental ecological process: Research advances and frontiers. J. Ecol. 2020, 108, 2047–2069. [Google Scholar] [CrossRef]
- Pivello, V.R.; Vieira, I.; Christianini, A.V.; Ribeiro, D.B.; Menezes, L.D.S.; Berlinck, C.N.; Melo, F.P.; Marengo, J.A.; Tornquist, C.G.; Tomas, W.M.; et al. Understanding Brazil’s catastrophic fires: Causes, consequences and policy needed to prevent future tragedies. Perspect. Ecol. Conserv. 2021, 19, 233–255. [Google Scholar] [CrossRef]
- Zylstra, P. Quantifying the direct fire threat to a critically endangered arboreal marsupial using biophysical, mechanistic modelling. Austral Ecol. 2022, 48, 266–288. [Google Scholar] [CrossRef]
- Sanderfoot, O.V.; Bassing, S.B.; Brusa, J.L.; Emmet, R.L.; Gillman, S.J.; Swift, K.; Gardner, B. A review of the effects of wildfire smoke on the health and behavior of wildlife. Environ. Res. Lett. 2021, 16, 123003. [Google Scholar] [CrossRef]
- Recher, H.F.; Lunney, D.; Matthews, A. Small mammal populations in a eucalypt forest affected by fire and drought. I. Long-term patterns in an era of climate change. Wildl. Res. 2009, 36, 143–158. [Google Scholar] [CrossRef]
- Doty, A.C.; Currie, S.E.; Stawski, C.; Geiser, F. Can bats sense smoke during deep torpor? Physiol. Behav. 2018, 185, 31–38. [Google Scholar] [CrossRef]
- Geiser, F.; Stawski, C.; Doty, A.C.; E Cooper, C.; Nowack, J. A burning question: What are the risks and benefits of mammalian torpor during and after fires? Conserv. Physiol. 2018, 6, coy057. [Google Scholar] [CrossRef]
- Mendyk, R.W.; Weisse, A.; Fullerton, W. A wake-up call for sleepy lizards: The olfactory-driven response of Tiliqua rugosa (Reptilia: Squamata: Sauria) to smoke and its implications for fire avoidance behavior. J. Ethol. 2019, 38, 161–166. [Google Scholar] [CrossRef]
- Moberg, G.P.; Mench, J.A. The Biology of Animal Stress: Basic Principles and Implications for Animal Welfare; CABI Publishing: New York, NY, USA, 2000. [Google Scholar] [CrossRef]
- Fagan, W.F.; Lewis, M.A.; Auger-Méthé, M.; Avgar, T.; Benhamou, S.; Breed, G.; LaDage, L.; Schlägel, U.E.; Tang, W.-W.; Papastamatiou, Y.P.; et al. Spatial memory and animal movement. Ecol. Lett. 2013, 16, 1316–1329. [Google Scholar] [CrossRef]
- Branco, T.; Redgrave, P. The Neural Basis of Escape Behavior in Vertebrates. Annu. Rev. Neurosci. 2020, 43, 417–439. [Google Scholar] [CrossRef] [Green Version]
- Massman, W.J.; Frank, J.M.; Mooney, S.J. Advancing Investigation and Physical Modeling of First-Order Fire Effects on Soils. Fire Ecol. 2010, 6, 36–54. [Google Scholar] [CrossRef]
- Cane, J.H.; Neff, J.L. Predicted fates of ground-nesting bees in soil heated by wildfire: Thermal tolerances of life stages and a survey of nesting depths. Biol. Conserv. 2011, 144, 2631–2636. [Google Scholar] [CrossRef]
- Love, B.G.; Cane, J.H. Limited direct effects of a massive wildfire on its sagebrush steppe bee community. Ecol. Èntomol. 2016, 41, 317–326. [Google Scholar] [CrossRef]
- Jordaan, P.R.; Steyl, J.C.A.; Hanekom, C.C.; Combrink, X. Fire-associated reptile mortality in Tembe Elephant Park, South Africa. Fire Ecol. 2020, 16, 4–9. [Google Scholar] [CrossRef]
- Costa, B.M.; Pantoja, D.L.; Vianna, M.C.M.; Colli, G.R. Direct and Short-Term Effects of Fire on Lizard Assemblages from a Neotropical Savanna Hotspot. J. Herpetol. 2013, 47, 502–510. [Google Scholar] [CrossRef]
- Miranda, H.S.; Sato, M.N.; Neto, W.N.; Aires, F.S. Fires in the cerrado, the Brazilian savanna. In Tropical Fire Ecology; Springer: Heidelberg, Germany, 2009; pp. 427–450. [Google Scholar] [CrossRef]
- Rodrigues, C.A.; Zirondi, H.L.; Fidelis, A. Fire frequency affects fire behavior in open savannas of the Cerrado. For. Ecol. Manag. 2021, 482, 118850. [Google Scholar] [CrossRef]
- Kauf, Z.; Damsohn, W.; Fangmeier, A. Do relationships between leaf traits and fire behaviour of leaf litter beds persist in time? PLoS ONE 2018, 13, e0209780. [Google Scholar] [CrossRef] [Green Version]
- Buckingham, S.; Murphy, N.; Gibb, H. Effects of fire severity on the composition and functional traits of litter-dwelling macroinvertebrates in a temperate forest. For. Ecol. Manag. 2018, 434, 279–288. [Google Scholar] [CrossRef]
- Rocha, C.F.D.; Van Sluys, M.; Alves, M.A.S.; Bergallo, H.; Vrcibradic, D. Activity of Leaf-Litter Frogs: When Should Frogs Be Sampled? J. Herpetol. 2000, 34, 285. [Google Scholar] [CrossRef]
- Dundas, S.J.; Ruthrof, K.X.; Hardy, G.E.S.; Fleming, P.A. Some like it hot: Drought-induced forest die-off influences reptile assemblages. Acta Oecologica 2021, 111, 103714. [Google Scholar] [CrossRef]
- Rosa, T.F.; Camarota, F.; Zuanon, L.A.; Tito, R.; Maravalhas, J.B.; Powell, S.; Vasconcelos, H.L. The effects of high-severity fires on the arboreal ant community of a Neotropical savanna. Oecologia 2021, 196, 951–961. [Google Scholar] [CrossRef] [PubMed]
- Markle, C.E.; Wilkinson, S.L.; Waddington, J.M. Initial Effects of Wildfire on Freshwater Turtle Nesting Habitat. J. Wildl. Manag. 2020, 84, 1373–1383. [Google Scholar] [CrossRef]
- Gomes, J.O.; Maciel, A.O.; Costa, J.C.L.; Andrade, G.V. Diet Composition in Two Sympatric Amphisbaenian Species (Amphisbaena ibijara and Leposternon polystegum) from the Brazilian Cerrado. J. Herpetol. 2009, 43, 377–384. [Google Scholar] [CrossRef]
- Certini, G.; Moya, D.; Lucas-Borja, M.E.; Mastrolonardo, G. The impact of fire on soil-dwelling biota: A review. For. Ecol. Manag. 2021, 488, 118989. [Google Scholar] [CrossRef]
- Avitabile, S.C.; Nimmo, D.G.; Bennett, A.F.; Clarke, M.F. Termites Are Resistant to the Effects of Fire at Multiple Spatial Scales. PLoS ONE 2015, 10, e0140114. [Google Scholar] [CrossRef] [PubMed]
- Simioni, F.; Campos, V.A.; Dorado-Rodrigues, T.F.; Penha, J.; Strüssmann, C. Crab burrows and termite thermal chimneys as refuges for anurans in a neotropical wetland. Salamandra 2014, 50, 133–138. [Google Scholar]
- Monteiro, I.; Viana-Junior, A.B.; Solar, R.R.D.C.; Neves, F.D.S.; DeSouza, O. Disturbance-modulated symbioses in termitophily. Ecol. Evol. 2017, 7, 10829–10838. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, I.B.; Fidelis, A.; Miranda, H.S.; Ticktin, T. How do the wets burn? Fire behavior and intensity in wet grasslands in the Brazilian savanna. Braz. J. Bot. 2016, 40, 167–175. [Google Scholar] [CrossRef] [Green Version]
- Costa, L.M.; de Freitas, G.H.S.; da Silva, P.H.V.B.P.; Ribeiro, L.C.; de Vasconcelos, M.F.; Rodrigues, M. Breeding biology of the Cipo Cinclodes Cinclodes espinhacensis, a micro-endemic furnariid of the southeastern Brazilian mountains. Rev. Bras. Ornitol. 2019, 27, 63–69. [Google Scholar] [CrossRef]
- Trochet, A.; Le Chevalier, H.; Calvez, O.; Barthe, L.; Isselin-Nondedeu, F.; Picard, D.; Debelgarric, M.; Pégourié, N.; Rocher, R.; Ribéron, A. Postbreeding Movements in Marbled Newts (Caudata, Salamandridae): A Comparative Radiotracking Study in Two Habitat Types. Herpetologica 2017, 73, 1–9. [Google Scholar] [CrossRef]
- Enriquez-Urzelai, U.; Bernardo, N.; Moreno-Rueda, G.; Montori, A.; Llorente, G. Are amphibians tracking their climatic niches in response to climate warming? A test with Iberian amphibians. Clim. Chang. 2019, 154, 289–301. [Google Scholar] [CrossRef]
- Brand, M.E.; Rechkemmer, W.T.; Clark, S.A.; McCravy, K.W.; Lydeard, C.; Meiers, S.T.; Jenkins, S.E. The Influence of Fire and Other Environmental Factors on Terrestrial Gastropod Species Composition in an Oak-Hickory Woodland of West-Central Illinois. Am. Malacol. Bull. 2020, 38, 39–49. [Google Scholar] [CrossRef]
- Labonte, D.; Federle, W. Scaling and biomechanics of surface attachment in climbing animals. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20140027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sutton, G.P.; Mendoza, E.; Azizi, E.; Longo, S.J.; Olberding, J.P.; Ilton, M.; Patek, S.N. Why do Large Animals Never Actuate Their Jumps with Latch-Mediated Springs? Because They can Jump Higher without Them. Integr. Comp. Biol. 2019, 59, 1609–1618. [Google Scholar] [CrossRef] [Green Version]
- Brandt, E.E.; Sasiharan, Y.; Elias, D.O.; Mhatre, N. Jump takeoff in a small jumping spider. J. Comp. Physiol. A 2021, 207, 153–164. [Google Scholar] [CrossRef]
- Sarremejane, R.; Mykrä, H.; Bonada, N.; Aroviita, J.; Muotka, T. Habitat connectivity and dispersal ability drive the assembly mechanisms of macroinvertebrate communities in river networks. Freshw. Biol. 2017, 62, 1073–1082. [Google Scholar] [CrossRef]
- Mardiastuti, A. Response and impact of fire on bird community in the tropical rainforest: A review. IOP Conf. Ser. Earth Environ. Sci. 2020, 504, 012001. [Google Scholar] [CrossRef]
- Hirt, M.R.; Jetz, W.; Rall, B.C.; Brose, U. A general scaling law reveals why the largest animals are not the fastest. Nat. Ecol. Evol. 2017, 1, 1116–1122. [Google Scholar] [CrossRef]
- Weyand, P.G.; Davis, J.A. Running performance has a structural basis. J. Exp. Biol. 2005, 208, 2625–2631. [Google Scholar] [CrossRef] [Green Version]
- Ruf, T.; Valencak, T.; Tataruch, F.; Arnold, W. Running Speed in Mammals Increases with Muscle n-6 Polyunsaturated Fatty Acid Content. PLoS ONE 2006, 1, e65. [Google Scholar] [CrossRef]
- Winchell, K.M.; Maayan, I.; Fredette, J.R.; Revell, L.J. Linking locomotor performance to morphological shifts in urban lizards. Proc. R. Soc. B Boil. Sci. 2018, 285, 20180229. [Google Scholar] [CrossRef] [Green Version]
- Griffiths, A.D.; Brook, B.W. Effect of fire on small mammals: A systematic review. Int. J. Wildland Fire 2014, 23, 1034–1043. [Google Scholar] [CrossRef] [Green Version]
- Silveira, L.; Henrique, F.; Rodrigues, G.; Jácomo, A.T.d.A.; Filho, J.A.F.D. Impact of wildfires on the megafauna of Emas National Park, central Brazil. Oryx 1999, 33, 108–114. [Google Scholar] [CrossRef] [Green Version]
- Mahony, M.; Gould, J.; Beranek, C.T.; Callen, A.; Clulow, J.; Clulow, S.; Klop-Toker, K.; Mahony, S.; Wallace, S.; Seeto, R.; et al. A trait-based analysis for predicting impact of wildfires on frogs. Aust. Zool. 2022, 42, 326–351. [Google Scholar] [CrossRef]
- Jofré, L.E.; Curth, M.D.T.; Farji-Brener, A.G. Unexpected costs of extended phenotypes: Nest features determine the effect of fires on leaf cutter ant’s demography. Proc. R. Soc. B Boil. Sci. 2022, 289, 20212333. [Google Scholar] [CrossRef] [PubMed]
- Moore, P.A.; Smolarz, A.G.; Markle, C.E.; Waddington, J.M. Hydrological and thermal properties of moss and lichen species on rock barrens: Implications for turtle nesting habitat. Ecohydrology 2018, 12, e2057. [Google Scholar] [CrossRef]
- Collias, N. Engineering aspects of nest building by birds. Endeavour 1986, 10, 9–16. [Google Scholar] [CrossRef]
- Costa, L.M.; de Freitas, G.H.S.; Rodrigues, M. Architecture, composition and placement of nests of the Cipo Canastero Asthenes luizae (Aves: Furnariidae), a bird endemic to Brazilian mountaintops. J. Nat. Hist. 2019, 53, 391–412. [Google Scholar] [CrossRef]
- Alambiaga, I.; Álvarez, E.; Diez-Méndez, D.; Verdejo, J.; Barba, E. “The tale of the three little tits”: Different nest building solutions under the same environmental pressures. Avian Biol. Res. 2020, 13, 49–56. [Google Scholar] [CrossRef]
- Rowley, I. The use of mud in nest-building—A review of the incidence and taxonomic importance. Ostrich 1969, 40, 139–148. [Google Scholar] [CrossRef]
- Abensperg-Traun, M.; Milewski, A.V. Abundance and diversity of termites (Isoptera) in imburnt versus burnt vegetation at the Barrens in Mediterranean Western Australia. Aust. J. Ecol. 1995, 20, 413–417. [Google Scholar] [CrossRef]
- Wijas, B.J.; Lim, S.; Cornwell, W.K. Continental-scale shifts in termite diversity and nesting and feeding strategies. Ecography 2021, 2022, e05902. [Google Scholar] [CrossRef]
- Smith, A.; Avitabile, S.C.; Leonard, S.W.J. Less fuel for the fire: Malleefowl (Leipoa ocellata) nesting activity affects fuel loads and fire behaviour. Wildl. Res. 2016, 43, 640. [Google Scholar] [CrossRef]
- Maisey, A.C.; Haslem, A.; Leonard, S.W.J.; Bennett, A.F. Foraging by an avian ecosystem engineer extensively modifies the litter and soil layer in forest ecosystems. Ecol. Appl. 2020, 31, e02219. [Google Scholar] [CrossRef]
- Cooper, W.E., Jr.; Vitt, L.J.; Hedges, R.; Huey, R. Locomotor impairment and defense in gravid lizards (Eumeces laticeps): Behavioral shift in activity may offset costs of reproduction in an active forager. Behav. Ecol. Sociobiol. 1990, 27, 153–157. [Google Scholar] [CrossRef]
- Smith, S.K.; Young, V.K.H. Balancing on a Limb: Effects of Gravidity on Locomotion in Arboreal, Limbed Vertebrates. Integr. Comp. Biol. 2021, 61, 573–578. [Google Scholar] [CrossRef]
- Veasey, J.S.; Houston, D.C.; Metcalfe, N.B. A hidden cost of reproduction: The trade-off between clutch size and escape take-off speed in female zebra finches. J. Anim. Ecol. 2001, 70, 20–24. [Google Scholar] [CrossRef]
- Guillemette, M.; Ouellet, J.-F. Temporary flightlessness as a potential cost of reproduction in pre-laying Common Eiders Somateria mollissima. IBIS 2005, 147, 301–306. [Google Scholar] [CrossRef]
- Fokidis, H.B.; Risch, T.S. The burden of motherhood: Gliding locomotion in mammals influences maternal reproductive investment. J. Mammal. 2008, 89, 617–625. [Google Scholar] [CrossRef]
- Noren, S.R.; Redfern, J.V.; Edwards, E.F. Pregnancy is a drag: Hydrodynamics, kinematics and performance in pre- and post-parturition bottlenose dolphins (Tursiops truncatus). J. Exp. Biol. 2011, 214, 4151–4159. [Google Scholar] [CrossRef] [Green Version]
- Miles, D.B.; Sinervo, B.; Frankino, W.A. Reproductive burden, locomotor performance, and the cost of reproduction in free ranging lizards. Evolution 2000, 54, 1386–1395. [Google Scholar] [CrossRef]
- Voigt, C.C. Reproductive energetics of the nectar-feeding bat Glossophaga soricina (Phyllostomidae). J. Comp. Physiol. B 2003, 173, 79–85. [Google Scholar] [CrossRef]
- Miller, K.E.; Bales, K.L.; Ramos, J.H.; Dietz, J.M. Energy intake, energy expenditure, and reproductive costs of female wild golden lion tamarins (Leontopithecus rosalia). Am. J. Primatol. 2006, 68, 1037–1053. [Google Scholar] [CrossRef]
- Speakman, J.R. The physiological costs of reproduction in small mammals. Philos. Trans. R. Soc. B Biol. Sci. 2007, 363, 375–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nascimento, A.R.; Malinov, I.K.; Freire, G.; Freitas, A.V.L.; Diniz, I.R. The Temporal Dynamics of Two Morpho Fabricius, 1807 Species (Lepidoptera: Nymphalidae) are Affected Differently by Fire in the Brazilian Savanna. Environ. Èntomol. 2020, 49, 1449–1454. [Google Scholar] [CrossRef]
- Blumstein, D.T.; Hayes, L.D.; Pinter-Wollman, N. Social consequences of rapid environmental change. Trends Ecol. Evol. 2022, 38, 337–345. [Google Scholar] [CrossRef]
- Daly, M.; Wilson, M. Sex, Evolution and Behavior, 2nd ed; Willard Grant Press: Boston, MA, USA, 1983. [Google Scholar]
- Grafe, T.U.; Döbler, S.; Linsenmair, K.E. Frogs flee from the sound of fire. Proc. R. Soc. B Boil. Sci. 2002, 269, 999–1003. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, H.; Bousack, H. Modelling a Historic Oil-Tank Fire Allows an Estimation of the Sensitivity of the Infrared Receptors in Pyrophilous Melanophila Beetles. PLoS ONE 2012, 7, e37627. [Google Scholar] [CrossRef] [Green Version]
- Álvarez-Ruiz, L.; Belliure, J.; Pausas, J.G. Fire-driven behavioral response to smoke in a Mediterranean lizard. Behav. Ecol. 2021, 32, 662–667. [Google Scholar] [CrossRef]
- Álvarez-Ruiz, L.; Pausas, J.G.; Blumstein, D.T.; Putman, B.J. Lizards’ response to the sound of fire is modified by fire history. Anim. Behav. 2023, 196, 91–102. [Google Scholar] [CrossRef]
- Prevedello, J.A.; Forero-Medina, G.; Vieira, M.V. Does land use affect perceptual range? Evidence from two marsupials of the Atlantic Forest. J. Zool. 2011, 284, 53–59. [Google Scholar] [CrossRef]
- Prevedello, J.A.; Forero-Medina, G.; Vieira, M.V. Movement behaviour within and beyond perceptual ranges in three small mammals: Effects of matrix type and body mass. J. Anim. Ecol. 2010, 79, 1315–1323. [Google Scholar] [CrossRef]
- Kiltie, R.A. Scaling of visual acuity with body size in mammals and birds. Funct. Ecol. 2000, 14, 226–234. [Google Scholar] [CrossRef]
- Álvarez, G.; Ammagarahalli, B.; Hall, D.R.; Pajares, J.A.; Gemeno, C. Smoke, pheromone and kairomone olfactory receptor neurons in males and females of the pine sawyer Monochamus galloprovincialis (Olivier) (Coleoptera: Cerambycidae). J. Insect Physiol. 2015, 82, 46–55. [Google Scholar] [CrossRef]
- Stawski, C.; Matthews, J.K.; Körtner, G.; Geiser, F. Physiological and behavioural responses of a small heterothermic mammal to fire stimuli. Physiol. Behav. 2015, 151, 617–622. [Google Scholar] [CrossRef]
- Nowack, J.; Delesalle, M.; Stawski, C.; Geiser, F. Can hibernators sense and evade fires? Olfactory acuity and locomotor performance during deep torpor. Sci. Nat. 2016, 103, 73. [Google Scholar] [CrossRef] [PubMed]
- Schütz, S.; Weissbecker, B.; Hummel, H.E.; Apel, K.-H.; Schmitz, H.; Bleckmann, H. Insect antenna as a smoke detector. Nature 1999, 398, 298–299. [Google Scholar] [CrossRef]
- Schmitz, H.; Schmitz, A.; Kreiss, E.; Gebhardt, M.; Gronenberg, W. Navigation to Forest Fires by Smoke and Infrared Reception: The Specialized Sensory Systems of “Fire-Loving” Beetles. Navigation 2008, 55, 137–145. [Google Scholar] [CrossRef]
- Kay, G.M.; Driscoll, D.A.; Lindenmayer, D.B.; Pulsford, S.A.; Mortelliti, A. Pasture height and crop direction influence reptile movement in an agricultural matrix. Agric. Ecosyst. Environ. 2016, 235, 164–171. [Google Scholar] [CrossRef]
- Beauchamp, G. On how risk and group size interact to influence vigilance. Biol. Rev. 2019, 94, 1918–1934. [Google Scholar] [CrossRef]
- van der Marel, A.; López-Darias, M.; Waterman, J.M. Group-enhanced predator detection and quality of vigilance in a social ground squirrel. Anim. Behav. 2019, 151, 43–52. [Google Scholar] [CrossRef]
- Decety, J.; Norman, G.J.; Berntson, G.G.; Cacioppo, J.T. A neurobehavioral evolutionary perspective on the mechanisms underlying empathy. Prog. Neurobiol. 2012, 98, 38–48. [Google Scholar] [CrossRef]
- Morelli, F.; Benedetti, Y.; Díaz, M.; Grim, T.; Ibáñez-Álamo, J.D.; Jokimäki, J.; Kaisanlahti-Jokimäki, M.; Tätte, K.; Markó, G.; Jiang, Y.; et al. Contagious fear: Escape behavior increases with flock size in European gregarious birds. Ecol. Evol. 2019, 9, 6096–6104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Socias-Martínez, L.; Kappeler, P.M. Catalyzing Transitions to Sociality: Ecology Builds on Parental Care. Front. Ecol. Evol. 2019, 7, 160. [Google Scholar] [CrossRef] [Green Version]
- Beauchamp, G.; Li, Z.; Yu, C.; A Bednekoff, P.; Blumstein, D.T. A meta-analysis of the group-size effect on vigilance in mammals. Behav. Ecol. 2021, 32, 919–925. [Google Scholar] [CrossRef]
- Pérez-Manrique, A.; Gomila, A. Emotional contagion in nonhuman animals: A review. WIREs Cogn. Sci. 2021, 13, e1560. [Google Scholar] [CrossRef]
- Clutton-Brock, T.H.; Gaynor, D.; McIlrath, G.M.; Maccoll, A.D.C.; Kansky, R.; Chadwick, P.; Manser, M.; Skinner, J.D.; Brotherton, P.N.M. Predation, group size and mortality in a cooperative mongoose, Suricata suricatta. J. Anim. Ecol. 1999, 68, 672–683. [Google Scholar] [CrossRef] [Green Version]
- Komarek, E.V. Fire and animal behavior. In Proceedings of the Tall Timbers Fire Ecology Conference, Tallahassee, FL, USA, 10–11 April 1969; pp. 160–207. [Google Scholar]
- Pruetz, J.D.; LaDuke, T.C. Brief communication: Reaction to fire by savanna chimpanzees (Pan troglodytes verus) at Fongoli, Senegal: Conceptualization of “fire behavior” and the case for a chimpanzee model. Am. J. Phys. Anthr. 2009, 141, 646–650. [Google Scholar] [CrossRef]
- Arganda, S.; Pérez-Escudero, A.; de Polavieja, G.G. A common rule for decision making in animal collectives across species. Proc. Natl. Acad. Sci. USA 2012, 109, 20508–20513. [Google Scholar] [CrossRef] [Green Version]
- Ellis, S.; Snyder-Mackler, N.; Ruiz-Lambides, A.; Platt, M.L.; Brent, L.J.N. Deconstructing sociality: The types of social connections that predict longevity in a group-living primate. Proc. R. Soc. B Boil. Sci. 2019, 286, 20191991. [Google Scholar] [CrossRef] [Green Version]
- Montero, A.P.; Williams, D.M.; Martin, J.G.; Blumstein, D.T. More social female yellow-bellied marmots, Marmota flaviventer, have enhanced summer survival. Anim. Behav. 2020, 160, 113–119. [Google Scholar] [CrossRef]
- Philson, C.S.; Blumstein, D.T. Emergent social structure is typically not associated with survival in a facultatively social mammal. Biol. Lett. 2023, 19, 20220511. [Google Scholar] [CrossRef]
- Pilon, N.A.L.; Cava, M.G.B.; Hoffmann, W.A.; Abreu, R.C.R.; Fidelis, A.; Durigan, G. The diversity of post-fire regeneration strategies in the cerrado ground layer. J. Ecol. 2020, 109, 154–166. [Google Scholar] [CrossRef]
- van Mantgem, E.F.; Keeley, J.E.; Witter, M. Faunal Responses to Fire in Chaparral and Sage Scrub in California, USA. Fire Ecol. 2015, 11, 128–148. [Google Scholar] [CrossRef] [Green Version]
- Gosper, C.R.; Prober, S.M.; Yates, C.J. Estimating fire interval bounds using vital attributes: Implications of uncertainty and among-population variability. Ecol. Appl. 2013, 23, 924–935. [Google Scholar] [CrossRef] [PubMed]
- Latif, Q.S.; Saab, V.A.; Dudley, J.G. Prescribed fire limits wildfire severity without altering ecological importance for birds. Fire Ecol. 2021, 17, 37. [Google Scholar] [CrossRef]
- Haslem, A.; Avitabile, S.C.; Taylor, R.S.; Kelly, L.T.; Watson, S.; Nimmo, D.G.; Kenny, S.A.; Callister, K.; Spence-Bailey, L.M.; Bennett, A.F.; et al. Time-since-fire and inter-fire interval influence hollow availability for fauna in a fire-prone system. Biol. Conserv. 2012, 152, 212–221. [Google Scholar] [CrossRef]
- Rush, S.; Klaus, N.; Keyes, T.; Petrick, J.; Cooper, R. Fire severity has mixed benefits to breeding bird species in the southern Appalachians. For. Ecol. Manag. 2012, 263, 94–100. [Google Scholar] [CrossRef]
- Chalmandrier, L.; Midgley, G.; Barnard, P.; Sirami, C. Effects of time since fire on birds in a plant diversity hotspot. Acta Oecologica 2013, 49, 99–106. [Google Scholar] [CrossRef]
- Lee, J.S.; Cornwell, W.K.; Kingsford, R.T. Rainforest bird communities threatened by extreme fire. Glob. Ecol. Conserv. 2021, 33, e01985. [Google Scholar] [CrossRef]
- Rainsford, F.W.; Kelly, L.T.; Leonard, S.W.; Bennett, A.F. Post-fire habitat relationships for birds differ among ecosystems. Biol. Conserv. 2021, 260, 109218. [Google Scholar] [CrossRef]
- Robinson, N.M.; Leonard, S.W.; Bennett, A.F.; Clarke, M.F. Refuges for birds in fire-prone landscapes: The influence of fire severity and fire history on the distribution of forest birds. For. Ecol. Manag. 2014, 318, 110–121. [Google Scholar] [CrossRef]
- Graf, M.; Lettenmaier, L.; Müller, J.; Hagge, J. Saproxylic beetles trace deadwood and differentiate between deadwood niches before their arrival on potential hosts. Insect Conserv. Divers. 2021, 15, 48–60. [Google Scholar] [CrossRef]
- Paviour-Smith, K. The Fruiting-Bodies of Macrofungi as Habitats for Beetles of the Family Ciidae (Coleoptera). Oikos 1960, 11, 43. [Google Scholar] [CrossRef]
- Hovick, T.J.; McGranahan, D.A.; Elmore, R.D.; Weir, J.R.; Fuhlendorf, S.D. Pyric-carnivory: Raptor use of prescribed fires. Ecol. Evol. 2017, 7, 9144–9150. [Google Scholar] [CrossRef] [Green Version]
- Lopes, L.E.; de Meireles, R.C.; Peixoto, H.J.C.; Teixeira, J.P.G.; Machado, T.L.d.S.S.; Lombardi, V.T. Movement ecology of the threatened Campo Miner Geositta poeciloptera and its implications for the conservation of tropical open grassland birds. Bird Conserv. Int. 2022, 33, E38. [Google Scholar] [CrossRef]
- Bagne, K.E.; Purcell, K.L. Short-term responses of birds to prescribed fire in fire-suppressed forests of California. J. Wildl. Manag. 2011, 75, 1051–1060. [Google Scholar] [CrossRef]
- Walesiak, M.; Mikusiński, G.; Borowski, Z.; Żmihorski, M. Large fire initially reduces bird diversity in Poland’s largest wetland biodiversity hotspot. Biodivers. Conserv. 2022, 31, 1037–1056. [Google Scholar] [CrossRef]
- McGee, J.M. Small Mammal Populations in an Unburned and Early Fire Successional Sagebrush Community. J. Range Manag. 1982, 35, 177. [Google Scholar] [CrossRef]
- Briani, D.C.; Palma, A.R.; Vieira, E.M.; Henriques, R.P. Post-fire succession of small mammals in the Cerrado of central Brazil. Biodivers. Conserv. 2004, 13, 1023–1037. [Google Scholar] [CrossRef]
- Holmes, A.L.; Robinson, W.D. Fire mediated patterns of population densities in mountain big sagebrush bird communities. J. Wildl. Manag. 2013, 77, 737–748. [Google Scholar] [CrossRef]
- Beal-Neves, M.; Chiarani, E.; Ferreira, P.M.A.; Fontana, C.S. The role of fire disturbance on habitat structure and bird communities in South Brazilian Highland Grasslands. Sci. Rep. 2020, 10, 19708. [Google Scholar] [CrossRef]
- Saab, V.A.; Latif, Q.R.; Block, W.M.; Dudley, J.G. Short-term benefits of prescribed fire to bird communities of dry forests. Fire Ecol. 2022, 18, 4. [Google Scholar] [CrossRef]
- Stawski, C.; Hume, T.; Körtner, G.; Currie, S.E.; Nowack, J.; Geiser, F. Post-fire recovery of torpor and activity patterns of a small mammal. Biol. Lett. 2017, 13, 20170036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matthews, J.K.; Stawski, C.; Körtner, G.; Parker, C.A.; Geiser, F. Torpor and basking after a severe wildfire: Mammalian survival strategies in a scorched landscape. J. Comp. Physiol. B 2016, 187, 385–393. [Google Scholar] [CrossRef] [PubMed]
- Stawski, C.; Körtner, G.; Nowack, J.; Geiser, F. The importance of mammalian torpor for survival in a post-fire landscape. Biol. Lett. 2015, 11, 20150134. [Google Scholar] [CrossRef]
- Geiser, F.; Drury, R.L.; Körtner, G.; Turbill, C.; Pavey, C.R.; Brigham, R.M. Passive rewarming from torpor in mammals and birds: Energetic, ecological and evolutionary implications. In Life in the Cold: Evolution, Mechanisms, Adaptation, and Application; Barnes, B.M., Carey, H.V., Eds.; Biological Papers of the University of Alaska: Fairbanks, AK, USA, 2004; pp. 51–62. [Google Scholar]
- Turbill, C.; Stojanovski, L. Torpor reduces predation risk by compensating for the energetic cost of antipredator foraging behaviours. Proc. R. Soc. B Boil. Sci. 2018, 285, 20182370. [Google Scholar] [CrossRef] [Green Version]
- Thorat, L.; Nath, B.B. Insects with Survival Kits for Desiccation Tolerance under Extreme Water Deficits. Front. Physiol. 2018, 9, 1843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Legge, S.; Murphy, S.; Heathcote, J.; Flaxman, E.; Augusteyn, J.; Crossman, M. The short-term effects of an extensive and high-intensity fire on vertebrates in the tropical savannas of the central Kimberley, northern Australia. Wildl. Res. 2008, 35, 33–43. [Google Scholar] [CrossRef]
- Lindsay, M.N.; Lewis, D.B.; Halstead, N.; Gainsbury, A.M. Fire severity effects on the herpetofaunal diversity of the Florida scrub, a biodiversity hotspot. Biodivers. Conserv. 2023, 32, 1857–1878. [Google Scholar] [CrossRef]
- Leahy, L.; Legge, S.; Tuft, K.; McGregor, H.; Barmuta, L.; Jones, M.; Johnson, C.N. Amplified predation after fire suppresses rodent populations in Australia’s tropical savannas. Wildl. Res. 2015, 42, 705–716. [Google Scholar] [CrossRef]
- Shaw, R.E.; James, A.I.; Tuft, K.; Legge, S.; Cary, G.J.; Peakall, R.; Banks, S.C. Unburnt habitat patches are critical for survival and in situ population recovery in a small mammal after fire. J. Appl. Ecol. 2021, 58, 1325–1335. [Google Scholar] [CrossRef]
- Doherty, T.S.; Geary, W.L.; Jolly, C.J.; Macdonald, K.J.; Miritis, V.; Watchorn, D.J.; Cherry, M.J.; Conner, L.M.; González, T.M.; Legge, S.M.; et al. Fire as a driver and mediator of predator–prey interactions. Biol. Rev. 2022, 97, 1539–1558. [Google Scholar] [CrossRef] [PubMed]
- Nordberg, E.J.; Schwarzkopf, L. Reduced competition may allow generalist species to benefit from habitat homogenization. J. Appl. Ecol. 2018, 56, 305–318. [Google Scholar] [CrossRef]
- Switzer, P.V. Site fidelity in predictable and unpredictable habitats. Evol. Ecol. 1993, 7, 533–555. [Google Scholar] [CrossRef]
- Kreling, S.E.; Gaynor, K.M.; McInturff, A.; Calhoun, K.L.; Brashares, J.S. Site fidelity and behavioral plasticity regulate an ungulate’s response to extreme disturbance. Ecol. Evol. 2021, 11, 15683–15694. [Google Scholar] [CrossRef]
- Klaassen, M.; Bauer, S.; Madsen, J.; Tombre, I. Modelling behavioural and fitness consequences of disturbance for geese along their spring flyway. J. Appl. Ecol. 2005, 43, 92–100. [Google Scholar] [CrossRef] [Green Version]
- Sagonas, K.; Meiri, S.; Valakos, E.D.; Pafilis, P. The effect of body size on the thermoregulation of lizards on hot, dry Mediterranean islands. J. Therm. Biol. 2013, 38, 92–97. [Google Scholar] [CrossRef]
- Zamora-Camacho, F.J.; Reguera, S.; Moreno-Rueda, G. Bergmann’s Rule rules body size in an ectotherm: Heat conservation in a lizard along a 2200-metre elevational gradient. J. Evol. Biol. 2014, 27, 2820–2828. [Google Scholar] [CrossRef]
- Prada, M.; Marinho-Filho, J. Effects of Fire on the Abundance of Xenarthrans in Mato Grosso, Brazil. Austral Ecol. 2004, 29, 568–573. [Google Scholar] [CrossRef]
- Turschak, G.; Rochester, C.; Hathaway, S.; Stokes, D.; Haas, C.; Fisher, R. Effects of Large-Scale Wildfire on Carnivores in San Diego County, California. San Diego, CA. 2010. Available online: https://sdmmp.com/view_article.php?cid=CiteID_1603251358357600 (accessed on 15 March 2021).
- Miranda, R.B.; Klaczko, J.; Tonini, J.F.; Brandão, R.A. Escaping from predators: A review of Neotropical lizards defence traits. Ethol. Ecol. Evol. 2022, 34, 1–31. [Google Scholar] [CrossRef]
- Céréghino, R.; Françoise, L.; Bonhomme, C.; Carrias, J.-F.; Compin, A.; Corbara, B.; Jassey, V.; Leflaive, J.; Rota, T.; Farjalla, V.; et al. Desiccation resistance traits predict freshwater invertebrate survival and community response to drought scenarios in a Neotropical ecosystem. Ecol. Indic. 2020, 119, 106839. [Google Scholar] [CrossRef]
- Santos, F.L.; Nogueira, J.; de Souza, R.A.F.; Falleiro, R.M.; Schmidt, I.B.; Libonati, R. Prescribed Burning Reduces Large, High-Intensity Wildfires and Emissions in the Brazilian Savanna. Fire 2021, 4, 56. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Fire Regime Parameter | Effect on Vulnerability/Score | ||||
|---|---|---|---|---|---|
| Negligible | Slight | Moderate | Important | Extreme | |
| 0.5 | 1.0 | 2.0 | 4.0 | 8.0 | |
| Fire return interval 1 | <10% | 10–30% | 31–50% | 51–70% | >70% |
| Fire extent 2 | 0 | 1 | 2 | 3 | ≥4 |
| Fire seasonality 3 | <10% | 10–30% | 31–50% | 51–70% | >70% |
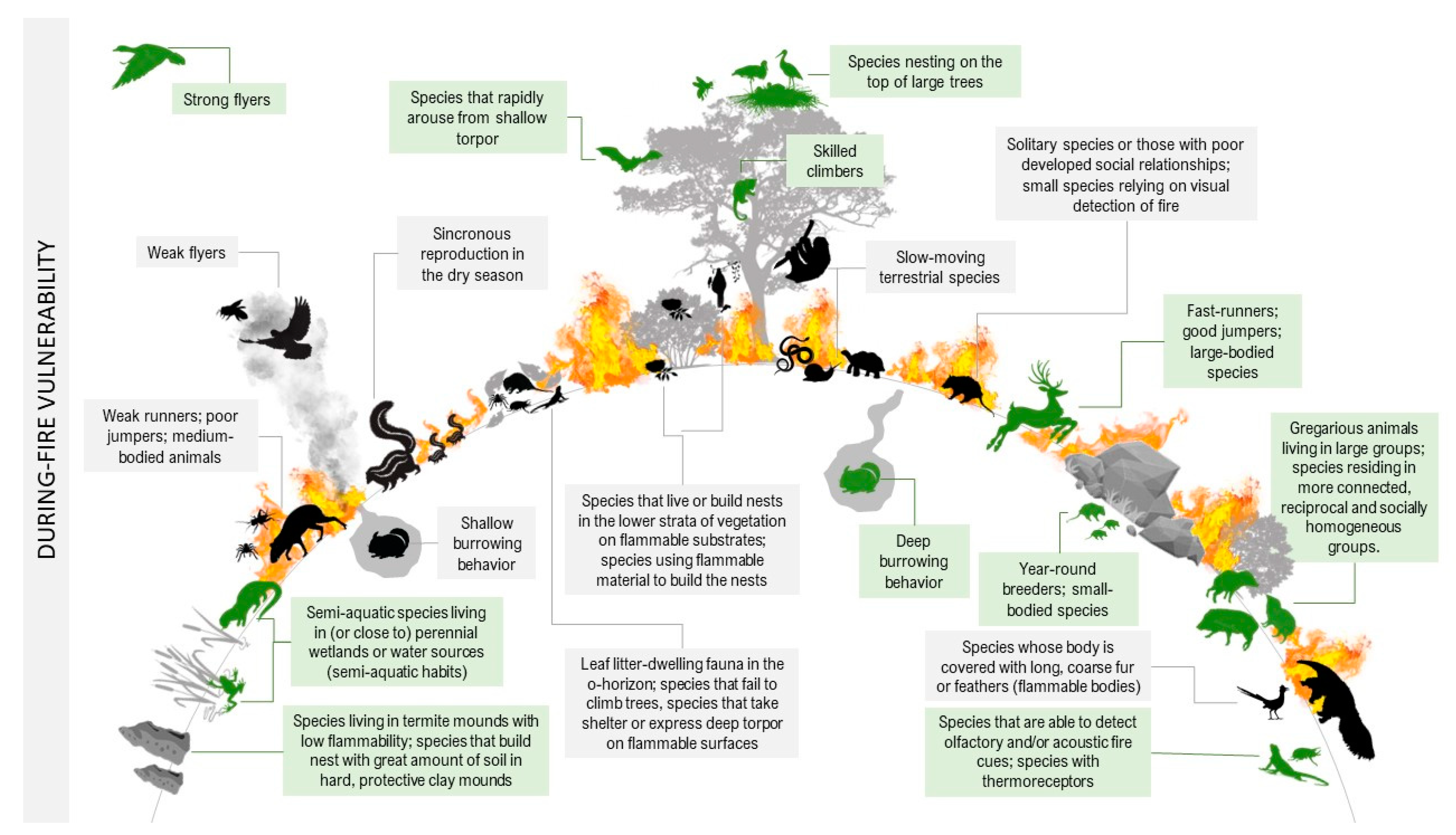
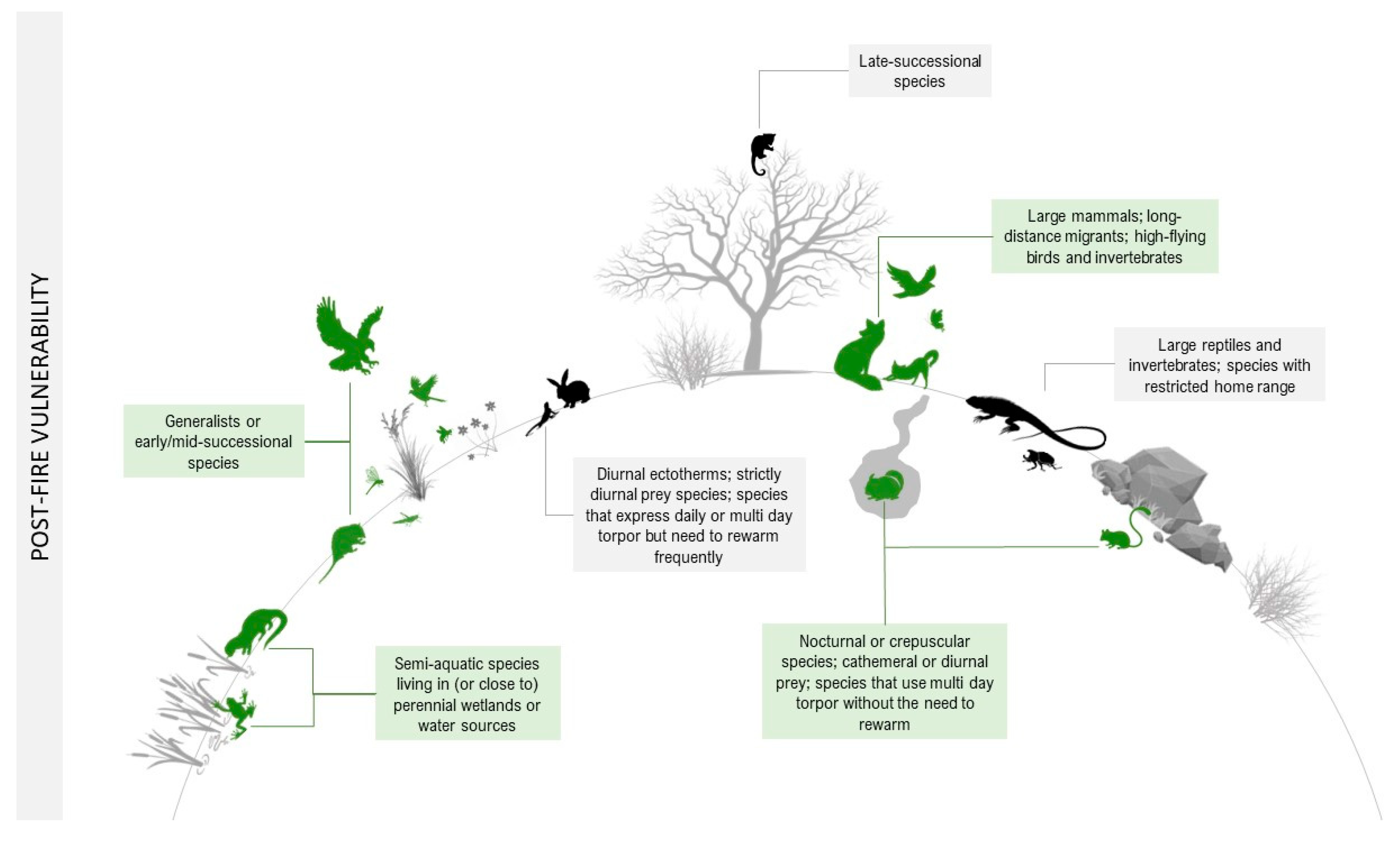
| Trait Group | Increase Vulnerability | Decrease Vulnerability | |
|---|---|---|---|
| DURING FIRE | Dormancy | Species that often express deep torpor on flammable surfaces in the flame zone. | Non-hibernators; species that rapidly arouse from shallow torpor when exposed to smoke or flame noises; species that remain in torpor in places protected from fire, such as in deeper soil layers |
| Escape decision | Animals that run away randomly when frightened; fossorial species with shallow burrowing behavior; species that take shelter in flammable or suffocating places, such as plants in the lower layers, litter, or cavities in small trees. | Animals that run toward nearby refuges when frightened; fossorial species with deep burrowing behavior; scansorial animals that seek refuge on top of tall trees during surface fires, in water, in termite mounds, or on rocky surfaces with little flammable material. | |
| Habitat use | Leaf litter-dwelling fauna in the o-horizon and other species that live or build nests in the lower strata of vegetation on flammable substrates, such as shrubs, grasses, dry and/or fallen trunks and branches, and small trees. | Soil-dwelling species that can burrow deeper into the ground; species that live or build nests close to perennial wetlands or water sources (semi-aquatic habits), below-ground, on rocky substrates, termite mounds with low flammability, and deep cavities inside massive tree trunks or in the upper strata of vegetation (on the top of tall trees). | |
| Mobility | Limited movement capability: slow-moving animals, weak flyers, ground-dwelling species that fail to climb trees, smaller jumpers with reduced effective jump height. | Good or excellent movement capability: fast runners, strong flyers, skilled climbers, larger jumpers with great effective jump height, and other jumping specialists that use catapult mechanisms. | |
| Morphology | Medium-bodied animals that may have difficulty fleeing or finding refuge; species whose bodies are covered with long, coarse fur or feathers. | Small-bodied animals that can find refuge more easily during a fire, while larger ones can flee or move away from affected areas; species with short fur, smooth skin, or covered with scales. | |
| Nest substrate | Species using flammable materials to build nests: thatched mounds, moss and lichen, fine grass or mammalian hair, and plant material such as bark, fiber, leaves, twigs, grasses, tussocks, and branches. | Species that use thermally insulating building materials: great amounts of soil in hard, protective clay mounds; species with deliberate behavior for modifying their surrounding environment causally reducing flammability; species that build subterranean nests without thatched mounds. | |
| Reproductive cycles | Synchronous reproduction, usually at the end of the dry season, exposing fragile life stages, pregnant, lactating, nesting, and brooding females to high-intensity fire. | Year-round breeders or species that reproduce during the wet season but decrease reproduction during the dry season. | |
| Sensory detection of fire cues | Species that spend most of their time in complex vegetation and rely primarily on the visual detection of fire (small-bodied animals could be even more vulnerable, as they usually have lower visual acuity). | Species that are able to detect olfactory and/or acoustic fire cues; species that can detect fire cues at lower thresholds; species that have thermoreceptors that can detect infrared radiation from fires; species relying primarily on the visual detection of fire, but that spend most of their time in the top of tall trees or open, low-stature vegetation and topographically simple landscapes. | |
| Social organization | Solitary animals or those that live in small family groups (parents and young); species with poorly developed social relationships (e.g., groups with weak connections) and whose individuals or groups lack effective communication skills. | Gregarious animals living in large groups; social species or those residing in more connected, reciprocal, and socially homogeneous groups. | |
| Behavioral plasticity | Late-successional species that require more structured habitats for nest sites and foraging, which take several years to recover. | Generalists that can temporarily adapt their diet and/or habitat preferences to the conditions and food resources available across the post-fire landscape; species that may benefit from fire-induced changes include early or mid-successional species. | |
| POST-FIRE | Dormancy | Species that express multi-day torpor but need to rewarm frequently; species that use daily torpor, which is not as deep as hibernation, lasts only some hours rather than days or weeks, and is usually—but not always—interrupted by daily foraging and feeding. | Invertebrates that remain inactive after a fire, allowing their tissues to become desiccated (anhydrobiosis); invertebrates that express aestivation and remain in an inactive stage remarkably resistant to water loss; species that use multi-day torpor for weeks or even months after a fire or during fire season without the need to rewarm. |
| Endogenous circadian rhythms | Diurnal ectotherms that depend on thermoregulation opportunities afforded by habitat structure; strictly diurnal prey species. | Nocturnal or crepuscular species; cathemeral or diurnal prey that can adjust their daily activity patterns. | |
| Mobility | Species with restricted home range; species with high site fidelity or territorial species; Migratory species (highly mobile), but with strong site fidelity. | Highly mobile species that travel long distances or show metapopulation dynamics; species with low site fidelity or non-territorial species. | |
| Morphology | Large ectotherms; invertebrates with thinner cuticles. | Large mammals; species capable of camouflaging in the scorched substrate; invertebrates with higher cuticle thickness. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Batista, E.K.L.; Figueira, J.E.C.; Solar, R.R.C.; de Azevedo, C.S.; Beirão, M.V.; Berlinck, C.N.; Brandão, R.A.; de Castro, F.S.; Costa, H.C.; Costa, L.M.; et al. In Case of Fire, Escape or Die: A Trait-Based Approach for Identifying Animal Species Threatened by Fire. Fire 2023, 6, 242. https://doi.org/10.3390/fire6060242
Batista EKL, Figueira JEC, Solar RRC, de Azevedo CS, Beirão MV, Berlinck CN, Brandão RA, de Castro FS, Costa HC, Costa LM, et al. In Case of Fire, Escape or Die: A Trait-Based Approach for Identifying Animal Species Threatened by Fire. Fire. 2023; 6(6):242. https://doi.org/10.3390/fire6060242
Chicago/Turabian StyleBatista, Eugênia K. L., José E. C. Figueira, Ricardo R. C. Solar, Cristiano S. de Azevedo, Marina V. Beirão, Christian N. Berlinck, Reuber A. Brandão, Flávio S. de Castro, Henrique C. Costa, Lílian M. Costa, and et al. 2023. "In Case of Fire, Escape or Die: A Trait-Based Approach for Identifying Animal Species Threatened by Fire" Fire 6, no. 6: 242. https://doi.org/10.3390/fire6060242