
Short-Term Response of Soil Bacterial Communities after Prescribed Fires in Semi-Arid Mediterranean Forests

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
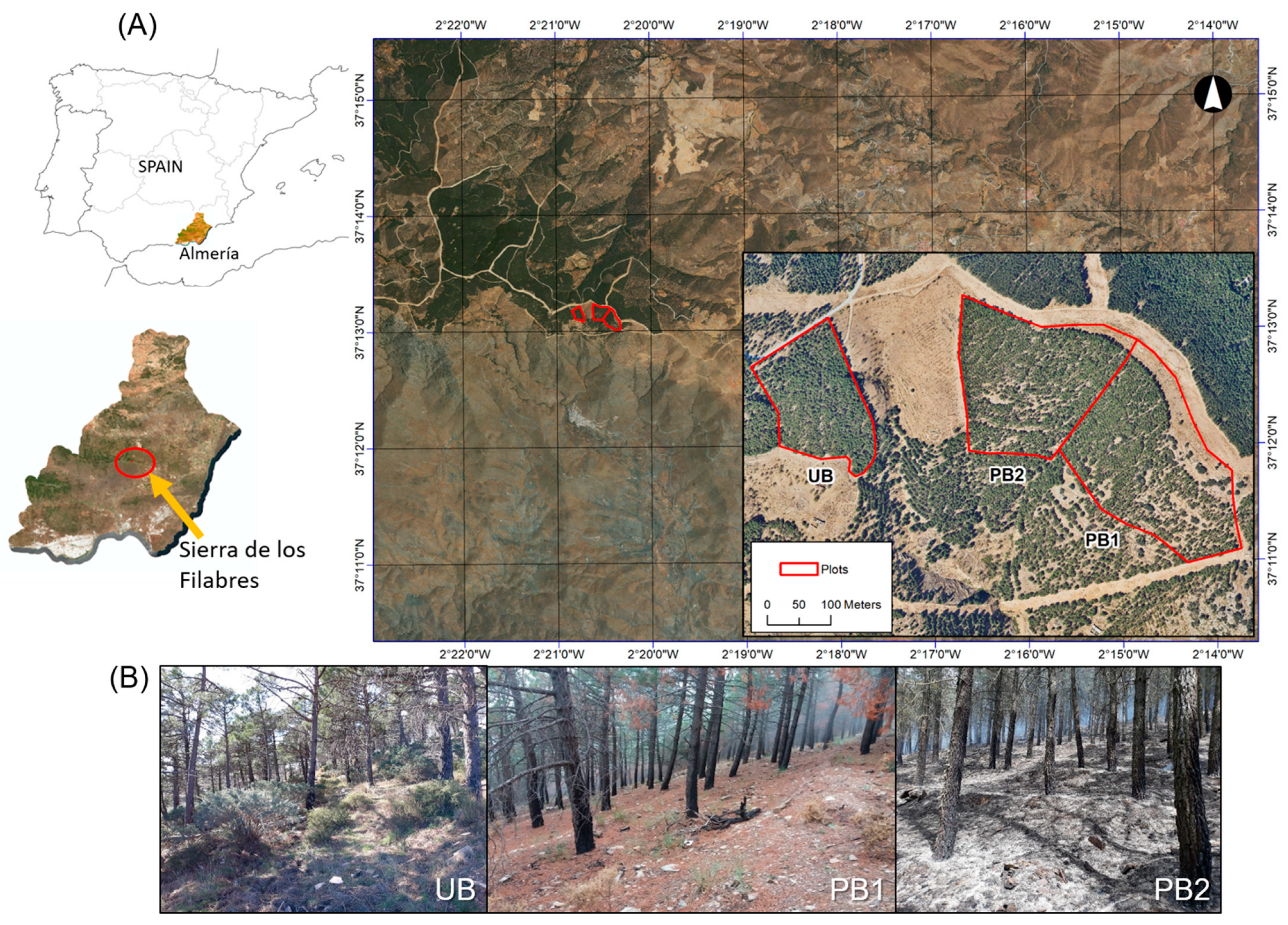
2.1. Study Site
2.2. Prescribed Burning Operations and Experimental Design
2.3. Chemical Soil Properties
2.4. Composition and Diversity of Bacterial Communities in Soils Treated and Untreated with Prescribed Fire
2.5. Statistical Analyses
3. Results
3.1. Prescribed Burning Effects on Soil Properties
3.2. Bacterial Community Richness and Diversity
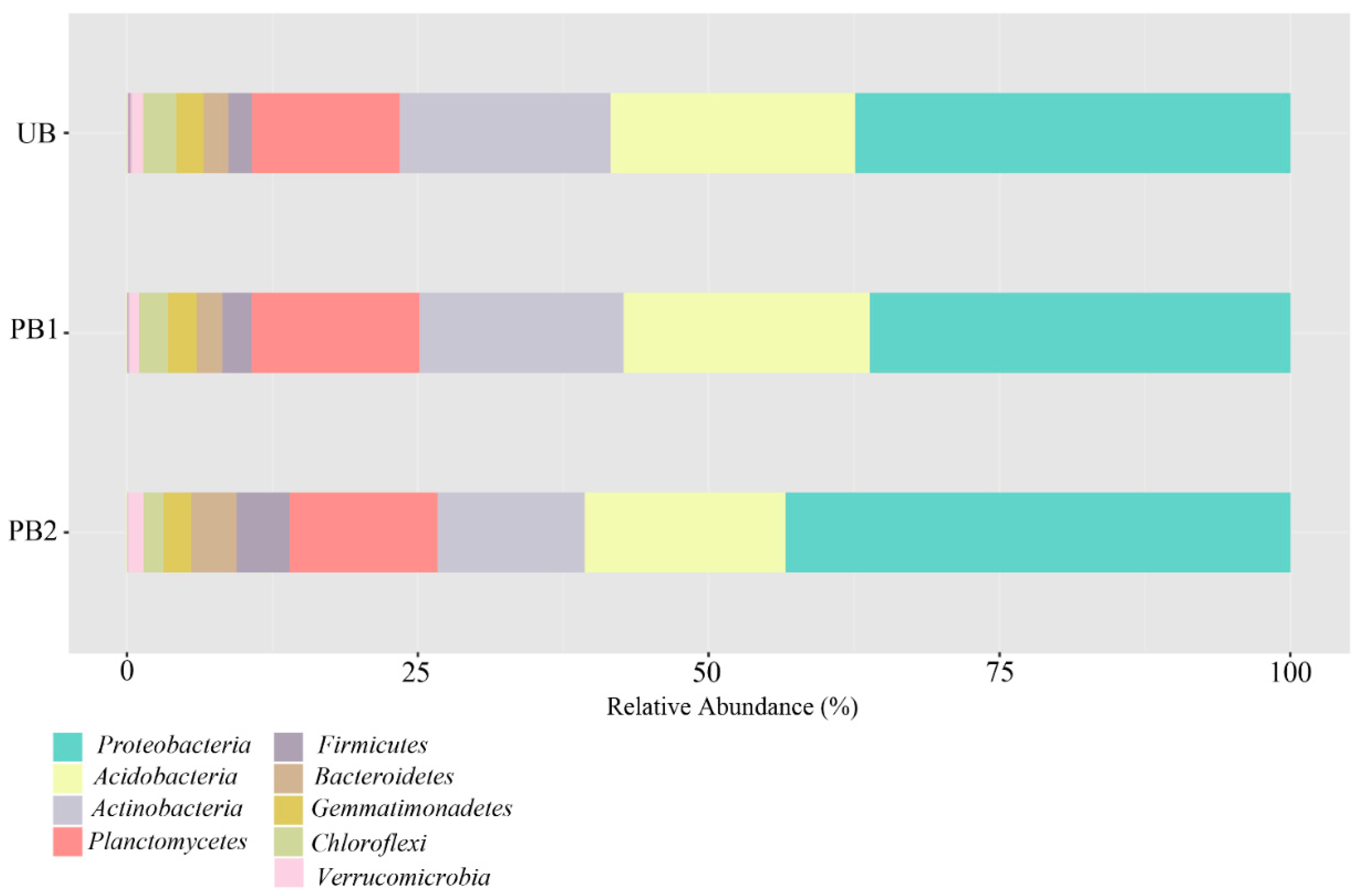
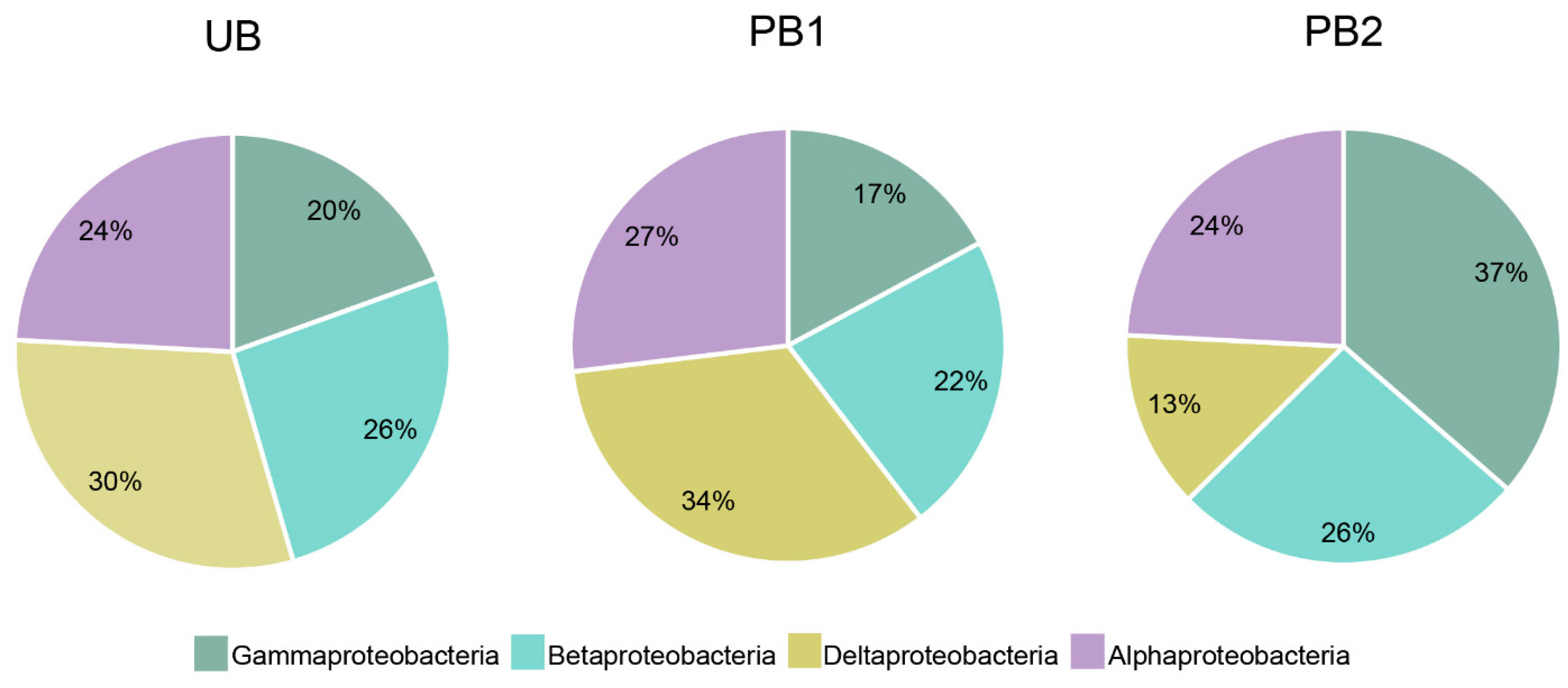
3.3. Analysis of Bacterial Communities after Prescribed Burning Treatment
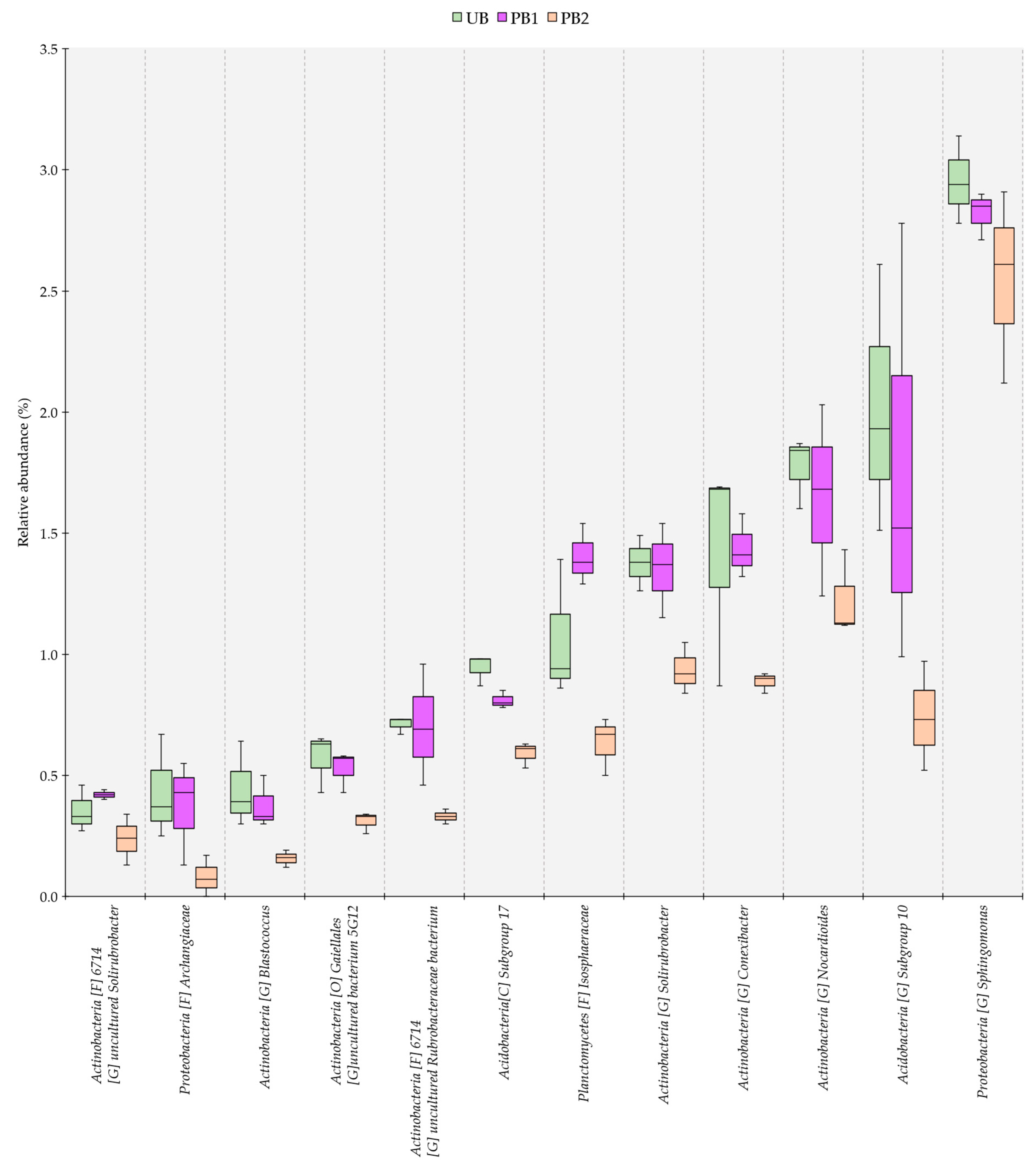
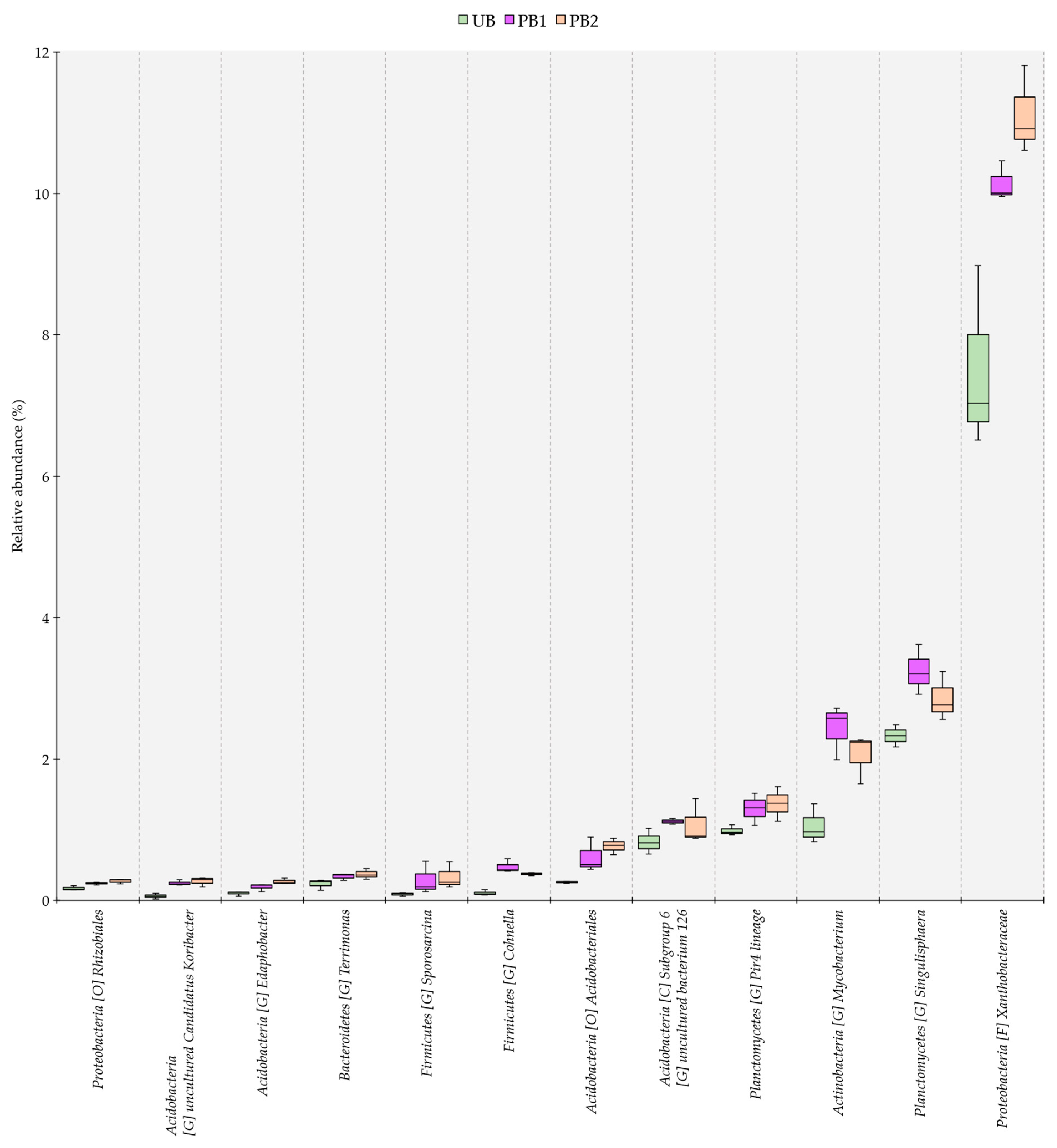
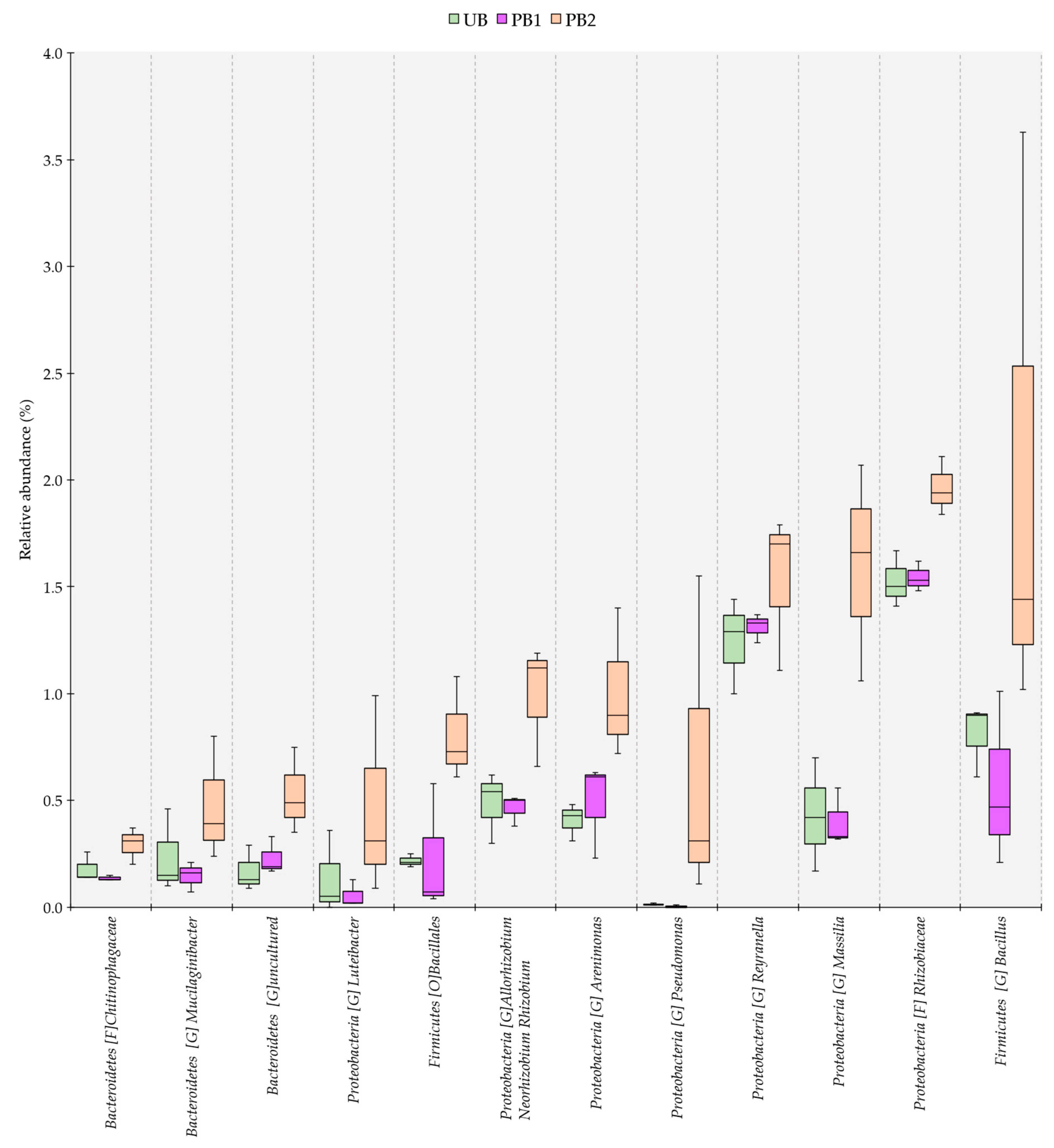
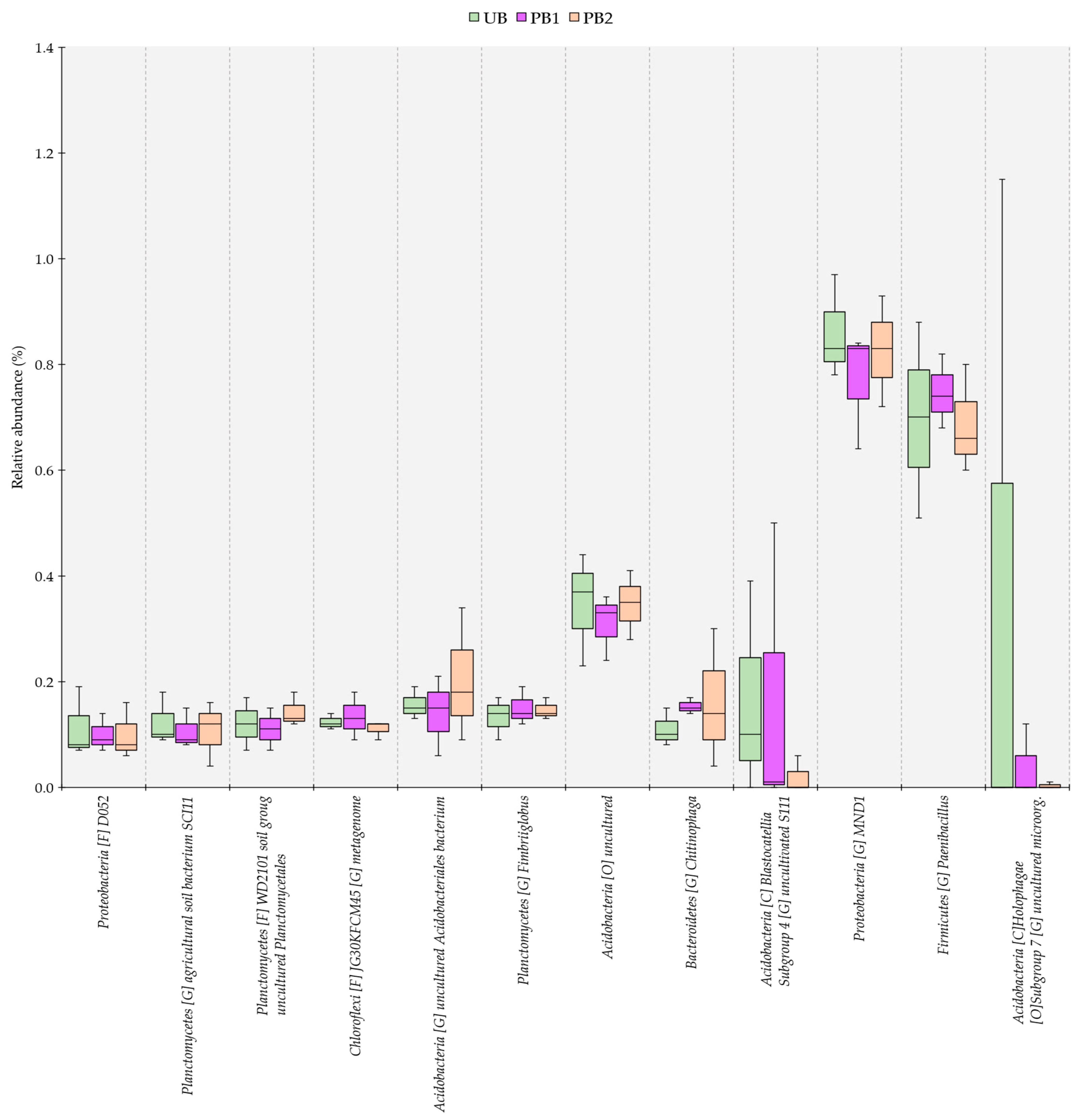
3.4. Genus-Level Dissimilarity Analysis of Bacterial Communities in Soils Treated with Prescribed Burning and Unburned Soils
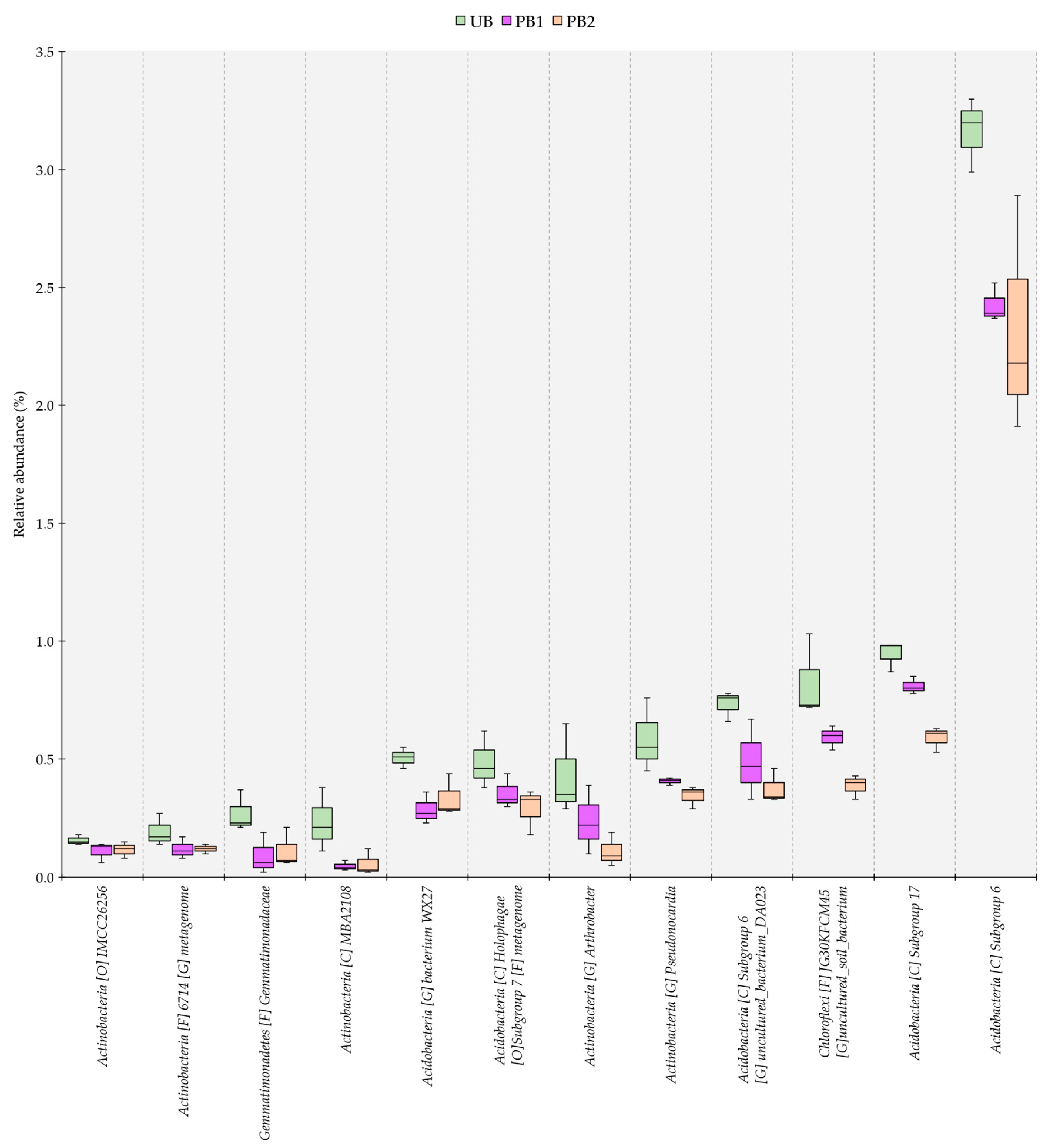
3.5. Relative Abundance of Bacterial Communities as a Function of Time since Prescribed Burning Treatment
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- San-Miguel-Ayanz, J.; Moreno, J.M.; Camia, A. Analysis of Large Fires in European Mediterranean Landscapes: Lessons Learned and Perspectives. For. Ecol. Manag. 2013, 294, 11–22. [Google Scholar] [CrossRef]
- Rodríguez y Silva, F.R.; González-Cabán, A. ‘SINAMI’: A Tool for the Economic Evaluation of Forest Fire Management Programs in Mediterranean Ecosystems. Int. J. Wildl. Fire 2010, 19, 927–936. [Google Scholar] [CrossRef]
- Pausas, J.G.; Keeley, J.E. A Burning Story: The Role of Fire in the History of Life. Bioscience 2009, 59, 593–601. [Google Scholar] [CrossRef] [Green Version]
- Turco, M.; Bedia, J.; Di Liberto, F.; Fiorucci, P.; Von Hardenberg, J.; Koutsias, N.; Llasat, M.C.; Xystrakis, F.; Provenzale, A. Decreasing Fires in Mediterranean Europe. PLoS ONE 2016, 11, e0150663. [Google Scholar] [CrossRef] [Green Version]
- Moreno Rodríguez, J.M.; Urbieta, I.R.; Bedia, J.; Gutiérrez, J.M.; Vallejo, V.R. Los Incendios Forestales En España Ante Al Cambio Climático.Los bosques y la Biodiversidad frente al Cambio Climático, Vulnerabilidad, y Adpaptación en España. Ministerio de Agricultura, Alimentación y Medio Ambiente, 2015. pp. 584–588. Available online: http://hdl.handle.net/10578/8179 (accessed on 28 February 2023).
- Neary, D.G.; Klopatek, C.C.; DeBano, L.F.; Ffolliott, P.F. Fire Effects on Belowground Sustainability: A Review and Synthesis. For. Ecol. Manag. 1999, 122, 51–71. [Google Scholar] [CrossRef]
- Certini, G. Effects of Fire on Properties of Forest Soils: A Review. Oecologia 2005, 143, 1–10. [Google Scholar] [CrossRef]
- Hinojosa, M.B.; Parra, A.; Laudicina, V.A.; Moreno, J.M. Post-Fire Soil Functionality and Microbial Community Structure in a Mediterranean Shrubland Subjected to Experimental Drought. Sci. Total Environ. 2016, 573, 1178–1189. [Google Scholar] [CrossRef]
- Certini, G. Fire as a Soil-Forming Factor. Ambio 2014, 43, 191–195. [Google Scholar] [CrossRef] [Green Version]
- Neary, D.G.; Ryan, K.C.; DeBano, L.F. Wildland Fire in Ecosystems: Effects of Fire on Soils and Water; General Technical Report RMRS-GTR-42-vol.4; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Ogden, UT, USA, 2005; 250p. [CrossRef] [Green Version]
- Andreu, V.; Imeson, A.C.; Rubio, J.L. Temporal Changes in Soil Aggregates and Water Erosion after a Wildfire in a Mediterranean Pine Forest. Catena 2001, 44, 69–84. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Berbel, N.; Ortega, R.; Lucas-Borja, M.E.; Solé-Benet, A.; Miralles, I. Long-Term Effects of Two Organic Amendments on Bacterial Communities of Calcareous Mediterranean Soils Degraded by Mining. J. Environ. Manag. 2020, 271, 110920. [Google Scholar] [CrossRef]
- Fontúrbel, M.T.; Barreiro, A.; Vega, J.A.; Martín, A.; Jiménez, E.; Carballas, T.; Fernández, C.; Díaz-Raviña, M. Effects of an Experimental Fire and Post-Fire Stabilization Treatments on Soil Microbial Communities. Geoderma 2012, 191, 51–60. [Google Scholar] [CrossRef]
- Lucas-Borja, M.E.; Miralles, I.; Ortega, R.; Plaza-Álvarez, P.A.; Gonzalez-Romero, J.; Sagra, J.; Soriano-Rodríguez, M.; Certini, G.; Moya, D.; Heras, J. Immediate Fire-Induced Changes in Soil Microbial Community Composition in an Outdoor Experimental Controlled System. Sci. Total Environ. 2019, 696, 134033. [Google Scholar] [CrossRef] [PubMed]
- Mataix-Solera, J.; Cerdà, A. Efectos de los Incendios Forestales Sobre los Suelos en España. El Estado de la Cuestión Visto por los Científicos Españoles; Cátedra de Divulgació de La Ciencia Universitat de Valencia: Valencia, Spain, 2009; pp. 491–532. [Google Scholar]
- Sáenz de Miera, L.E.; Pinto, R.; Gutierrez-Gonzalez, J.J.; Calvo, L.; Ansola, G. Wildfire Effects on Diversity and Composition in Soil Bacterial Communities. Sci. Total Environ. 2020, 726, 138636. [Google Scholar] [CrossRef] [PubMed]
- Molina, J.R.; García, J.P.; Fernández, J.J.; Rodríguez y Silva, F. Prescribed Fire Experiences on Crop Residue Removal for Biomass Exploitations. Application to the Maritime Pine Forests in the Mediterranean Basin. Sci. Total Environ. 2018, 612, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Plaza-Álvarez, P.A.; Lucas-Borja, M.E.; Sagra, J.; Moya, D.; Alfaro-Sánchez, R.; González-Romero, J.; De las Heras, J. Changes in Soil Water Repellency after Prescribed Burnings in Three Different Mediterranean Forest Ecosystems. Sci. Total Environ. 2018, 644, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Santín, C.; Doerr, S.H. Fire Effects on Soils: The Human Dimension. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150171. [Google Scholar] [CrossRef] [Green Version]
- Alcañiz, M.; Outeiro, L.; Francos, M.; Úbeda, X. Effects of Prescribed Fires on Soil Properties: A Review. Sci. Total Environ. 2018, 613–614, 944–957. [Google Scholar] [CrossRef]
- Carra, B.G.; Bombino, G.; Lucas-Borja, M.E.; Muscolo, A.; Romeo, F.; Zema, D.A. Short-Term Changes in Soil Properties after Prescribed Fire and Mulching with Fern in Mediterranean Forests. J. For. Res. 2022, 33, 1271–1289. [Google Scholar] [CrossRef]
- Fernandes, P.M.; Davies, G.M.; Ascoli, D.; Fernández, C.; Moreira, F.; Rigolot, E.; Stoof, C.R.; Vega, J.A.; Molina, D. Prescribed Burning in Southern Europe: Developing Fire Management in a Dynamic Landscape. Front. Ecol. Environ. 2013, 11, e4–e14. [Google Scholar] [CrossRef] [Green Version]
- Robichaud, P.R.; Lewis, S.A.; Brown, R.E.; Bone, E.D.; Brooks, E.S. Evaluating Post-Wildfire Logging-Slash Cover Treatment to Reduce Hillslope Erosion after Salvage Logging Using Ground Measurements and Remote Sensing. Hydrol. Process. 2020, 34, 4431–4445. [Google Scholar] [CrossRef]
- Vega, J.A.; Fontúrbel, T.; Merino, A.; Fernández, C.; Ferreiro, A.; Jiménez, E. Testing the Ability of Visual Indicators of Soil Burn Severity to Reflect Changes in Soil Chemical and Microbial Properties in Pine Forests and Shrubland. Plant Soil 2013, 369, 73–91. [Google Scholar] [CrossRef]
- Zapata-Sierra, A.J.; Zapata-Castillo, L.; Manzano-Agugliaro, F. Water Resources Availability in Southern Europe at the Basin Scale in Response to Climate Change Scenarios. Environ. Sci. Eur. 2022, 34, 75. [Google Scholar] [CrossRef]
- Peinado, F.M.; Sierra, A.M. Proyecto LUCDEME. Mapa de Suelos a Escala 1:100.000 Provincia de Almería; Ministerio de Agricultura, Pesca y Alimentación: Madrid, Spain, 2004.
- Walkey, A.; Black, I.A. An Examination of Degtjareff Method for Determining Soil Organic Matter and a Proposed Modification of the Chromic Acid Titration Method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Mingorance, M.D. Guidelines for Improving Organic Carbon Recovery by the Wet Oxidation Method. Chemosphere 2007, 68, 409–413. [Google Scholar] [CrossRef]
- Walters, W.; Hyde, E.R.; Berg-Lyons, D.; Ackermann, G.; Humphrey, G.; Parada, A.; Gilbert, J.A.; Jansson, J.K.; Caporaso, J.G.; Fuhrman, J.A.; et al. Improved Bacterial 16S RRNA Gene (V4 and V4-5) and Fungal Internal Transcribed Spacer Marker Gene Primers for Microbial Community Surveys. mSystems 2016, 1, e00009-15. [Google Scholar] [CrossRef] [Green Version]
- Comeau, A.M.; Douglas, G.M.; Langille, M.G.I. Microbiome Helper: A Custom and Streamlined Workflow for Microbiome Research. mSystems 2017, 2, e00127-16. [Google Scholar] [CrossRef] [Green Version]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, Interactive, Scalable and Extensible Microbiome Data Science Using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Anderson, M.J. Permutation Tests for Univariate or Multivariate Analysis of Variance and Regression. Can. J. Fish. Aquat. Sci. 2001, 58, 626–639. [Google Scholar] [CrossRef]
- Clarke, K.R. Non-Parametric Multivariate Analyses of Changes in Community Structure. Austral Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Anderson, M.; Gorley, R.; Clarke, K. PERMANOVA+ for PRIMER. Guide to Software and Statistical Methods; PRIMER-E: Plymouth, UK, 2008. [Google Scholar]
- Plaza-Álvarez, P.A.; Lucas-Borja, M.E.; Sagra, J.; Zema, D.A.; González-Romero, J.; Moya, D.; De las Heras, J. Changes in Soil Hydraulic Conductivity after Prescribed Fires in Mediterranean Pine Forests. J. Environ. Manag. 2019, 232, 1021–1027. [Google Scholar] [CrossRef]
- Sagra, J.; Moya, D.; Plaza-Álvarez, P.A.; Lucas-Borja, M.E.; Alfaro-Sánchez, R.; De Las Heras, J.; Ferrandis, P. Predation on Early Recruitment in Mediterranean Forests after Prescribed Fires. Forests 2017, 8, 243. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, J.; González-Pérez, J.A.; Turmero, A.; Hernández, M.; Ball, A.S.; González-Vila, F.J.; Arias, M.E. Physico-Chemical and Microbial Perturbations of Andalusian Pine Forest Soils Following a Wildfire. Sci. Total Environ. 2018, 634, 650–660. [Google Scholar] [CrossRef] [PubMed]
- Bárcenas-Moreno, G.; García-Orenes, F.; Mataix-Solera, J.; Mataix-Beneyto, J.; Bååth, E. Soil Microbial Recolonisation after a Fire in a Mediterranean Forest. Biol. Fertil. Soils 2011, 47, 261–272. [Google Scholar] [CrossRef]
- Scharenbroch, B.C.; Nix, B.; Jacobs, K.A.; Bowles, M.L. Two Decades of Low-Severity Prescribed Fire Increases Soil Nutrient Availability in a Midwestern, USA Oak (Quercus) Forest. Geoderma 2012, 183–184, 80–91. [Google Scholar] [CrossRef]
- Knelman, J.E.; Graham, E.B.; Trahan, N.A.; Schmidt, S.K.; Nemergut, D.R. Fire Severity Shapes Plant Colonization Effects on Bacterial Community Structure, Microbial Biomass, and Soil Enzyme Activity in Secondary Succession of a Burned Forest. Soil Biol. Biochem. 2015, 90, 161–168. [Google Scholar] [CrossRef]
- Pressler, Y.; Moore, J.C.; Cotrufo, M.F. Belowground Community Responses to Fire: Meta-Analysis Reveals Contrasting Responses of Soil Microorganisms and Mesofauna. Oikos 2019, 128, 309–327. [Google Scholar] [CrossRef]
- D’Ascoli, R.; Rutigliano, F.A.; De Pascale, R.A.; Gentile, A.; De Santo, A.V. Functional Diversity of the Microbial Community in Mediterranean Maquis Soils as Affected by Fires. Int. J. Wildl. Fire 2005, 14, 355–363. [Google Scholar] [CrossRef]
- Cerda, A.; Robichaud, P.R. Forest Fire Effects on Soil Microbiology; CRC Press: Boca Raton, FL, USA, 2009; pp. 149–192. [Google Scholar] [CrossRef]
- Rodríguez, J.; González-Pérez, J.A.; Turmero, A.; Ball, A.S.; González-Vila, F.J.; Enriqueta Arias, M. Wildfire Effects on the Microbial Activity and Diversity in a Mediterranean Forest Soil. Catena 2017, 158, 82–88. [Google Scholar] [CrossRef] [Green Version]
- Dworkin, M. Prokaryotic Life Cycles. Prokaryotes 2006, 2, 140–166. [Google Scholar] [CrossRef]
- Pérez-Valera, E.; Goberna, M.; Faust, K.; Raes, J.; García, C.; Verdú, M. Fire Modifies the Phylogenetic Structure of Soil Bacterial Co-Occurrence Networks. Environ. Microbiol. 2017, 19, 317–327. [Google Scholar] [CrossRef]
- Smith, N.R.; Kishchuk, B.E.; Mohn, W.W. Effects of Wildfire and Harvest Disturbances on Forest Soil Bacterial Communities. Appl. Environ. Microbiol. 2008, 74, 216–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiang, X.; Shi, Y.; Yang, J.; Kong, J.; Lin, X.; Zhang, H.; Zeng, J.; Chu, H. Rapid Recovery of Soil Bacterial Communities after Wildfire in a Chinese Boreal Forest. Sci. Rep. 2014, 4, 3829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bukar, M.; Sodipo, O.; Dawkins, K.; Ramirez, R.; Kaldapa, J.T.; Tarfa, M.; Esiobu, N.; Bukar, M.; Sodipo, O.; Dawkins, K.; et al. Microbiomes of Top and Sub-Layers of Semi-Arid Soils in North-Eastern Nigeria Are Rich in Firmicutes and Proteobacteria with Surprisingly High Diversity of Rare Species. Adv. Microbiol. 2019, 9, 102–118. [Google Scholar] [CrossRef] [Green Version]
- Song, H.-G.; Kim, O.-S.; Yoo, J.-J.; Jeon, S.-O.; Hong, S.-H.; Lee, D.-H.; Ahn, T.-S. Monitoring of Soil Bacterial Community and Some Inoculated Bacteria After Prescribed Fire in Microcosm. J. Microbiol. 2004, 42, 285–291. [Google Scholar] [CrossRef]
- Dooley, S.R.; Treseder, K.K. The Effect of Fire on Microbial Biomass: A Meta-Analysis of Field Studies. Biogeochemistry 2012, 109, 49–61. [Google Scholar] [CrossRef] [Green Version]
- Whitman, T.; Whitman, E.; Woolet, J.; Flannigan, M.D.; Thompson, D.K.; Parisien, M.A. Soil Bacterial and Fungal Response to Wildfires in the Canadian Boreal Forest across a Burn Severity Gradient. Soil Biol. Biochem. 2019, 138, 107571. [Google Scholar] [CrossRef]
- Santacruz-Calvo, L.; González-López, J.; Manzanera, M. Arthrobacter Sp. Nov., a Highly Desiccation-Tolerant, Xeroprotectant-Producing Strain Isolated from Dry Soil. Int. J. Syst. Evol. Microbiol. 2013, 63, 4174. [Google Scholar] [CrossRef]
- Cobo-Díaz, J.F.; Fernández-González, A.J.; Villadas, P.J.; Robles, A.B.; Toro, N.; Fernández-López, M. Metagenomic Assessment of the Potential Microbial Nitrogen Pathways in the Rhizosphere of a Mediterranean Forest After a Wildfire. Microb. Ecol. 2015, 69, 895–904. [Google Scholar] [CrossRef] [Green Version]
- Fernández-González, A.J.; Martínez-Hidalgo, P.; Cobo-Díaz, J.F.; Villadas, P.J.; Martínez-Molina, E.; Toro, N.; Tringe, S.G.; Fernández-López, M. The Rhizosphere Microbiome of Burned Holm-Oak: Potential Role of the Genus Arthrobacter in the Recovery of Burned Soils. Sci. Rep. 2017, 7, 6008. [Google Scholar] [CrossRef]
- Manzanera, M.; Narváez-Reinaldo, J.J.; García-Fontana, C.; Vílchez, J.I.; González-López, J. Genome Sequence of Arthrobacter Koreensis 5J12A, a Plant Growth-Promoting and Desiccation-Tolerant Strain. Genome Announc. 2015, 3, 648–663. [Google Scholar] [CrossRef] [Green Version]
- Mongodin, E.F.; Shapir, N.; Daugherty, S.C.; DeBoy, R.T.; Emerson, J.B.; Shvartzbeyn, A.; Radune, D.; Vamathevan, J.; Riggs, F.; Grinberg, V.; et al. Secrets of Soil Survival Revealed by the Genome Sequence of Arthrobacter Aurescens TC1. PLoS Genet. 2006, 2, e214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, J.; Yang, H.; Long, X.; Liu, Z.; Rengel, Z. Pepino (Solanum muricatum) Planting Increased Diversity and Abundance of Bacterial Communities in Karst Area. Sci. Rep. 2016, 6, 21938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, C.R.; Condron, L.M.; Clough, T.J.; Fiers, M.; Stewart, A.; Hill, R.A.; Sherlock, R.R. Biochar Induced Soil Microbial Community Change: Implications for Biogeochemical Cycling of Carbon, Nitrogen and Phosphorus. Pedobiologia (Jena) 2011, 54, 309–320. [Google Scholar] [CrossRef]
- Soria, R.; Rodríguez-Berbel, N.; Ortega, R.; Lucas-Borja, M.E.; Miralles, I. Soil Amendments from Recycled Waste Differently Affect CO2 Soil Emissions in Restored Mining Soils under Semiarid Conditions. J. Environ. Manag. 2021, 294, 112894. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Air Temperature (°C) | Relative Air Humidity (%) | Wind Velocity (Km h−1) | Spread Rate (m/min) | Flame Length (m) | Mean Surface Temperature (°C) | Max Surface Temperature (°C) | Mean Residence Time (s) | |
|---|---|---|---|---|---|---|---|---|
| PB1 | 18.95 ± 1.98 | 35.6 ± 3.5 | 5.99 ± 4.56 | 1.6–1.87 | 0.2–1.0 | 632.5 | 759.0 | 73.5 |
| PB2 | 20.17 ± 2.95 | 22.20 ± 3.03 | 2.75 ± 3.26 | 0.81–1.71 | 0.5–0.9 | 589.9 | 832.5 | 61.7 |
| EC (mS/cm−1) | pH | TOC (%) | |
|---|---|---|---|
| UB | 0.038 ± 0.00 a | 7.246 ± 0.02 a | 1.370 ± 0.35 a |
| PB1 | 0.030 ± 0.01 a | 7.593 ± 0.20 a | 1.652 ± 0.16 a |
| PB2 | 0.058 ± 0.02 a | 7.393 ± 0.41 a | 1.595 ± 0.02 a |
| ASV Observed | Faith | Shannon | Pielou | |
|---|---|---|---|---|
| UB | 1044 ± 4.58 a | 53.58 ± 0.78 a | 8.97 ± 0.03 a | 0.89 ± 0.00 a |
| PB1 | 1052 ± 17.43 a | 53.40 ± 0.65 a | 8.74 ± 0.03 b | 0.87 ± 0.00 b |
| PB2 | 1047 ± 18.87 a | 54.43 ± 0.85 a | 8.65 ± 0.03 c | 0.86 ± 0.00 c |
| Phylum | Bacterial Taxa | Abundance (%) | Contribution (%) | Cumulative Contribution (%) |
|---|---|---|---|---|
| UB average similarity: 90% | ||||
| Proteobacteria | [F] Xanthobacteraceae | 2.73 | 2.68 | 2.68 |
| Acidobacteria | [C] Subgroup_6 | 1.78 | 1.82 | 4.5 |
| Proteobacteria | Sphingomonas | 1.72 | 1.75 | 6.25 |
| Acidobacteria | RB41 | 1.59 | 1.57 | 7.82 |
| Planctomycetes | Singulisphaera | 1.53 | 1.55 | 9.37 |
| PB1 average similarity: 92% | ||||
| Proteobacteria | [F] Xanthobacteraceae | 3.18 | 3.27 | 3.27 |
| Acidobacteria | RB41 | 1.95 | 1.96 | 5.23 |
| Planctomycetes | Singulisphaera | 1.8 | 1.8 | 7.02 |
| Proteobacteria | Sphingomonas | 1.68 | 1.72 | 8.74 |
| Acidobacteria | [C] Subgroup_6 | 1.56 | 1.59 | 10.33 |
| Actinobacteria | Mycobacterium | 1.56 | 1.53 | 11.86 |
| PB2 average similarity: 90% | ||||
| Proteobacteria | [O] Xanthobacteraceae | 3.33 | 3.49 | 3.49 |
| Planctomycetes | Singulisphaer | 1.69 | 1.73 | 5.22 |
| Proteobacteria | Sphingomonas | 1.59 | 1.61 | 6.83 |
| Acidobacteria | RB41 | 1.55 | 1.57 | 8.4 |
| Acidobacteria | Subgroup_6 | 1.52 | 1.51 | 9.91 |
| Phylum | Bacterial Taxa | Av. UB | Av. PB1 | Contribution (%) | Cumulative Contribution (%) |
|---|---|---|---|---|---|
| UB and PB1 dissimilarity = 9% | |||||
| Actinobacteria | Mycobacterium | 1.02 | 1.56 | 2.53 | 2.53 |
| Proteobacteria | [F] Xanthobacteraceae | 2.73 | 3.18 | 2.14 | 4.68 |
| Acidobacteria | [O] Subgroup 7 | 0.36 | 0.12 | 1.88 | 6.55 |
| Firmicutes | Cohnella | 0.32 | 0.69 | 1.75 | 8.30 |
| Acidobacteria | RB41 Subgroup 4 uncult. | 1.59 | 1.95 | 1.73 | 10.03 |
| Acidobacteria | soil bacterium clone S111 | 0.31 | 0.27 | 1.56 | 11.59 |
| UB and PB2 dissimilarity 13% | |||||
| Proteobacteria | Massilia | 0.63 | 1.25 | 2.28 | 2.28 |
| Proteobacteria | [F] Xanthobacteraceae | 2.73 | 3.33 | 2.20 | 4.48 |
| Proteobacteria | Pseudomonas | 0.11 | 0.71 | 2.19 | 6.67 |
| Acidobacteria | Subgroup 10 | 1.41 | 0.85 | 2.05 | 8.73 |
| Firmicutes | Bacillus | 0.89 | 1.37 | 1.75 | 10.47 |
| Firmicutes | [O] Bacillales | 0.46 | 0.89 | 1.57 | 12.04 |
| Proteobacteria | Luteibacter | 0.27 | 0.62 | 1.54 | 13.58 |
| Proteobacteria | [F] Archangiaceae | 0.64 | 0.23 | 1.53 | 15.11 |
| Actinobacteria | Mycobacterium | 1.02 | 1.43 | 1.50 | 16.61 |
| Proteobacteria | Massilia | 0.63 | 1.25 | 2.28 | 2.28 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soria, R.; Tortosa, A.; Rodríguez-Berbel, N.; Lucas-Borja, M.E.; Ortega, R.; Miralles, I. Short-Term Response of Soil Bacterial Communities after Prescribed Fires in Semi-Arid Mediterranean Forests. Fire 2023, 6, 145. https://doi.org/10.3390/fire6040145
Soria R, Tortosa A, Rodríguez-Berbel N, Lucas-Borja ME, Ortega R, Miralles I. Short-Term Response of Soil Bacterial Communities after Prescribed Fires in Semi-Arid Mediterranean Forests. Fire. 2023; 6(4):145. https://doi.org/10.3390/fire6040145
Chicago/Turabian StyleSoria, Rocío, Antonio Tortosa, Natalia Rodríguez-Berbel, Manuel Esteban Lucas-Borja, Raúl Ortega, and Isabel Miralles. 2023. "Short-Term Response of Soil Bacterial Communities after Prescribed Fires in Semi-Arid Mediterranean Forests" Fire 6, no. 4: 145. https://doi.org/10.3390/fire6040145