
Torulaspora delbrueckii Improves Organoleptic Properties and Promotes Malolactic Fermentation in Carbonic Maceration Wines


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Carbonic Maceration and Alcoholic Fermentation
2.2. Malolactic Fermentation
2.3. Area under the Curve (AUC)
2.4. Physicochemical Analysis
2.4.1. General Oenological Parameters
2.4.2. Volatile Composition
2.4.3. Color and Anthocyanin Analysis
2.5. Sensory Analysis
2.6. Statistical Analysis
3. Results
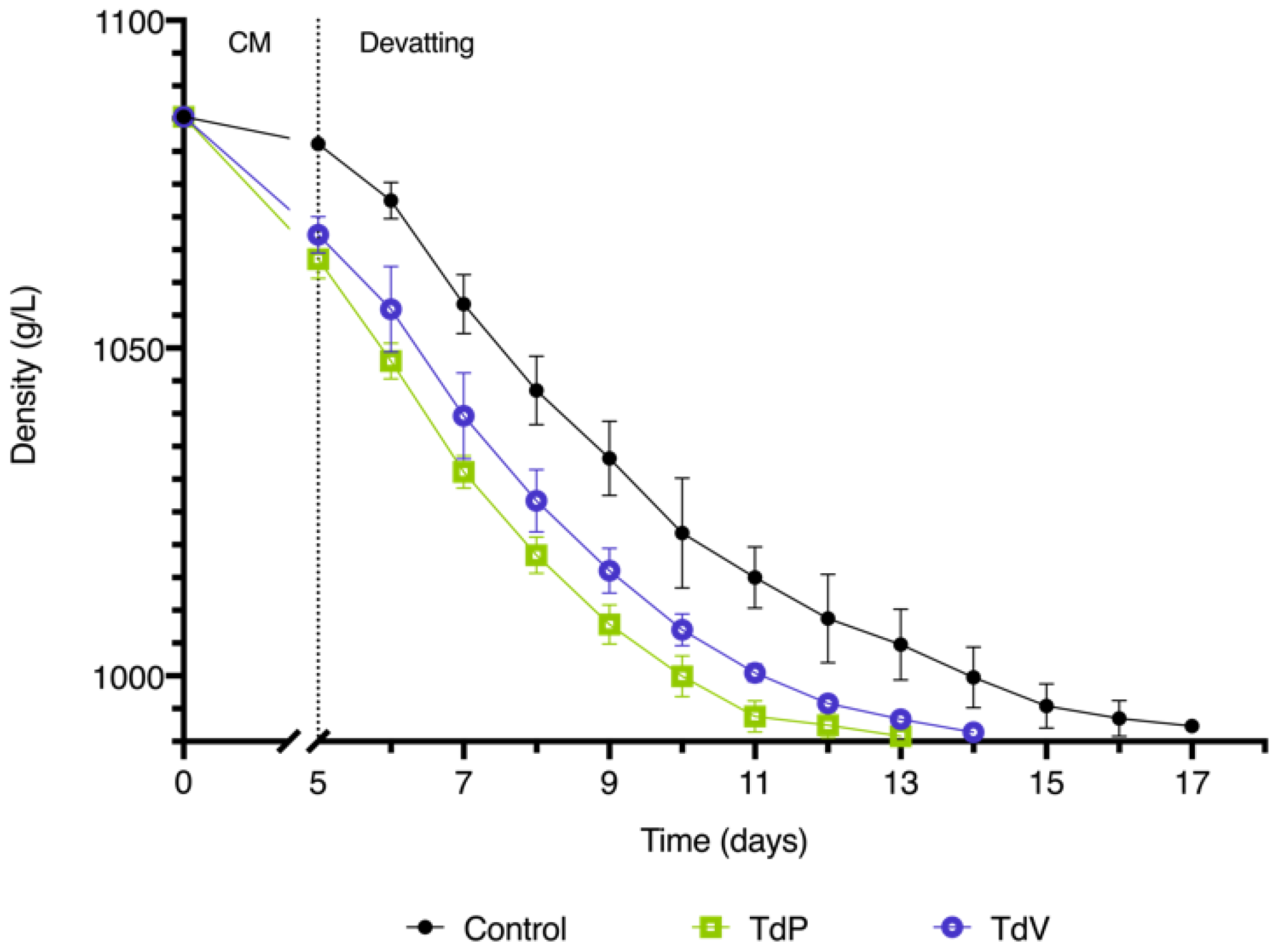
3.1. Alcoholic Fermentation
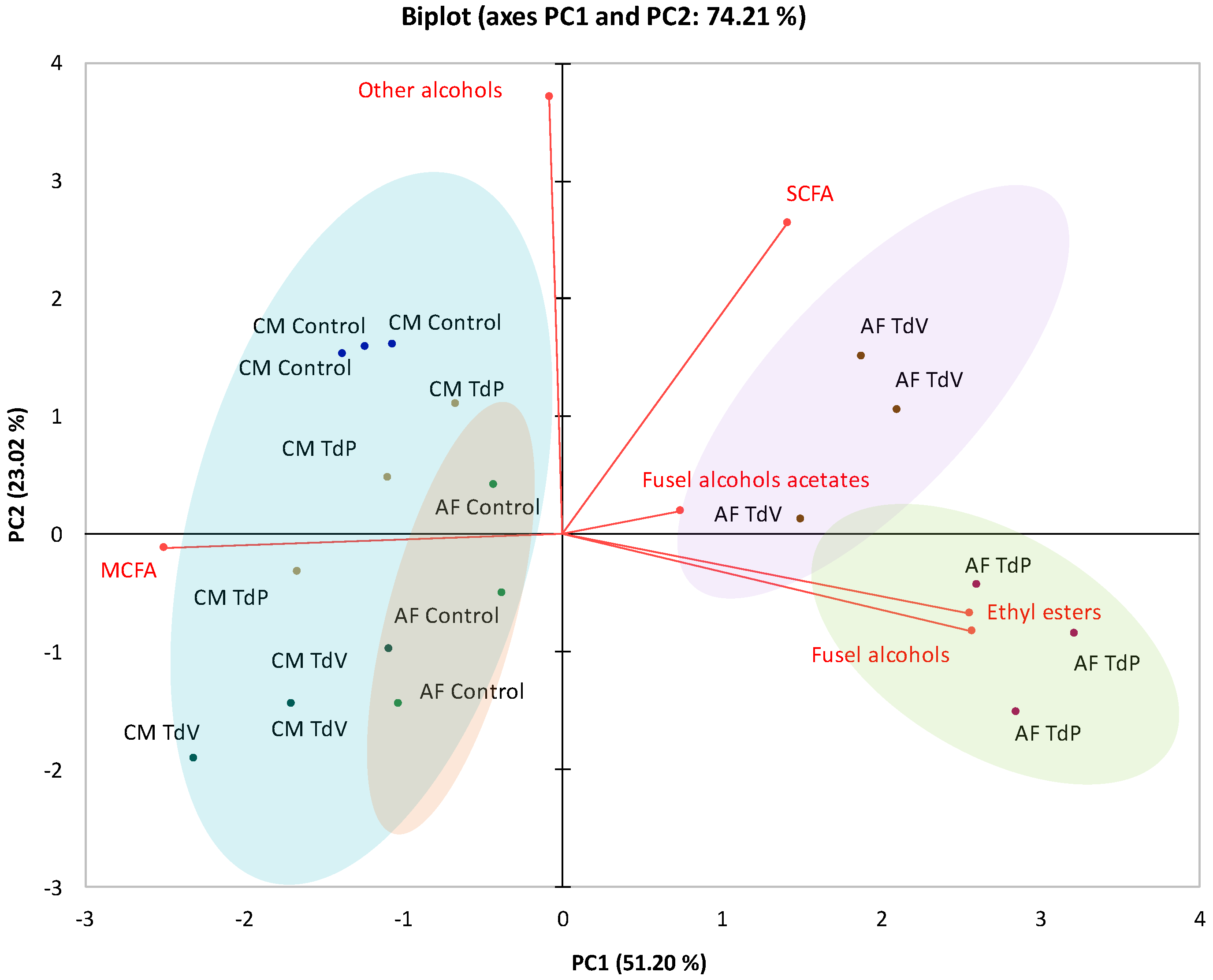
3.2. Volatile Composition
3.3. Color Parameters and Anthocyanins Composition
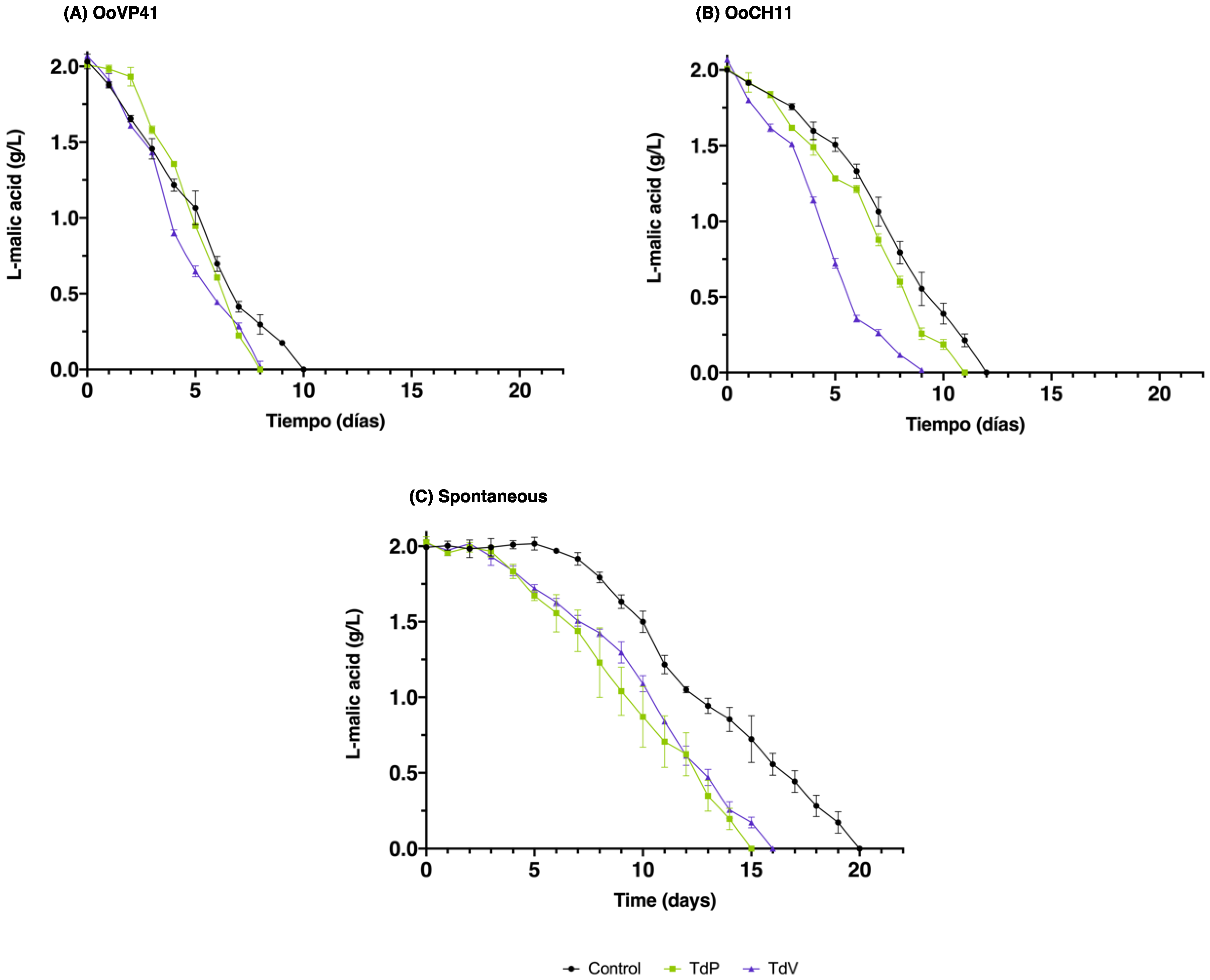
3.4. Malolactic Fermentation
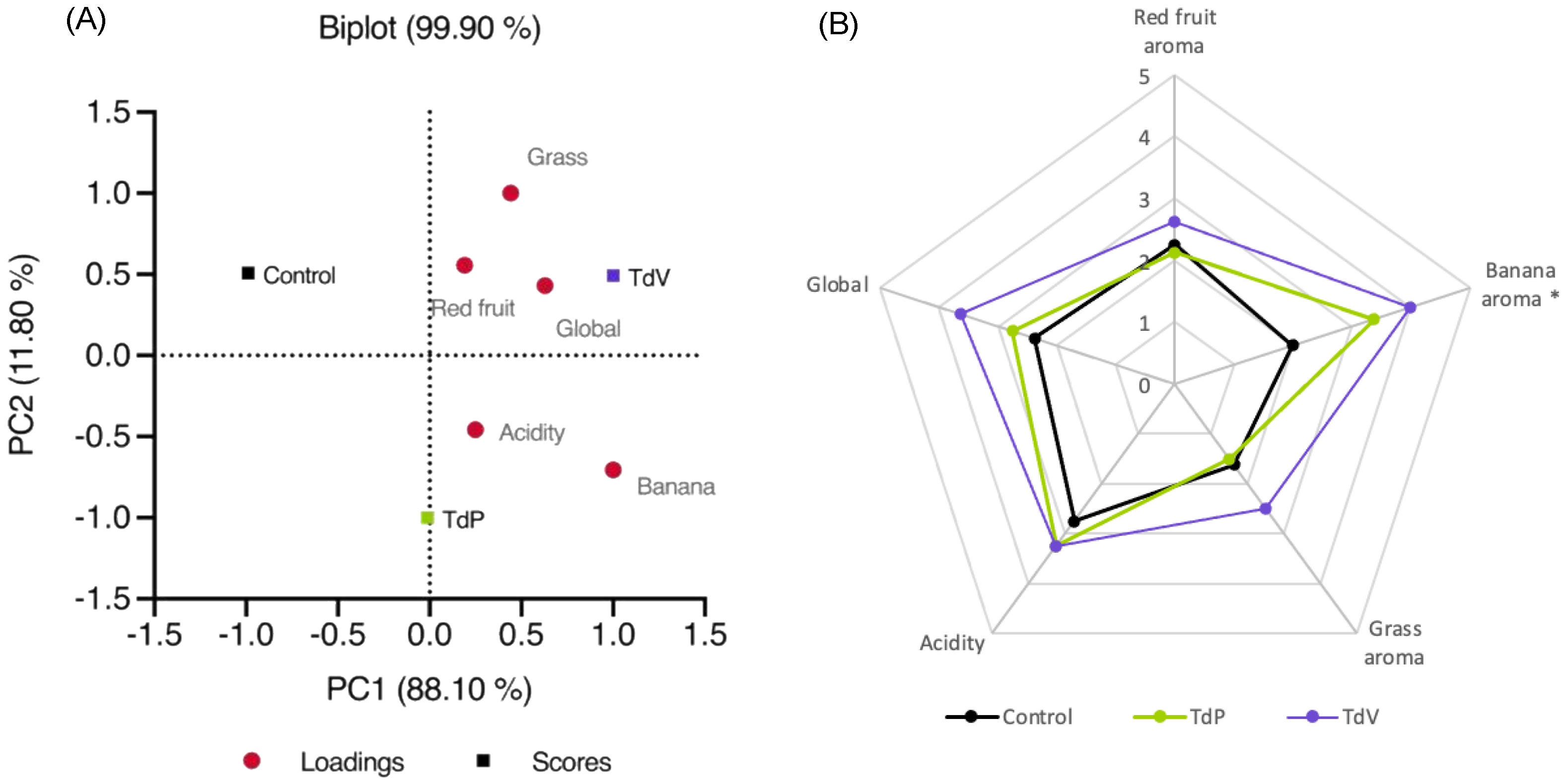
3.5. Organoleptic Characteristics
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Tesnière, C.; Flanzy, C. Carbonic maceration wines: Characteristics and winemaking process. In Advances in Food and Nutrition Research; Academic Press Inc.: Cambridge, MA, USA, 2011; Volume 63, pp. 1–15. [Google Scholar] [CrossRef]
- Portu, J.; Gutiérrez-Viguera, A.R.; González-Arenzana, L.; Santamaría, P. Characterization of the color parameters and monomeric phenolic composition of ‘Tempranillo’ and ‘Graciano’ wines made by carbonic maceration. Food Chem. 2023, 406, 134327. [Google Scholar] [CrossRef]
- Antalick, G.; Perello, M.C.; de Revel, G. Esters in wines: New insight through the establishment of a database of French wines. Am. J. Enol. Vitic. 2014, 65, 293–304. [Google Scholar] [CrossRef]
- Etaio, I.; Meillon, S.; Pérez-Elortondo, F.J.; Schlich, P. Dynamic sensory description of Rioja Alavesa red wines made by different winemaking practices by using Temporal Dominance of Sensations. J. Sci. Food Agric. 2016, 96, 3492–3499. [Google Scholar] [CrossRef]
- González-Arenzana, L.; Santamaría, R.; Escribano-Viana, R.; Portu, J.; Garijo, P.; López-Alfaro, I.; López, R.; Santamaría, P.; Gutiérrez, A.R. Influence of the carbonic maceration winemaking method on the physicochemical, color, aromatic and microbiological features of tempranillo red wines. Food Chem. 2020, 319, 126569. [Google Scholar] [CrossRef]
- Chinnici, F.; Sonni, F.; Natali, N.; Galassi, S.; Riponi, C. Color Features and Pigment Composition of Italian Carbonic Macerated Red Wines. Food Chem. 2009, 113, 651–657. [Google Scholar] [CrossRef]
- Guzzon, R.; Malacarne, M.; Larcher, R.; Franciosi, E.; Toffanin, A. The impact of grape processing and carbonic maceration on the microbiota of early stages of winemaking. J. Appl. Microbiol. 2022, 128, 209–224. [Google Scholar] [CrossRef]
- Gutiérrez, A.R.; Santamaría, P.; Olarte, C.; López-Alfaro, I.; Garijo, P.; González-Arenzana, L.; Sanz, S. Influence of microbial population on the characteristics of carbonic maceration wines. LWT 2022, 166, 113783. [Google Scholar] [CrossRef]
- Renault, P.; Coulon, J.; de Revel, G.; Barbe, J.C.; Bely, M. Increase of fruity aroma during mixed T. delbrueckii/S. cerevisiae wine fermentation is linked to specific esters enhancement. Int. J. Food Microbiol. 2015, 207, 40–48. [Google Scholar] [CrossRef]
- Azzolini, M.; Tosi, E.; Lorenzini, M.; Finato, F.; Zapparoli, G. Contribution to the aroma of white wines by controlled Torulaspora delbrueckii cultures in association with Saccharomyces cerevisiae. World J. Microbiol. Biotechnol. 2015, 31, 277–293. [Google Scholar] [CrossRef]
- Chen, K.; Escott, C.; Loira, I.; del Fresno, J.M.; Morata, A.; Tesfaye, W.; Calderon, F.; Suárez-Lepe, J.A.; Han, S.; Benito, S. Use of non-Saccharomyces yeasts and oenological tannin in red winemaking: Influence on color, aroma and sensorial properties of young wines. Food Microbiol. 2018, 69, 51–63. [Google Scholar] [CrossRef]
- Balmaseda, A.; Rozès, N.; Bordons, A.; Reguant, C. Torulaspora delbrueckii promotes malolactic fermentation in high polyphenolic red wines. LWT 2021, 148, 111777. [Google Scholar] [CrossRef]
- Muñoz-Redondo, J.M.; Puertas, B.; Cantos-Villar, E.; Jiménez-Hierro, M.J.; Carbú, M.; Garrido, C.; Ruiz-Moreno, M.J.; Moreno-Rojas, J.M. Impact of Sequential Inoculation with the Non-Saccharomyces T. delbrueckii and M. pulcherrima Combined with Saccharomyces cerevisiae Strains on Chemicals and Sensory Profile of Rosé Wines. J. Agric. Food Chem. 2021, 69, 1598–1609. [Google Scholar] [CrossRef]
- Bely, M.; Stoeckle, P.; Masneuf-Pomarède, I.; Dubourdieu, D. Impact of mixed Torulaspora delbrueckii-Saccharomyces cerevisiae culture on high-sugar fermentation. Int. J. Food Microbiol. 2008, 122, 312–320. [Google Scholar] [CrossRef]
- Benito, S. The impact of Torulaspora delbrueckii yeast in winemaking. Appl. Microbiol. Biotechnol. 2018, 102, 3081–3094. [Google Scholar] [CrossRef]
- Belda, I.; Navascués, E.; Marquina, D.; Santos, A.; Calderon, F.; Benito, S. Dynamic analysis of physiological properties of Torulaspora delbrueckii in wine fermentations and its incidence on wine quality. Appl. Microbiol. Biotechnol. 2015, 99, 1911–1922. [Google Scholar] [CrossRef]
- Martín-García, A.; Balmaseda, A.; Bordons, A.; Reguant, C. Effect of the inoculation strategy of non-Saccharomyces yeasts on wine malolactic fermentation. Oeno One 2020, 54, 101–108. [Google Scholar] [CrossRef]
- Ferrando, N.; Araque, I.; Ortís, A.; Thornes, G.; Bautista-Gallego, J.; Bordons, A.; Reguant, C. Evaluating the effect of using non-Saccharomyces on Oenococcus oeni and wine malolactic fermentation. Food Res. Int. 2020, 138, 109779. [Google Scholar] [CrossRef]
- Ruiz-de-Villa, C.; Poblet, M.; Bordons, A.; Reguant, C.; Rozès, N. Comparative Study of Inoculation Strategies of Torulaspora delbrueckii and Saccharomyces cerevisiae on the Performance of Alcoholic and Malolactic Fermentations in an Optimized Synthetic Grape Must. Int. J. Food Microbiol. 2023, 404, 110367. [Google Scholar] [CrossRef]
- Gutiérrez, A.R.; Santamaría, P.; González-Arenzana, L.; Garijo, P.; Olarte, C.; Sanz, S. Yeasts Inoculation Effect on Bacterial Development in Carbonic Maceration Wines Elaboration. Foods 2023, 12, 2755. [Google Scholar] [CrossRef]
- De Man, J.C.; Rogosa, M.; Sharpe, M.E. A medium for the cultivation of Lactobacilli. J. Appl. Bacteriol. 1960, 23, 130–135. [Google Scholar] [CrossRef]
- Quirós, M.; Rojas, V.; Gonzalez, R.; Morales, P. Selection of non-Saccharomyces yeast strains for reducing alcohol levels in wine by sugar respiration. Int. J. Food Microbiol. 2014, 181, 85–91. [Google Scholar] [CrossRef]
- Balmaseda, A.; Aniballi, L.; Rozès, N.; Bordons, A.; Reguant, C. Use of yeast mannoproteins by Oenococcus oeni during malolactic fermentation under different oenological conditions. Foods 2021, 10, 1540. [Google Scholar] [CrossRef]
- Ortega, C.; Lopez, R.; Cacho, J.; Ferreira, V. Fast analysis of important wine volatile compounds Development and validation of a new method based on gas chromatographic-flame ionisation detection analysis of dichloromethane microextracts. J. Chromatogr. A 2001, 923, 205–214. [Google Scholar] [CrossRef]
- Ayala, F.; Echavarri, J.F.; Negueruela, A.I. A New Simplified Method for Measuring the Color of Wines. I. Red and Ros Wines. Am. J. Enol. Vitic. 1997, 48, 357–363. [Google Scholar] [CrossRef]
- Glories, Y. La couleur des vins rouges. 2ème partie: Mesure, origine et interprétation. Connaiss. Vigne Vin 1984, 18, 253–271. [Google Scholar]
- Niketic-Aleksic, G.K.; Hrazdina, G. Quantitative analysis of the anthocyanin content in grape juices and wines. Lebensm. Wiss. Technol. 1972, 5, 163–165. [Google Scholar]
- Roca-Mesa, H.; Delgado-Yuste, E.; Mas, A.; Torija, M.J.; Beltran, G. Importance of micronutrients and organic nitrogen in fermentations with Torulaspora delbrueckii and Saccharomyces cerevisiae. Int. J. Food Microbiol. 2022, 381, 109915. [Google Scholar] [CrossRef]
- Contreras, A.; Hidalgo, C.; Henschke, P.A.; Chambers, P.J.; Curtin, C.; Varela, C. Evaluation of non-Saccharomyces yeasts for the reduction of alcohol content in wine. Appl. Environ. Microbiol. 2014, 80, 1670–1678. [Google Scholar] [CrossRef]
- Zhu, X.; Navarro, Y.; Mas, A.; Torija, M.J.; Beltran, G. A rapid method for selecting non-Saccharomyces strains with a low ethanol yield. Microorganisms 2020, 8, 658. [Google Scholar] [CrossRef]
- Ribéreau-Gayon, P.; Dubourdieu, D.; Donèche, B.; Lonvaud, A. Handbook of Enology, Volume 1: The Microbiology of Wine and Vinification, 2nd ed.; John Wiley & Sons Ltd.: Chichester, UK, 2006. [Google Scholar]
- Escribano-Viana, R.; Portu, J.; Garijo, P.; López, R.; Santamaría, P.; López-Alfaro, I.; Gutiérrez, A.R.; González-Arenzana, L. Effect of the sequential inoculation of non-Saccharomyces/Saccharomyces on the anthocyans and stilbenes composition of Tempranillo wines. Front. Microbiol. 2019, 10, 773. [Google Scholar] [CrossRef]
- Domizio, P.; Liu, Y.; Bisson, L.F.; Barile, D. Use of non-Saccharomyces wine yeasts as novel sources of mannoproteins in wine. Food Microbiol. 2014, 43, 5–15. [Google Scholar] [CrossRef]
- Salinas, M.R.; Alonso, G.L.; Navarro, G.; Pardo, F.; Jimeno, J.; Huerta, M.D. Evolution of the Aromatic Composition of Wines Undergoing Carbonic Maceration Under Different Aging Conditions. Am. J. Enol. Vitic. 1996, 47, 134–144. [Google Scholar] [CrossRef]
- Sumby, K.M.; Grbin, P.R.; Jiranel, V. Microbial modulation of aromatic esters in wine: Current knowledge and future prospects. Food Chem. 2010, 121, 1–16. [Google Scholar] [CrossRef]
- Ruiz-de-Villa, C.; Poblet, M.; Bordons, A.; Reguant, C.; Rozès, N. Screening of Saccharomyces cerevisiae and Torulaspora delbrueckii strains in relation to their effect on malolactic fermentation. LWT 2023, 112, 104212. [Google Scholar]
- Gombau, J.; Pons-Mercadé, P.; Conde, M.; Asbiro, L.; Pascual, O.; Gómez-Alonso, S.; García-Romero, E.; Miquel Canals, J.; Hermosín-Gutiérrez, I.; Zamora, F. Influence of grape seeds on wine composition and astringency of Tempranillo, Garnacha, Merlot and Cabernet Sauvignon wines. Food Sci. Nutr. 2020, 8, 3442–3455. [Google Scholar] [CrossRef]
- Minnaar, P.; Nyobo, L.; Jolly, N.; Ntushelo, N.; Meiring, S. Anthocyanins and polyphenols in Cabernet Franc wines produced with Saccharomyces cerevisiae and Torulaspora delbrueckii yeast strains: Spectrophotometric analysis and effect on selected sensory attributes. Food Chem. 2018, 268, 287–291. [Google Scholar] [CrossRef]
- Belda, I.; Conchillo, L.B.; Ruiz, J.; Navascués, E.; Marquina, D.; Santos, A. Selection and Use of Pectinolytic Yeasts for Improving Clarification and Phenolic Extraction in Winemaking. Int. J. Food Microbiol. 2016, 223, 1–8. [Google Scholar] [CrossRef]
- Vidana Gamage, G.C.; Lim, Y.Y.; Choo, W.S. Sources and relative stabilities of acylated and nonacylated anthocyanins in beverage systems. J. Food Sci. Technol. 2022, 59, 831–845. [Google Scholar] [CrossRef]
- Maturano, Y.P.; Rodríguez Assaf, L.A.; Toro, M.E.; Nally, M.C.; Vallejo, M.; Castellanos de Figueroa, L.I.; Combina, M.; Vazquez, F. Multi-enzyme production by pure and mixed cultures of Saccharomyces and non-Saccharomyces yeasts during wine fermentation. Int. J. Food Microbiol. 2012, 155, 43–50. [Google Scholar] [CrossRef]
- Morata, A.; Gómez-Cordovés, M.C.; Suberviola, J.; Bartolomé, B.; Colomo, B.; Suárez, J.A. Adsorption of anthocyanins by yeast cell walls during the fermentation of red wines. J. Agric. Food Chem. 2003, 51, 4084–4088. [Google Scholar] [CrossRef]
- Tofalo, R.; Suzzi, G.; Perpetuini, G. Discovering the Influence of Microorganisms on Wine Color. Front. Microbiol. 2021, 12, 790935. [Google Scholar] [CrossRef]
- Rivas-Gonzalo, J.C.; Gutierrez, T.; Hebrero, E.; Santos-Buelga, C. Comparisons of Methods for the Determination of Anthocyanins in Red Wines. Am. J. Enol. Vitic. 1992, 43, 210–214. [Google Scholar] [CrossRef]
- Fulcrand, H.; Benabdeljalil, C.; Rigaud, J.; Cheynier, R.; Moljtounet, M. A new class of wine pigments generated by reaction between pyruvic acid and grape anthocyanins. Phytochemistry 1997, 47, 1401–1407. [Google Scholar] [CrossRef]
- Capucho, I.; San Romão, M.V. Effect of ethanol and fatty acids on malolactic activity of Leuconostoc oenos. Appl. Microbiol. Biotechnol. 1994, 42, 391–395. [Google Scholar] [CrossRef]
- Lafon-Lafourcade, S.; Geneix, C.; Ribéreau-Gayon, P. Inhibition of Alcoholic Fermentation of Grape Must by Fatty Acids Produced by Yeasts and Their Elimination by Yeast Ghosts. Appl. Environ. Microbiol. 1984, 47, 1246–1249. [Google Scholar] [CrossRef]
- Verdugo-Vásquez, N.; Orrego, R.; Gutiérrez-Gamboa, G.; Reyes, M.; Zurita Silva, A.; Balbontín, C.; Gaete, N.; Salazar-Parra, C. Trends and climatic variability in the Chilean viticultural production zones: Three decades of climatic data (1985–2015). OENO One 2023, 57, 345–362. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| General Oenological Parameters | End of CM | End of AF | ||||
|---|---|---|---|---|---|---|
| CM Control | CM TdP | CM TdV | AF Control | AF TdP | AF TdV | |
| AUCs | 5416 ± 1 a | 5372 ± 7 b | 5381 ± 7 b | 11,287 ± 49 B | 8119 ± 17 A | 9165 ± 28 A |
| Glycerol (g/L) | 2.04 ± 0.12 a | 3.43 ± 0.79 a | 3.38 ± 0.55 a | 7.51 ± 0.76 AB | 6.56 ± 0.59 A | 7.55 ± 0.09 B |
| Ethanol (% v/v) | 1.06 ± 0.04 a | 2.99 ± 0.28 b | 2.84 ± 0.16 b | 11.06 ± 0.21 A | 11.58 ± 0.16 A | 11.70 ± 0.09 A |
| Ammonia (mg/L) | 68 ± 9 b | 40 ± 11 a | 46 ± 6 a | n.d. | 11 ± 4 B | 7 ± 5 AB |
| α-amino nitrogen (mg/L) | 109 ± 3 b | 60 ± 7 a | 97 ± 7 b | 19 ± 7 A | 27 ± 4 A | 22 ± 3 A |
| Succinic acid (g/L) | 0.42 ± 0.02 b | 0.27 ± 0.07 a | 0.31 ± 0.10 ab | 0.34 ± 0.04 A | 0.31 ± 0.05 A | 0.29 ± 0.03 A |
| Citric acid (g/L) | 0.26 ± 0.04 a | 0.34 ± 0.10 a | 0.28 ± 0.04 a | 0.23 ± 0.02 A | 0.25 ± 0.03 A | 0.25 ± 0.05 A |
| Acetic acid (g/L) | 0.34 ± 0.07 b | 0. ± 0.02 a | 0.61 ± 0.06 ab | 0.54 ± 0.10 A | 0.47 ± 0.05 A | 0.31 ± 0.06 A |
| L-malic acid (g/L) | n.a. | n.a. | n.a. | 1.73 ± 0.04 A | 1.64 ± 0.04 B | 1.58 ± 0.04 C |
| pH | n.a. | n.a. | n.a. | 3.21 ± 0.03 A | 3.25 ± 0.03 A | 3.27 ± 0.04 A |
| Eq. mannose (mg/L) | n.a. | n.a. | n.a. | 114 ± 13 A | 301 ± 7 B | 311 ± 8 B |
| Phenolic Compounds and Color Parameters | End of CM | End of AF | ||||
|---|---|---|---|---|---|---|
| CM Control | CM TdP | CM TdV | AF Control | AF TdP | AF TdV | |
| Total anthocyanins (mg/L) quantified with spectrophotometry | 61 ± 2 a | 106 ± 4 b | 118 ± 3 b | 49 ± 3 A | 114 ± 17 B | 112 ± 7 B |
| Free anthocyanins (mg/L) quantified with HPLC | 20 ± 7 a | 85 ± 13 bc | 97 ± 23 c | 24 ± 2 A | 69 ± 3 BC | 66 ± 7 C |
| Pyranoanthocyanins (mg/L) | n.d. | 2.43 ± 0.10 d | 2.46 ± 0.12 d | 1.76 ± 0.01 B | 2.08 ± 0.06 C | 2.06 ± 0.03 C |
| L* | 53 ± 2 bc | 45 ± 1 a | 45 ± 3 a | 59 ± 1 C | 48 ± 2 AB | 48 ± 4 AB |
| C* | 48 ± 2 b | 51 ± 1 b | 53 ± 5.63 b | 33 ± 3 A | 53 ± 4 B | 52 ± 3 B |
| H* | 46.2 ± 4.8 c | 22.5 ± 6.4 b | 23.0 ± 1.9 b | 17.9 ± 0.4 AB | 12.6 ± 1.7 A | 11.1 ± 1.6 A |
| a* | 33 ± 3 a | 47 ± 3 b | 50 ± 4 b | 33 ± 2 A | 49 ± 7 B | 51 ± 3 B |
| Bb* | 34.6 ± 2.8 c | 19.3 ± 4.9 b | 21.2 ± 0.1 b | 10.6 ± 0.4 A | 10.8 ± 1.9 A | 9.9 ± 0.8 A |
| CI | 2.45 ± 0.18 b | 2.79 ± 0.09 b | 2.76 ± 0.13 b | 1.48 ± 0.34 a | 2.55 ± 0.26 b | 2.52 ± 0.23 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruiz-de-Villa, C.; Gombau, J.; Poblet, M.; Bordons, A.; Canals, J.M.; Zamora, F.; Reguant, C.; Rozès, N. Torulaspora delbrueckii Improves Organoleptic Properties and Promotes Malolactic Fermentation in Carbonic Maceration Wines. Fermentation 2023, 9, 1021. https://doi.org/10.3390/fermentation9121021
Ruiz-de-Villa C, Gombau J, Poblet M, Bordons A, Canals JM, Zamora F, Reguant C, Rozès N. Torulaspora delbrueckii Improves Organoleptic Properties and Promotes Malolactic Fermentation in Carbonic Maceration Wines. Fermentation. 2023; 9(12):1021. https://doi.org/10.3390/fermentation9121021
Chicago/Turabian StyleRuiz-de-Villa, Candela, Jordi Gombau, Montse Poblet, Albert Bordons, Joan Miquel Canals, Fernando Zamora, Cristina Reguant, and Nicolas Rozès. 2023. "Torulaspora delbrueckii Improves Organoleptic Properties and Promotes Malolactic Fermentation in Carbonic Maceration Wines" Fermentation 9, no. 12: 1021. https://doi.org/10.3390/fermentation9121021