
Effect of Spring Maize Genotypes on Fermentation and Nutritional Value of Whole Plant Maize Silage in Northern Pakistan
 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
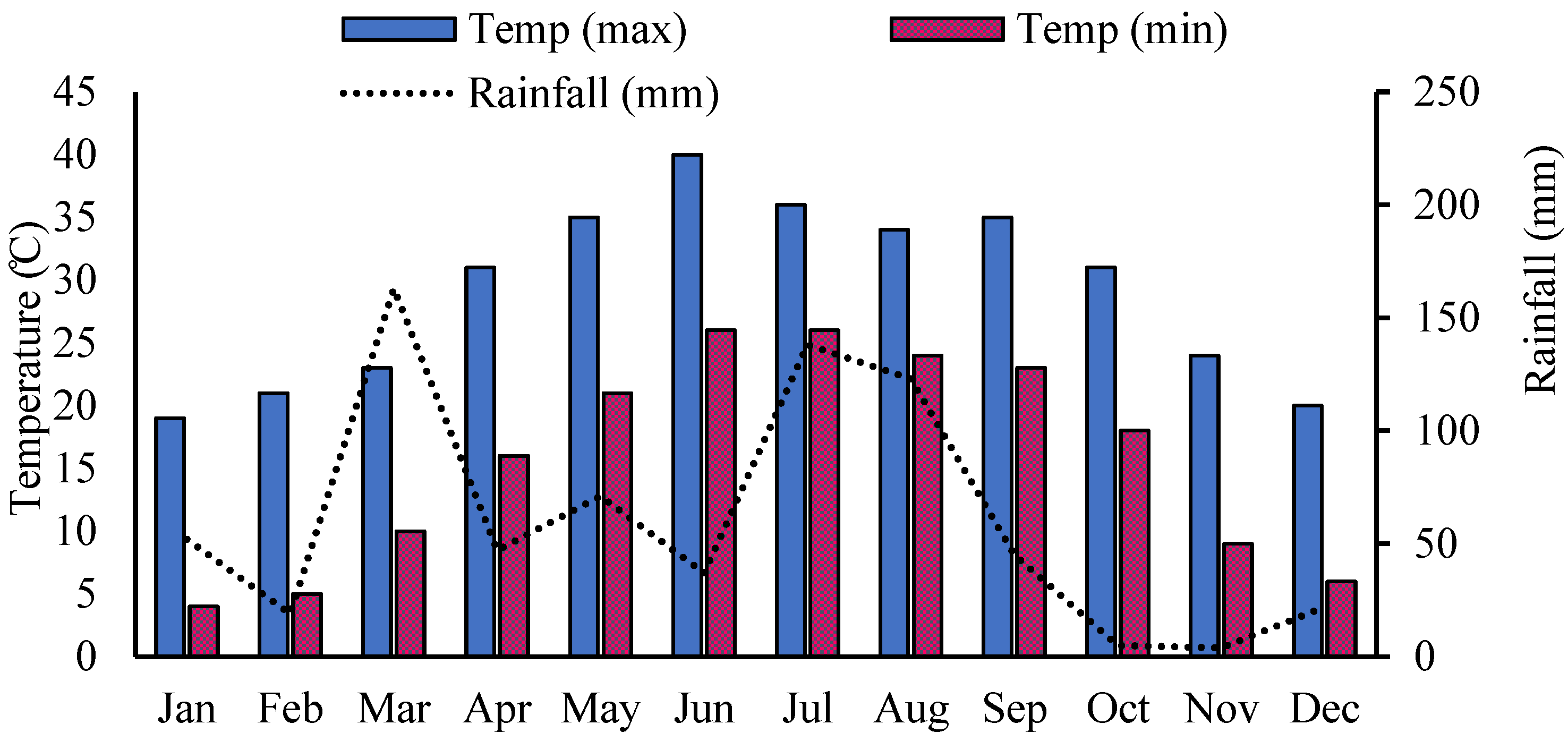
2.1. Experimental Design and Crop Management
2.2. Land Preparation and Crop Management
2.3. Monitoring of Maize Growth, Maturity, and Sampling
2.4. Laboratory-Scale Silage Preparation
2.5. Fermentation Quality
2.6. In Sacco Digestibility and Degradability
2.7. The CNCPS Carbohydrate Subfractions, Digestible Nutrients, and Energy Values
2.8. Statistical Analysis
3. Results
3.1. Growth and Phonological Characteristics
3.2. Biomass and Nutrient Yield
3.3. Chemical Composition
3.4. Rumen Degradation Characteristics of Maize Silages of Different Maize Genotypes
3.5. Fermentation Characteristics
3.6. Total Digestible Nutrients and Estimated Energy Values
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Johnson, L.M.; Harrison, J.H.; Davidson, D.; Swift, M.; Mahanna, W.C. Corn silage management II: Effects of hybrid, maturity, and mechanical processing on digestion and energy content. J. Dairy Sci. 2002, 85, 2913–2927. [Google Scholar] [CrossRef]
- Khan, N.A.; Yu, P.Q.; Ali, M.; Cone, J.W.; Hendriks, W.H. Nutritive value of maize silage in relation to dairy cow performance and milk quality. J. Sci. Food Agric. 2015, 95, 238–252. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Ding, W.; Ke, W.; Li, F.; Zhang, P.; Guo, X. Modulation of metabolome and bacterial community in whole crop corn silage by inoculating homofermentative Lactobacillus plantarum and heterofermentative Lactobacillus buchneri. Front. Microbiol. 2019, 9, 3299. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Zhou, X.; Gu, Q.; Liang, M.; Mu, S.; Zhou, B. Analysis of the correlation between bacteria and fungi in sugarcane tops silage prior to and after aerobic exposure. Bioresour. Technol. 2019, 291, 121835. [Google Scholar] [CrossRef] [PubMed]
- Kung, L., Jr.; Shaver, R.D.; Grant, R.J.; Schmidt, R.J. Silage review: Interpretation of chemical, microbial, and organoleptic components of silages. J. Dairy Sci. 2018, 101, 4020–4033. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.A.; Cone, J.W.; Fievez, V.; Hendriks, W.H. Causes of variation in fatty acid content and composition in grass and maize silages. Anim. Feed Sci. Technol. 2012, 174, 3–45. [Google Scholar] [CrossRef]
- Abeysekara, S.; Khan, N.A.; Yu, P. Relationship between protein molecular structural makeup and metabolizable protein supply to dairy cattle from new cool-season forage corn cultivars. Acta Mol. Biomol. Spectrosc. 2018, 191, 303–314. [Google Scholar] [CrossRef] [PubMed]
- Bernardes, T.F.; Daniel, J.L.P.; Adesogan, A.T.; McAllister, T.A.; Drouin, P.; Nussio, L.G.; Huhtanen, P.; TremblayN, G.F.; Bélanger, G.; Cai, Y. Silage review: Unique challenges of silages made in hot and cold regions. J. Dairy Sci. 2018, 101, 4001–4019. [Google Scholar] [CrossRef] [PubMed]
- Guan, H.; Yan, Y.; Li, X.; Li, X.; Shuai, Y.; Feng, G.; Ran, Q.; Cai, Y.; Li, Y.; Zhang, X. Microbial communities and natural fermentation of corn silages prepared with farm bunker-silo in Southwest China. Bioresour. Technol. 2018, 265, 282–290. [Google Scholar] [CrossRef]
- Bal, M.A.; Shaver, R.D.; Shinners, K.J.; Coors, J.G.; Lauer, J.G.; Straub, R.J.; Koegel, R.G. Stage of maturity, processing, and genotype effects on ruminal in situ disappearance of whole plant corn silage. Anim. Feed Sci. Technol. 2000, 86, 83–94. [Google Scholar] [CrossRef]
- Masoero, F.; Rossi, F.; Pulimeno, A.M. Chemical composition and in vitro digestibility of stalks, leaves and cobs of four corn hybrids at different phonological stages. Ital. J. Anim. Sci. 2006, 5, 215–227. [Google Scholar] [CrossRef]
- Der Bedrosian, M.C.; Kung, L.; Nestor, K.E. The effects of hybrid, maturity, and length of storage on the composition and nutritive value of corn silage. J. Dairy Sci. 2012, 5, 5115–5126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, N.A.; Alam, M.; Khan, R.; Khan, K.; Rahman, S.U. Evaluating the nutritional value of the newly developed quality protein maize in Pakistan: Impact on broiler performance and profitability. Pak. J. Zool. 2020, 52, 585–592. [Google Scholar] [CrossRef]
- Hangshu, X.; Khan, N.A.; Sun, K.; Sun, F.; Sadeeq, U.R.; Fu, Q.; Li, Y.; Zhang, Y.; Hu, G. Batch-to-batch variation in carbohydrates molecular structures, nutritive value and ruminal metabolism in corn coproducts. Anim. Feed. Sci. Technol. 2020, 263, 114458. [Google Scholar]
- Refat, B.; Prates, L.L.; Khan, N.A.; Lei, Y.; Christensen, D.A.; McKinnon, J.J.; Yu, P. Physiochemical characteristics and molecular structures for digestible carbohydrates of silages. J. Agric. Food Chem. 2017, 65, 8979–8991. [Google Scholar] [CrossRef]
- Association of Official and Analytical Chemist (AOAC). Official Methods of Analysis, 18th ed.; AOAC: Washington DC, USA, 2000. [Google Scholar]
- Van-Soest, P.J.; Robertson, H.B.; Lewis, B.A. Methods of dietary fiber, NDF and nonstarch polysaccharides determination in relation to animal material. J. Dairy Sci. 1991, 74, 3583–3591. [Google Scholar] [CrossRef]
- Licitra, G.; Hernandez, T.M.; Van Soest, V.J. Standardization of procedures for nitrogen fractionation of ruminant feeds. Anim. Feed. Sci. Technol. 1996, 57, 347–358. [Google Scholar] [CrossRef]
- National Research Council (NRC). Nutrient Requirements of Dairy Cattle, 7th ed.; National Academic Press: Washington, DC, USA, 2001. [Google Scholar]
- Khan, N.A.; Booker, H.; Yu, P. Molecular structures and metabolic characteristics of protein in brown and yellow flaxseed with altered nutrient traits. J. Agric. Food Chem. 2014, 62, 6556–6564. [Google Scholar] [CrossRef]
- Tilley, J.M.A.; Terry, R.A. A two stage technique for in vitro digestion of forage crops. J. Br. Grassl. 1963, 18, 104–111. [Google Scholar] [CrossRef]
- Van Amburgh, M.E.; Collao-Saenz, E.A.; Higgs, R.J.; Ross, D.A.; Recktenwald, E.B. The Cornell Net Carbohydrate and Protein System: Updates to the model and evaluation of version 6.5. J. Dairy Sci. 2015, 98, 6361–6380. [Google Scholar] [CrossRef]
- Cone, J.W.; Van Gelder, A.H.; Van Schooten, H.A.; Groten, J.A.M. Effects of forage maize type and maturity stage on in vitro rumen fermentation characteristics. NJAS—Wagening. J. Life Sci. 2008, 55, 139–154. [Google Scholar] [CrossRef]
- Ali, M.; Cone, J.W.; Khan, N.A.; Hendriks, W.H.; Struik, P.C. Effect of temperature and duration of ensiling on in vitro degradation of maize silages in rumen fluid. J. Anim. Physiol. Anim. Nutr. 2015, 92, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Zamir, M.S.I.; Ahmad, A.H.; Javeed, H.M.R.; Latif, T. Growth and yield behavior of two maize hybrids (Zea mays L.) towards different plant spacing. Cercet. Agron. Mold. 2011, 44, 33–40. [Google Scholar]
- Pradeep, M.G.; Patil, R.H. Evaluation of maize hybrids for their maturity groups and their effect on yield. Int. J. Pure Appl. Biosci. 2018, 6, 382–387. [Google Scholar]
- Santiago López, U.; Rosales Nieto, C.A.; Santiago López, E.; Santiago López, N.; Preciado Rangel, P.; Palmo Gil, A.; Real, D. Yield of forage, grain, and biomass in eight hybrids of maize with different sowing dates and environmental conditions. Rev. Mex. Cienc. Pecu. 2018, 9, 86–104. [Google Scholar] [CrossRef] [Green Version]
- Millner, J.P.; Vill Aver, R.; Hardacre, A.K. The yield and nutritive value of maize genotypes grown for silage, New Zealand. J. Agric. Res. 2010, 48, 101–108. [Google Scholar] [CrossRef]
- Khan, N.A.; Tewoldebrahn, T.A.; Cone, J.W.; Zom, R.L.G.; Hendriks, W.H. Effect of silage maize maturity and concentrate type on milk fatty acid composition of dairy cows. J. Dairy Sci. 2012, 95, 1472–1483. [Google Scholar] [CrossRef] [Green Version]
- Guyader, J.; Baron, S.V.; Beauchemin, K.A. Corn forage yield and quality for silage in short growing season areas of the Canadian Prairies. Agronomy 2018, 8, 164. [Google Scholar] [CrossRef] [Green Version]
- Khan, N.A.; Cone, J.W.; Pellikaan, W.F.; Khan, M.A.; Struik, P.C.; Hendriks, W.H. Changes in fatty acid content and composition in silage maize during grain filling. J. Sci. Food Agric. 2011, 91, 1041–1049. [Google Scholar] [CrossRef]
- Johnson, L.M.; Harrison, J.H.; Davidson, D.; Robutti, J.L.; Swift, M.; Mahanna, W.C.; Shinners, K. Corn silage management. I. Effects of hybrid, maturity, and mechanical processing on chemical and physical characteristic. J. Dairy Sci. 2002, 85, 833–853. [Google Scholar] [CrossRef]
- Schwab, E.C.; Shaver, R.D.; Lauer, J.G.; Coors, J.G. Estimating silage energy value and milk yield to rank corn hybrids. Anim. Feed Sci. Technol. 2003, 109, 1–18. [Google Scholar] [CrossRef]
- Kung, J.L.; Lim, J.M.; Hudson, D.J.; Smith, J.M.; Joerger, R.D. Chemical composition and nutritive value of corn silage harvested in the northeastern United States after Tropical Storm Irene. J. Dairy Sci. 2015, 98, 2055–2062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loučka, R.; Tyrolová, Y.; Jančík, F.; Kubelková, P.; Homolka, P.; Jambor, V. Variation for In Vivo Digestibility in Two Maize Hybrid Silages. Czech J. Anim. Sci. 2018, 63, 17–23. [Google Scholar] [CrossRef]
- Higgs, R.J.; Chase, L.E.; Ross, D.A.; Van Amburgh, M.E. Updating the Cornell Net Carbohydrate and Protein System feed library and analyzing model sensitivity to feed inputs. J. Dairy Sci. 2015, 98, 6340–6360. [Google Scholar] [CrossRef] [Green Version]
- Khan, N.A.; Habib, G.; Ullah, G. Chemical composition, rumen degradability, protein utilization and lactation response to selected tree leaves as substitute of cottonseed cake in the diet of dairy goats. Anim. Feed Sci. Technol. 2009, 154, 160–168. [Google Scholar] [CrossRef]
- Habib, G.; Khan, N.A.; Sultan, A.; Ali, M. Nutritive value of common tree foliages for livestock in the semi-arid and arid rangelands of Northern Pakistan. Livest. Sci. 2016, 184, 64–70. [Google Scholar] [CrossRef]
- Nazli, M.; Halim, R.A.; Abdullah, A.M.; Hussin, G.; Samsudin, A.A. Potential of four corn varieties at different harvest stages for silage production in Malaysia. Asian Australas. J. Anim. Sci. 2019, 32, 224–232. [Google Scholar] [CrossRef] [PubMed]
- Shao, T.; Zhang, Z.X.; Shimojo, M.; Wang, T.; Masuda, Y. Comparison of fermentation characteristics of Italian ryegrass (Lolium multiflorum Lam.) and guineagrass (Panicum maximum Jacq.) during the early stage of ensiling. Asian Australas. J. Anim. 2005, 18, 1727–1734. [Google Scholar] [CrossRef]
- Anders, B.; Karin, J.; Katrin, S.M.; Johan, S. Metabolite profiles of lactic acid bacteria in grass silage. Appl. Environ. Microbiol. 2007, 73, 5547–5552. [Google Scholar]
- NRC (National Research Council). Nutrient Requirements of Beef Cattle, 8th ed.; National Academy of Sciences: Washington, DC, USA, 2016. [Google Scholar]
- Nennich, T.D.; Linn, J.G.; Johnson, D.G.; Endres, M.I.; Jung, H.G. Comparison of feeding corn silages from leafy or conventional corn hybrids to lactating dairy cows. J. Dairy Sci. 2003, 86, 2932–2939. [Google Scholar] [CrossRef] [Green Version]
- Ali, M.; Cone, J.W.; Khan, N.A.; Hendriks, W.H. Variation between individual cows in in situ rumen degradation characteristics of maize and grass silages. NJAS—Wagening. J. Life Sci. 2016, 78, 167–173. [Google Scholar] [CrossRef]


{kind=link}
| Genotypes | Growth Characteristics | Phonological Parameters | |||
|---|---|---|---|---|---|
| DTF | DTS | Height (m) | LS/P | C/P | |
| P1429 | 59 bc | 64 b | 2.14 d | 3.90 bc | 1.07 d |
| Azam | 57 d | 61 d | 1.83 e | 4.50 a | 1.14 cd |
| DK9108 | 59 c | 62 c | 2.45 a | 3.80 c | 1.30 bc |
| QPM200 | 61 b | 64 ab | 2.22 c | 3.80 c | 1.33 b |
| QPM300 | 60 ab | 64 ab | 2.37 b | 3.30 d | 1.58 a |
| P1543 | 62 a | 65 a | 2.15 d | 4.10 b | 1.25 c |
| SEM | 0.40 | 0.3 | 0.11 | 0.18 | 0.06 |
| Significance | *** | *** | *** | *** | *** |
| Genotypes | DM (g/kg) | Yield (tons/ha) | |||
|---|---|---|---|---|---|
| DM | CP | Starch | NDF | ||
| P1429 | 326 | 14.9 b | 1.02 c | 5.25 d | 6.60 d |
| Azam | 339 | 13.0 c | 0.83 d | 4.16 e | 6.12 e |
| DK9108 | 329 | 16.0 b | 1.06 c | 5.76 b | 7.29 b |
| QPM200 | 323 | 16.3 ab | 1.20 b | 5.41 c | 7.04 c |
| QPM300 | 320 | 17.9 a | 1.24 a | 6.67 a | 7.42 a |
| P1543 | 335 | 15.9 b | 1.06 c | 5.54 bc | 7.31 b |
| SEM | 0.31 | 0.34 | 0.03 | 0.11 | 0.37 |
| Significance | NS | *** | *** | *** | *** |
| Genotypes | g/100 g Dry Matter | g/100 g CP | IVDMD (g/100 g) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| CP | CHO | Starch | ADF | NFC | NDF | SCP | NDICP | ADICP | ||
| P1429 | 6.30 d | 84.2 | 35.3 b | 25.7 a | 42.0 b | 44.3 bc | 44.1 a | 13.2 abc | 4.70 c | 65.7 b |
| Azam | 6.04 e | 84.7 | 32.0 d | 27.2 a | 40.3 c | 47.1 a | 42.1 bc | 11.5 c | 5.70 b | 61.5 c |
| DK9108 | 6.60 c | 84.9 | 36.0 a | 26.6 a | 41.9 bc | 45.6 ab | 43.8 a | 13.3 ab | 5.65 b | 65.8 b |
| QPM200 | 7.51 b | 83.8 | 33.6 c | 26.1 a | 42.6 ab | 43.2 c | 43.6 a | 12.9 abc | 6.00 b | 65.8 b |
| QPM300 | 8.05 a | 84.0 | 37.3 a | 23.6 b | 43.8 a | 41.5 d | 41.1 c | 12.3 bc | 7.01 a | 67.9 a |
| P1543 | 6.60 c | 85.3 | 34.9 bc | 25.6 a | 42.3 ab | 44.9 bc | 43.0 ab | 20.1 c | 5.42 b | 66.7 ab |
| SEM | 0.11 | 0.33 | 0.36 | 0.40 | 0.42 | 0.37 | 0.67 | 0.44 | 0.22 | 0.47 |
| Significance | *** | NS | *** | *** | *** | *** | *** | *** | *** | *** |
| Genotypes | Rumen Degradation Characteristics | EDstarch | UDstarch | |||
|---|---|---|---|---|---|---|
| W | D | U | Kd | |||
| P1429 | 33.6 cd | 53.0 ab | 5.71 a | 19.4 a | 88.0 b | 12.0 b |
| Azam | 37.9 c | 50.0 b | 2.55 c | 09.7 c | 85.7 b | 14.6 ab |
| DK9108 | 40.4 b | 37.8 c | 2.80 c | 14.5 b | 94.2 a | 05.81 d |
| QPM200 | 31.0 d | 55.4 a | 5.85 a | 19.1 a | 83.8 c | 16.2 a |
| QPM300 | 46.2 a | 37. 6 c | 4.66 b | 22.1 a | 95. 8 a | 09.22 c |
| P1543 | 41.6 b | 21.5 d | 1.28 d | 11.2 c | 94.4 a | 02.60 e |
| SEM | 1.12 | 0.56 | 0.89 | 0.57 | 2.22 | 1.60 |
| p value | ** | *** | * | *** | *** | *** |
| Genotypes | pH | Lactic Acid% | Acetic Acid% | Propionic Acid | NH3-N (g/100 g N) |
|---|---|---|---|---|---|
| P1429 | 4.00 ab | 4.72 b | 2.55 c | 0.90 c | 9.20 |
| Azam | 4.20 a | 3.95 d | 3.03 a | 1.18 a | 10.1 |
| DK9108 | 3.90 b | 4.71 bc | 2.85 b | 1.02 b | 9.60 |
| QPM200 | 3.90 b | 4.17 c | 2.70 bc | 0.98 bc | 9.40 |
| QPM300 | 3.60 c | 5.19 a | 2.24 d | 0.80 d | 9.10 |
| P1543 | 3.60 c | 4.95 ab | 2.69 bc | 0.90 c | 9.60 |
| SEM | 0.31 | 0.40 | 0.11 | 0.10 | 0.43 |
| Significance | ** | ** | * | * | NS |
| Genotypes | Carbohydrate Subfraction | ||||
|---|---|---|---|---|---|
| CA4 | CB1 | CB2 | CB3 | CC | |
| P1429 | 5.32 b | 35.3 b | 10.77 ab | 40.8 b | 3.66 |
| Azam | 5.77 ab | 32.0 d | 11.35 a | 43.3 a | 3.80 |
| DK9108 | 5.16 b | 36.0 ab | 9.84 ab | 42.0 ab | 3.64 |
| QPM200 | 5.40 b | 33.6 c | 11.48 a | 39.62 c | 3.59 |
| QPM300 | 6.79 a | 37.3 a | 8.80 b | 38.05 d | 3.50 |
| P1543 | 6.11 ab | 34.9 bc | 11.26 a | 41.3 b | 3.46 |
| SEM | 0.098 | 0.36 | 0.12 | 0.37 | 0.09 |
| Significance | ** | *** | ** | *** | NS |
| tdNDF | tdNFC | tdCP | tdFA | TDN | DE | ME | |
|---|---|---|---|---|---|---|---|
| P1429 | 24.4 b | 41.2 b | 6.59 c | 2.22 | 70.2 ab | 3.03 ab | 2.41 bc |
| Azam | 25.8 a | 39.5 c | 6.43 c | 2.08 | 68.1 c | 2.80 b | 2.28 c |
| DK9108 | 25.0 ab | 40.1 bc | 6.65 c | 2.22 | 69.8 ab | 3.02 | 2.41 bc |
| QPM200 | 22.9 cd | 41.7 ab | 7.50 b | 1.98 | 69.7 ab | 3.02 b | 2.40 bc |
| QPM300 | 21.8 d | 42.9 a | 8.04 a | 2.16 | 70.6 a | 3.07 a | 2.44 a |
| P1543 | 24.0 bc | 41.5 ab | 6.89 c | 2.12 | 70.3 ab | 3.04 ab | 2.42 ab |
| SEM | 0.32 | 0.40 | 0.11 | 0.10 | 0.24 | 0.01 | 0.99 |
| Significance | *** | *** | *** | NS | * | ** | ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, M.; Ma, Y.; Khan, N.; Khan, M.Z.; Akbar, A.; Khan, R.U.; Kamran, M.; Khan, N.A. Effect of Spring Maize Genotypes on Fermentation and Nutritional Value of Whole Plant Maize Silage in Northern Pakistan. Fermentation 2022, 8, 587. https://doi.org/10.3390/fermentation8110587
Jiang M, Ma Y, Khan N, Khan MZ, Akbar A, Khan RU, Kamran M, Khan NA. Effect of Spring Maize Genotypes on Fermentation and Nutritional Value of Whole Plant Maize Silage in Northern Pakistan. Fermentation. 2022; 8(11):587. https://doi.org/10.3390/fermentation8110587
Chicago/Turabian StyleJiang, Mingming, Yulin Ma, Nadar Khan, Muhammad Zahoor Khan, Ali Akbar, Rifat Ullah Khan, Muhammad Kamran, and Nazir Ahmad Khan. 2022. "Effect of Spring Maize Genotypes on Fermentation and Nutritional Value of Whole Plant Maize Silage in Northern Pakistan" Fermentation 8, no. 11: 587. https://doi.org/10.3390/fermentation8110587