
Optimized Operating Conditions for a Biological Treatment Process of Industrial Residual Process Brine Using a Halophilic Mixed Culture


Abstract
:1. Introduction
2. Materials and Methods
2.1. Strain and Medium
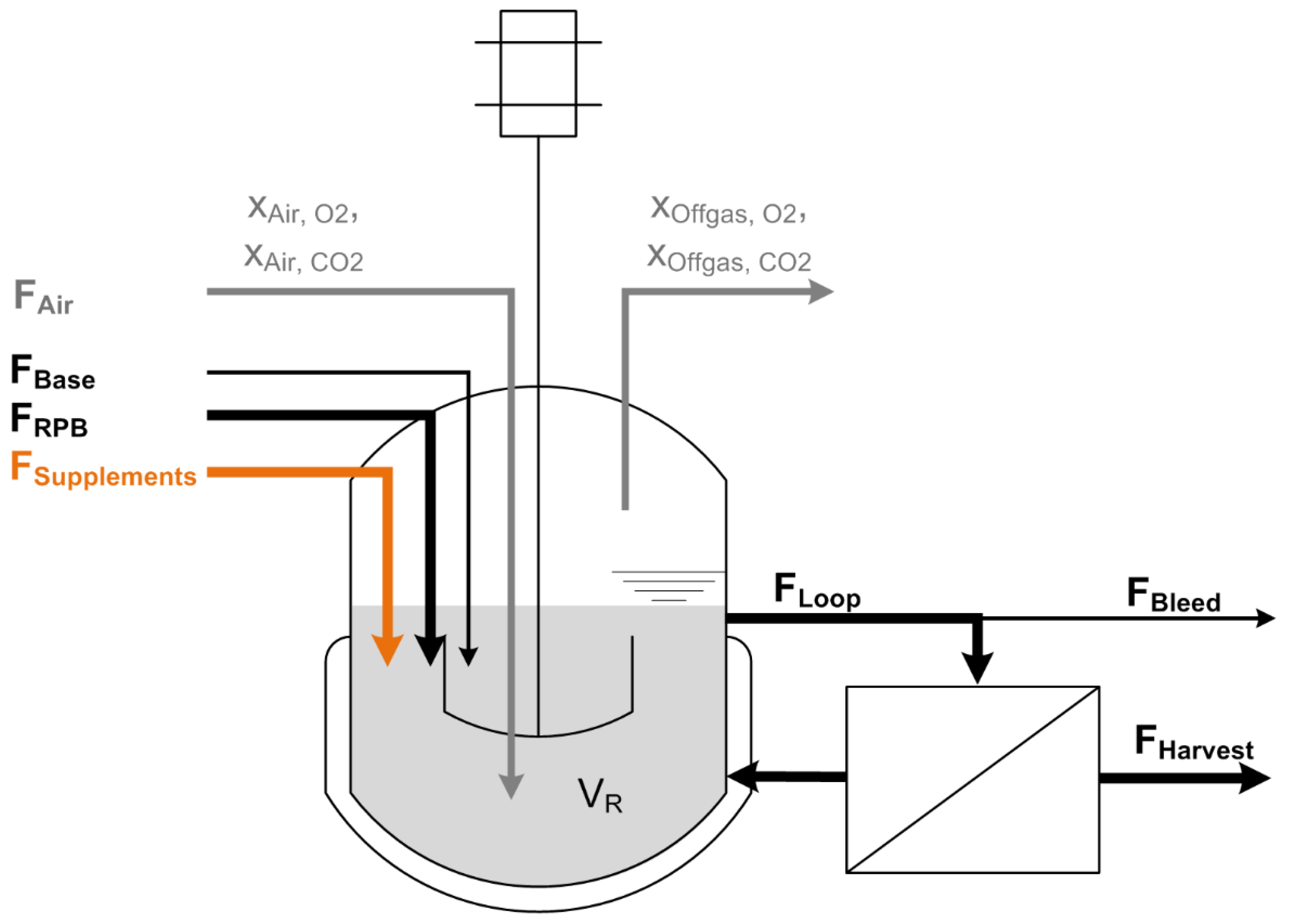
2.2. Bioreactor Setup—Continuous Stirred-Tank Reactor
2.3. Calculations
2.3.1. Steady-State Cultivation
2.3.2. Multiple Linear Regression
2.4. Analytical Procedures
2.4.1. HPLC Analysis
2.4.2. Media Composition Analysis
2.4.3. Determination of the Chloride Ion Concentration
3. Results and Discussion
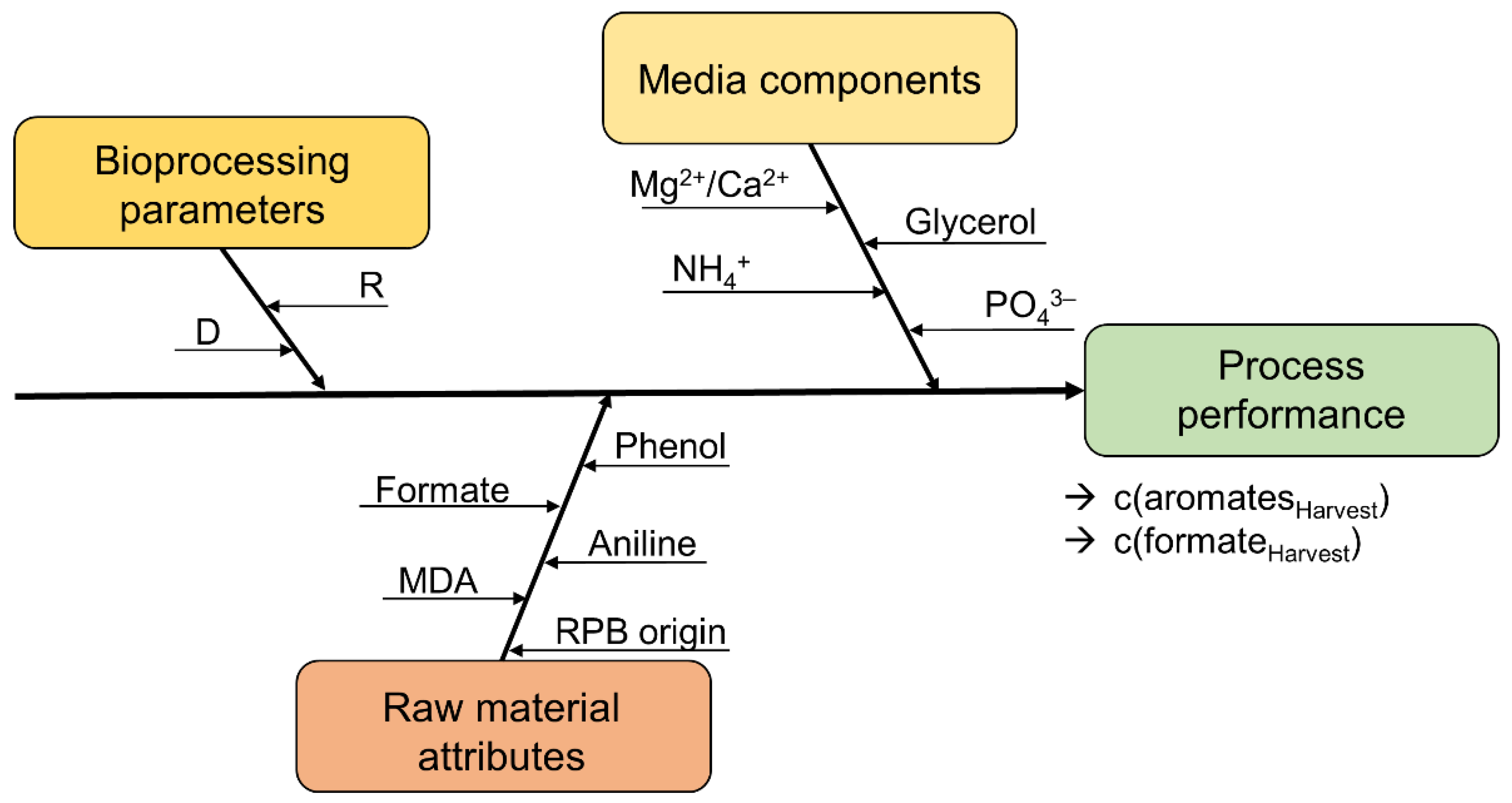
3.1. Definition of Process Performance Variables and Critical Process Parameters
- the residual aromatic concentration in the harvest c(aromatesHarvest) [mg L−1];
- the residual formate concentration in the harvest c(formateHarvest) [g L−1];
- Bioprocess parameters (dilution rate, retention rate, and the biomass concentration (indicated by the optical density));
- Media components (glycerol, ammonium, phosphate, magnesium, and calcium feed concentrations);
- Raw material attributes (RPB origin, formate, aniline, phenol, and 4,4′-MDA feed concentrations).
3.2. Influence of Critical Raw Material Attributes on Process Performance
3.2.1. RPB Origin
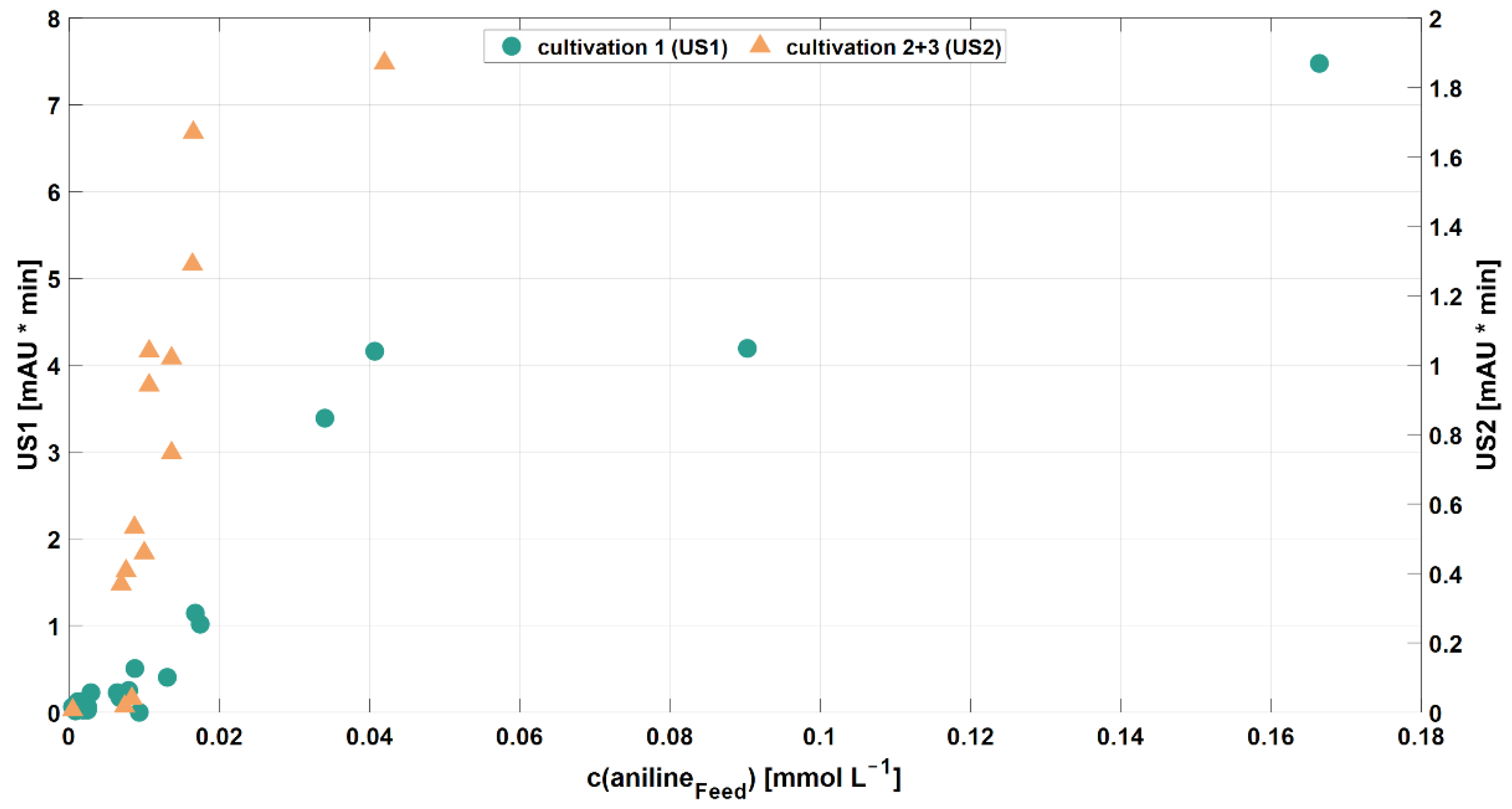
3.2.2. Aromatic Compounds
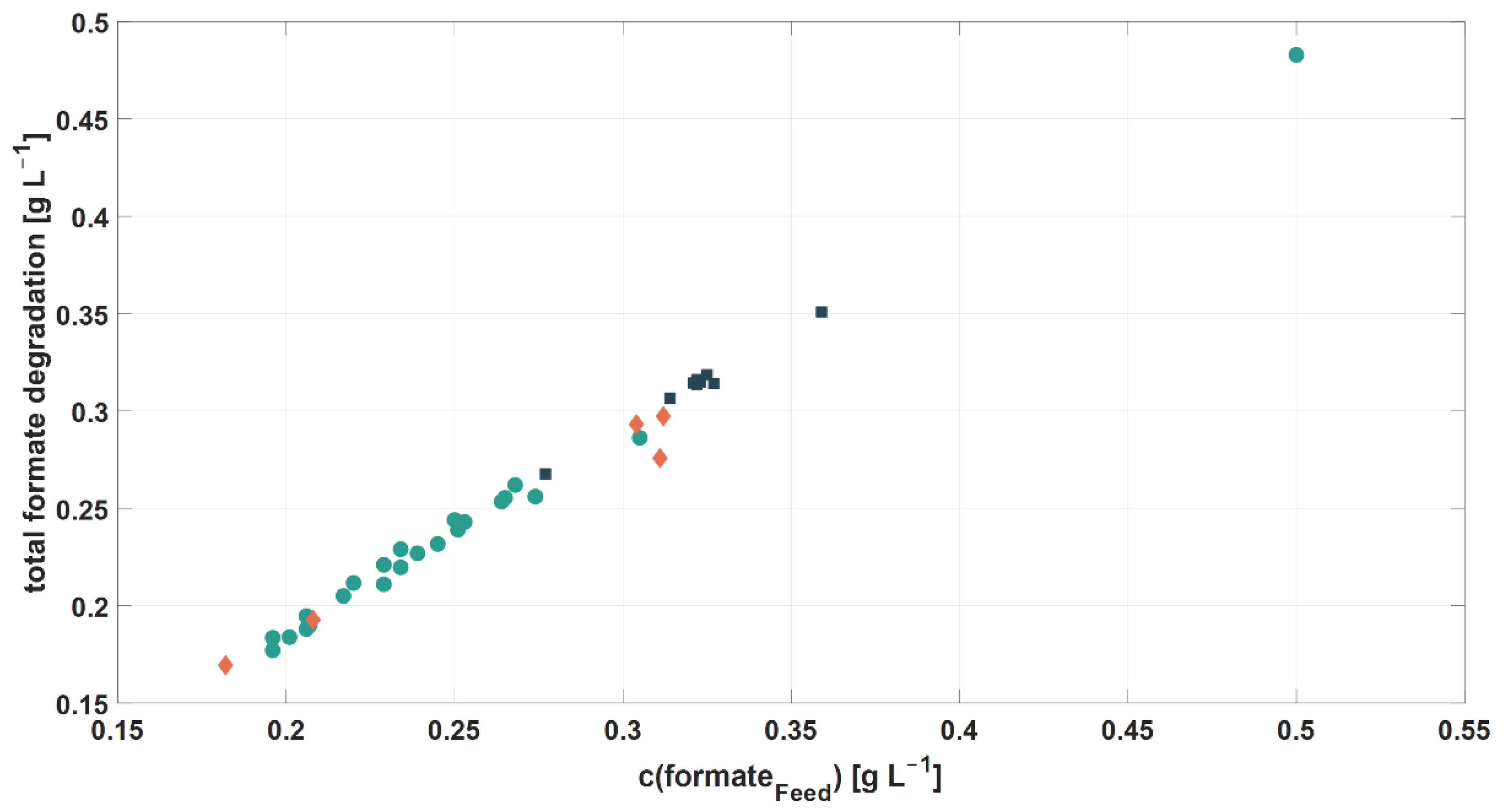
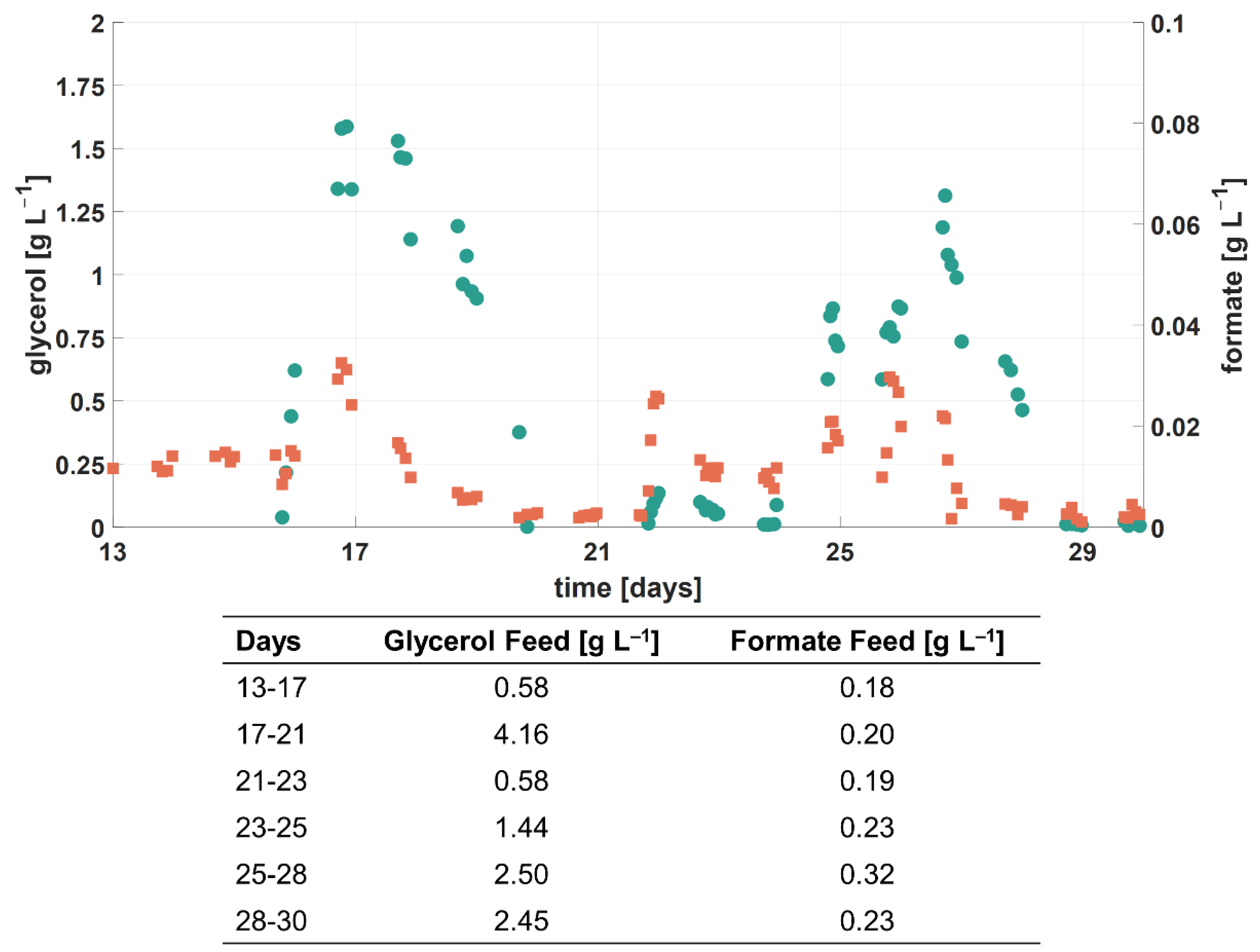
3.2.3. Formate
3.3. Influence of Media Components on Process Performance
3.3.1. Ammonium and Phosphate
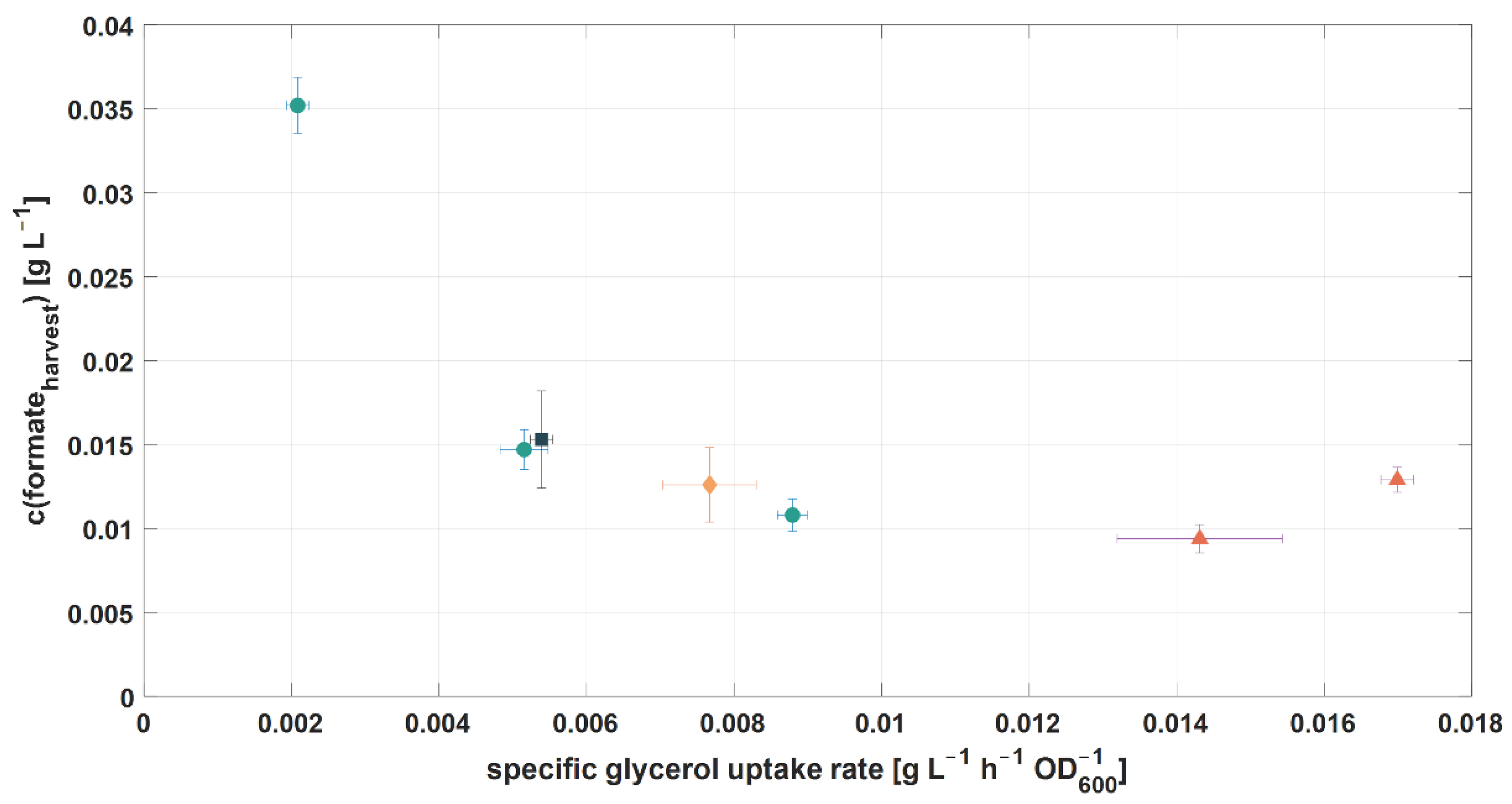
3.3.2. Glycerol
3.4. Defining Optimized Operating Conditions
4. Conclusions
5. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Crook, J.; Mousavi, A. The chlor-alkali process: A review of history and pollution. Environ. Forensics 2016, 17, 211–217. [Google Scholar] [CrossRef]
- Lakshmanan, S.; Murugesan, T. The chlor-alkali process: Work in progress. Clean Technol. Environ. Policy 2014, 16, 225–234. [Google Scholar] [CrossRef]
- Bergner, D. Membrane cells for chlor-alkali electrolysis. J. Appl. Electrochem. 1982, 12, 631–644. [Google Scholar] [CrossRef]
- Garcia-Herrero, I.; Margallo, M.; Onandía, R.; Aldaco, R.; Irabien, A. Environmental challenges of the chlor-alkali production: Seeking answers from a life cycle approach. Sci. Total Environ. 2017, 580, 147–157. [Google Scholar] [CrossRef] [Green Version]
- Reig, M.; Casas, S.; Aladjem, C.; Valderrama, C.; Gibert, O.; Valero, F.; Centeno, C.M.; Larrotcha, E.; Cortina, J.L. Concentration of NaCl from seawater reverse osmosis brines for the chlor-alkali industry by electrodialysis. Desalination 2014, 342, 107–117. [Google Scholar] [CrossRef]
- Muddemann, T.; Bulan, A.; Sievers, M.; Kunz, U. Avoidance of Chlorine Formation during Electrolysis at Boron-Doped Diamond Anodes in Highly Sodium Chloride Containing and Organic-Polluted Wastewater. J. Electrochem. Soc. 2018, 165, J3281–J3287. [Google Scholar] [CrossRef]
- Brinkmann, T.; Giner Santonja, G.; Schorcht, F.; Roudier, S.; Delgado Sancho, L. Best Available Techniques (BAT) Reference Document for the Production of Chlor-Alkali; Publications Office of the European Union: Luxembourg, 2014. [Google Scholar]
- Keating, J.T.; Behling, K.-J. Brine, Impurities, and Membrane Chlor-Alkali Cell Performance, in Modern Chlor-Alkali Technology; Prout, N.M., Moorhouse, J.S., Eds.; Springer: Dordrecht, The Netherlands, 1990; Volume 4, pp. 125–139. [Google Scholar]
- Silva, J.M.; Soloveichik, G.L.; Novak, D. Effects of Organic Impurities on Chloralkali Membrane Electrolyzer Performance. Ind. Eng. Chem. Res. 2008, 48, 983–987. [Google Scholar] [CrossRef]
- Thiel, G.P.; Kumar, A.; Gómez-González, A.; Lienhard, V.J.H. Utilization of Desalination Brine for Sodium Hydroxide Production: Technologies, Engineering Principles, Recovery Limits, and Future Directions. ACS Sustain. Chem. Eng. 2017, 5, 11147–11162. [Google Scholar] [CrossRef]
- Madaeni, S.; Kazemi, V. Treatment of saturated brine in chlor-alkali process using membranes. Sep. Purif. Technol. 2008, 61, 68–74. [Google Scholar] [CrossRef]
- Zhuo, Y.; Sheng, M.; Liang, X.; Cao, G. Treatment of high salinity wastewater using CWPO process for reuse. J. Adv. Oxid. Technol. 2017, 20. [Google Scholar] [CrossRef] [Green Version]
- Mainka, T.; Mahler, N.; Herwig, C.; Pflügl, S. Soft Sensor-Based Monitoring and Efficient Control Strategies of Biomass Concentration for Continuous Cultures of Haloferax mediterranei and Their Application to an Industrial Production Chain. Microorganisms 2019, 7, 648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castillo-Carvajal, L.C.; Sanz-Martín, J.L.; Barragán-Huerta, B.E. Biodegradation of organic pollutants in saline wastewater by halophilic microorganisms: A review. Environ. Sci. Pollut. Res. Int. 2014, 21, 9578–9588. [Google Scholar] [CrossRef] [PubMed]
- Fathepure, B.Z. Recent studies in microbial degradation of petroleum hydrocarbons in hypersaline environments. Front. Microbiol. 2014, 5, 173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mainka, T.; Herwig, C.; Pflügl, S. Reducing Organic Load From Industrial Residual Process Brine with a Novel Halophilic Mixed Culture: Scale-Up and Long-Term Piloting of an Integrated Bioprocess. Front. Bioeng. Biotechnol. 2022, 10, 896576. [Google Scholar]
- Tan, X.; Acquah, I.; Liu, H.; Li, W.; Tan, S. A critical review on saline wastewater treatment by membrane bioreactor (MBR) from a microbial perspective. Chemosphere 2019, 220, 1150–1162. [Google Scholar] [CrossRef]
- Al-Shaikh, A.J.; Jamal, M.T. Bioaugmentation of halophilic consortia for the degradation of petroleum hydrocarbons and petroleum wastewater treatment. Int. J. Adv. Res. Biol. Sci. 2020, 7, 97–112. [Google Scholar]
- Jamal, M.T.; Pugazhendi, A. Degradation of petroleum hydrocarbons and treatment of refinery wastewater under saline condition by a halophilic bacterial consortium enriched from marine environment (Red Sea), Jeddah, Saudi Arabia. 3 Biotech 2018, 8, 276. [Google Scholar] [CrossRef]
- Kreye, O.; Mutlu, H.; Meier, M.A.R. Sustainable routes to polyurethane precursors. Green Chem. 2013, 15, 1431–1455. [Google Scholar] [CrossRef]
- Boros, R.Z.; Koós, T.; Wafaa, C.; Nehéz, K.; Farkas, L.; Viskolcz, B.; Szőri, M. A theoretical study on the phosgenation of methylene diphenyl diamine (MDA). Chem. Phys. Lett. 2018, 706, 568–576. [Google Scholar] [CrossRef]
- Schupp, T.; Allmendinger, H.; Boegi, C.; Bossuyt, B.T.A.; Hidding, B.; Shen, S.; Tury, B.; West, R.J. The Environmental Behavior of Methylene-4, 4′-dianiline. In Reviews of Environmental Contamination and Toxicology; Springer: Cham, Germany, 2018; Volume 246, pp. 91–132. [Google Scholar]
- Bulan, A.; Weber, R.; Muddemann, T. Process for the Electrochemical Purification of Chloride-Containing Process Solutions. US20190177186A1, 13 June 2019. [Google Scholar]
- Merenov, A.S.; Jewell, D.W.; Gillis, P.A.; Jansma, G.I.; Breed, A.W.; Anderson, J.J.; Reed, D.J. Process for the Production of Methylene Diphenyl Diisocyanate Isomer Mixtures with Specific Isomer Distributions and New Products Derived. US9090540B2, 28 July 2015. [Google Scholar]
- Thornton, P.G. Process for the Preparation of 4, 4′-Methylenedianiline. US3367969A, 6 February 1968. [Google Scholar]
- Eberly, L.E. Multiple Linear Regression; Topics in Biostatistics; Humana Press: Totowa, NJ, USA, 2007; pp. 165–187. [Google Scholar]
- Seber, G.A.; Lee, A.J. Linear Regression Analysis; John Wiley & Sons: Hoboken, NJ, USA, 2012. [Google Scholar]
- Erian, A.M.; Gibisch, M.; Pflügl, S.; Engineered, E. coli W enables efficient 2,3-butanediol production from glucose and sugar beet molasses using defined minimal medium as economic basis. Microb. Cell Fact. 2018, 17, 190. [Google Scholar] [CrossRef]
- Kolthoff, I.M.; Lauer, W.M.; Sunde, C.J. The use of dichlorofluorescein as an adsorption indicator for the argentometric titration of chlorides. J. Am. Chem. Soc. 1929, 51, 3273–3277. [Google Scholar] [CrossRef]
- Alva, V.A.; Peyton, B.M. Phenol and Catechol Biodegradation by the Haloalkaliphile Halomonas campisalis: Influence of pH and Salinity. Environ. Sci. Technol. 2003, 37, 4397–4402. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Meng, F.; Duan, W.; Lin, Y.; Zheng, Y. Biodegradation of phenol in saline or hypersaline environments by bacteria: A review. Ecotoxicol. Environ. Saf. 2019, 184, 109658. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, G.; Boll, M.; Heider, J. Microbial degradation of aromatic compounds—From one strategy to four. Nat. Rev. Genet. 2011, 9, 803–816. [Google Scholar] [CrossRef]
- Mainka, T.; Weirathmüller, D.; Herwig, C.; Pflügl, S. Potential applications of halophilic microorganisms for biological treatment of industrial process brines contaminated with aromatics. J. Ind. Microbiol. Biotechnol. 2021, 48, kuab015. [Google Scholar] [CrossRef]
- Niks, D.; Hille, R. Molybdenum-and tungsten-containing formate dehydrogenases and formylmethanofuran dehydrogenases: Structure, mechanism, and cofactor insertion. Protein Sci. 2019, 28, 111–122. [Google Scholar] [CrossRef] [Green Version]
- Alpdağtaş, S.; Turunen, O.; Valjakka, J.; Binay, B. The challenges of using NAD+-dependent formate dehydrogenases for CO2 conversion. Crit. Rev. Biotechnol. 2021, 42, 953–972. [Google Scholar] [CrossRef]
- Tishkov, V.I.; Popov, V. Protein engineering of formate dehydrogenase. Biomol. Eng. 2006, 23, 89–110. [Google Scholar] [CrossRef]
- Maia, L.B.; Moura, I.; Moura, J.J.G. Molybdenum and tungsten-containing formate dehydrogenases: Aiming to inspire a catalyst for carbon dioxide utilization. Inorg. Chim. Acta 2016, 455, 350–363. [Google Scholar] [CrossRef]
- Yu, X.; Niks, D.; Mulchandani, A.; Hille, R. Efficient reduction of CO2 by the molybdenum-containing formate dehydrogenase from Cupriavidus necator (Ralstonia eutropha). J. Biol. Chem. 2017, 292, 16872–16879. [Google Scholar] [CrossRef] [Green Version]
- Mills, M.M.; Moore, C.M.; Langlois, R.; Milne, A.; Achterberg, E.; Nachtigall, K.; Lochte, K.; Geider, R.J.; La, R.J. Nitrogen and phosphorus co-limitation of bacterial productivity and growth in the oligotrophic subtropical North Atlantic. Limnol. Oceanogr. 2008, 53, 824–834. [Google Scholar] [CrossRef] [Green Version]
- Bracken, M.E.S.; Hillebrand, H.; Borer, E.T.; Seabloom, E.W.; Cebrian, J.; Cleland, E.E.; Elser, J.J.; Gruner, D.S.; Harpole, W.S.; Ngai, J.T.; et al. Signatures of nutrient limitation and co-limitation: Responses of autotroph internal nutrient concentrations to nitrogen and phosphorus additions. Oikos 2014, 124, 113–121. [Google Scholar] [CrossRef]
- Maldonado, M.; Boyd, P.; Harrison, P.J.; Price, N. Co-limitation of phytoplankton growth by light and Fe during winter in the NE subarctic Pacific Ocean. Deep Sea Res. Part II Top. Stud. Oceanogr. 1999, 46, 2475–2485. [Google Scholar] [CrossRef]
- Morel, F.M.M.; Reinfelder, J.; Roberts, S.B.; Chamberlain, C.P.; Lee, J.G.; Yee, D. Zinc and carbon co-limitation of marine phytoplankton. Nature 1994, 369, 740–742. [Google Scholar] [CrossRef]
- Price, N.M.; Morel, F.M.M. Colimitation of phytoplankton growth by nickel and nitrogen. Limnol. Oceanogr. 1991, 36, 1071–1077. [Google Scholar] [CrossRef]
- Sunda, W.G.; Huntsman, S.A. Interrelated influence of iron, light and cell size on marine phytoplankton growth. Nature 1997, 390, 389–392. [Google Scholar] [CrossRef]
- Frank, S.E. Growth Characteristics of Acinetobacter Johnsonii 210A under Single and Dual Nutrient Limitation with Special Reference to Carbon, Nitrogen and Phosphorus; ETH Zurich: Zürich, Switzerland, 1999; p. 146. [Google Scholar]
- Egli, T. On multiple-nutrient-limited growth of microorganisms, with special reference to dual limitation by carbon and nitrogen substrates. Antonie van Leeuwenhoek 1991, 60, 225–234. [Google Scholar] [CrossRef]
- Egli, T.; Quayle, J.R. Influence of the Carbon: Nitrogen Ratio of the Growth Medium on the Cellular Composition and the Ability of the Methylotrophic Yeast Hansenula polymorpha to Utilize Mixed Carbon Sources. Microbiology 1986, 132, 1779–1788. [Google Scholar] [CrossRef] [Green Version]
- Egli, T.; Zinn, M. The concept of multiple-nutrient-limited growth of microorganisms and its application in biotechnological processes. Biotechnol. Adv. 2003, 22, 35–43. [Google Scholar] [CrossRef]
- Zinn, M.; Witholt, B.; Egli, T. Dual nutrient limited growth: Models, experimental observations, and applications. J. Biotechnol. 2004, 113, 263–279. [Google Scholar] [CrossRef]
- Poblete-Castro, I.; Escapa, I.F.; Jäger, C.; Puchalka, J.; Lam, C.M.C.; Schomburg, D.; Prieto, M.A.; Santos, V.A.P.M.D. The metabolic response of P. putida KT2442 producing high levels of polyhydroxyalkanoate under single- and multiple-nutrient-limited growth: Highlights from a multi-level omics approach. Microb. Cell Fact. 2012, 11, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karmann, S.; Panke, S.; Zinn, M. Fed-Batch Cultivations of Rhodospirillum rubrum Under Multiple Nutrient-Limited Growth Conditions on Syngas as a Novel Option to Produce Poly(3-Hydroxybutyrate) (PHB). Front. Bioeng. Biotechnol. 2019, 7, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavaillé, L.; Albuquerque, M.; Grousseau, E.; Lepeuple, A.-S.; Uribelarrea, J.-L.; Hernandez-Raquet, G.; Paul, E. Understanding of polyhydroxybutyrate production under carbon and phosphorus-limited growth conditions in non-axenic continuous culture. Bioresour. Technol. 2016, 201, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Luna, M.F.; Ochsner, A.M.; Amstutz, V.; von Blarer, D.; Sokolov, M.; Arosio, P.; Zinn, M. Modeling of Continuous PHA Production by a Hybrid Approach Based on First Principles and Machine Learning. Processes 2021, 9, 1560. [Google Scholar] [CrossRef]
- Rüegg, I.; Hafner, T.; Bucheli-Witschel, M.; Egli, T. Dynamics of Benzene and Toluene Degradation inPseudomonas putida F1 in the Presence of the Alternative Substrate Succinate. Eng. Life Sci. 2007, 7, 331–342. [Google Scholar] [CrossRef]
- Duetz, W.A.; Marqués, S.; de Jong, C.; Ramos, J.L.; van Andel, J.G. Inducibility of the TOL catabolic pathway in Pseudomonas putida (pWW0) growing on succinate in continuous culture: Evidence of carbon catabolite repression control. J. Bacteriol. 1994, 176, 2354–2361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahler, N.; Tschirren, S.; Pflügl, S.; Herwig, C. Optimized bioreactor setup for scale-up studies of extreme halophilic cultures. Biochem. Eng. J. 2018, 130, 39–46. [Google Scholar] [CrossRef]
- Marozava, S.; Röling, W.F.; Seifert, J.; Küffner, R.; von Bergen, M.; Meckenstock, R.U. Physiology of Geobacter metallireducens under excess and limitation of electron donors. Part II. Mimicking environmental conditions during cultivation in retentostats. Syst. Appl. Microbiol. 2014, 37, 287–295. [Google Scholar] [CrossRef] [Green Version]
- Marozava, S.; Röling, W.F.; Seifert, J.; Küffner, R.; von Bergen, M.; Meckenstock, R.U. Physiology of Geobacter metallireducens under excess and limitation of electron donors. Part I. Batch cultivation with excess of carbon sources. Syst. Appl. Microbiol. 2014, 37, 277–286. [Google Scholar] [CrossRef] [Green Version]
- Deowan, S.A.; Bouhadjar, S.I.; Hoinkis, J. 5-Membrane bioreactors for water treatment. In Advances in Membrane Technologies for Water Treatment; Basile, A., Cassano, A., Rastogi, N.K., Eds.; Woodhead Publishing: Oxford, UK, 2015; pp. 155–184. [Google Scholar]
- Ding, Y.; Guo, Z.; Ma, B.; Wang, F.; You, H.; Mei, J.; Hou, X.; Liang, Z.; Li, Z.; Jin, C. The Influence of Different Operation Conditions on the Treatment of Mariculture Wastewater by the Combined System of Anoxic Filter and Membrane Bioreactor. Membranes 2021, 11, 729. [Google Scholar] [CrossRef]
- Cao, S.M.; Fontoura, G.A.T.; Dezotti, M.; Bassin, J.P. Combined organic matter and nitrogen removal from a chemical industry wastewater in a two-stage MBBR system. Environ. Technol. 2016, 37, 96–107. [Google Scholar] [CrossRef] [PubMed]
- Lorantfy, B.; Ruschitzka, P.; Herwig, C. Investigation of physiological limits and conditions for robust bioprocessing of an extreme halophilic archaeon using external cell retention system. Biochem. Eng. J. 2014, 90, 140–148. [Google Scholar] [CrossRef]
- Doran, P.M. Bioprocess Engineering Principles; Elsevier: Amsterdam, The Netherlands, 1995. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Critical Process Parameter | Unit | Critical Process Parameter | Unit |
|---|---|---|---|
| Glycerol | g L−1 | Formate | mg L−1 |
| Ammonium (NH4+) | mmol L−1 | Aniline | mg L−1 |
| Phosphate (PO43−) | mmol L−1 | Phenol | mg L−1 |
| Magnesium (Mg2+) | g L−1 | 4,4′-MDA | mg L−1 |
| Calcium (Ca2+) | g L−1 | Optical Density OD600 | - |
| Dilution rate D | h−1 | Retention rate R | - |
| Parameter | Cultivation 1 | Cultivation 2 | Cultivation 3 |
|---|---|---|---|
| Reference | [16] | This study | This study |
| Feed system | Feed: RPB and glycerol | Feed: RPB and glycerol | Feed 1: RPB Feed 2: glycerol |
| Reactor volume | 16 L | 1 L | 1 L |
| Range of D | 0.06–0.20 h−1 | 0.09 h−1 | 0.10 h−1 |
| Range of R | 0.75–0.95 | 0.92–0.98 | 0.8–0.91 |
| RPB origins | 1 | 1 | 1, 2, 3 |
| Cultivation time [days] | >200 | 39 | 35 |
| Contaminant | RPB 1 [mg L−1] * | RPB 2 [mg L−1] | RPB 3 [mg L−1] |
|---|---|---|---|
| Aniline | 0.04–15.50 | 3.91 | 0.04 |
| Phenol | 1.37–7.71 | 15.00 | 2.99 |
| 4,4′-MDA | 0.02–0.74 | 0.21 | 0.06 |
| Exp. | c(NH4+Feed) [mM] | c(NH4+Harv.) [mM] | c(PO43−Feed) [mM] | c(PO43−Harv.) [mM] | YNH4+/Glycerol [molN molgly.−1] | YPO43-/Glycerol [molP molgly] | c(formateFeed) [mg L−1] | c(formateHarvest) [mg L−1] |
|---|---|---|---|---|---|---|---|---|
| 2.1 | 19.75 | 10.87 | 1.11 | 0.76 | 0.426 | 0.017 | 314 | 7.44 |
| 2.2 | 19.90 | 12.86 | 0.07 | 0 | 0.340 | 0.003 | 323 | 8.15 |
| 2.3 | 4.63 | 0 | 1.02 | 0.30 | 0.199 | 0.031 | 359 | 8.11 |
| 2.4 | 4.69 | 0 | 0.07 | 0 | 0.219 | 0.004 | 322 | 8.52 |
| 2.5 | 9.65 | 1.28 | 0.34 | 0 | 0.412 | 0.017 | 322 | 5.89 |
| 2.6 | 8.72 | 0.92 | 0.37 | 0 | 0.390 | 0.019 | 325 | 6.40 |
| 2.7 | 9.75 | 0.52 | 0.35 | 0 | 0.434 | 0.009 | 321 | 6.66 |
| Exp. | RPB | D [h−1] | R [–] | qS,gly [g L−1 h−1 OD600−1] | c(Gly.Feed) [g L−1] | c(NH4+Feed) [mmol L−1] | c(PO43−Feed) [mmol L−1] |
|---|---|---|---|---|---|---|---|
| 2.8 | 1 | 0.095 | 0.800 | 17.0 × 10−3 | 3.74 | 5.10 | 0.36 |
| 2.9 | 1 | 0.095 | 0.800 | 15.9 × 10−3 | 1.79 | 4.90 | 0.29 |
| 3.1 | 1 | 0.088 | 0.917 | 8.7 × 10−3 | 2.07 | 5.23 | 0.33 |
| 3.2 | 1 | 0.088 | 0.980 | 2.1 × 10−3 | 0.30 | 0.73 | 0.05 |
| 3.3 | 1 | 0.088 | 0.965 | 5.2 × 10−3 | 0.57 | 1.52 | 0.10 |
| 3.4 | 2 | 0.088 | 0.965 | 7.7 × 10−3 | 0.58 | 1.41 | 0.10 |
| 3.5 | 3 | 0.088 | 0.965 | 5.4 × 10−3 | 0.59 | 1.54 | 0.10 |
| Parameter | Control Parameter | Proposed Parameter Values |
|---|---|---|
| Specific glycerol uptake rate [g L−1 h−1 OD600−1] | - | 8.0–16.0 × 10−3 |
| Ammonium (NH4+) conc. feed | Ratio to glycerol [molN molgly−1] | 0.20–0.22 |
| Phosphate (PO43−) conc. feed | Ratio to glycerol [molP molgly−1] | 0.003–0.004 |
| Magnesium (Mg2+) conc. feed | Feed concentration for: MgSO4 × 7 H2O [g L−1] | 0.26 (equals 2.16 mmol L−1) |
| Calcium (Ca2+) conc. feed | Feed concentration for: CaCl2 × 2 H2O [g L−1] | 0.055 (equals 0.51 mmol L−1) |
| Dilution rate D [h−1] | - | 0.2 |
| Retention rate R [–] | - | 0.98 |
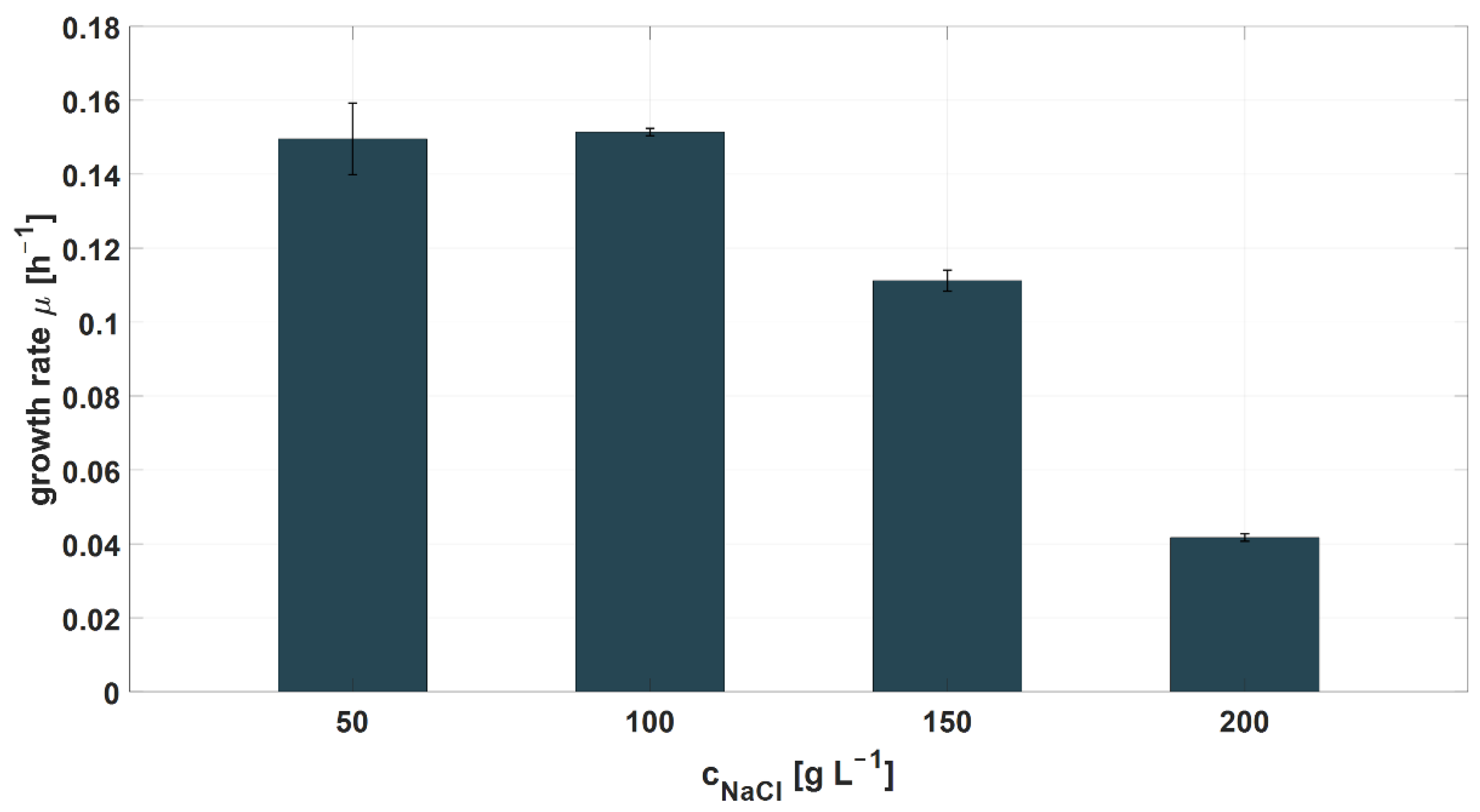
| NaCl [g L−1] | - | 50–100 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mainka, T.; Herwig, C.; Pflügl, S. Optimized Operating Conditions for a Biological Treatment Process of Industrial Residual Process Brine Using a Halophilic Mixed Culture. Fermentation 2022, 8, 246. https://doi.org/10.3390/fermentation8060246
Mainka T, Herwig C, Pflügl S. Optimized Operating Conditions for a Biological Treatment Process of Industrial Residual Process Brine Using a Halophilic Mixed Culture. Fermentation. 2022; 8(6):246. https://doi.org/10.3390/fermentation8060246
Chicago/Turabian StyleMainka, Thomas, Christoph Herwig, and Stefan Pflügl. 2022. "Optimized Operating Conditions for a Biological Treatment Process of Industrial Residual Process Brine Using a Halophilic Mixed Culture" Fermentation 8, no. 6: 246. https://doi.org/10.3390/fermentation8060246