
Mechanism and Effect of Amino Acids on Lactic Acid Production in Acidic Fermentation of Food Waste


1
School of Environment and Ecology, Jiangnan University, Wuxi 214122, China
2
Jiangsu Collaborative Innovation Center of Water Treatment Technology and Material, Suzhou University of Science and Technology, Suzhou 215011, China
*
Author to whom correspondence should be addressed.
Fermentation 2024, 10(4), 179; https://doi.org/10.3390/fermentation10040179
Submission received: 2 March 2024
/
Revised: 20 March 2024
/
Accepted: 22 March 2024
/
Published: 25 March 2024
(This article belongs to the Section Industrial Fermentation)
Abstract
:Amino acids, particularly the ones that cannot be synthesised during fermentation, are reportedly to be key nutrients for anaerobic fermentation processes, and some of the acids are also intermediate products of anaerobic fermentation of protein-rich waste. To date, particularly, there is a lack of research on the effects of some amino acids, such as cysteine, glycine, aspartic acid, and valine, on lactic production from the fermentation of food waste and also the mechanisms involved in the process. Thus, this study investigated the effects of the four different amino acids on lactic acid production during the acidic anaerobic fermentation of food waste. Firstly, batch experiments on synthetic food waste at different pHs (4.0, 5.0, and 6.0) were executed. The results harvested in this study showed that higher LA concentrations and yields could be obtained at pH 5.0 and pH 6.0, compared with those at pH 4.0. The yield of lactic acid was slightly lower at pH 5.0 than at pH 6.0. Furthermore, caustic consumption at pH 5.0 was much lower. Therefore, we conducted batch experiments with additions of different amino acids (cysteine, glycine, aspartic acid, and valine) under pH 5.0. The additions of the four different amino acids showed different or even opposite influences on LA production. Glycine and aspartic acids presented no noticeable effects on lactic acid production, but cysteine evidently enhanced the lactic acid yield of food waste by 13%. Cysteine addition increased α-glucosidase activity and hydrolysis rate and simultaneously enhanced the abundance of Lactobacillus at the acidification stage as well as lactate dehydrogenase, which also all favoured lactic acid production. However, the addition of valine evidently reduced lactic acid yield by 18%, and the results implied that valine seemingly inhibited the conversion of carbohydrate. In addition, the low abundance of Lactobacillus was observed in the tests with valine, which appeared to be detrimental to lactic acid production. Overall, this study provides a novel insight into the regulation of lactic acid production from anaerobic fermentation of food waste by adding amino acids under acidic fermentation conditions.
1. Introduction
It is estimated that about 2.5 × 109 tons of food waste (FW) will be generated globally by 2025 [1]. If the bulky waste is improperly treated, a public food and environmental security crisis could take place due to the potential pollution of the environment caused by the waste [2]. Currently, one of the most widely implemented resource recovery methods for food waste is anaerobic digestion for methane production, but the approach has two evident drawbacks: a long retention time and relatively poor stability [3]. Therefore, an effective food waste management system is impending to provide cost-effective resource and energy recovery, further achieving a circular bio-economy and fulfilling the goal of sustainability [4]. Thus far, several promising fermentation technologies have emerged and been employed to produce value-added products, such as biohydrogen, biohythane, lactic acid (LA), volatile fatty acids (VFAs), and medium chain carboxylic acids (MCCAs), from various food or industrial organic waste (Table 1) [5,6,7].
The anaerobic digestion (AD) process consists of four steps: hydrolysis, acidogenesis, acetogenesis, and methanogenesis. LA as an intermediate product is produced during acidogenesis [4], and has a broad range of applications such as a flavour enhancer, antacid, and preservative in the food, chemical, cosmetic, and pharmaceutical industries [8,9]. LA, especially, can be used in the plastics sector to produce biodegradable polylactic acid (PLA) plastics and polyhydroxyalkanoates (PHA) [10]. The demand for lactic acid gradually increases every year. The global demand for LA produced from non-fossil fuel-based processes has reached an annual growth rate of 15% [11]. The production of LA from fermentation processes is conducive to reconciling the consumption of fossil energy and reaching the goal of carbon neutrality [12].
To obtain a high LA yield, the operating conditions of fermentation processes such as pH, temperature, and substrate composition should be optimised [13]. pH, as a key influencing factor, significantly affects enzymatic activity and bacterial metabolism [14]. However, reportedly, lactic acid fermentation has a rather wider pH range from 3.5 to 11 [6,15,16,17,18,19,20]. Seemingly, acidic conditions are more beneficial for LA production than alkaline conditions [6,21], and meanwhile, LA derived from FW at pH 6 was a very suitable substrate as an electron donor to facilitate the chain elongation process for caproate production [22,23]. Furthermore, the natural acidification of food waste also produces lactic acid, which indicates that food waste usually carries microorganisms that can carry out lactic acid production. Evidently, lactic acid fermentation can proceed without inoculation under anaerobic conditions [6]. Therefore, it is necessary to explore the natural fermentation of food waste under different acidic conditions.
During the AD process, organic matter such as food waste is commonly degraded, and amino acids (AAs) are the main intermediates of protein degradation. Reportedly, amino acids are also nutrients required by microorganisms, and many of the AAs cannot be synthesised during AD, and a number of the AAs must be harvested from the environment [24]. Therefore, it is important to supply certain amino acids that have important biological functions in anaerobic fermentation. For example, L-AAs (such as L-cysteine, L-alanine, and L-histidine) can serve as a source of nitrogen, promote microbial growth, and/or act as electron transporters in microbial biochemical reactions [25,26]. Certain D-AAs such as D-leucine and D-methionine can act as signalling molecules to regulate cell wall biogenesis, biofilm integrity, and spore germination, trigger microbial biofilm disintegration at low concentrations to regulate the distribution of bacterial populations, and influence the surrounding ecosystem [25,27,28]. To date, the focus on free amino acids in anaerobic digestion is mainly on the biodegradability of amino acids, which are commonly produced from substrates such as gelatin, sewage sludge, and food waste during fermentation [24,29,30,31]. Furthermore, some amino acids have been reported to play a positive role in enhancing the anaerobic digestion process. For example, L-Cysteine promoted L-glucose degradation and influenced enzymatic and microbial activity that are related to VFAs and methane production [27]. Cysteine contains sulfur groups that can promote the anaerobic conversion of unsaturated long-chain fatty acids to methane [32]. Reportedly, the contents of L-aspartate and glycine are correlated with lactic acid yield in anaerobic fermentation [33]. Valine degradation releases electrons, which can be used to steer selective production of VFAs by controlling the oxidation reduction potential (ORP) [34]. Thus, the addition of the amino acids to anaerobic fermentation systems of food waste presumably can substantially enhance the production of lactic acid or other VFAs.
However, thus far, studies examining the effects of AAs on AD have mainly focused on VFAs and methane, and little has been explored on their effects on lactic acid production. Therefore, this study aimed to investigate the effects of acidic conditions and the addition of different amino acids on lactic acid production with indigenous microorganisms in FW. Firstly, the effect on lactic acid production was examined at pH levels of 4.0, 5.0, and 6.0. Subsequently, changes in lactic acid and metabolic intermediates involved in the AD pathways by different amino acids were investigated. Furthermore, the microbial community profiles of the test groups were examined in-depth to reveal the influence of AA on LA yields and community structure.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Fermentation processes for production of high value-added products.
| Product | Substrate | Inoculum | Reactor | Operation Condition | Outcomes | Reference |
|---|---|---|---|---|---|---|
| Biohydrogen | Synthetic food waste | Clostridium acetobutulicum | Batch fermentation reactor (working volume: 1000 mL) | Temperature: 32 ± 1 °C; pH: 6.85; addition of Fe3O4 (100 mg) and TiO2 (50 mg) | Volume: 3392 mL; yield: 213.66 mL H2/g VS | [35] |
| Biohythane | Synthetic organic wastewater | Anaerobic sludge | The reactor: a feeding tank; two anaerobic reactors (R1 (1.5 L): hydrogen producing reactor, R2 (1.5 L): methane-producing reactor); two gas collectors; a final effluent tank | Sudden shock load phase: OLR: 25.0 g COD/L/d (R1); 15.7 g COD/L/d (R2) | 1.641 mol H2/mol glucose; methane production rate: 1.003 L CH4/L/d | [5] |
| Lactic acid | Coffee waste | Lactiplantibacillus plantarum WiKim0126 | Deman, rogosa and sharpe (MRS) broth | Pretreatment with hydrogen peroxide and acetic acid, along with a combination of Viscozyme L, Celluclast 1.5 L, and Pectinex Ultra SP-L. Lactiplantibacillus plantarum WiKim0126 induced fermentation with a 4% solid loading | Concentration: 22.8 g/L; productivity: 0.95 g/L/h within 24 h | [18] |
| Canteen FW | Enterococcus mundtii CGMCC 22227 | Fermenters with a working volume of 400 mL | Pretreatment of food waste: Glycosylase (2 g/kg FW mixture) was used for saccharification; reactor pH: 6.8 | Concentration: 115 g/L; yield: 0.97 g LA/g total sugar | [19] | |
| Modelled FW | Mixture of commercial yoghurt and solid compost from an industrial platform | Fed-batch fermentation at a pilot scale (12 L) | Temperature: 35 °C; pH: 5; Organic loading rate (OLR): 25 g TS/L/d for the first four days | Concentration: 68 g/L; yield: 0.38 g LA/g TS | [36] | |
| Canteen FW | Indigenous microbiota | Batch fermentation bioreactor (working volume: 5 L) | pH: 6; temperature: 25 °C; mechanically stirred: 120 rpm; TS of FW: 7% | Yield: 0.46 g LA/g TS | [6] | |
| VFAs | Vegetable waste | Anaerobic sludge | Continuous stirred tank reactors (working volume: 1 L) | Hydraulic retention time (HRT): 20 d; OLR: 3 g VS/L/d; a minor pH: 5.8 | VFAs concentration: 29.6 ± 2.1 g/L (47.0 ± 2.1 g COD/L); bioconversion yield: 49.2 ± 2.0% | [37] |
| VFAs and biohythane | Canteen FW | Anaerobic digestion sludge | Microbial electrolysis cell and anaerobic reactor (MEC-AR): working volume: 1 L, the two electrodes: carbon rod (8 × 3 × 50 mm), six square carbon felts (50 × 50 × 5 mm) | Applied voltage: 1.2 V; HRT: 10 d; pH: 5.04 ± 0.10 | Biohythane yield: 2.73 ± 0.05 L/L/d (with 34.80 ± 1.09% CH4, 14.95 ± 0.86% H2); VFAs concentration: 19.39 ± 0.44 g COD/L | [7] |
| MCCAs | Canteen FW | Indigenous microbiota | Batch fermentation (total volume: 500 mL) | VS of reactor FW: 75 g/L; pH: 6 temperature: 37 °C | Caproic acid concentration: 88.24 mM | [38] |
| FW from FW treatment plant | Acclimated pit mud obtained from a CE reactor | Continuous stirred tank reactor (working volume: 1 L) | pH: FW was adjusted to pH 6.0 ± 0.5 before feeding, reactor no control; OLR: 9.24 g COD/L/d; HRT: 20 d | MCCA concentration: 29,886.10 mg COD/L; caproic acid concentration: 28,191.66 mg COD/L | [23] |
2. Materials and Methods
2.1. Materials
In order to ensure the consistency of food waste as substrate during the experiment, food waste was prepared based on the previous studies [20,39]. A specific ingredient recipe was listed in Table 2. The ingredients were mixed, cooked, then crushed and homogenised in a grinder. The prepared food waste was stored at 4 °C for less than 10 d until further use. In addition, the naturally present microbes in the food waste were used as inoculum without external inoculation.
2.2. Effects of pH on Food Waste Fermentation
Three test groups of different pHs (4.0, 5.0, and 6.0) were set up to explore the effects of pH on lactic acid production during the fermentation of food waste. In this study, the food waste was diluted with deionised water to a TS of 8.0%, and the characteristics of the food waste are depicted in Table 3. The tests were carried out in triplicate in serum bottles with a working volume of 300 mL. The bottles were purged with nitrogen gas for 10 min to maintain strict anaerobic niches. Then the serum bottles were incubated in an incubator shaker at 150 rpm, and the temperature was maintained at 37 ± 2 °C. The pH was adjusted to 4.0, 5.0, and 6.0 with sodium hydroxide (6 M) and hydrochloric acid (6 M) solutions daily. Daily samples were taken to observe the changes in soluble chemical oxygen demand (SCOD), soluble carbohydrates, and lactic acid in the test bottles, and finally to define an appropriate pH for subsequent fermentation trials.
2.3. Effects of Amino Acid Additions on Food Waste Fermentation
According to the results harvested with the tests carried out in Section 2.2, a pH of 5.0 was set for subsequent fermentation trials, and other operational conditions were the same as those mentioned above. In the trials with AA additions, 5 groups of experiments were carried out, namely, the control group without adding any amino acid (Control), the group with adding cysteine (Cys), the group with adding aspartic acid (Asp), the group with adding glycine (Gly), and the group with adding valine (Val). The trials were all executed in triplicate in the serum bottles with a working volume of 300 mL, and for each test, 300 mg of amino acids were added. The pH was adjusted to 5.0 with sodium hydroxide (6 M) and hydrochloric acid (6 M) every 12 h. During the trials, samples were taken and analysed every 24 h.
2.4. Analytical Methods
The pH was measured using a digital meter equipped with a pH probe (ST310). TS and VS were determined using a weighing method (APHA.1998) [40]. Samples were centrifuged at 8000× g for 10 min and then filtered by membrane filters with a pore size of 0.45 μm to assay soluble components, including SCOD, soluble carbohydrate, soluble proteins, and fermentation products. COD was determined by the hash method (DRB200, HACH Company, Maricopa, Arizona, USA) [40]. Soluble carbohydrates and soluble proteins were determined using the phenol-sulfuric acid method and the Caulmers Brilliant Blue method [41], respectively. Amino acids were analysed using a fully automated amino acid analyser (EClassical 3100, Dalian Elite Analytical Instruments Co., Ltd., Dalian, China). VFAs were detected by a gas chromatograph (GC-2010, Shimadzu (Shanghai) Global Laboratory Consumables Co., Ltd., Shanghai, China), a flame ionisation detector (FID), and a fused-silica capillary column (PEG-20 M, Dalian Replete Science and Technilogy Co., Ltd., Dalian, China). The temperatures of the detector and injection port were 250 °C and 210 °C, respectively. Samples need to be pretreated before injection. Each sample was mixed with phosphoric acid (3 M) in a 1:1 ratio (v:v) and then filtered using a 0.22 μm microporous membrane to determine VFA concentrations. Lactic acid was measured using a high-performance liquid chromatograph (Ultimate 3000, Thermo Fisher Scientific (China) Co., Ltd., Shanghai, China) equipped with a Shim-pack GIST C18 AQ column (5 μm, 4.6 mm × 250 mm) and a diode array detector, with a mobile phase of 50 mmol/L NaH2PO4 (pH was adjusted to 2.1 with 85% H3PO4) at a flow rate of 1.0 mL/min, a column temperature of 40 °C, and a detector UV of 210 nm. α-Glucosidase activity was determined according to the method employed in the study of Goel et al. [42]. Lactate dehydrogenase activity was determined spectrophotometrically using a lactate dehydrogenase test kit (ELISA), and all the procedures were carried out according to the instructions on the kit. Specific methods can be found in Supplementary Materials. DNA was extracted using the PowerSoil® DNA kit (Mo Bio Laboratories, San Diego, CA, USA), and the amplification products were purified using a PCR amplifier with primer pairs 338F and 806R, respectively, and then sequenced. Microbial samples were analyzed for high-throughput sequencing (Shanghai Majorbio Bio-pharm Technology Co., Ltd., Shanghai, China).
2.5. Calculation of the Hydrolysis Rate
The hydrolysis rate was calculated using the following formula [6]:
where SCOD represents the soluble COD of the fermentation broth by the end of the tests, SCOD0 and TCOD0 denote the soluble and total COD of the influent FW, and SCODAA represents the theoretical COD of added amino acids.
2.6. Statistical Analysis of Data
All assays were conducted in triplicate, and the results were expressed as the mean ± standard deviation of biological parallel experiments. Significant differences were identified through a t-test (ANOVA), and a p < 0.05 was considered to be statistically significant.
3. Results and Discussion
3.1. Effects of pH on Lactic Acid Production from Fermentation of Food Waste
3.1.1. Effect of pH on Hydrolysis and Acid Production
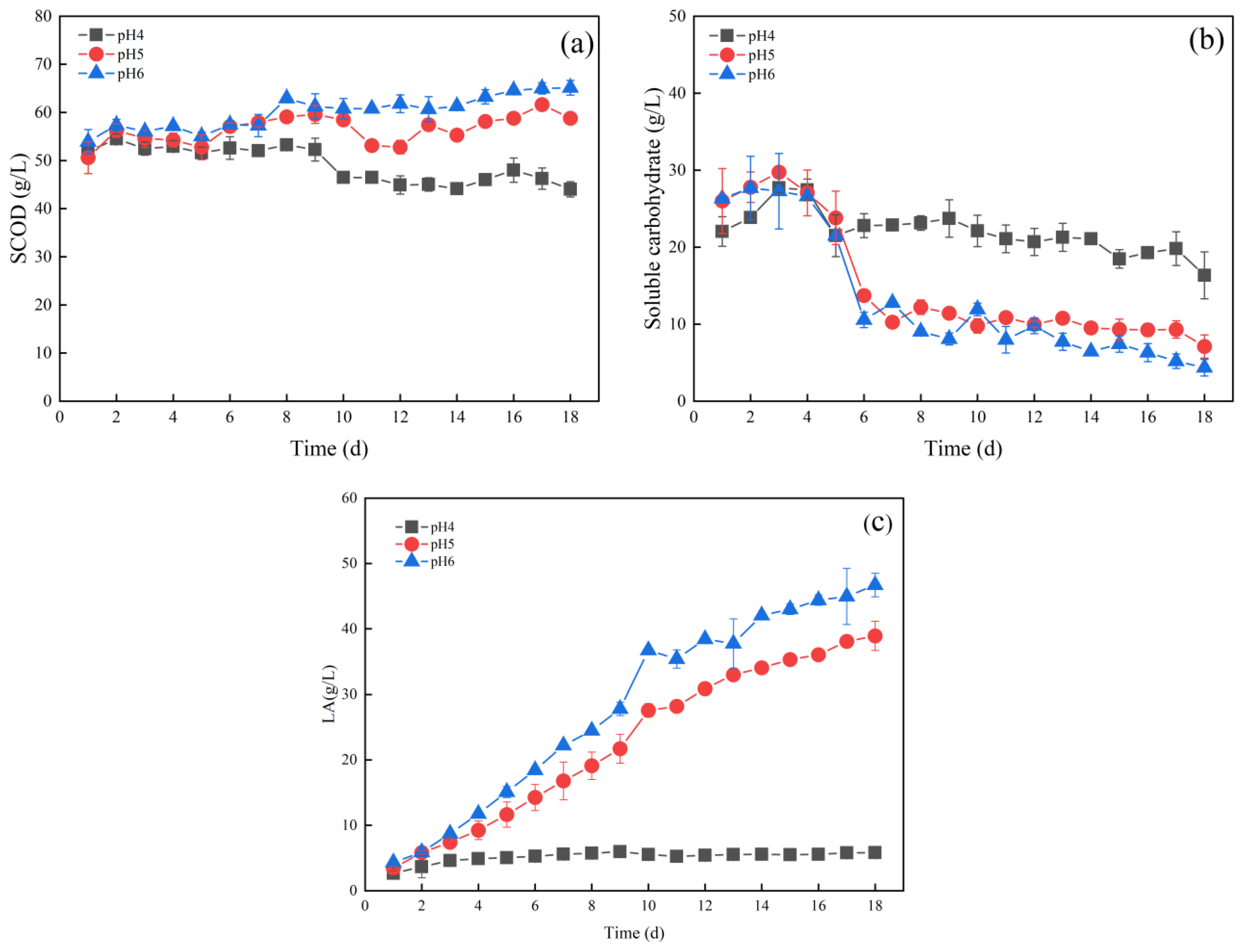
Figure 1a–c shows the changes in SCOD, soluble carbohydrates, and lactic acid during the fermentation of food waste at different pHs. In Figure 1a, SCOD increased in the first 2 days, which indicated that the leaching of solid organic matter in the initial phase was greater than the consumption of the soluble components by microbes. The SCOD of the group with a pH of 4.0 decreased to 46.67 g COD/L on the 10th day and then remained stable. However, the SCOD of the other two test groups (pH 5.0 and pH 6.0) maintained a relatively slow increase and reached a maximum of 61.63 g COD/L and 65.20 g COD/L, respectively, on the 17th day. The SCOD of the two groups (pH 5.0 and pH 6.0) increased by 41% and 33%, compared with that of the group of pH 4.0, respectively. The increase in SCOD at pH 5.0 and 6.0 proved that higher pH seemed to be more favourable for solids dissolution during the fermentation processes, which well agreed with the results reported by Tang et al. [43]. Acidic fermentation of food waste with an increase in pH was more conducive to the solubilisation of food waste.
In Figure 1b, in the first 3 days, an increase in the content of soluble carbohydrates under the three pH conditions was observed, indicating that the conversion of insoluble substances to soluble carbohydrates was seemingly greater than that of soluble carbohydrates to small molecules. Then the content of soluble carbohydrates began to decline from day 3 onwards, whereas the soluble carbohydrates concentration in the group of pH 4.0 remained almost unchanged after 5 days, which might be attributed to the severe inhibitory effects on acidogenic bacteria under the condition. The groups of pH 5.0 and pH 6.0 showed a continuous decrease in soluble carbohydrates content, down to 7.10 g/L and 4.35 g/L at 18 d, respectively, as higher pH seemingly alleviated acid inhibition and significantly promoted bacterial activity [6,44]. The degradation rate of soluble carbohydrates significantly increased with increasing pH, and higher pH at acidic conditions increased the reaction rate and shortened the fermentation period.
Figure 1c indicates no substantial increase in lactic acid concentration in the group of pH 4.0, which merely reached 5.81 g COD/L at 18 d. Evidently, lactic acid fermentation in the group was inhibited, whereas the lactic acid concentration continued to increase at pH 5.0 and pH 6.0, reaching 38.92 g COD/L and 46.70 g COD/L at 18 d. The result was also in agreement with the result of the degradation of soluble carbohydrates. The lower pH condition (pH 4.0) apparently led to the accumulation of non-protonated fatty acids and further substantially undermined the metabolic pathways of the lactobacilli. Hydrolysis and acidification were presumably completely inhibited under the condition, resulting in lower lactic acid yields [45,46]. Compared with that of the group at pH 4.0, lactic acid yield could be promoted at pH 5.0 and pH 6.0. The group at pH 5.0 is more economical in the use of sodium hydroxide compared to pH 6.0, and thus the subsequent pH conditions of the fermentation tests were defined as pH 5.0.
3.1.2. Variations of Amino Acids during Fermentation at pH 5.0
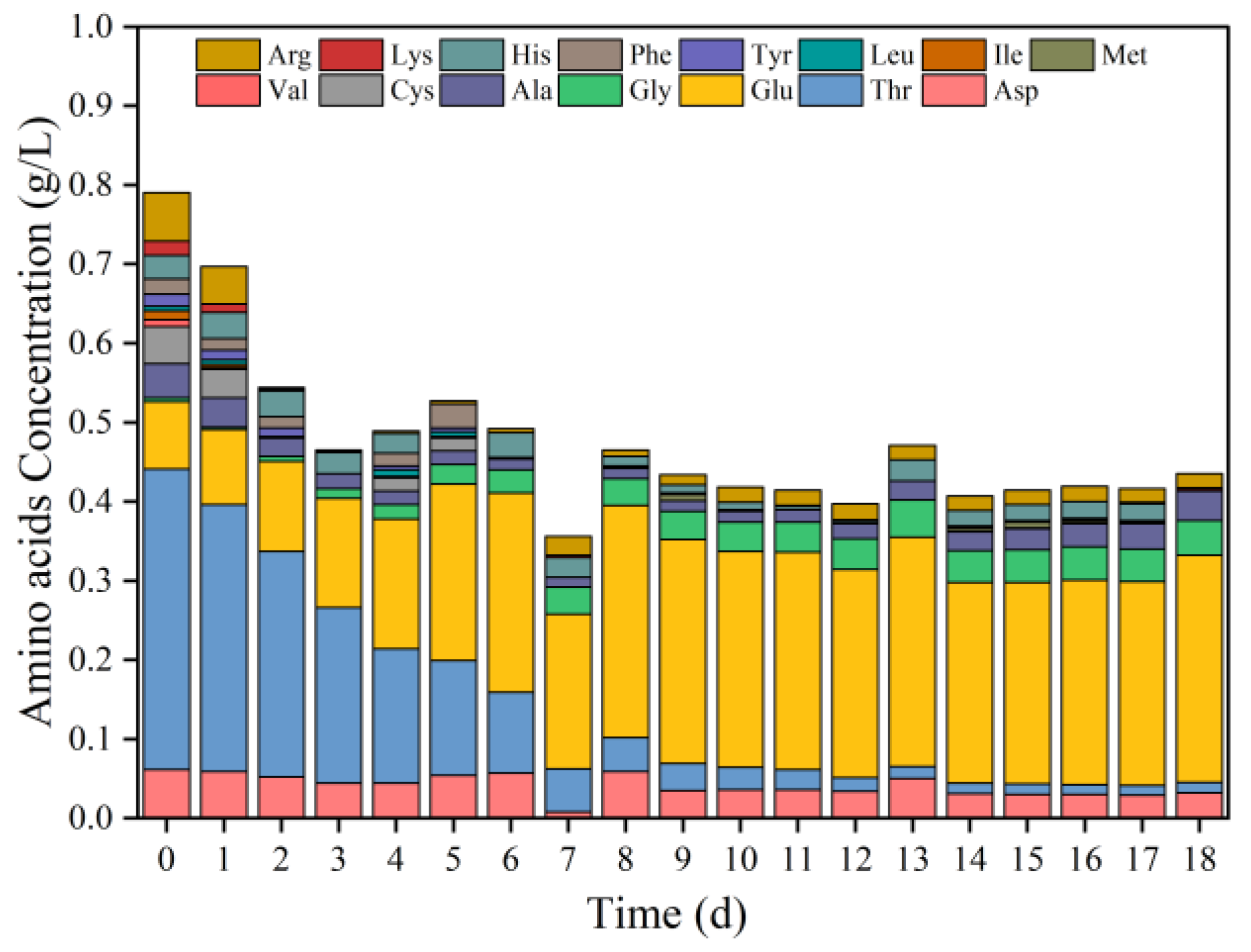
Commonly, amino acids are also key growth factors, just such as pyrimidines, purines, and vitamins, which are essential for LAB growth and reproduction [8,33]. The results of Section 3.1.1 indicate that fermentation of food waste without exogenous inoculation presented better LA yields at pH 5.0. Thus, ionic amino acids were measured to understand the changes in amino acids during the fermentation of food waste at pH 5.0. Figure 2 shows the changes of various soluble amino acids under pH 5.0. Rich amino acid species existed in the original FW, and aspartic acid (Asp), threonine (The), glutamic acid (Glu), alanine (Ala), cysteine (Cys), and arginine (Arg) were the main components of amino acids in the food waste. These six amino acids accounted for about 85% of the total amino acids. As the fermentation proceeded, glutamic acid presented evident accumulation, increasing from the initial 0.085 g/L to 0.287 g/L, accounting for 65% of the content of all amino acids, and becoming the predominant amino acid at the end of fermentation. The degradation of proteins in the food waste led to the production of a higher concentration of glutamate, and seemingly it was difficult to be degraded by microbes in the group, thus leading to its accumulation. Notably, Glu and Ala were not only key cytoarchitectural components and central metabolites but could be biosynthesised at a relatively low energy cost [24,31]. In addition, threonine declined from the initial 0.38 g/L to 0.013 g/L, and cysteine was quickly degraded and utilised, while valine, iso-leucine, leucine, and other amino acids with low initial content were quickly consumed and completely degraded. The amino acids were presumably utilised by microorganisms for conversion to VFAs. During anaerobic fermentation, amino acids are degraded into smaller organic matter, such as VFAs.
3.2. Effects of Adding Different Amino Acids on Food Waste Fermentation
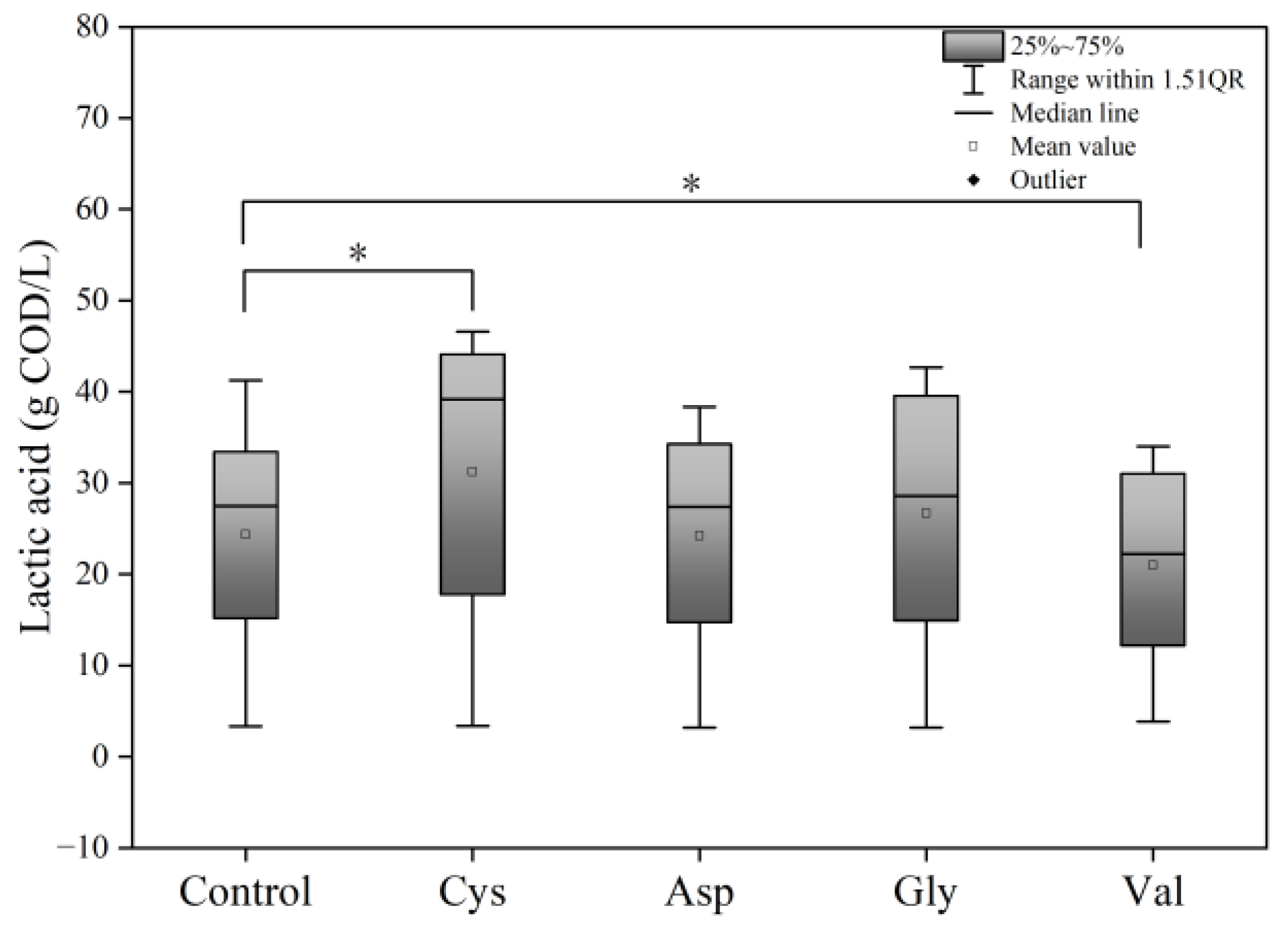
Accordingly, cysteine, aspartic acid, glycine, and valine were selected as exogenously added amino acids, which were based on the changes in amino acid concentrations in food waste fermentation at pH 5.0 in this current study. A few studies showed that four amino acids had special biological effects in anaerobic systems [28,34,47]. The effects of the four amino acids on lactic acid production from food waste fermentation were first explored. The effects of pH variations on amino acids can be ruled out by Figure S1. Figure 3 represents the ANOVA analysis of the results of the LA concentration every 24 h and indicates Asp and Gly had no significant effects on lactic acid production at the same dose, compared to that of the control group (p > 0.05). The addition of Cys and Val presented distinctive effects on lactic acid production within the trials compared to that of the control group (p < 0.05). Therefore, the subsequent investigation mainly focused on how Cys and Val affected the fermentation of food waste. During anaerobic fermentation, amino acids were degraded from proteins, which were first hydrolysed proteases into peptides and amino acids [48]. Amino acids were then broken down in two main ways: (1) pairs of amino acids can be degraded by bacteria within the genus Clostridium (obligate anaerobes) via Stickland reaction. In the reactor, an amino acid acts as an electron donor, while another amino acid acts as an electron acceptor, offering cells approximately 0.5 moles of ATP per mole of amino acid transformed during the reaction [30]; (2) individual amino acids can also be fermented in processes that require the presence of hydrogen utilising bacteria. Commonly, the degradation products of these amino acids under anaerobic conditions are organic compounds (short-chain and branched-chain organic acids), ammonia, carbon dioxide, and small amounts of hydrogen and sulfur compounds [24,42]. However, certain amino acids can also be degraded during anaerobic fermentation to generate lactic acid. Thus, to determine the impact of the added amino acids on the lactic acid content due to its conversion, the theoretical amount of lactic acid produced due to the added amino acids was estimated. When all degraded AAs were converted to lactic acid, the theoretical lactic acid yield was calculated using the reaction equation. The corresponding theoretical lactic acid yields for the degradation of 1.00 g of Cys and Val were 0.74 and 0.77 g, respectively (Equations (1)–(3)), which was negligible for lactic acid yield compared with the degradation of sugars.
Cys: C3H7NO2S + H2O → C3H4O3 + H2S + NH3
C3H4O3 + NADH + [H] → C3H6O3 + [NAD+]
Val: C5H11NO2 + 5H2O → C3H6O3 + NH3 + 2CO2 + 6H2
3.2.1. Effects of Different Amino Acids on Hydrolysis
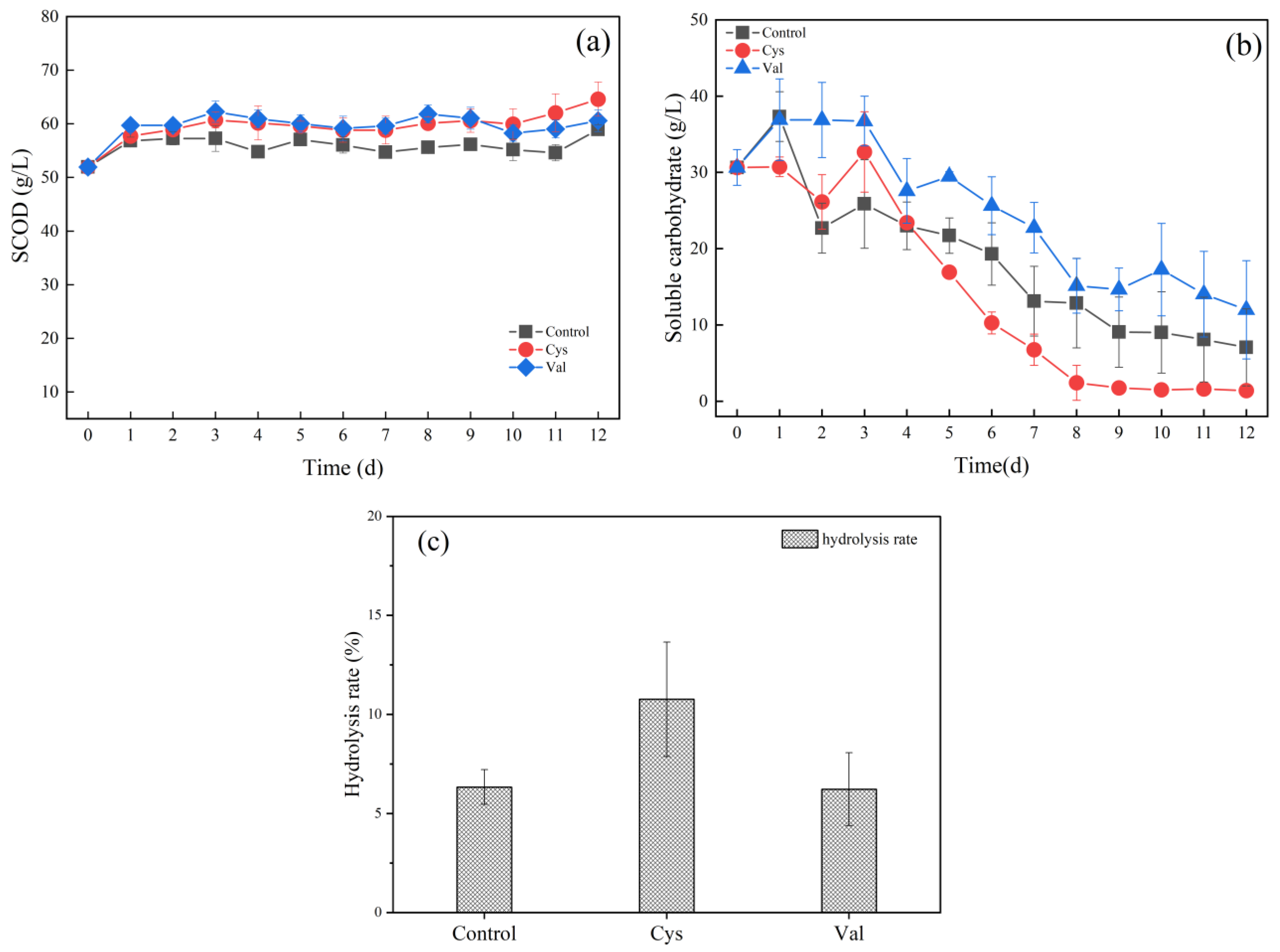
SCOD was the sum of soluble organic matter in the supernatant, and a higher SCOD indicated more potential fermentation substrate in the reactor and thereby provided more accessible pathways for microbes and enzymes to utilise [15]. Figure 4a shows the variation of SCOD with time for groups with additions of different amino acids. On day 1, there was a significant increase in SCOD for each group, which indicated that the insoluble solids in the reactor were gradually solubilised, and the increase of soluble matter can also be seen from the change in soluble carbohydrates content in Figure 4b. The subsequent SCOD remained nearly stable during the fermentation process, and that of the Control, Cys, and Val groups reached 58.93 g/L, 64.60 g/L, and 60.60 g/L, respectively. Based on the results of the ANOVA, the differences between the three groups of SCOD were also significant (p < 0.05). From the overall trend, the SCOD of the Cys and Val groups was higher than that of the control group. Figure 4c indicates that the soluble matter increased, and the hydrolysis rates of the three groups were 6.34%, 10.76%, and 6.23%, respectively. The addition of cysteine presented a positive effect on the dissolution of insoluble solids from food waste.
Carbohydrates are more abundant components of food waste because they are commonly easier to degrade and utilise than proteins and lipids. Soluble carbohydrates can be hydrolysed to monosaccharides during hydrolysis. The degradation of monosaccharides usually proceeds faster than that of amino acids under anaerobic conditions [49]. From the overall trend of soluble carbohydrates (Figure 4b), the content of soluble carbohydrates in the Cys group was almost completely consumed in the first 8 days, whereas in the Control and Val groups in the 12 d the content still had 7.05 g/L and 12.00 g/L unutilised, respectively, suggesting that the addition of Cys accelerated the utilisation of soluble carbohydrates and increased the rate of hydrolysis of carbohydrates in food waste. α-Glucosidase is a hydrolytic enzyme for the degradation of carbohydrates [42,43]. The α-glucosidase activity of the groups of different amino acids at the first 5 days was analyzed, and the mean values of the α-glucosidase activity of different amino acid groups of the 5 days are listed in Table 4. Table 4 shows that the addition of cysteine increased the enzyme activity of α-glucosidase by 22% compared with that of the control group, which was consistent with the soluble carbohydrate content (Figure 4b). Pyruvate was an important metabolic intermediate, and monosaccharides can be converted to pyruvate and then utilised by microorganisms to produce lactic acid, and α-glucosidase played a crucial role in the degradation of soluble carbohydrates. Lactic acid production in food waste can reportedly be altered by modulating the activity of some key enzymes, such as α-glucosidase [42]. The content of soluble carbohydrates in the Val Group was lower than that of the Control Group, which suggests Val seemingly inhibited the hydrolysis of soluble carbohydrates. During degradation, valine could release more electrons and make the system more reductive (i.e., lower ORP) [34]. The ORP below zero is often referred to as an intermediate transition between an oxidative and reductive environment, which reportedly could affect the hydrolysis of proteins and carbohydrates in various substrates [50]. Figure S2 represents the changes in soluble protein, where all three groups first underwent a slight decrease in protein, and then the protein content increased to about 1.0 g/L. The differences between the three groups were not significant due to the low soluble protein content during fermentation.
3.2.2. Effects of Different Amino Acids on Acidification
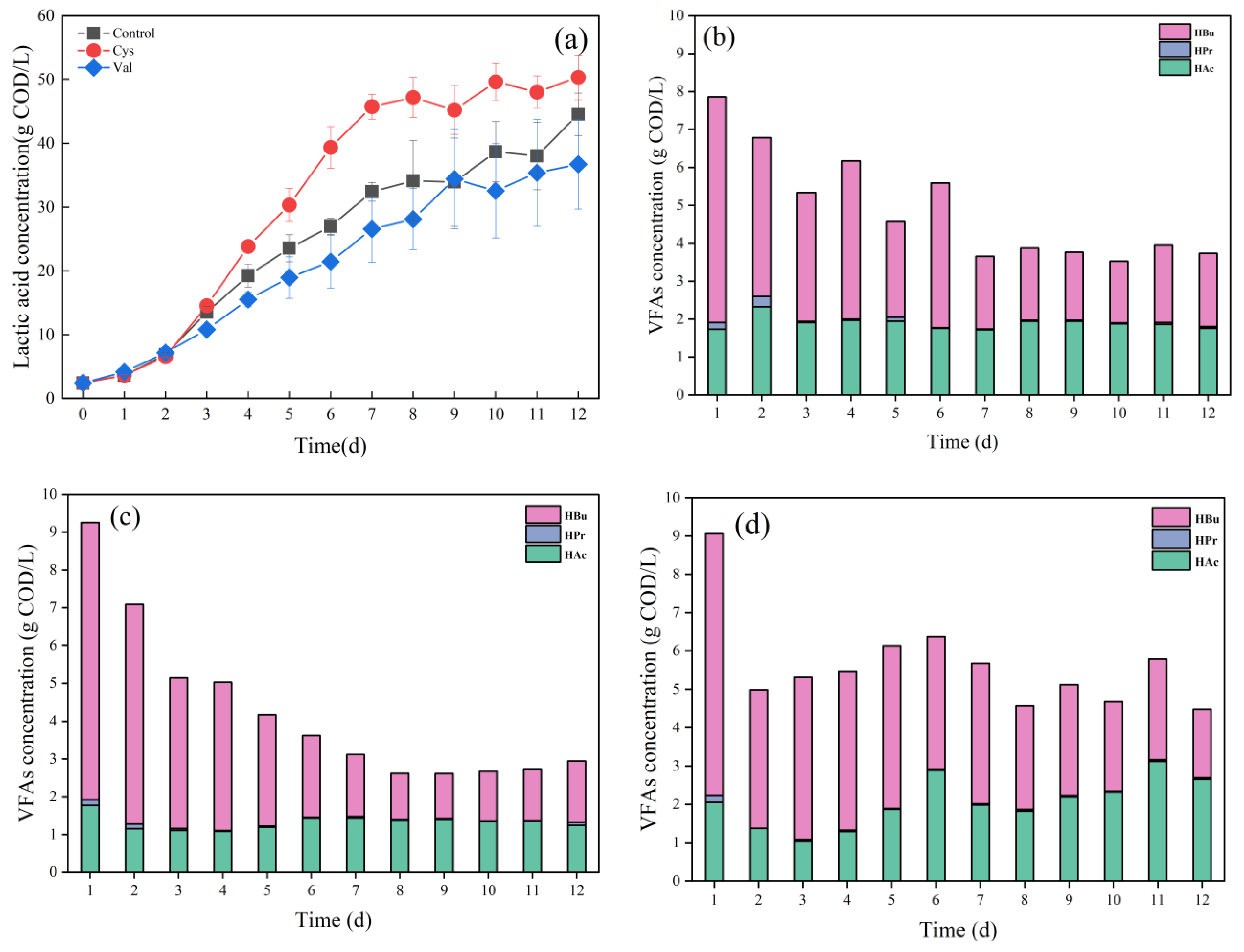
During acidification, various organic acids, such as lactic acid and VFAs, were produced. Pyruvate produced by the metabolism of soluble carbohydrates and proteins can be converted to lactic acid under the action of lactate dehydrogenase (LDH). Figure 5a shows the changes of lactic acid after the addition of different amino acids. The maximum concentration of lactic acid was 50.34 g COD/L in the Cys group, reaching the maximum on the 7th day, and the lactic acid concentration in the Control and Val groups were 32.40 g COD/L and 26.56 g COD/L, respectively, on the 7th day. The yields of lactic acid in the Control and Val groups were significantly lower than those of the Cys group (Table 4), indicating the addition of cysteine not only promoted the production of lactic acid but also seemingly shortened the period of lactic acid fermentation. The trend of the LA concentration well agreed with the trend of soluble carbohydrate degradation in the Cys group. The addition of cysteine substantially promoted the conversion of soluble carbohydrates, and thus more monosaccharides were available in the Cys group in the early stage, which might be more potent for lactic acid production. The lactate dehydrogenase activity was analyzed on day 7 (Table 4), which demonstrated that the lactate dehydrogenase activity was the highest in the Cys group. However, the Val group presented a certain inhibitory effect on the lactate dehydrogenase, which suggested that the addition of cysteine likely improved the rate of degradation of soluble carbohydrates and enhanced the activity of α-glucosidase. In turn, the addition of cysteine presented a certain positive effect on the activity of lactate dehydrogenase, shortened the productive period of lactic acid, and thus subsequently promoted lactic acid production.
In addition, the composition of VFAs varied among the groups with different amino acids. Figure 5b–d show the changes in VFAs in the control group, Cys, and Val groups, respectively. The two groups with the addition of amino acids presented much higher levels of VFAs than the control group, and the dominant VFA species during fermentation in the three groups were acetic acid (HAc) and butyric acid (HBu), accompanied by a low concentration of propionic acid (HPr) throughout the entire fermentation period. Butyric acid contents of three groups on day 2 were significantly higher, and perhaps the microorganisms present in the food waste were more favourable for butyric acid production at the beginning. However, as the fermentation proceeded, the dominant microorganisms changed to be more favourable for lactic acid production. The final acetic acid content was higher in the Val group, and valine reportedly could release more electrons during degradation, making the niche of the group more reducing (lower ORP) and more favourable for the accumulation of VFAs [34]. Probably the added Val facilitated the conversion of lactic acid to acetic acid, which led to a higher acetic acid content in the Val group.
3.2.3. Microbial Community Analysis
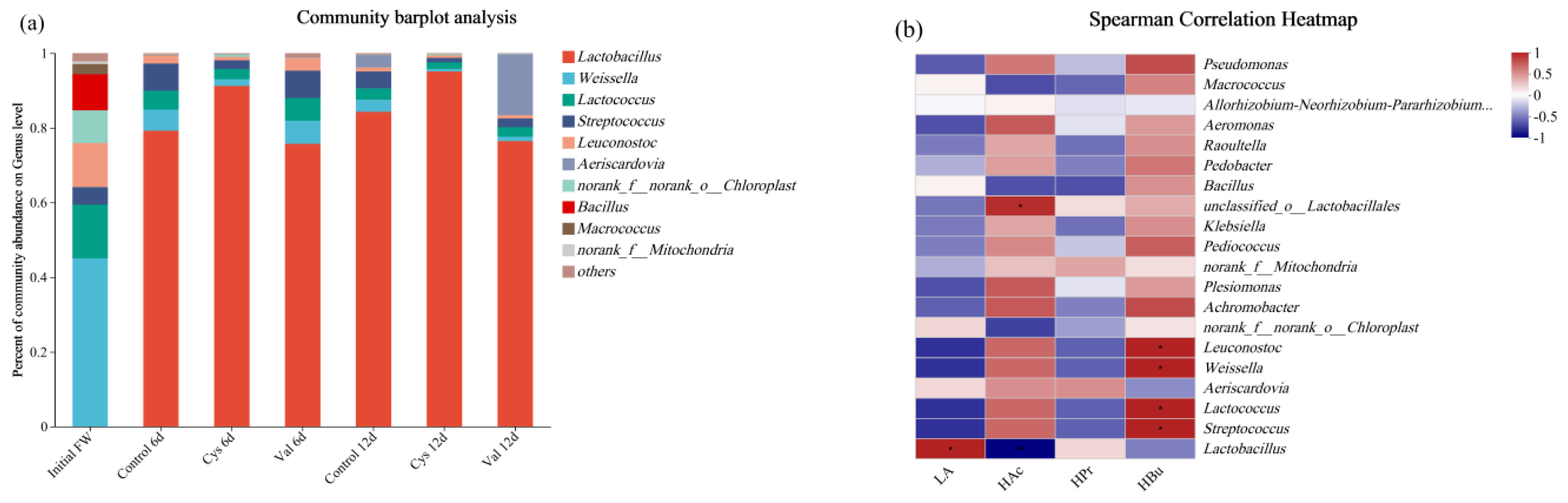
In order to decipher the effects of additions of different amino acids on the microbial community, samples from days 6 and 12 of each reactor were collected and analysed (Figure 6). Figure 6b shows that Lactobacillus spp. was positively correlated with lactic acid production. Lactic acid bacteria (LAB), such as Lactobacillus spp., have been reported to exist at very low pH conditions and produce lactic acid at a pH of 4.0 [51]. Lactobacillus spp. was the main contributor to the production of lactic acid, as it was evidently dominant in the whole process (Figure 6b). The most dominant genus in the raw food waste was the genus Weissella (45%), and Lactobacillus was not the most dominant microorganism in the raw food waste. However, during the fermentation over time, Lactobacillus dominated in all three reactors, with an increase of 84%, 95%, and 76% in the control, Cys, and Val groups in the end. The changes in the relative abundance of Lactobacillus and the enhanced lactic acid yields were also in good agreement. Butyric acid production was seemingly related to Streptococcus, Lactococcus, Weissella, and Leuconostoc. These bacteria can produce other organic acids, such as acetic and butyric acids, from organic matter and also perform lactic acid fermentation in certain niches [52,53,54]. As shown in Figure 6a, on day 6, the proportion of these bacteria in the valine group was evidently higher than that in the control group. The percentage in the group with cysteine was evidently lower than that in the control group, which was consistent with the organic acid content of the three groups, implying that cysteine and valine affected the distribution of microbial species in the reactors. Amino acids in anaerobic media can affect microbial growth as inhibitory or stimulatory compounds [45]. Thus, the increase in the rate of lactic acid production due to the addition of cysteine and valine might be related to their function in the basic metabolic pathways. These results revealed the selectivity of microorganisms in bioreactors and food waste as a suitable substrate for lactic acid fermentation. Furthermore, the addition of cysteine to the fermentation of food waste could increase the abundance of Lactobacillus and thus promote the production of lactic acid.
4. Conclusions
The present study demonstrated that higher LA concentrations (38.92–46.7 g/L) were obtained at pH 5.0 and pH 6.0, implying higher substrate degradation rates compared with those at pH 4.0. Different amino acids presented distinctive effects on the production of lactic acid from food waste. Cysteine promoted substrate solubilisation and carbohydrate degradation and subsequently facilitated lactic acid production, in which enhanced key enzyme (LDH) activity and enriched Lactobacillus played an essential role during the acidification phase. However, valine evidently inhibited the degradation of carbohydrate due to the mitigated α-glucosidase activity, limited the reduction of available pyruvate in cells, and thus subsequently suppressed lactic acid production. This study examined in-depth the mechanisms and significance of pH regulation and the addition of amino acids in lactic acid production from food waste. Furthermore, this current study offers a feasible way to improve lactic acid production from food waste. Using cysteine-rich substrates is worth exploring for lactic acid fermentation instead of pure amino acids. In addition, attention should be paid to the production of amino acids as intermediate products of anaerobic fermentation in future studies. Different amino acids can have distinctive biological functions, which should be diligently explored in order to efficiently regulate anaerobic digestion through the addition of specific AAs.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/fermentation10040179/s1, Figure S1: Variations of pH in the reactors with pH adjustment every 12 h; Figure S2: Soluble protein for three groups (Control, Cys, and Val).
Author Contributions
Methodology, Data curation, Writing—original draft, Investigation, Y.Z.; Funding acquisition, Conceptualization, Writing—review and editing, X.Z. and H.L.; Methodology, Formal analysis, Y.W. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Taihu Talent Program—Innovative Individual (2022) and “Shuang Chuang Doctoral Talent” of Jiangsu Province (2022) (NO. JSSCBS20221049).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data generated or analyzed during this study are included in this published article.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Zhang, W.; Li, X.; He, Y.; Xu, X.; Chen, H.; Zhang, A.; Liu, Y.; Xue, G.; Makinia, J. Ammonia amendment promotes high rate lactate production and recovery from semi-continuous food waste fermentation. Bioresour. Technol. 2020, 302, 122881. [Google Scholar] [CrossRef]
- Zhou, M.M.; Yan, B.H.; Wong, J.W.C.; Zhang, Y. Enhanced volatile fatty acids production from anaerobic fermentation of food waste: A mini-review focusing on acidogenic metabolic pathways. Bioresour. Technol. 2018, 248, 68–78. [Google Scholar] [CrossRef]
- Wang, Q.; Feng, K.; Li, H. Nano iron materials enhance food waste fermentation. Bioresour. Technol. 2020, 315, 123804. [Google Scholar] [CrossRef] [PubMed]
- De Groof, V.; Coma, M.; Arnot, T.; Leak, D.J.; Lanham, A.B. Selecting fermentation products for food waste valorisation with HRT and OLR as the key operational parameters. Waste Manag. 2021, 127, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Wirasembada, Y.C.; Shin, B.; Shin, J.; Kurniawan, A.; Cho, J. Effects of sudden shock load on simultaneous biohythane production in two-stage anerobic digestion of high-strength organic wastewater. Bioresour. Technol. 2024, 394, 130186. [Google Scholar] [CrossRef]
- Tang, J.L.; Wang, X.C.; Hu, Y.S.; Zhang, Y.M.; Li, Y.Y. Lactic acid fermentation from food waste with indigenous microbiota: Effects of pH, temperature and high OLR. Waste Manag. 2016, 52, 278–285. [Google Scholar] [CrossRef] [PubMed]
- Bian, C.L.; Chen, X.Y.; Wang, J.; Xiao, B.Y.; Liu, R.Z.; Li, L.; Liu, J.G. Simultaneous biohythane and volatile fatty acids production from food waste in microbial electrolysis cell-assisted acidogenic reactor. J. Clean. Prod. 2023, 420, 138370. [Google Scholar] [CrossRef]
- Sunhoon, K.; Pyung, C.L.; Eun, G.L.; Yong, K.C.; Nam, C. Production of lactic acid by Lactobacillusrhamnosus with vitamin-supplemented soybean hydrolysate. Enzym. Microb. Technol. 2000, 26, 209–215. [Google Scholar]
- Daly, S.E.; Usack, J.G.; Harroff, L.A.; Booth, J.G.; Keleman, M.P.; Angenent, L.T. Systematic Analysis of Factors That Affect Food-Waste Storage: Toward Maximizing Lactate Accumulation for Resource Recovery. ACS Sustain. Chem. Eng. 2020, 8, 13934–13944. [Google Scholar] [CrossRef]
- Ma, X.; Gao, M.; Li, C.; Wang, N.; Wang, Q.; Sun, X.; Ma, X. Effects of different lignocellulosic wastes on alleviating acidification of L-lactic acid production from food waste fermentation. Bioresour. Technol. 2021, 342, 126043. [Google Scholar] [CrossRef]
- Ye, T.; Li, X.; Zhang, T.; Su, Y.; Zhang, W.; Li, J.; Gan, Y.; Zhang, A.; Liu, Y.; Xue, G. Copper (II) addition to accelerate lactic acid production from co-fermentation of food waste and waste activated sludge: Understanding of the corresponding metabolisms, microbial community and predictive functional profiling. Waste Manag. 2018, 76, 414–422. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.X.; Cui, M.; Wang, B.; Zhang, C.H.; Zhu, Y.L.; Jin, X.; Wang, R.; Zhang, X.Q. Electrospun Environment-Friendly Poly (L-Lactic Acid)/CO2-Based Polyurea Nanofiber Film for Piezoelectric Sensor. Adv. Sustain. Syst. 2023, 7, 2200441. [Google Scholar] [CrossRef]
- Xu, X.; Zhang, W.; Gu, X.; Guo, Z.; Song, J.; Zhu, D.; Liu, Y.; Liu, Y.; Xue, G.; Makinia, J. Stabilizing lactate production through repeated batch fermentation of food waste and waste activated sludge. Bioresour. Technol. 2020, 300, 122709. [Google Scholar] [CrossRef] [PubMed]
- Bao, X.Y.; Feng, H.R.; Guo, G.; Huo, W.J.; Li, Q.H.; Xu, Q.F.; Liu, Q.; Wang, C.; Chen, L. Effects of laccase and lactic acid bacteria on the fermentation quality, nutrient composition, enzymatic hydrolysis, and bacterial community of alfalfa silage. Front. Microbiol. 2022, 13, 1035942. [Google Scholar] [CrossRef] [PubMed]
- Feng, K.; Li, H.; Zheng, C.Z. Shifting product spectrum by pH adjustment during long-term continuous anaerobic fermentation of food waste. Bioresour. Technol. 2018, 270, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Yue, L.; Chuan, S.; Wu, Y.Y.; Han, D.; Li, K.; Ma, J.Y. Effect of pH dynamic control on ethanol-lactic type fermentation (ELTF) performance of glucose. Environ. Technol. 2022, 43, 4102–4114. [Google Scholar] [CrossRef]
- Wu, Y.Y.; Ma, H.L.; Zheng, M.Y.; Wang, K.J. Lactic acid production from acidogenic fermentation of fruit and vegetable wastes. Bioresour. Technol. 2015, 191, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Kim, J.C.; Kim, Y.Y.; Yang, J.E.; Lee, H.M.; Hwang, I.M.; Park, H.W.; Kim, H.M. Utilization of coffee waste as a sustainable feedstock for high-yield lactic acid production through microbial fermentation. Sci. Total Environ. 2024, 912, 169521. [Google Scholar] [CrossRef]
- Liu, S.; Wang, Q.H.; Li, Y.; Zhu, W.B.; Wang, N.H.; Sun, H.S.; Gao, M.; Ma, X.Y. Highly efficient oriented bioconversion of food waste to lactic acid in an open system: Microbial community analysis and biological carbon fixation evaluation. Bioresour. Technol. 2023, 370, 128398. [Google Scholar] [CrossRef]
- Yang, L.; Chen, L.; Li, H.; Deng, Z.; Liu, J. Lactic acid production from mesophilic and thermophilic fermentation of food waste at different pH. J. Environ. Manag. 2022, 304, 114312. [Google Scholar] [CrossRef]
- Bonk, F.; Bastidas, O.J.R.; Yousef, A.F.; Schmidt, J.E.; Bonk, F. Exploring the selective lactic acid production from food waste in uncontrolled pH mixed culture fermentations using different reactor configurations. Bioresour. Technol. 2017, 238, 416–424. [Google Scholar] [CrossRef] [PubMed]
- Montecchio, D.; Gazzola, G.; Gallipoli, A.; Gianico, A.; Braguglia, C.M. Medium chain Fatty acids production from Food Waste via homolactic fermentation and lactate/ethanol elongation: Electron balance and thermodynamic assessment. Waste Manag. 2024, 177, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.Y.; Bai, J.Z.; Zuo, J.E. Performance and mechanisms of medium-chain fatty acid production by anaerobic fermentation of food waste without external electron donors. Bioresour. Technol. 2023, 374, 128735. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Yin, Q.D.; Zhou, X.Z.; Guo, Q.N.; Wu, G.X. Distribution of extracellular amino acids and their potential functions in microbial cross-feeding in anaerobic digestion systems. Bioresour. Technol. 2022, 360, 127535. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Xu, H.T.; Ma, C.W.; Zhu, Z.H.; Xu, T.C.; Guo, Y.L.; Ye, J.F. Review on the intermediate amino acids and their enantiomers during the anaerobic digestion: The distribution, biofunctions and mechanisms. Rev. Environ. Sci. Bio-Technol. 2022, 21, 469–482. [Google Scholar] [CrossRef]
- Helmstetter, F.; Arnold, P.; Höger, B.; Petersen, L.M.; Beitz, E. Formate-nitrite transporters carrying nonprotonatable amide amino acids instead of a central histidine maintain pH-dependent transport. J. Biol. Chem. 2019, 294, 623–631. [Google Scholar] [CrossRef]
- Liu, H.; Chen, Y.G. Enhanced Methane Production from Food Waste Using Cysteine To Increase Biotransformation of L-Monosaccharide, Volatile Fatty Acids, and Biohydrogen. Environ. Sci. Technol. 2018, 52, 3777–3785. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhang, X.; Huang, H.; Qin, Z.; Liu, C.; Chen, Y. Amino Acid Configuration Affects Volatile Fatty Acid Production during Proteinaceous Waste Valorization: Chemotaxis, Quorum Sensing, and Metabolism. Environ. Sci. Technol. 2022, 56, 8702–8711. [Google Scholar] [CrossRef]
- Reggiani, R.; Bertani, A. Anaerobic amino acid metabolism. Russ. J. Plant Physiol. 2003, 50, 733–736. [Google Scholar] [CrossRef]
- Ramsay, I.R.; Pullammanappallil, P.C. Protein degradation during anaerobic wastewater treatment: Derivation of stoichiometry. Biodegradation 2001, 12, 247–257. [Google Scholar] [CrossRef]
- Chen, S.S.; Dong, B.; Dai, X.H.; Wang, H.Y.; Li, N.; Yang, D.H. Effects of thermal hydrolysis on the metabolism of amino acids in sewage sludge in anaerobic digestion. Waste Manag. 2019, 88, 309–318. [Google Scholar] [CrossRef]
- Yekta, S.S.; Elreedy, A.; Liu, T.; Hedenström, M.; Isaksson, S.; Fujii, M.; Schnürer, A. Influence of cysteine, serine, sulfate, and sulfide on anaerobic conversion of unsaturated long-chain fatty acid, oleate, to methane. Sci. Total Environ. 2022, 817, 152967. [Google Scholar] [CrossRef]
- Song, L.; Yang, D.; Liu, R.; Liu, S.; Dai, X. The dissolution of polysaccharides and amino acids enhanced lactic acid production from household food waste during pretreatment process. Sci. Total Environ. 2023, 864, 161068. [Google Scholar] [CrossRef]
- Chen, B.Y.; Rupani, P.F.; Azman, S.; Dewil, R.; Appels, L. A redox-based strategy to enhance propionic and butyric acid production during anaerobic fermentation. Bioresour. Technol. 2022, 361, 127672. [Google Scholar] [CrossRef]
- Masihi, F.; Rezaeitavabe, F.; Karimi-Jashni, A.; Riefler, G. Optimization and enhancement of biohydrogen production in a single-stage hybrid (dark/photo) fermentation reactor using Fe3O4 and TiO2 nanoparticles. Int. J. Hydrogen Energy 2024, 52, 295–305. [Google Scholar] [CrossRef]
- Chenebault, C.; Moscoviz, R.; Trably, E.; Escudié, R.; Percheron, B. Lactic acid production from food waste using a microbial consortium: Focus on key parameters for process upscaling and fermentation residues valorization. Bioresour. Technol. 2022, 354, 127230. [Google Scholar] [CrossRef]
- Gonçalves, M.J.; González-Fernández, C.; Greses, S. Long hydraulic retention time mediates stable volatile fatty acids production against slight pH oscillations. Waste Manag. 2024, 176, 140–148. [Google Scholar] [CrossRef]
- Tang, J.L.; Yang, H.; Pu, Y.H.; Hu, Y.S.; Huang, J.; Jin, N.; He, X.R.; Wang, X.C. Caproic acid production from food waste using indigenous microbiota: Performance and mechanisms. Bioresour. Technol. 2023, 387, 129687. [Google Scholar] [CrossRef]
- Wang, Q.; Yang, L.; Feng, K.; Li, H.; Deng, Z.; Liu, J. Promote lactic acid production from food waste fermentation using biogas slurry recirculation. Bioresour. Technol. 2021, 337, 125393. [Google Scholar] [CrossRef]
- AHPA. Standard Methods for the Examination of Water and Wastewater; American Public Health Association: Washington, DC, USA, 1998. [Google Scholar]
- Lowry, O.; Rosebrough, N.; Farr, A.L.; Randall, R. Protein Measurement with the Folin Phenol Reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Goel, R.; Mino, T.; Satoh, H.; Matsuo, T. Enzyme activities under anaerobic and aerobic conditions inactivated sludge sequencing batch reactor. Water Res. 1998, 32, 2081–2088. [Google Scholar] [CrossRef]
- Tang, J.; Wang, X.C.; Hu, Y.; Zhang, Y.; Li, Y. Effect of pH on lactic acid production from acidogenic fermentation of food waste with different types of inocula. Bioresour. Technol. 2017, 224, 544–552. [Google Scholar] [CrossRef]
- Wu, Q.L.; Guo, W.Q.; Zheng, H.S.; Chao, L.H.; Chi, F.X.; Li, Y.R.; Ren, N.Q. Enhancement of volatile fatty acid production by co-fermentation of food waste and excess sludge without pH control: The mechanism and microbial community analyses. Bioresour. Technol. 2016, 216, 653–660. [Google Scholar] [CrossRef]
- Aljundi, I.H.; Belovich, J.M.; Talu, O. Adsorption of lactic acid from fermentation broth and aqueous solutions on Zeolite molecular sieves. Chem. Eng. Sci. 2005, 60, 5004–5009. [Google Scholar] [CrossRef]
- Dalie, D.K.D.; Deschamps, A.M.; Richard-Forget, F. Lactic acid bacteria—Potential for control of mould growth and mycotoxins: A review. Food Control 2010, 21, 370–380. [Google Scholar] [CrossRef]
- Liu, H.; Chen, Y.; Ye, J.; Xu, H.; Zhu, Z.; Xu, T. Effects of different amino acids and their configurations on methane yield and biotransformation of intermediate metabolites during anaerobic digestion. J. Environ. Manag. 2021, 296, 113152. [Google Scholar] [CrossRef]
- Hassan, M.A.; Abol-Fotouh, D.; Omer, A.M.; Tamer, T.M.; Abbas, E. Comprehensive insights into microbial keratinases and their implication in various biotechnological and industrial sectors: A review. Int. J. Biol. Macromol. 2020, 154, 567–583. [Google Scholar] [CrossRef]
- Xia, A.; Cheng, J.; Ding, L.K.; Lin, R.C.; Song, W.L.; Su, H.B.; Zhou, J.H.; Cen, K.F. Substrate consumption and hydrogen production via co-fermentation of monomers derived from carbohydrates and proteins in biomass wastes. Appl. Energy 2015, 139, 9–16. [Google Scholar] [CrossRef]
- Nguyen, D.; Khanal, S.K. A little breath of fresh air into an anaerobic system: How microaeration facilitates anaerobic digestion process. Biotechnol. Adv. 2018, 36, 1971–1983. [Google Scholar] [CrossRef]
- Itoh, Y.; Tada, K.; Kanno, T.; Horiuchi, J.I. Selective production of lactic acid in continuous anaerobic acidogenesis by extremely low pH operation. J. Biosci. Bioeng. 2012, 114, 537–539. [Google Scholar] [CrossRef]
- Park, B.H.; Kim, I.S.; Park, J.K.; Zhi, Z.; Lee, H.M.; Kwon, O.W.; Lee, B.C. Probiotic effect of Lactococcus lactis subsp. cremoris RPG-HL-0136 on intestinal mucosal immunity in mice. Appl. Biol. Chem. 2021, 64, 1–9. [Google Scholar]
- Yu, H.S.; Jang, H.J.; Lee, N.K.; Paik, H.D. Evaluation of the probiotic characteristics and prophylactic potential of Weissella cibaria strains isolated from kimchi. LWT-Food Sci. Technol. 2019, 112, 108229. [Google Scholar] [CrossRef]
- Cai, Y.M.; Benno, Y.; Ogawa, M.; Ohmomo, S.; Kumai, S.; Nakase, T. Influence of Lactobacillus spp. from an inoculant and of Weissella and Leuconostoc spp. from forage crops on silage fermentation. Appl. Environ. Microbiol. 1998, 64, 2982–2987. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
(a) SCOD, (b) soluble carbohydrates, and (c) lactic acid under different pHs.

Figure 2.
Amino acids during fermentation at pH 5.0 (The full name of amino acids in the legend is: Argnine (Arg); Lysine (Lys); Histidine (His); Phenylalanine (Phe); Tyrosine (Tyr); Leucine (Leu); Isoleucine (Ile); Methionine (Met); Valine (Val); Cysteine (Cys); Alanine (Ala); Glycine (Gly); Glutamic acid (Glu); Threonine (Thr); Aspartic acid (Asp)).
Figure 2.
Amino acids during fermentation at pH 5.0 (The full name of amino acids in the legend is: Argnine (Arg); Lysine (Lys); Histidine (His); Phenylalanine (Phe); Tyrosine (Tyr); Leucine (Leu); Isoleucine (Ile); Methionine (Met); Valine (Val); Cysteine (Cys); Alanine (Ala); Glycine (Gly); Glutamic acid (Glu); Threonine (Thr); Aspartic acid (Asp)).

Figure 3.
Box plot of LA concentration every 24 h in five groups (Control, Cys, Asp, Gly, and Val). * p < 0.05.
Figure 3.
Box plot of LA concentration every 24 h in five groups (Control, Cys, Asp, Gly, and Val). * p < 0.05.

Figure 4.
(a) SCOD, (b) soluble carbohydrates, and (c) hydrolysis rate for three groups (Control, Cys, and Val).
Figure 4.
(a) SCOD, (b) soluble carbohydrates, and (c) hydrolysis rate for three groups (Control, Cys, and Val).

Figure 5.
(a) Lactic acid concentration of three groups (Control, Cys, and Val); VFA concentration of (b) Control, (c) Cys, (d) Val.
Figure 5.
(a) Lactic acid concentration of three groups (Control, Cys, and Val); VFA concentration of (b) Control, (c) Cys, (d) Val.

Figure 6.
(a) The relative abundance at the genus level of microbial communities of the groups Control, Cys, and Val, and (b) a heat map of the correlation between microbial communities and fermentation products.
Figure 6.
(a) The relative abundance at the genus level of microbial communities of the groups Control, Cys, and Val, and (b) a heat map of the correlation between microbial communities and fermentation products.

Table 2.
Ingredient recipe for food waste.
| Ingredient | Mass ratio (%) |
|---|---|
| Rice | 21.8 |
| Noodles | 20.0 |
| Cabbage | 12.0 |
| Carrot | 5.6 |
| Onion | 5.0 |
| Potato | 12.0 |
| Egg | 4.6 |
| Tofu | 4.6 |
| Mushroom | 3.0 |
| Pork | 8.2 |
| Spices | 3.2 |
Table 3.
Characteristics of food waste: pH, total solids (TS), volatile solids (VS), total chemical oxygen demand (TCOD), and soluble chemical oxygen demand (SCOD).
Table 3.
Characteristics of food waste: pH, total solids (TS), volatile solids (VS), total chemical oxygen demand (TCOD), and soluble chemical oxygen demand (SCOD).
| Index | Food Waste |
|---|---|
| pH | 5.02 ± 0.2 |
| TS (g/L) | 82.92 ± 0.41 |
| VS (g/L) | 81.19 ± 0.35 |
| TCOD (g/L) | 110.40 ± 3.53 |
| SCOD (g/L) | 51.93 ± 0.33 |
Table 4.
Lactic acid yield, α-glucosidase, and lactate dehydrogenase activity of Control, Cys, and Val groups.
Table 4.
Lactic acid yield, α-glucosidase, and lactate dehydrogenase activity of Control, Cys, and Val groups.
| Control | Cys | Val | |
|---|---|---|---|
| α-glucosidase activity (U/g·VS) | 138.56 ± 2.9 | 169.36 ± 2.4 | 134.31 ± 2.4 |
| Lactic acid yield (g/g TS) | 0.50 ± 0.04 | 0.56 ± 0.04 | 0.41 ± 0.08 |
| lactate dehydrogenase activity (μmol/min/104 cell) | 1.76 ± 0.08 | 1.89 ± 0.05 | 1.28 ± 0.13 |
Note: α-glucosidase activity was the average of the previous five days, lactate dehydrogenase activity was measured on the 7th day.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhou, Y.; Zhang, X.; Wang, Y.; Liu, H. Mechanism and Effect of Amino Acids on Lactic Acid Production in Acidic Fermentation of Food Waste. Fermentation 2024, 10, 179. https://doi.org/10.3390/fermentation10040179
AMA Style
Zhou Y, Zhang X, Wang Y, Liu H. Mechanism and Effect of Amino Acids on Lactic Acid Production in Acidic Fermentation of Food Waste. Fermentation. 2024; 10(4):179. https://doi.org/10.3390/fermentation10040179
Chicago/Turabian StyleZhou, Yan, Xuedong Zhang, Yue Wang, and Hongbo Liu. 2024. "Mechanism and Effect of Amino Acids on Lactic Acid Production in Acidic Fermentation of Food Waste" Fermentation 10, no. 4: 179. https://doi.org/10.3390/fermentation10040179
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.