
The Short-Term Responses of Forest Soil Invertebrate Communities to Typhoon Disturbances
 and
and
Abstract
:1. Introduction
2. Materials and Methods
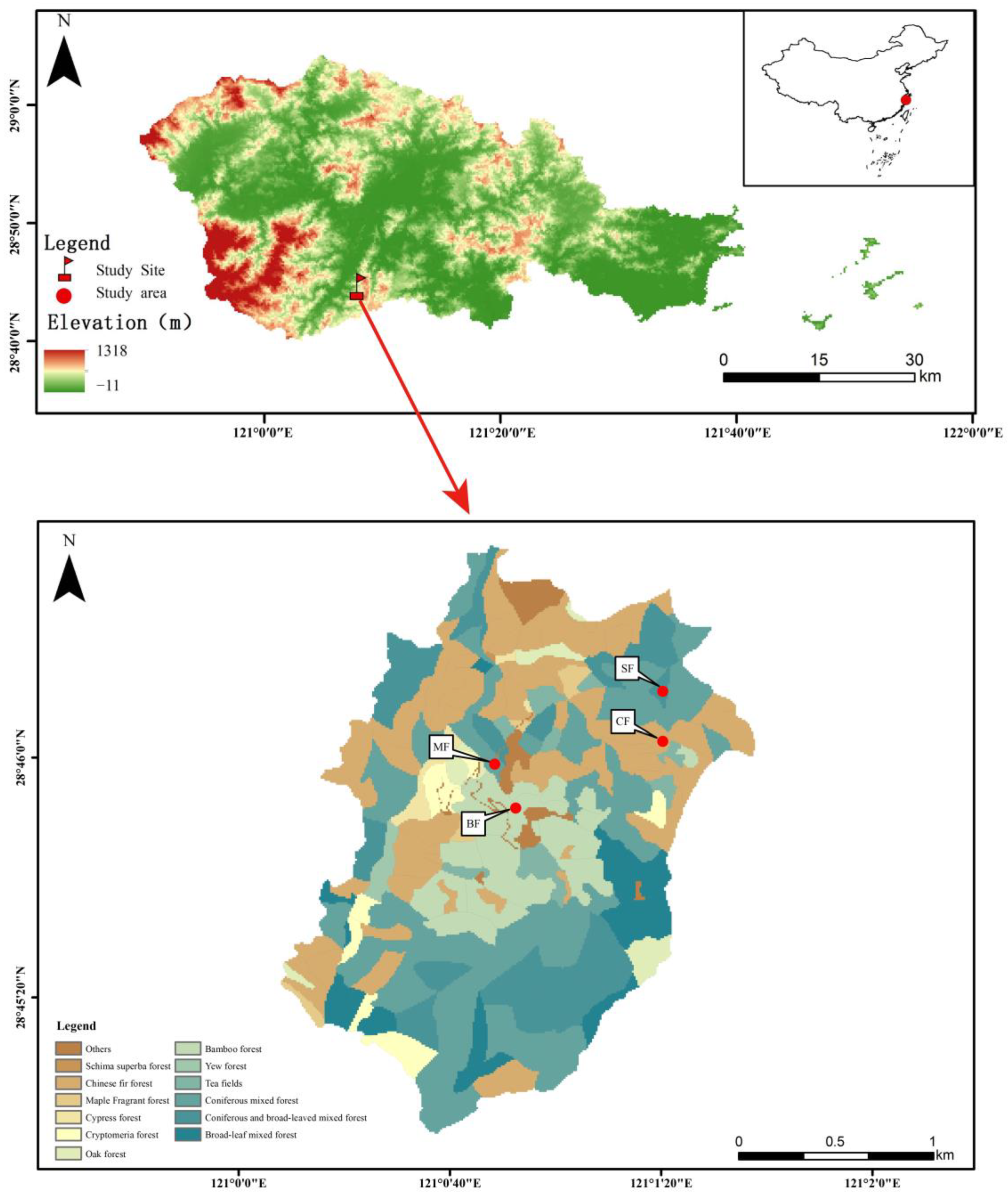
2.1. Site Description
2.2. Experimental Design
2.3. Sample Handling and Environmental Characterization
2.4. Data Analysis and Statistics
3. Results
3.1. Compositions and Densities of Soil Invertebrate Communities
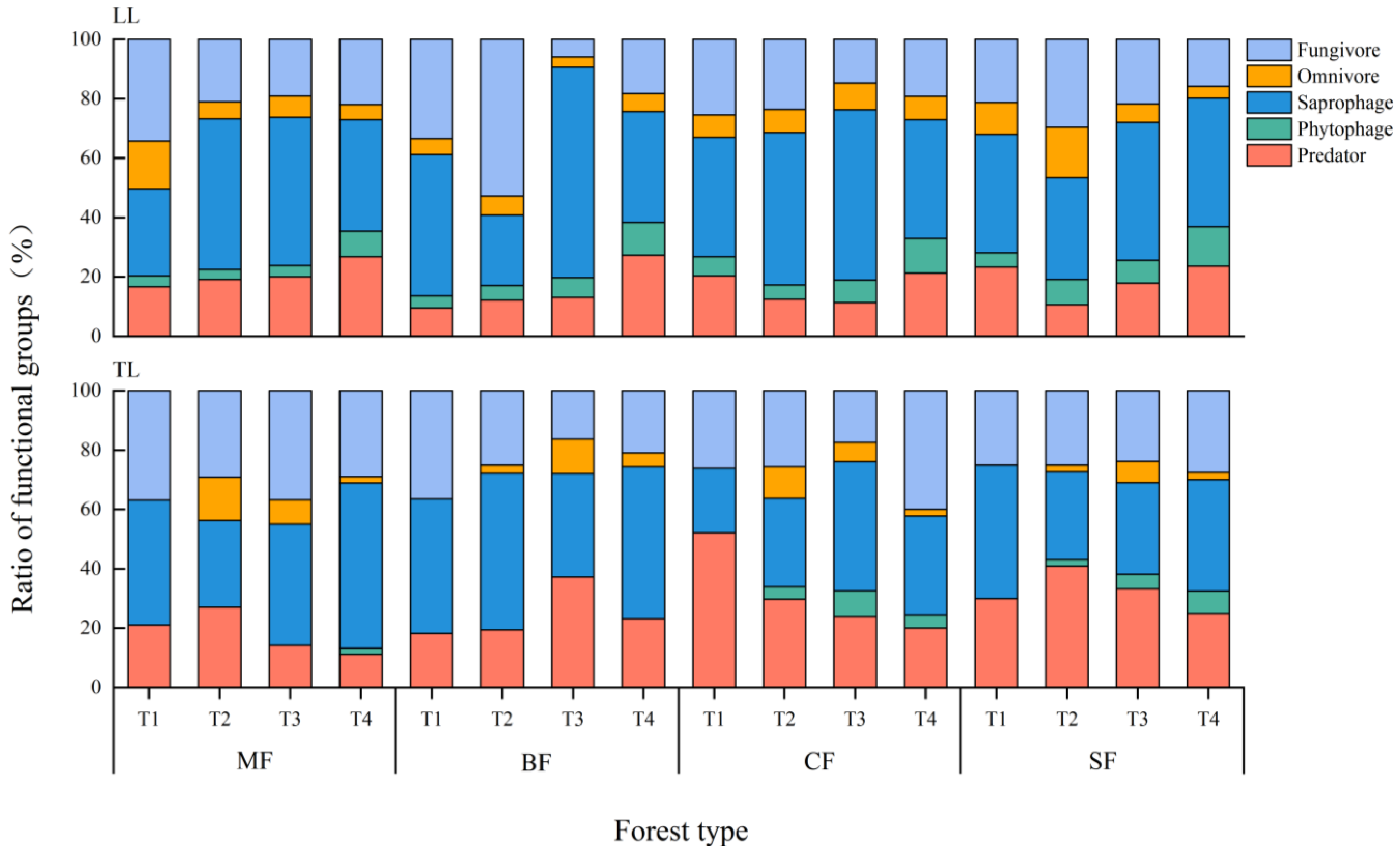
3.2. Soil Invertebrate Functional Groups and Detrital Food Webs
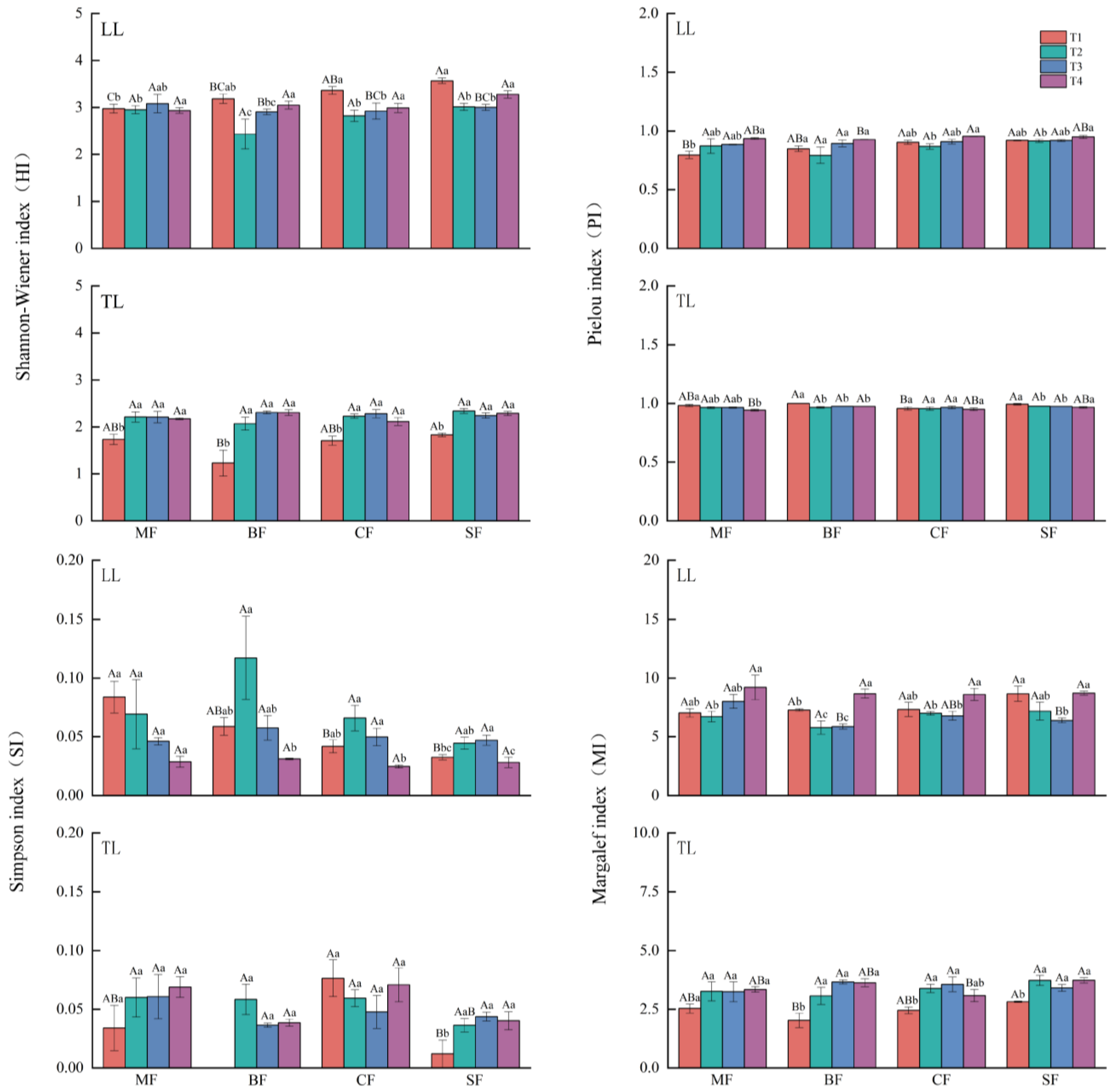
3.3. Soil Invertebrate Diversity
3.4. Relationships between Soil Invertebrate Communities and Soil Environmental Factors
4. Discussion
4.1. The Short-Term Response and Restoration of Soil Invertebrate Communities to Typhoon Disturbances
4.2. The Effects of Typhoon Disturbances on Detrital Food Webs
4.3. The Effects of Non-Disastrous Typhoon Disturbances on Soil Invertebrate Biodiversity
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gilliam, F.S. Impacts of tropical cyclones on longleaf pine ecosystems of Florida: Tropical cyclogenesis, landfall frequencies, and climate change. Front. Ecol. Evol. 2021, 9, 595791. [Google Scholar] [CrossRef]
- Gong, Y.; Staudhammer, C.L.; Kenney, G.; Wiesner, S.; Zhang, Y.; Starr, G. Vegetation structure drives forest phenological recovery after hurricane. Sci. Total Environ. 2021, 774, 145651. [Google Scholar] [CrossRef]
- Eppinga, M.B.; Pucko, C.A. The impact of hurricanes Irma and Maria on the forest ecosystems of Saba and St. Eustatius, northern Caribbean. Biotropica 2018, 50, 723–728. [Google Scholar] [CrossRef]
- Van Beusekom, A.E.; González, G.; Stankavich, S.; Zimmerman, J.K.; Ramírez, A. Understanding tropical forest abiotic response to hurricanes using experimental manipulations, field observations, and satellite data. Biogeosciences 2020, 17, 3149–3163. [Google Scholar] [CrossRef]
- Schowalter, T.D.; Willig, M.R.; Presley, S.J. Post-hurricane successional dynamics in abundance and diversity of canopy arthropods in a tropical rainforest. Environ. Entomol. 2017, 46, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Stork, G. How do beetle assemblages respond to cyclonic disturbance of a fragmented tropical rainforest landscape? Oecologia 2009, 161, 591–599. [Google Scholar]
- Liu, X.; Zeng, X.; Zou, X.; González, G.; Wang, C.; Yang, S. Litterfall production prior to and during Hurricanes Irma and Maria in four Puerto Rican forests. Forests 2018, 9, 367. [Google Scholar] [CrossRef]
- Jaramillo, V.J.; Martínez-Yrízar, A.; Maass, M.; Nava-Mendoza, M.; Castañeda-Gómez, L.; Ahedo-Hernández, R.; Araiza, S.; Verduzco, A. Hurricane impact on biogeochemical processes in a tropical dry forest in western Mexico. For. Ecol. Manag. 2018, 426, 72–80. [Google Scholar] [CrossRef]
- Wu, Z.; Li, Y.; Zhou, G.; Chen, B. Abnormal Litterfall and lts Ecological Significance. Sci. Silvae Sin. 2008, 44, 28–31. [Google Scholar]
- IPCC. Climate Change 2021: The Physical Science Basis; Cambridge University Press: London, UK, 2021; pp. 1–195. [Google Scholar]
- Lin, K.C.; Hamburg, S.P.; Wang, L.; Duh, C.T.; Huang, C.M.; Chang, C.T.; Lin, T.C. Impacts of increasing typhoons on the structure and function of a subtropical forest: Reflections of a changing climate. Sci. Rep. 2017, 7, 4911. [Google Scholar] [CrossRef]
- Chapman, E.L.; Chambers, J.Q.; Ribbeck, K.F.; Baker, D.B.; Tobler, M.A.; Zeng, H.; White, D.A. Hurricane Katrina impacts on forest trees of Louisiana’s Pearl River basin. For. Ecol. Manag. 2008, 256, 883–889. [Google Scholar] [CrossRef]
- Willig, M.R.; Camilo, G.R. The effect of Hurricane Hugo on six invertebrate species in the Luquillo Experimental Forest of Puerto Rico. Biotropica 1991, 23, 455–461. [Google Scholar] [CrossRef]
- Elmquist, D.C.; Kahl, K.B.; Johnson-Maynard, J.L.; Eigenbrode, S.D. Linking agricultural diversification practices, soil arthropod communities and soil health. J. Appl. Ecol. 2023, 60, 1952–1963. [Google Scholar] [CrossRef]
- Gergócs, V.; Flórián, N.; Tóth, Z.; Sipőcz, L.; Dombos, M. Detangling ecosystem services: Open-field manipulation of soil-dwelling microarthropods provides new opportunities to investigate their effects on nitrogen cycling. Ecol. Evol. 2022, 12, e9134. [Google Scholar] [CrossRef] [PubMed]
- Kataja-Aho, S.; Hannonen, P.; Liukkonen, T.; Rosten, H.; Haimi, J. The arthropod community of boreal Norway spruce forests responds variably to stump harvesting. For. Ecol. Manag. 2016, 371, 75–83. [Google Scholar] [CrossRef]
- Kudrin, A.; Perminova, E.; Taskaeva, A.; Ditts, A.; Konakova, T. A meta-analysis of the effects of harvesting on the abundance and richness of soil fauna in boreal and temperate forests. Forests 2023, 14, 923. [Google Scholar] [CrossRef]
- Peguero, G.; Folch, E.; Liu, L.; Ogaya, R.; Penuelas, J. Divergent effects of drought and nitrogen deposition on microbial and arthropod soil communities in a Mediterranean forest. Eur. J. Soil Biol. 2021, 103, 103275. [Google Scholar] [CrossRef]
- Çakır, M.; Akburak, S.; Makineci, E.; Bolat, F. Recovery of soil biological quality (QBS-ar) and soil microarthropod abundance following a prescribed fire in the Quercus frainetto forest. Appl. Soil Ecol. 2023, 184, 104768. [Google Scholar] [CrossRef]
- Schowalter, T. Insect responses to major landscape-level disturbance. Annu. Rev. Entomol. 2012, 57, 1–20. [Google Scholar] [CrossRef]
- Shiels, A.B.; González, G. Understanding the key mechanisms of tropical forest responses to canopy loss and biomass deposition from experimental hurricane effects. For. Ecol. Manag. 2014, 332, 1–10. [Google Scholar] [CrossRef]
- Bouget, C.; Duelli, P. The effects of windthrow on forest insect communities: A literature review. Biol. Conserv. 2004, 118, 281–299. [Google Scholar] [CrossRef]
- Cours, J.; Bouget, C.; Barsoum, N.; Horak, J.; Le Souchu, E.; Leverkus, A.B.; Pincebourde, S.; Thorn, S.; Salle, A. Surviving in Changing Forests: Abiotic Disturbance Legacy Effects on Arthropod Communities of Temperate Forests. Curr. For. Rep. 2023, 9, 189–218. [Google Scholar] [CrossRef]
- Foster, M.S.; Terborgh, J. Impact of a rare storm event on an Amazonian forest. Biotropica 1998, 30, 470–473. [Google Scholar] [CrossRef]
- Chen, W.; Liu, W.; Liang, H.; Jiang, M.; Dai, Z. Response of storm surge and M2 tide to typhoon speeds along coastal Zhejiang Province. Ocean Eng. 2023, 270, 113646. [Google Scholar] [CrossRef]
- Pan, Y.; Cao, A.; Wu, Y.; Lu, S.; Fan, L.; Li, P. On the Response of Zhejiang Coastal Waters to 12 Typhoons from 2011 to 2015. J. Mar. Sci. Eng. 2022, 10, 543. [Google Scholar] [CrossRef]
- Dong, J.; Huang, X. Typhoon track classification and analysis of rainstorm area landing on Zhejiang. J. Zhejiang Meteorol. 2019, 13–19. [Google Scholar]
- Wang, H.; Xu, X.; Yang, W.; Cao, R.; Wang, Z.; Zheng, B.; Lv, H.; Liu, T. The ecological stoichiometry of carbon, nitrogen and phosphorus in urban garden plants with different life forms and its response to typhoon Hagupit. Acta Ecol. Sin. 2021, 41, 8931–8938. [Google Scholar]
- Wang, Z.; Wang, X.; Shen, G. Effects of typhoon disturbance on the litter production in an evergreen broad-leaved forest in the Tiantong, Zhejiang. J. East China Norm. Univ. 2014, 2014, 79–89. [Google Scholar]
- Krab, E.J.; Cornelissen, J.; Berg, M.P.; Freckleton, R. A simple experimental set-up to disentangle the effects of altered temperature and moisture regimes on soil organisms. Methods Ecol. Evol. 2015, 6, 1159–1168. [Google Scholar] [CrossRef]
- Edwards, C.A. The assessment of populations of soil-inhabiting invertebrates. Agric. Ecosyst. Environ. 1991, 34, 145–176. [Google Scholar] [CrossRef]
- Yin, W. Subtropical Soil Animals of China; Science Press: Beijing, China, 1992. [Google Scholar]
- Yin, W. Pictorial Keys to Soil Animals of China; Science Press: Beijing, China, 1998. [Google Scholar]
- Lu, R. Analytical Methods of Soil Agricultural Chemistry; China Agricultural Science Press: Beijing, China, 1999. [Google Scholar]
- Jackson, M.L. Soil Chemical Analysis—Advanced Course; UW-Madison Libraries Parallel Press: Madison, WI, USA, 2005. [Google Scholar]
- Bao, S. Soil Agrochemical Analysis, 3rd ed.; China Agriculture Press: Beijing, China, 2000. [Google Scholar]
- Klotzbücher, T.; Kaiser, K.; Guggenberger, G.; Gatzek, C.; Kalbitz, K. A new conceptual model for the fate of lignin in decomposing plant litter. Ecology 2011, 92, 1052–1062. [Google Scholar] [CrossRef]
- Rowland, A.; Roberts, J. Lignin and cellulose fractionation in decomposition studies using acid-detergent fibre methods. Commun. Soil Sci. Plant Anal. 1994, 25, 269–277. [Google Scholar] [CrossRef]
- Deng, Y.; Bai, Y.; Cao, R.; Jiang, Y.; Wang, Z.; Li, F.; Gong, H.; Yang, W. Key drivers of soil arthropod community shift across a subalpine forest series vary greatly with litter and topsoil layers. Eur. J. Soil Biol. 2022, 111, 103421. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.; Wagner, H. Package “vegan”: Community Ecology package. Time Int. 2012, 1997, 15–17. [Google Scholar]
- Arbuckle, J.L. IBM SPSS Amos 20 User’s Guide; Amos Development Corporation, SPSS Inc.: Chicago, IL, USA, 2011; pp. 226–229. [Google Scholar]
- Schowalter, T.D.; Pandey, M.; Presley, S.J.; Willig, M.R.; Zimmerman, J.K. Arthropods are not declining but are responsive to disturbance in the Luquillo Experimental Forest, Puerto Rico. Proc. Natl. Acad. Sci. USA 2021, 118, e2002556117. [Google Scholar] [CrossRef] [PubMed]
- Angulo-Sandoval, P.; Fernández-Marín, H.; Zimmerman, J.; Alde, T. Changes in patterns of understory leaf phenology and herbivory following hurricane damage. Biotropica 2004, 36, 60–67. [Google Scholar] [CrossRef]
- Prather, C. Divergent responses of leaf herbivory to simulated hurricane effects in a rainforest understory. For. Ecol. Manag. 2014, 332, 87–92. [Google Scholar] [CrossRef]
- Novais, S.; Macedo-Reis, L.E.; Cristobal-Peréz, E.J.; Sánchez-Montoya, G.; Janda, M.; Neves, F.; Quesada, M. Positive effects of the catastrophic Hurricane Patricia on insect communities. Sci. Rep. 2018, 8, 15042. [Google Scholar] [CrossRef]
- Ding, Z.; Xu, G.; Zhang, Y.X.; Zhang, S.; Ma, K. Altitudinal patterns of dominant invertebrates in forest soil and litter are scale-different. Ecol. Inform. 2023, 77, 102238. [Google Scholar] [CrossRef]
- Fekete, I.; Varga, C.; Biro, B.; Toth, J.A.; Varbiro, G.; Lajtha, K.; Szabo, G.; Kotroczo, Z. The effects of litter production and litter depth on soil microclimate in a central european deciduous forest. Plant Soil 2016, 398, 291–300. [Google Scholar] [CrossRef]
- Canepuccia, A.D.; Farias, A.A.; Escalante, A.H.; Iribarne, O.; Novaro, A.; Isacch, J.P. Differential responses of marsh predators to rainfall-induced habitat loss and subsequent variations in prey availability. Can. J. Zool. 2008, 86, 407–418. [Google Scholar] [CrossRef]
- Alejandro, D.; Canepuccia; Cicchino, A.; Escalante, A.; Novaro, A.; Isacch, J.P. Differential responses of marsh arthropods to rainfall-induced habitat loss. Zool. Stud. 2009, 48, 174–183. [Google Scholar]
- Perry, K.I.; Sivakoff, F.S.; Wallin, K.F.; Wenzel, J.W.; Herms, D.A. Forest disturbance and arthropods: Small-scale canopy and understory disturbances alter movement of mobile arthropods. Ecosphere 2021, 12, e03771. [Google Scholar] [CrossRef]
- Dell, J.; O’Brien, J.; Doan, L.; Richards, L.; Dyer, L. An arthropod survival strategy in a frequently burned forest. Ecology 2017, 98, 2972–2974. [Google Scholar] [CrossRef]
- Seibold, S.; Bässler, C.; Brandl, R.; Gossner, M.M.; Thorn, S.; Ulyshen, M.D.; Müller, J. Experimental studies of dead-wood biodiversity—A review identifying global gaps in knowledge. Biol. Conserv. 2015, 191, 139–149. [Google Scholar] [CrossRef]
- Salamon, J.A.; Scheu, S.; Schaefer, M. The Collembola community of pure and mixed stands of beech (Fagus sylvatica) and spruce (Picea abies) of different age. Pedobiologia 2008, 51, 385–396. [Google Scholar] [CrossRef]
- Debnath, P.; Karmakar, K. Garlic mite, Aceria tulipae (Keifer) (Acari: Eriophyoidea)–a threat for garlic in West Bengal, India. Int. J. Acarol. 2013, 39, 89–96. [Google Scholar] [CrossRef]
- Amin, M.R.; Islam, M.A.; Suh, S.J.; Kwon, O.; Lee, K.Y. Relationship between abiotic factors and the incidence of sucking pests on rose plants. Entomol. Res. 2020, 50, 475–482. [Google Scholar] [CrossRef]
- Wermelinger, B.; Obrist, M.K.; Baur, H.; Jakoby, O.; Duelli, P. Synchronous rise and fall of bark beetle and parasitoid populations in windthrow areas. Agric. For. Entomol. 2013, 15, 301–309. [Google Scholar] [CrossRef]
- Wermelinger, B.; Moretti, M.; Duelli, P.; Lachat, T.; Pezzatti, G.B.; Obrist, M.K. Impact of windthrow and savage-logging on taxonomic and functional diversity of forest arthropods. For. Ecol. Manag. 2017, 391, 9–18. [Google Scholar] [CrossRef]
- Schowalter, T.D.; Ganio, L.M. Invertebrate communities in a tropical rain forest canopy in Puerto Rico following Hurricane Hugo. Ecol. Entomol. 2010, 24, 191–201. [Google Scholar] [CrossRef]
- Bloch, C.P.; Willig, M.R. Context-dependence of long-term responses of terrestrial gastropod populations to large-scale disturbance. J. Trop. Ecol. 2006, 22, 111–122. [Google Scholar] [CrossRef]
- Schowalter, T.D. Invertebrate community structure and herbivory in a tropical rain forest canopy in Puerto Rico following Hurricane Hugo. Biotropica 1994, 26, 312–319. [Google Scholar] [CrossRef]
- Torres, J.A. Lepidoptera outbreaks in response to successional changes after the passage of Hurricane Hugo in Puerto Rico. J. Trop. Ecol. 1992, 8, 285–298. [Google Scholar] [CrossRef]
- Fischer, C.; Gerstmeier, R.; Wagner, T.C. Seasonal and temporal patterns of rainfall shape arthropod community composition and multi-trophic interactions in an arid environment. Sci. Rep. 2022, 12, 3742. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forest Type | Slope Aspect | Slope (°) | Plant Composition |
|---|---|---|---|
| MF | SE | 15–20 | Cunninghamia lanceolata, Liquidambar formosana, Camellia sinensis |
| BF | NW | 20–25 | Phyllostachys pubescns, Camellia sinensis, Camellia cuspidate, Eurya japonica, Lilium speciosum, Dioscorea japonica |
| CF | NW | 20–25 | Cunninghami lanceolata, Schima superba, Lilium formosana, Symplocos setchuensis, Litsea rotundifolia |
| SF | NW | 15–20 | Bothrocaryum controversum, Lindera rubronervia, Camellia cuspidata, Pterocarya stenoptera, Cyclobalanopsis gracilis |
| Site | Factor | Shannon–WIENER Index | Pielou Index | Simpson Index | Margalef Index | Density | Group |
|---|---|---|---|---|---|---|---|
| Litter layer | Forest type | 3.964 * | 4.183 * | 3.535 | 2.378 | 89.497 *** | 3.552 * |
| Typhoon/Period | 24.562 *** | 6.839 ** | 8.21 *** | 14.822 *** | 305.944 *** | 22.349 *** | |
| Forest type ∗ Typhoon/Period | 3.085 ** | 1.182 | 1.667 | 1.53 | 1.344 | 1.622 | |
| Topsoil layer | Forest type | 2.363 | 7.107 *** | 7.108 *** | 1.684 | 4.36 * | 1.464 |
| Typhoon/Period | 34.976 *** | 6.697 ** | 3.573 * | 15.732 *** | 59.356 *** | 41.262 *** | |
| Forest type ∗ Typhoon/Period | 2.032 | 1.138 | 1.688 | 1.112 | 0.734 | 1.307 |
| Factor | Fungivores | Omnivores | Saprophages | Phytophages | Predators | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| r² | p | r² | p | r² | p | r² | p | r² | p | |
| Typhoon/Time | 0.07 | *** | 0.11 | *** | 0.05 | *** | 0.23 | *** | 0.05 | *** |
| Forests type | 0.04 | * | 0.04 | ns | 0.05 | *** | 0.11 | *** | 0.05 | *** |
| Layer | 0.19 | *** | 0.12 | *** | 0.20 | *** | 0.11 | *** | 0.17 | *** |
| Typhoon ∗ Forests type | 0.10 | ** | 0.12 | ns | 0.08 | * | 0.09 | ns | 0.09 | ** |
| Typhoon ∗ Layer | 0.06 | *** | 0.04 | * | 0.05 | *** | 0.05 | *** | 0.04 | *** |
| Forests type ∗ Layer | 0.03 | * | 0.05 | * | 0.04 | ** | 0.02 | ns | 0.05 | ** |
| Typhoon ∗ Forests type ∗ Layer | 0.09 | ** | 0.12 | *** | 0.09 | * | 0.03 | ns | 0.10 | *** |
| Litter Layer | Topsoil Layer | ||||
|---|---|---|---|---|---|
| Environmental Factor | Pseudo-F | p | Environmental Factor | Pseudo-F | p |
| AT | 0.24 | ** | ST | 0.26 | ** |
| TC | 0.18 | ** | AT | 0.10 | ** |
| TP | 0.06 | ** | TN | 0.04 | ** |
| TN | 0.03 | ns | TP | 0.01 | ns |
| Li | 0.03 | * | TC | 0.01 | ns |
| Ce | 0.02 | ns | SWC | 0.04 | ns |
| LWC | 0.01 | ns | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xuan, F.; Pan, X.; Qin, M.; Cao, R.; Shen, H.; Jiang, Y.; Hou, J.; Wang, Z.; Wang, Z.; Wu, Q.; et al. The Short-Term Responses of Forest Soil Invertebrate Communities to Typhoon Disturbances. Forests 2024, 15, 583. https://doi.org/10.3390/f15040583
Xuan F, Pan X, Qin M, Cao R, Shen H, Jiang Y, Hou J, Wang Z, Wang Z, Wu Q, et al. The Short-Term Responses of Forest Soil Invertebrate Communities to Typhoon Disturbances. Forests. 2024; 15(4):583. https://doi.org/10.3390/f15040583
Chicago/Turabian StyleXuan, Fei, Xuebiao Pan, Mei Qin, Rui Cao, Hongye Shen, Yurui Jiang, Jianfeng Hou, Zhihui Wang, Zhuang Wang, Qiqian Wu, and et al. 2024. "The Short-Term Responses of Forest Soil Invertebrate Communities to Typhoon Disturbances" Forests 15, no. 4: 583. https://doi.org/10.3390/f15040583