
Exceptionally Uniform Bat Assemblages across Different Forest Habitats Are Dominated by Single Hyperabundant Generalist Species
Abstract
:1. Introduction
2. Materials and Methods
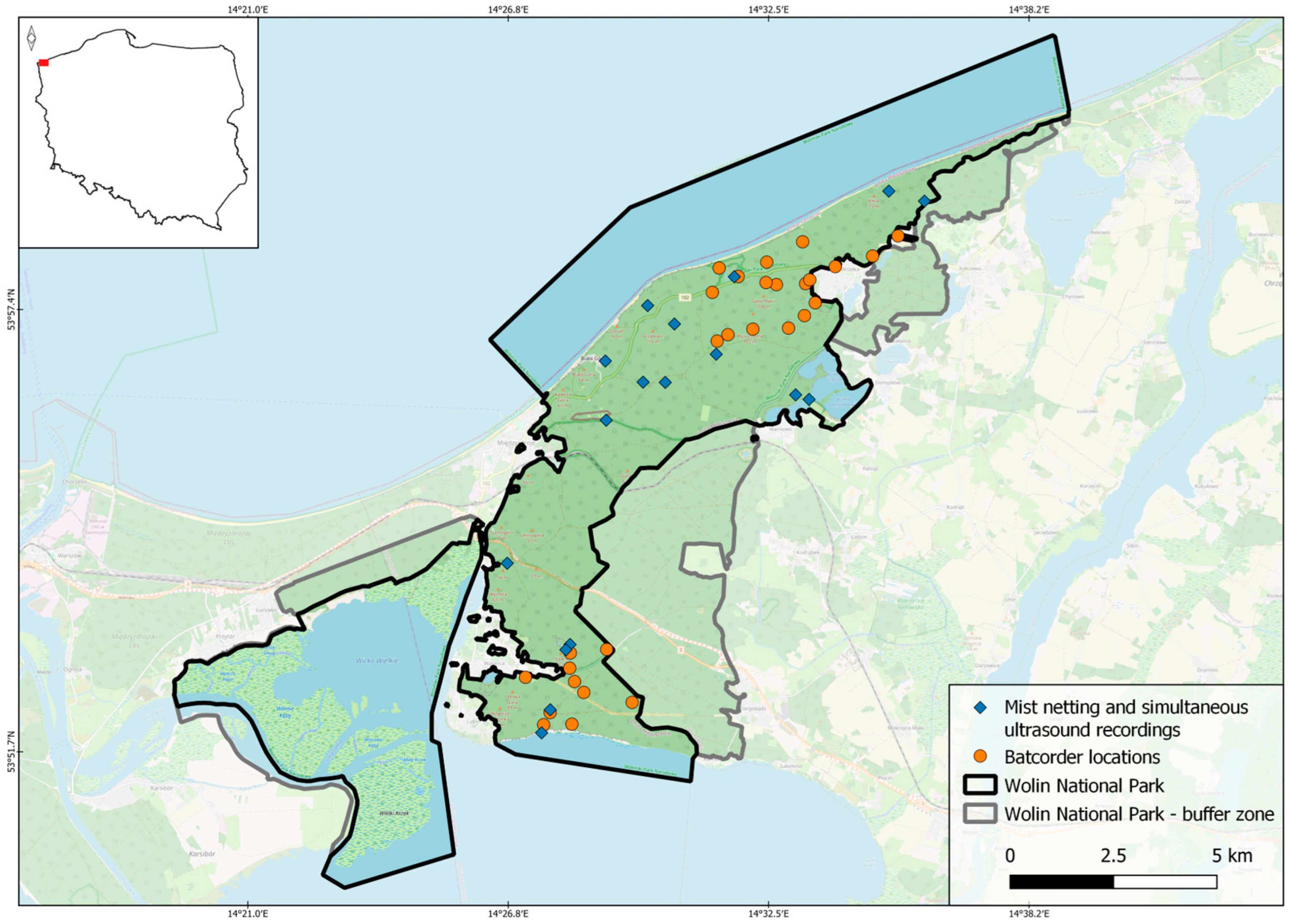
2.1. Study Area
2.2. Data Collection
2.3. Statistical Analysis
3. Results
3.1. General Composition of Bat Fauna
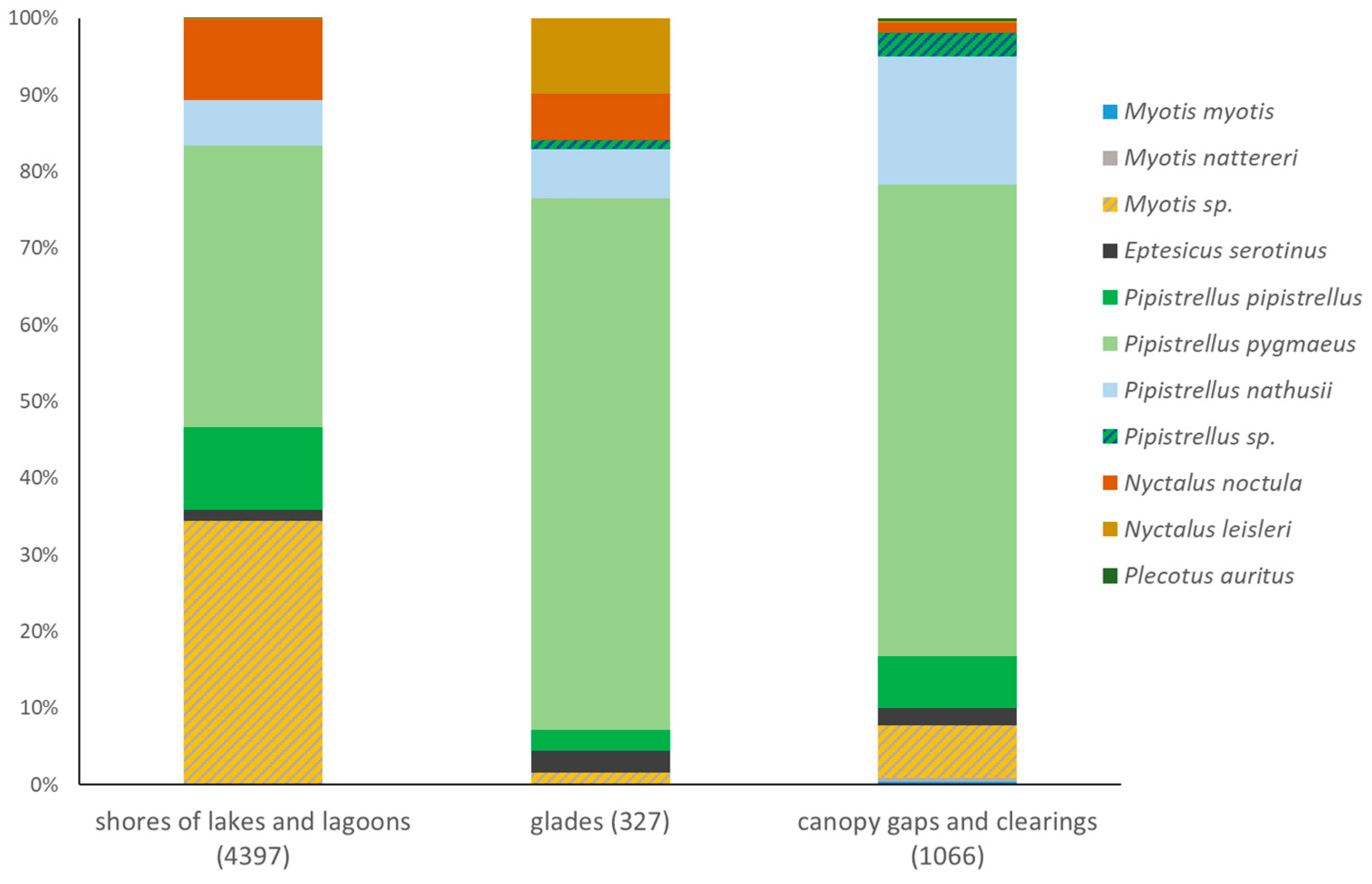
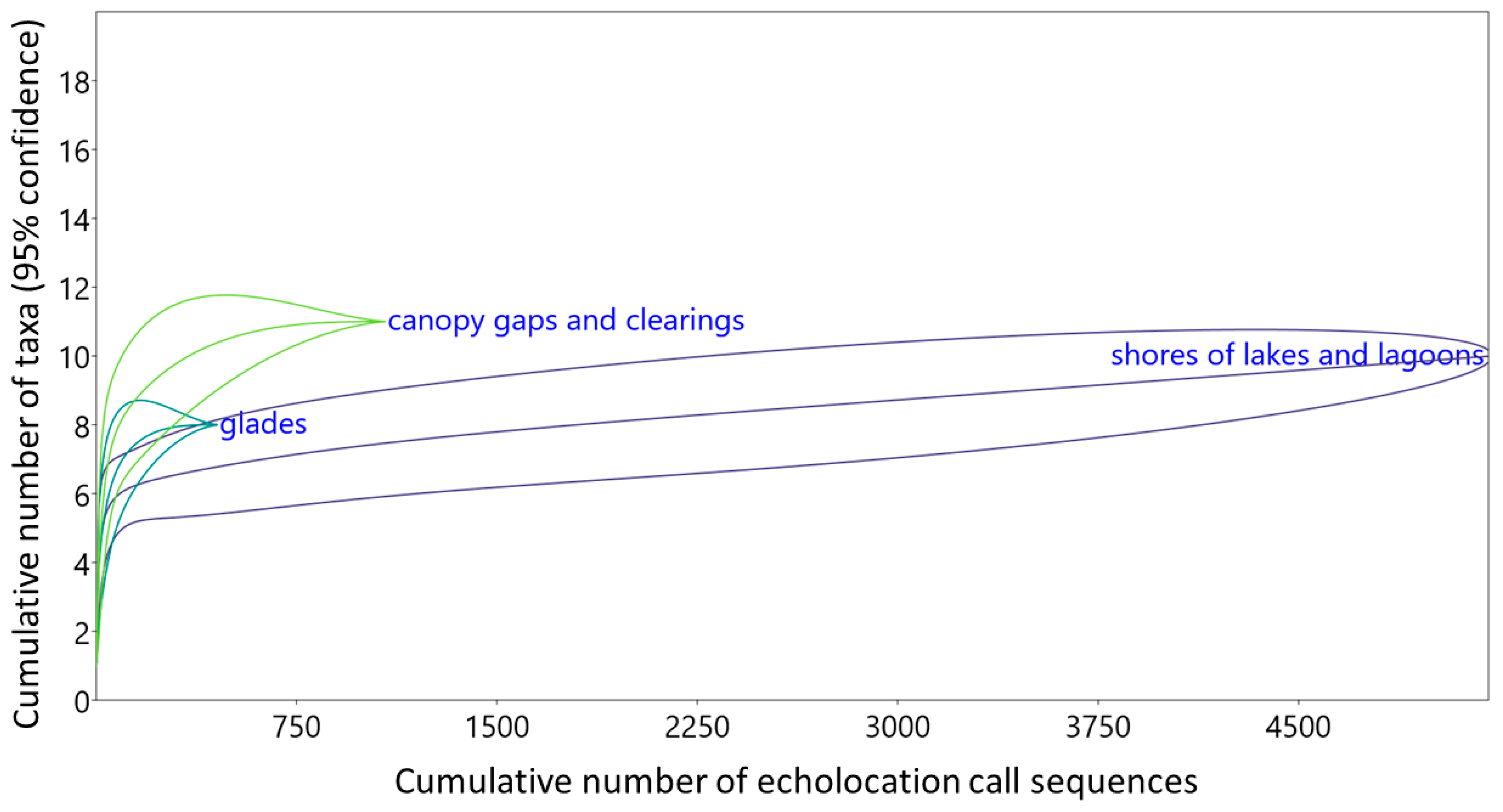
3.2. Species Composition Revealed Using Different Methods
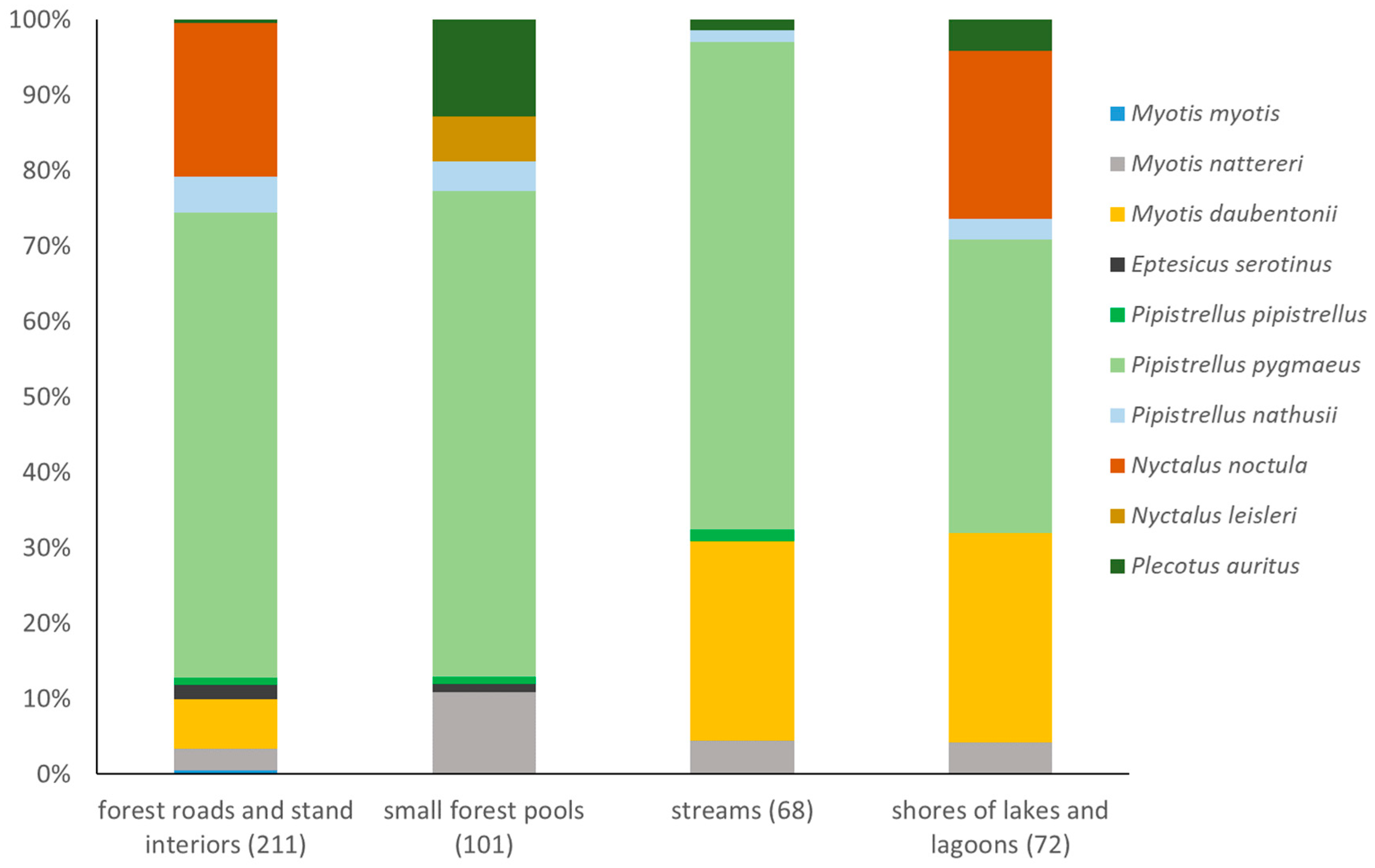
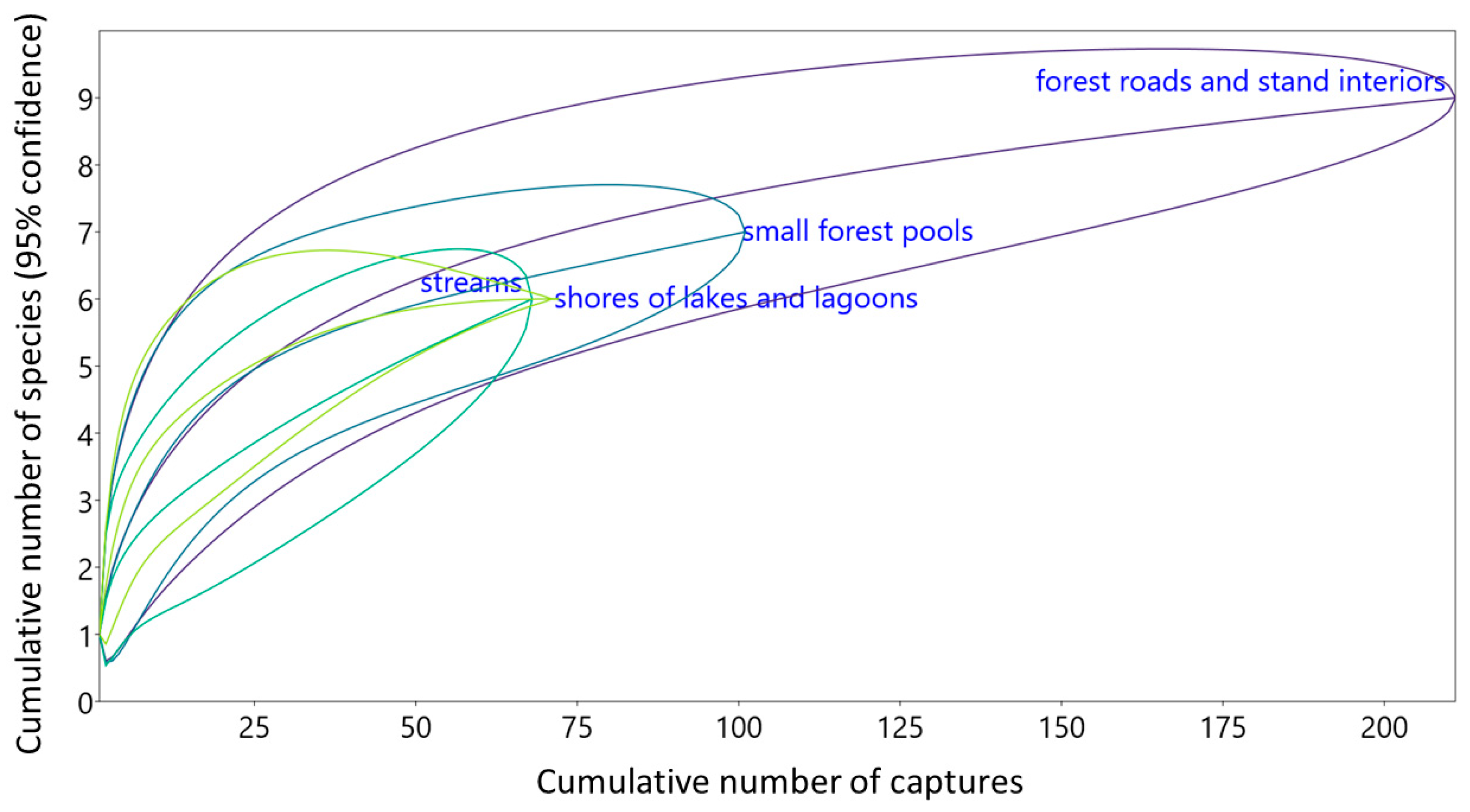
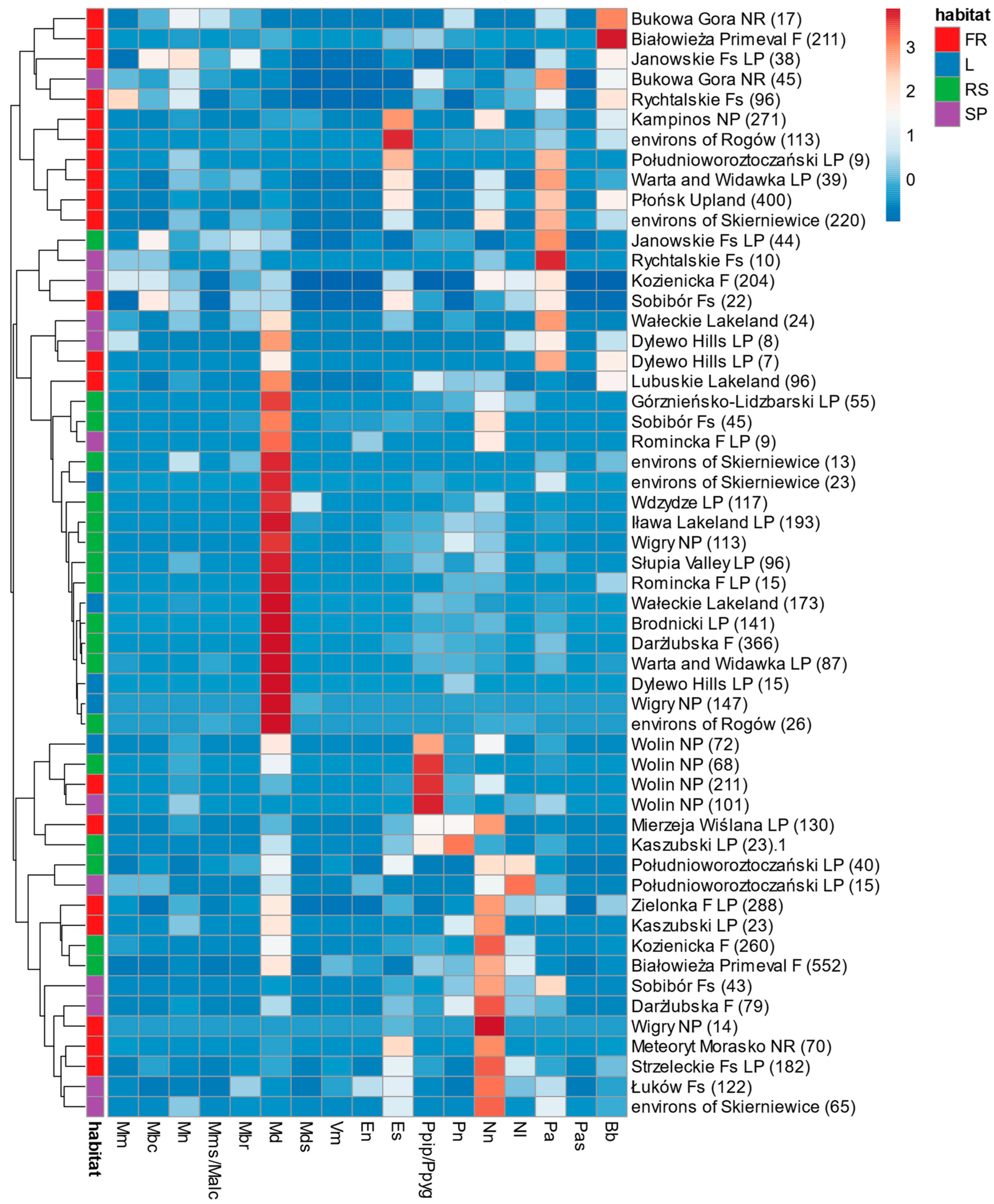
3.3. Structures of Bat Assemblages in Different Habitats
3.4. Comparison with the Other Polish Lowland Forests
4. Discussion
4.1. General Composition of Bat Fauna
4.2. Species Composition Revealed Using Different Methods
4.3. Uniformization of Bat Assemblages across Habitats—A Unique Feature of Wolin National Park?
4.4. Factors behind the Hyperabundance of Pipistrellus Pygmaeus and the Scarcity of Forest Specialists
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Law, B.; Park, K.J.; Lacki, M.J. Insectivorous Bats and Silviculture: Balancing Timber Production and Bat Conservation. In Bats in the Anthropocene: Conservation of Bats in a Changing World; Voigt, C., Kingston, T., Eds.; Springer: Cham, Germany, 2016; pp. 105–150. [Google Scholar] [CrossRef]
- Böhm, S.M.; Wells, K.; Kalko, E.K.V. Top-Down Control of Herbivory by Birds and Bats in the Canopy of Temperate Broad-Leaved Oaks (Quercus robur). PLoS ONE 2011, 6, e17ila857. [Google Scholar] [CrossRef]
- Beilke, E.A.; O’Keefe, J.M. Bats Reduce Insect Density and Defoliation in Temperate Forests: An Exclusion Experiment. Ecology 2023, 104, e3903. [Google Scholar] [CrossRef]
- Charbonnier, Y.; Barbaro, L.; Theillout, A.; Jactel, H. Numerical and Functional Responses of Forest Bats to a Major Insect Pest in Pine Plantations. PLoS ONE 2014, 9, e109488. [Google Scholar] [CrossRef]
- Ancillotto, L.; Rummo, R.; Agostinetto, G.; Tommasi, N.; Garonna, A.P.; Benedetta, F.; Bernardo, U.; Galimberti, A.; Russo, D. Bats as suppressors of agroforestry pests in beech forests. For. Ecol. Manag. 2022, 522, 120467. [Google Scholar] [CrossRef]
- Vaughan, N. The diets of British bats (Chiroptera). Mammal Rev. 1997, 27, 77–94. [Google Scholar] [CrossRef]
- Vesterinen, E.; Puisto, A.; Blomberg, A.; Lilley, T. Table for five, please: Dietary partitioning in boreal bats. Ecol. Evol. 2018, 8, 10914–10937. [Google Scholar] [CrossRef]
- Findley, J. Bats: A Community Perspective; Cambridge Studies in Ecology; Cambridge University Press: New York, NY, USA, 1995; 167p. [Google Scholar]
- Jung, K.; Kaiser, S.; Böhm, S.; Nieschulze, J.; Kalko, E.K.V. Moving in three dimensions: Effects of structural complexity on occurrence and activity of insectivorous bats in managed forest stands. J. Appl. Ecol. 2012, 49, 523–531. [Google Scholar] [CrossRef]
- Allegrini, C.; Korine, C.; Krasnov, B.R. Insectivorous Bats in Eastern Mediterranean Planted Pine Forests—Effects of Forest Structure on Foraging Activity, Diversity, and Implications for Management Practices. Forests 2022, 13, 1411. [Google Scholar] [CrossRef]
- Baagøe, H.J. The Scandinavian bat fauna: Adaptive wing morphology, and free flight in the field. In Recent Advances in the Study of Bats; Fenton, M.B., Racey, P.A., Rayner, J.M.V., Eds.; Cambridge University Press: New York, NY, USA, 1987; pp. 57–74. [Google Scholar]
- Neuweiler, G. Foraging Ecology and Audition in Echolocating Bats. Tree 1989, 6, 160–166. [Google Scholar] [CrossRef] [PubMed]
- Węgiel, A.; Grzywiński, W.; Ciechanowski, M. The foraging activity of bats in managed pine forests of different ages. Eur. J. For. Res. 2019, 138, 383–396. [Google Scholar] [CrossRef]
- Vlaschenko, A.; Kravchenko, K.; Yatsiuk, Y.; Hukov, V.; Kramer-Schadt, S.; Radchuk, V. Bat Assemblages Are Shaped by Land Cover Types and Forest Age: A Case Study from Eastern Ukraine. Forests 2022, 13, 1732. [Google Scholar] [CrossRef]
- Piksa, K.; Brzuskowski, T.; Zwijacz-Kozica, T. Distribution, Dominance Structure, Species Richness, and Diversity of Bats in Disturbed and Undisturbed Temperate Mountain Forests. Forests 2022, 13, 56. [Google Scholar] [CrossRef]
- Kaňuch, P.; Danko, Š.; Celuch, M.; Krištín, A.; Pjenčák, P.; Matis, Š.; Šmídt, J. Relating bat species presence to habitat features in natural forests of Slovakia (Central Europe). Mamm. Biol. 2008, 73, 147–155. [Google Scholar] [CrossRef]
- Węgiel, A.; Grzywiński, W.; Jaros, R.; Łacka, A.; Węgiel, J. Comparison of the Foraging Activity of Bats in Coniferous, Mixed, and Deciduous Managed Forests. Forests 2023, 14, 481. [Google Scholar] [CrossRef]
- Tillon, L.; Bouget, C.; Paillet, Y.; Aulagnier, S. How does deadwood structure temperate forest bat assemblages? Eur. J. For. Res. 2016, 135, 433–449. [Google Scholar] [CrossRef]
- Lesinski, G.; Kowalski, M.; Wojtowicz, B.; Gulatowska, J.; Lisowska, A. Bats on forest islands of different size in an agricultural landscape. Folia Zool. 2007, 56, 153–161. [Google Scholar]
- Ceľuch, M.; Kropil, R. Bats in a Carpathian beech-oak forest (Central Europe): Habitat use, foraging assemblages and activity patterns. Folia Zool. 2008, 57, 358–372. [Google Scholar]
- Humphrey, S.R. Nursery Roosts and Community Diversity of Nearctic Bats. J. Mammal. 1975, 56, 321–346. [Google Scholar] [CrossRef]
- Ruczyński, I.; Bogdanowicz, W. Roost Cavity Selection by Nyctalus noctula and N. leisleri (Vespertilionidae, Chiroptera) in Białowieża Primeval Forest, Eastern Poland. J. Mammal. 2005, 86, 921–930. [Google Scholar] [CrossRef]
- Lučan, R.K.; Andreas, M.; Benda, P.; Bartonička, T.; Březinová, T.; Hoffmannová, A.; Hulová, Š.; Hulva, P.; Neckářová, J.; Reiter, A.; et al. Alcathoe Bat (Myotis alcathoe) in the Czech Republic: Distributional Status, Roosting and Feeding Ecology. Acta Chiropterologica 2009, 11, 61–69. [Google Scholar] [CrossRef]
- Lesiński, G.; Olszewski, A.; Popczyk, B. Forest roads used by commuting and foraging bats in edge and interior zones. Pol. J. Ecol. 2011, 59, 611–616. [Google Scholar]
- Seibold, S.; Buchner, J.; Bässler, C.; Müller, J. Ponds in acidic mountains are more important for bats in providing drinking water than insect prey. J. Zool. 2013, 290, 302–308. [Google Scholar] [CrossRef]
- Russo, D.; Ancillotto, L.; Cistrone, L.; Libralato, N.; Domer, A.; Cohen, S.; Korine, C. Effects of artificial illumination on drinking bats: A field test in forest and desert habitats. Anim. Conserv. 2019, 22, 124–133. [Google Scholar] [CrossRef]
- Kowalski, M.; Krasnodębski, I.; Sachanowicz, K.; Dróżdż, R.; Wojtowicz, B. Skład gatunkowy, wybiórczość kryjówek i miejsc żerowania nietoperzy w Puszczy Kozienickiej. Kulon 1996, 1, 25–41. [Google Scholar]
- Ciechanowski, M. Community structure and activity of bats (Chiroptera) over different water bodies. Mamm. Biol. 2002, 67, 276–285. [Google Scholar] [CrossRef]
- Woliński Park Narodowy 2022. O Parku. Ogólnie—Położenie, Powierzchnia, Historia. Available online: https://wolinpn.pl/o-parku/ogolnie-polozenie-powierzchnia-historia/ (accessed on 18 November 2023).
- Woliński Park Narodowy 2022. Ekosystemy Leśne Wolińskiego Parku Narodowego. Statystyka. Available online: https://wolinpn.pl/przyroda-parku/statystyka/ (accessed on 18 November 2023).
- Dietz, C.; Kiefer, A. Bats of Britain and Europe; Bloomsbury Natural History: London, UK, 2016; 398p. [Google Scholar]
- Coleman, L.S.; Ford, W.M.; Dobony, C.A.; Britzke, E.R. A comparison of passive and active acoustic sampling for a bat community impacted by white-nose syndrome. J. Fish Wildl. Manag. 2014, 5, 217–226. [Google Scholar] [CrossRef]
- Barataud, M. Acoustic Ecology of European Bats. In Species Identification, Study of Their Habitats and Foraging Behaviour; Biotope—Muséum National D’historie Naturelle: Paris, France, 2014; 352p. [Google Scholar]
- Brabant, R.; Laurent, Y.; Dolap, U.; Degraer, S.; Poerink, B.J. Comparing the results of four widely used automated bat identification software programs to identify nine bat species in coastal Western Europe. Belg. J. Zool. 2018, 148, 119–128. [Google Scholar] [CrossRef]
- Parsons, K.N.; Jones, G. Acoustic identification of twelve species of echolocating bat by discriminant function analysis and artificial neural networks. J. Exp. Biol. 2000, 203 Pt 17, 2641–2656. [Google Scholar] [CrossRef] [PubMed]
- Hammer, R.; Harper, D.A.T.; Ryan, P.D. Past: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Häussler, U.; Nagel, A.; Braun, M.; Arnold, A. External characters discriminating sibling species of European pipistrelles, Pipistrellus pipistrellus (Schreber, 1774) and P. pygmaeus (Leach, 1825). Myotis 1999, 37, 27–40. [Google Scholar]
- Helversen, O.V.; Holderied, M. Zur Unterscheidung von Zwergfledermaus (Pipistrellus pipistrellus) und Mückenfledermaus (Pipistrellus mediterraneus/pygmaeus) im Feld. Nyctalus 2003, 8, 420–426. [Google Scholar]
- Metsalu, T.; Vilo, J. ClustVis: A web tool for visualizing clustering of multivariate data using Principal Component Analysis and heatmap. Nucleic Acids Res. 2015, 43, 566–570. [Google Scholar] [CrossRef]
- ClustVis. Available online: https://biit.cs.ut.ee/clustvis/ (accessed on 23 October 2023).
- Sachanowicz, K.; Ciechanowski, M.; Piksa, K. 2006. Distribution patterns, species richness and status of bats in Poland. Vespertilio 2006, 9–10, 151–173. [Google Scholar]
- Bidziński, K.; Ciechanowski, M.; Jankowska-Jarek, M.; Wikar, Z. Department of Vertebrate Ecology and Zoology. University of Gdańsk: Gdańsk, Poland, 2023; unpublished data. [Google Scholar]
- Gaisler, J.; Hanák, V.; Hanzal, V.; Jarský, V. Results of bat banding in the Czech and Slovak Republics, 1948–2000. Vespertilio 2003, 7, 3–61. [Google Scholar]
- Hutterer, R.; Ivanova, T.; Meyer-Cords, C.H.; Rodrigues, L. Bat Migration in Europe. A Review of Banding Data and Literature; Federal Agency for Nature Conservation: Bonn, Germany, 2005; pp. 1–176. [Google Scholar]
- Zyska, W.; Dylawerski, M.; Mackiewicz, R.; Skórkowski, R.; Szwarc, M.; Walczak, M.; Zyska, M.; Zyska, W. Występowanie nietoperzy w siedliskach leśnych Wolińskiego Parku Narodowego w latach 2018–2019 w tle wyników ocen prowadzonych w ostatnich 30 latach. Przegląd Przyr. 2020, 31, 64–81. [Google Scholar]
- Ruprecht, A.L. Bats (Chiroptera). In Atlas of Polish Mammals; Pucek, Z., Raczyński, J., Eds.; PWN: Warsaw, Poland, 1983; pp. 62–82, maps 27–67. [Google Scholar]
- Schaub, A.; Schnitzler, H.U. Echolocation behavior of the bat Vespertilio murinus reveals the border between the habitat types “edge” and “open space”. Behav. Ecol. Sociobiol. 2007, 61, 513–523. [Google Scholar] [CrossRef]
- Zingg, P.E. Akustische Artidentifikation von Fledermäusen (Mammalia: Chiroptera) in der Schweiz. Rev. Suisse De Zool. 1990, 97, 263–294. [Google Scholar] [CrossRef]
- Baagøe, H.J. Danish Bats (Mammalia: Chiroptera): Atlas and Analysis of Distribution, Occurrence and Abundance. Steenstrupia; Zoological Museum, University of Copenhagen: Copenhagen, Denmark, 2001; Volume 26, pp. 1–117. [Google Scholar]
- Ciechanowski, M.; Jakusz-Gostomska, A.; Żmihorski, M. Empty in summer, crowded during migration? Structure of assemblage, distribution pattern and habitat use by bats (Chiroptera: Vespertilionidae) in a narrow, marine peninsula. Mammal Res. 2016, 61, 45–55. [Google Scholar] [CrossRef]
- Keišs, O.; Spalis, D.; Pētersons, G. Funnel trap as a method for capture migrating bats in Pape, Latvia. Environ. Exp. Biol. 2021, 19, 7–10. [Google Scholar] [CrossRef]
- Ciechanowski, M.; Szkudlarek, R. Pierwsze stwierdzenie mroczka pozłocistego Eptesicus nilssonii (Keyserling & Blasius, 1839) na Pomorzu. Nietoperze 2003, 4, 105–107. [Google Scholar]
- Ciechanowski, M.; Sachanowicz, K.; Kokurewicz, T. Rare or underestimated?—The distribution and abundance of the pond bat (Myotis dasycneme) in Poland. Lutra 2007, 50, 107–134. [Google Scholar]
- Ciechanowski, M. Myotis Bechsteinii (Kuhl, 1817). Atlas Ssaków Polski. Instytut Ochrony Przyrody PAN, Kraków, Poland, 2021. Available online: https://www.iop.krakow.pl/Ssaki/gatunek/160 (accessed on 6 November 2023).
- Murray, K.; Britzke, E.; Hadley, B.; Robbins, L. Surveying bat communities: A comparison between mist nets and the Anabat II bat detector system. Acta Chiropterologica 1999, 1, 105–112. [Google Scholar]
- Flaquer, C.; Torre, I.; Arrizabalaga, A. Comparison of Sampling Methods for Inventory of Bat Communities. J. Mammal. 2007, 88, 526–533. [Google Scholar] [CrossRef]
- MacSwiney G, M.C.; Clarke, F.M.; Racey, P.A. What you see is not what you get: The role of ultrasonic detectors in increasing inventory completeness in Neotropical bat assemblages. J. Appl. Ecol. 2008, 45, 1364–1371. [Google Scholar] [CrossRef]
- Rachwald, A.; Boratyński, P.; Nowakowski, W.K. Species composition and night-time activity of bats flying over rivers in Białowieża Primeval Forest (Eastern Poland). Acta Theriol. 2001, 46, 235–242. [Google Scholar] [CrossRef]
- Ciechanowski, M.; Zając, T.; Biłas, A.; Dunajski, R. Spatiotemporal variation in activity of bat species differing in hunting tactics: Effects of weather, moonlight, food abundance, and structural clutter. Can. J. Zool. 2007, 85, 1249–1263. [Google Scholar] [CrossRef]
- Chaves-Ramírez, S.; Castillo-Salazar, C.; Sánchez-Chavarría, M.; Solís-Hernández, H.; Chaverri, G. Comparing the efficiency of monofilament and regular nets for capturing bats. R. Soc. Open Sci. 2021, 8, 211404. [Google Scholar] [CrossRef]
- Siemers, B.; Schnitzler, H.U. Echolocation signals reflect niche differentiation in five sympatric congeneric bat species. Nature 2004, 429, 657–661. [Google Scholar] [CrossRef]
- Vaughan, N.; Jones, G.; Harris, S. Identification of British bat species by multivariate analysis of echolocation call parameters. Bioacoustics 1997, 7, 189–207. [Google Scholar] [CrossRef]
- Walters, C.L.; Freeman, R.; Collen, A.; Dietz, C.; Brock Fenton, M.; Jones, G.; Obrist, M.K.; Puechmaille, S.J.; Sattler, T.; Siemers, B.M.; et al. A continental-scale tool for acoustic identification of European bats. J. Appl. Ecol. 2012, 49, 1064–1074. [Google Scholar] [CrossRef]
- Fenton, M.B. Choosing the ‘correct’ bat detector. Acta Chiropterologica 2000, 2, 215–224. [Google Scholar]
- Adams, A.M.; Jantzen, M.K.; Hamilton, R.M.; Fenton, M.B. Do you hear what I hear? Implications of detector selection for acoustic monitoring of bats. Methods Ecol. Evol. 2012, 3, 992–998. [Google Scholar] [CrossRef]
- Rachwald, A.; Wodecka, K.; Malzahn, E.; Kluziński, L. Bat activity in coniferous forest areas and the impact of air pollution. Mammalia 2004, 68, 445–453. [Google Scholar] [CrossRef]
- Ciechanowski, M. Habitat preferences of bats in anthropogenically altered, mosaic landscapes of northern Poland. Eur. J. Wildl. Res. 2015, 61, 415–428. [Google Scholar] [CrossRef]
- Rachwald, A.; Boratyński, J.S.; Krawczyk, J.; Szurlej, M.; Nowakowski, W.K. Natural and anthropogenic factors influencing the bat community in commercial tree stands in a temperate lowland forest of natural origin (Białowieża Forest). For. Ecol. Manag. 2021, 479, 118544. [Google Scholar] [CrossRef]
- Dietz, M.; Encarnação, J.A.; Kalko, E.K.V. Small scale distribution patterns of female and male Daubenton’s bats (Myotis daubentonii). Acta Chiropterologica 2006, 8, 403–415. [Google Scholar] [CrossRef]
- Kurek, K.; Tołkacz, K.; Mysłajek, R. Low abundance of the whiskered bat Myotis mystacinus (Kuhl, 1817) in Poland—Consequence of competition with pipistrelle bats? Appl. Ecol. Environ. Res. 2017, 15, 241–248. [Google Scholar] [CrossRef]
- Rachwald, A.; Bradford, T.; Borowski, Z.; Racey, P.A. Habitat Preferences of Soprano Pipistrelle Pipistrellus pygmaeus (Leach, 1825) and Common Pipistrelle Pipistrellus pipistrellus (Schreber, 1774) in Two Different Woodlands in North East Scotland. Zool. Stud. 2016, 55, e22. [Google Scholar] [CrossRef]
- Rachwald, A. Występowanie mopka zachodniego Barbastella barbastellus (Schreber, 1744) w Puszczy Białowieskiej na tle innych gatunków nietoperz. In Inwentaryzacja Wybranych Elementów Przyrodniczych i Kulturowych Puszczy Białowieskiej; Matuszkiewicz, J.M., Tabor, J., Eds.; Instytut Badawczy Leśnictwa: Sękocin Stary, Poland, 2022; pp. 713–737. [Google Scholar] [CrossRef]
- Gaisler, J. The r-K selection model and life history strategies in bats. In European Bat Research; Hanak, V., Horácek, I., Gaisler, J., Eds.; Charles University Press: Prague, Czech Republic, 1989; pp. 117–124. [Google Scholar]
- Arlettaz, R.; Godat, S.; Meyer, H. Competition for food by expanding pipistrelle bat populations (Pipistrellus pipistrellus) might contribute to the decline of lesser horseshoe bats (Rhinolophus hipposideros). Biol. Conserv. 2000, 93, 55–60. [Google Scholar] [CrossRef]
- Mickleburgh, S. Distribution and status of bats in the London area. In European Bat Research; Hanak, V., Horácek, I., Gaisler, J., Eds.; Charles University Press: Prague, Czech Republic, 1989; pp. 327–329. [Google Scholar]
- Glendell, M.; Vaughan, N. Foraging activity of bats in historic landscape parks in relation to habitat composition and park management. Anim. Conserv. 2002, 5, 309–316. [Google Scholar] [CrossRef]
- Vaughan, N.; Jones, G.; Harris, S. Habitat Use by Bats (Chiroptera) Assessed by Means of a Broad-Band Acoustic Method. J. Appl. Ecol. 1997, 34, 716–730. [Google Scholar] [CrossRef]
- Russ, J.M.; Montgomery, W.I. Habitat associations of bats in Northern Ireland: Implications for conservation. Biol. Conserv. 2002, 108, 49–58. [Google Scholar] [CrossRef]
- Sattler, T.; Bontadina, F.; Hirzel, A.H.; Arlettaz, R. Ecological niche modelling of two cryptic bat species calls for a reassessment of their conservation status. J. Appl. Ecol. 2007, 44, 1188–1199. [Google Scholar] [CrossRef]
- Stone, E.L.; Zeale, M.R.K.; Newson, S.E.; Browne, W.J.; Harris, S.; Jones, G. Managing conflict between bats and humans: The response of soprano pipistrelles (Pipistrellus pygmaeus) to exclusion from roosts in houses. PLoS ONE 2015, 10, e0131825. [Google Scholar] [CrossRef] [PubMed]
- Michaelsen, T.C.; Jensen, K.H.; Högstedt, G. Roost Site Selection in Pregnant and Lactating Soprano Pipistrelles (Pipistrellus pygmaeus Leach, 1825) at the Species Northern Extreme: The Importance of Warm and Safe Roosts. Acta Chiropterologica 2014, 16, 349–357. [Google Scholar] [CrossRef]
- Dietz, M.; Brombacher, M.; Erasmy, M.; Fenchuk, V.; Simon, O. Bat Community and Roost Site Selection of Tree-Dwelling Bats in a Well-Preserved European Lowland Forest. Acta Chiropterologica 2018, 20, 117–127. [Google Scholar] [CrossRef]
- Bashta, A.-T. Bridge structures and habitats of bats (Chiroptera): Species and spatial diversity. Theriol. Ukr. 2022, 24, 86–103. [Google Scholar] [CrossRef]
- Jenkins, E.V.; Laine, T.; Morgan, S.E.; Cole, K.R.; Speakman, J.R. Roost selection in the pipistrelle bat, Pipistrellus pipistrellus (Chiroptera: Vespertilionidae), in northeast Scotland. Anim. Behav. 1998, 56, 909–917. [Google Scholar] [CrossRef] [PubMed]
- Barlow, K.E.; Jones, G. Roosts, echolocation calls and wing morphology of two phonic types of Pipistrellus pipistrellus. Z. Säugetierkunde 1999, 64, 257–268. [Google Scholar]
- Kirkpatrick, L.; Graham, J.; McGregor, S.; Munro, L.; Scoarize, M. Flexible foraging strategies in Pipistrellus pygmaeus in response to abundant but ephemeral prey. PLoS ONE 2018, 13, e0204511. [Google Scholar] [CrossRef]
- Carr, J.F.; Hiltunen, J.K. Changes in the Bottom Fauna of Western Lake Erie from 1930 to 1961. Limnol. Oceanogr. 2003, 10, 551–569. [Google Scholar] [CrossRef]
- Rachwald, A.; Ciesielski, M.; Szurlej, M.; Żmihorski, M. Following the damage: Increasing western barbastelle bat activity in bark beetle infested stands in Białowieża Primeval forest. For. Ecol. Manag. 2022, 503, 119803. [Google Scholar] [CrossRef]
- Russo, D.; Cistrone, L.; Jones, G.; Mazzoleni, S. Roost selection by barbastelle bats (Barbastella barbastellus, Chiroptera: Vespertilionidae) in beech woodlands of central Italy: Consequences for conservation. Biol. Conserv. 2004, 117, 73–81. [Google Scholar] [CrossRef]
- Cel’uch, M.; Danko, S.; Kaňuch, P. On urbanisation of Nyctalus noctula and Pipistrellus pygmaeus in Slovakia. Vespertilio 2006, 9–10, 219–221. [Google Scholar]
- Friedland, R.; Schernewski, G.; Gräwe, U.; Greipsland, I.; Palazzo, D.; Pastuszak, M. Managing Eutrophication in the Szczecin (Oder) Lagoon-Development, Present State and Future Perspectives. Front. Mar. Sci. 2019, 5, 521. [Google Scholar] [CrossRef]
- Andreas, M.; Reiter, A.; Benda, P. Prey Selection and Seasonal Diet Changes in the Western Barbastelle Bat (Barbastella barbastellus). Acta Chiropterologica 2012, 14, 81–92. [Google Scholar] [CrossRef]
- Moore, J.H.; Gibson, L.; Amir, Z.; Chanthorn, W.; Ahmad, A.H.; Jansen, P.A.; Mendes, C.P.; Onuma, M.; Peres, C.A.; Luskin, M.S. The rise of hyperabundant native generalists threatens both humans and nature. Biol. Rev. 2023, 98, 1829–1844. [Google Scholar] [CrossRef] [PubMed]
- Szyp, E. Nietoperze Brodnickiego Parku Krajobrazowego. Master’s Thesis, Department of Vertebrate Zoology, Institute of Biology, Nicolaus Copernicus University, Toruń, Poland, 1996. [Google Scholar]
- Ciechanowski, M.; Koziróg, L.; Dariusz, J.; Przesmycka, A.; Świątkowska, A.; Kisicka, I.; Kasprzyk, K. Bat fauna of the Iława Lakeland Landscape Park (northern Poland). Myotis 2002, 40, 33–45. [Google Scholar]
- Bugajna, B. Wstępne badania nad nietoperzami (Chiroptera) rezerwatu „Meteoryt Morasko”. Rocz. Nauk. Pol. Tow. Ochr. Przyr. Salamandra 1996, 1, 217–218. [Google Scholar]
- Ciechanowski, M.; Czablewska, A.; Mączyńska, M.; Narczyński, T.; Przesmycka, A.; Zapart, A.; Jarzembowski, T.; Rachwald, A. Nietoperze (Chiroptera) Parku Krajobrazowego „Mierzeja Wiślana”. Nietoperze 2008, 9, 203–224. [Google Scholar]
- Piskorski, M.; Urban, P. Nietoperze Południoworoztoczańskiego Parku Krajobrazowego. Nietoperze 2003, 4, 21–25. [Google Scholar]
- Sachanowicz, K.; Krasnodębski, I. Skład gatunkowy i antropogeniczne kryjówki nietoperzy w Lasach Łukowskich. Nietoperze 2003, 4, 27–38. [Google Scholar]
- Ciechanowski, M. Chiropterofauna Puszczy Darżlubskiej. Nietoperze 2003, 4, 45–59. [Google Scholar]
- Ciechanowski, M. Struktura Przestrzenna Zespołu i Dynamika Aktywności Nietoperzy (Chiroptera) w Krajobrazie Leśno-rolniczym Północnej Polski. Ph.D. Thesis, Department of Vertebrate Ecology and Zoology, University of Gdańsk, Gdańsk, Poland, 2005. [Google Scholar]
- Ignaczak, M. Nietoperze rezerwatu “Bukowa Góra”. Nietoperze 2003, 4, 101–102. [Google Scholar]
- Ciechanowski, M.; Duriasz, J. Nietoperze (Chiroptera) Parku Krajobrazowego Wzgórz Dylewskich. Nietoperze 2005, 6, 25–36. [Google Scholar]
- Ciechanowski, M.; Anikowska, U.; Nalewaja, A.; Przesmycka, A.; Biała, A. Nietoperze (Chiroptera) Parku Krajobrazowego “Dolina Słupi”. Nietoperze 2006, 7, 19–37. [Google Scholar]
- Lesiński, G.; Gulatowska, J.; Kowalski, M.; Fuszara, E.; Fuszara, M.; Wojtowicz, B. Nietoperze Wysoczyzny Płońskiej. Nietoperze 2006, 7, 39–55. [Google Scholar]
- Piskorski, M. Fauna nietoperzy Parku Krajobrazowego Lasy Janowskie. Nietoperze 2007, 8, 3–11. [Google Scholar]
- Piskorski, M.; Gwardjan, M.; Kowalski, M.; Wojtowicz, B.; Urban, M.; Bochen, R. Fauna nietoperzy Parku Krajobrazowego Lasy Strzeleckie. Nietoperze 2009, 10, 15–22. [Google Scholar]
- Łochyński, M.; Grzywiński, W. Nietoperze Parku Krajobrazowego Puszcza Zielonka. Nietoperze 2009, 10, 23–35. [Google Scholar]
- Piskorski, M. Fauna nietoperzy Lasów Sobiborskich. Nietoperze 2008, 9, 3–17. [Google Scholar]
- Kmiecik, A.; Kmiecik, P.; Grzywiński, W. 2010. Chiropterofauna Wigierskiego Parku Narodowego. Nietoperze 2010, 11, 11–29. [Google Scholar]
- Postawa, T.; Gas, A. Fauna nietoperzy Wigierskiego Parku Narodowego (północno-wschodnia Polska). Stud. Chiropterologica 2003, 3–4, 31–42. [Google Scholar]
- Sachanowicz, K.; Marzec, M.; Ciechanowski, M.; Rachwald, A. Nietoperze Puszczy Rominckiej. Nietoperze 2001, 2, 109–115. [Google Scholar]
- Kowalski, M.; Ostrach-Kowalska, A.; Krasnodębski, I.; Sachanowicz, K.; Ignaczak, M.; Rusin, A. Nietoperze Parków Krajobrazowych: Górznieńsko-Lidzbarskiego i Welskiego. Nietoperze 2001, 2, 117–124. [Google Scholar]
- Ignaczak, M.; Radzicki, G.; Domański, J. Nietoperze Parku Krajobrazowego Międzyrzecza Warty i Widawki. Nietoperze 2001, 2, 125–134. [Google Scholar]
- Wikar, Z.; Ciechanowski, M. Ssaki rezerwatu przyrody „Studnica” i jego otoczenia. Chrońmy Przyr. Ojczystą 2019, 75, 374–387. [Google Scholar]
- Ciechanowski, M.; Przesmycka, A.; Sachanowicz, K. 2006. Nietoperze (Chiroptera) Wdzydzkiego Parku Krajobrazowego. Park. Nar. Rezerwaty Przyr. 2006, 25, 85–100. [Google Scholar]
- Apoznański, G.; Kokurewicz, T.; Błesznowska, J.; Kwasiborska, E.; Marszałek, T.; Górska, M. Use of Coniferous Plantations by Bats in Western Poland During Summer Months. Balt. For. 2020, 26, 1–8. [Google Scholar] [CrossRef]
- Ciechanowski, M. Ssaki (Mammalia). In Przyroda Rezerwatów Kurze Grzędy i Staniszewskie Błoto na Pojezierzu Kaszubskim; Herbich, J., Ciechanowski, M., Eds.; Fundacja Rozwoju Uniwersytetu Gdańskiego: Gdańsk, Poland, 2009; pp. 256–264. [Google Scholar]
- Jarzembowski, T.; Ostrach-Kowalska, A.; Rymarzak, G. Chiropterofauna Kaszubskiego Parku Krajobrazowego. Przegląd Przyr. 1997, 8, 123–127. [Google Scholar]
- Ciechanowski, M. Ssaki (Mammalia). In Przyroda Projektowanego Rezerwatu “Dolina Mirachowskiej Strugi” na Pojezierzu Kaszubskim; Ciechanowski, M., Fałtynowicz, W., Zieliński, S., Eds.; Acta Botanica Cassubica: Gdańsk, Poland, 2004; Volume 4, pp. 110–121. [Google Scholar]
- Wojtaszyn, G. Nietoperze Pojezierza Wałeckiego. Przegląd Przyr. 2002, 13, 199–211. [Google Scholar]
- Lesiński, G.; Gryz, J.; Rachwald, A.; Krauze-Gryz, D. Bat assemblages in fragmented forest complexes near Rogów (central Poland). For. Res. Pap. 2019, 79, 253–260. [Google Scholar] [CrossRef]
- Górecki, M.T. Nietoperze Chiroptera okolic Skierniewic. Przegląd Przyr. 1998, IX, 101–108. [Google Scholar]
- Lesiński, G.; Hejduk, J.; Gajęcka, K.; Górecki, M.; Janus, K.; Zieleniak, A. Nietoperze Bolimowskiego Parku Krajobrazowego i terenów otaczających. Park. Nar. Rezerwaty Przyr. 2018, 37, 65–80. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | Detectability Coefficient |
|---|---|
| Myotis myotis | 1.25 |
| Small Myotis 1 | 1.67 |
| Eptesicus serotinus | 0.63 |
| Pipistrellus spp. | 1.00 |
| Nyctalus noctula | 0.25 |
| Nyctalus leisleri | 0.31 |
| Plecotus auritus | 1.25 |
| Species | Mist Netting | D-1000X, MB, and EMT Detectors | Batcorder Detectors | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Raw | Adjusted | NL | Raw | Adjusted | NL | ||||||||
| n | % | NL | n | % | n | % | n | % | n | % | |||
| M. myotis | 1 | 0.2 | 1 | 4 | 0.06 | 5 | 0.09 | 3 | 3 | 0.02 | 4 | 0.03 | 2 |
| M. nattereri | 23 | 5.1 | 13 | 3 | 0.04 | 5 | 0.09 | 2 | 20 | 0.14 | 33 | 0.23 | 6 |
| M. daubentonii | 51 | 11.2 | 8 | - | - | - | - | - | - | - | - | - | - |
| Myotis sp. | - | - | - | 952 | 14.04 | 1590 | 27.46 | 9 | 869 | 5.98 | 1451 | 10.03 | 22 |
| E. serotinus | 5 | 1.1 | 4 | 156 | 2.30 | 98 | 1.70 | 10 | 81 | 0.56 | 51 | 0.35 | 12 |
| P. pipistrellus | 5 | 1.1 | 3 | 554 | 8.17 | 554 | 9.57 | 13 | 840 | 5.78 | 840 | 5.81 | 23 |
| P. pygmaeus | 273 | 60.0 | 14 | 2501 | 36.89 | 2501 | 43.19 | 16 | 7724 | 53.11 | 7724 | 53.40 | 28 |
| P. nathusii | 12 | 2.6 | 9 | 460 | 6.78 | 460 | 7.94 | 15 | 1075 | 7.39 | 1075 | 7.43 | 26 |
| Pipistrellus sp. | 2 | 0.4 | 1 | 38 | 0.56 | 38 | 0.66 | 6 | 3059 | 21.03 | 3059 | 21.15 | 30 |
| N. noctula | 59 | 13.0 | 5 | 1988 | 29.32 | 497 | 8.58 | 12 | 853 | 5.87 | 213 | 1.47 | 22 |
| N. leisleri | 6 | 1.3 | 1 | 120 | 1.77 | 37 | 0.64 | 8 | 12 | 0.08 | 4 | 0.03 | 5 |
| P. auritus | 18 | 4.0 | 4 | 4 | 0.06 | 5 | 0.09 | 3 | 7 | 0.05 | 9 | 0.06 | 5 |
| Total | 455 | 100.0 | 17 | 6780 | 100.00 | 5790 | 100.00 | 16 | 14,543 | 100.00 | 14,463 | 100.00 | 30 |
| NEV | - | - | - | 18 | - | - | - | - | 646 | - | - | - | - |
| Indet. | - | - | - | 30 | - | - | - | - | 1619 | - | - | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ciechanowski, M.; Wikar, Z.; Borzym, K.; Janikowska, E.; Brachman, J.; Jankowska-Jarek, M.; Bidziński, K. Exceptionally Uniform Bat Assemblages across Different Forest Habitats Are Dominated by Single Hyperabundant Generalist Species. Forests 2024, 15, 337. https://doi.org/10.3390/f15020337
Ciechanowski M, Wikar Z, Borzym K, Janikowska E, Brachman J, Jankowska-Jarek M, Bidziński K. Exceptionally Uniform Bat Assemblages across Different Forest Habitats Are Dominated by Single Hyperabundant Generalist Species. Forests. 2024; 15(2):337. https://doi.org/10.3390/f15020337
Chicago/Turabian StyleCiechanowski, Mateusz, Zuzanna Wikar, Katarzyna Borzym, Emilia Janikowska, Julia Brachman, Martyna Jankowska-Jarek, and Konrad Bidziński. 2024. "Exceptionally Uniform Bat Assemblages across Different Forest Habitats Are Dominated by Single Hyperabundant Generalist Species" Forests 15, no. 2: 337. https://doi.org/10.3390/f15020337