

Solar and Climatic Factors Affecting Tree-Ring Growth of Mountain Birch (Betula pubescens) beyond the Northern Timberline on Kola Peninsula, Northwestern Russia

Abstract
:1. Introduction
2. Materials and Methods
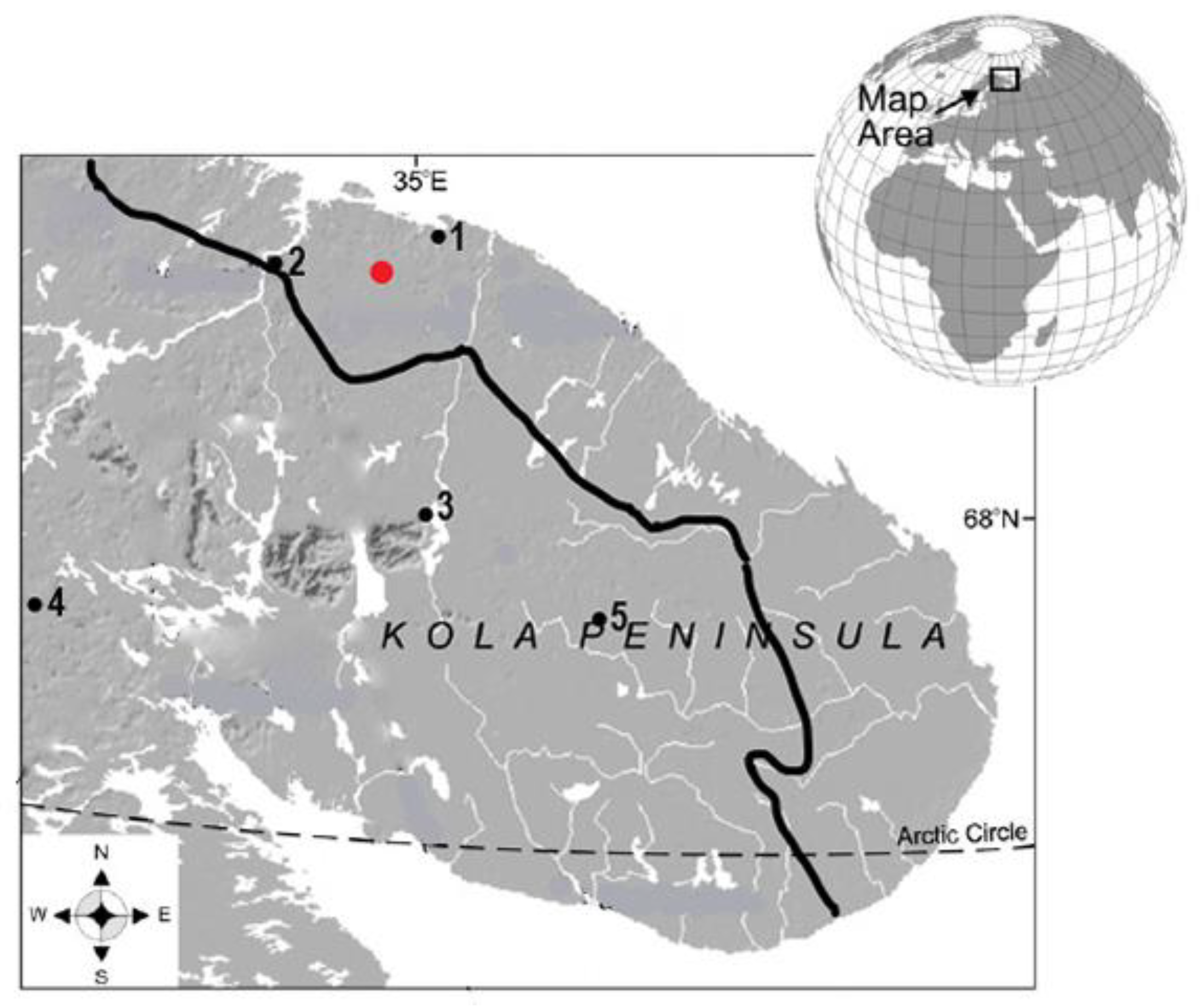
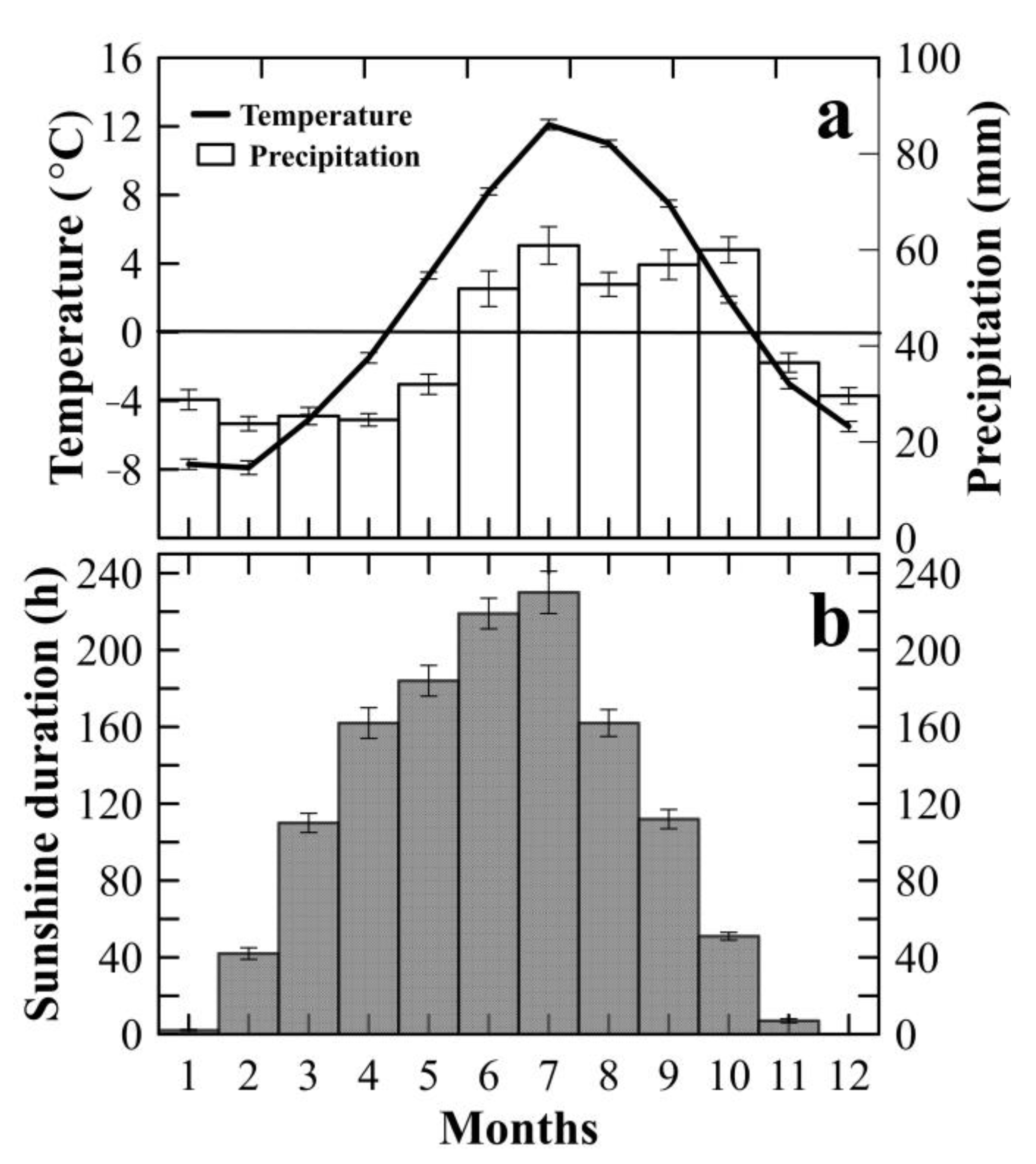
2.1. Study Site
2.2. Chronology Development
2.3. Data Sets
2.4. Statistical, Spectral and Wavelet Analysis
3. Results
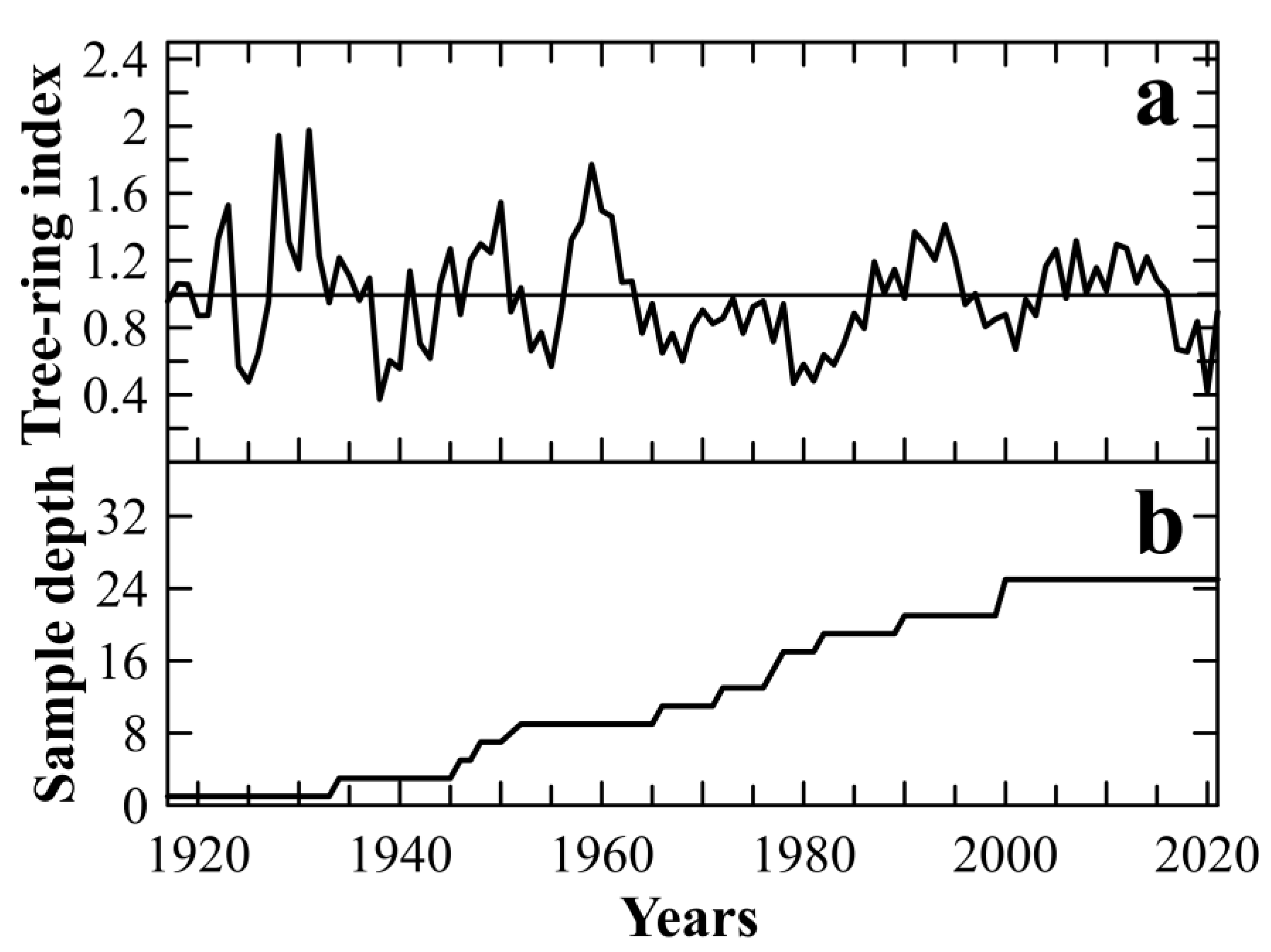
3.1. Chronology Statistics
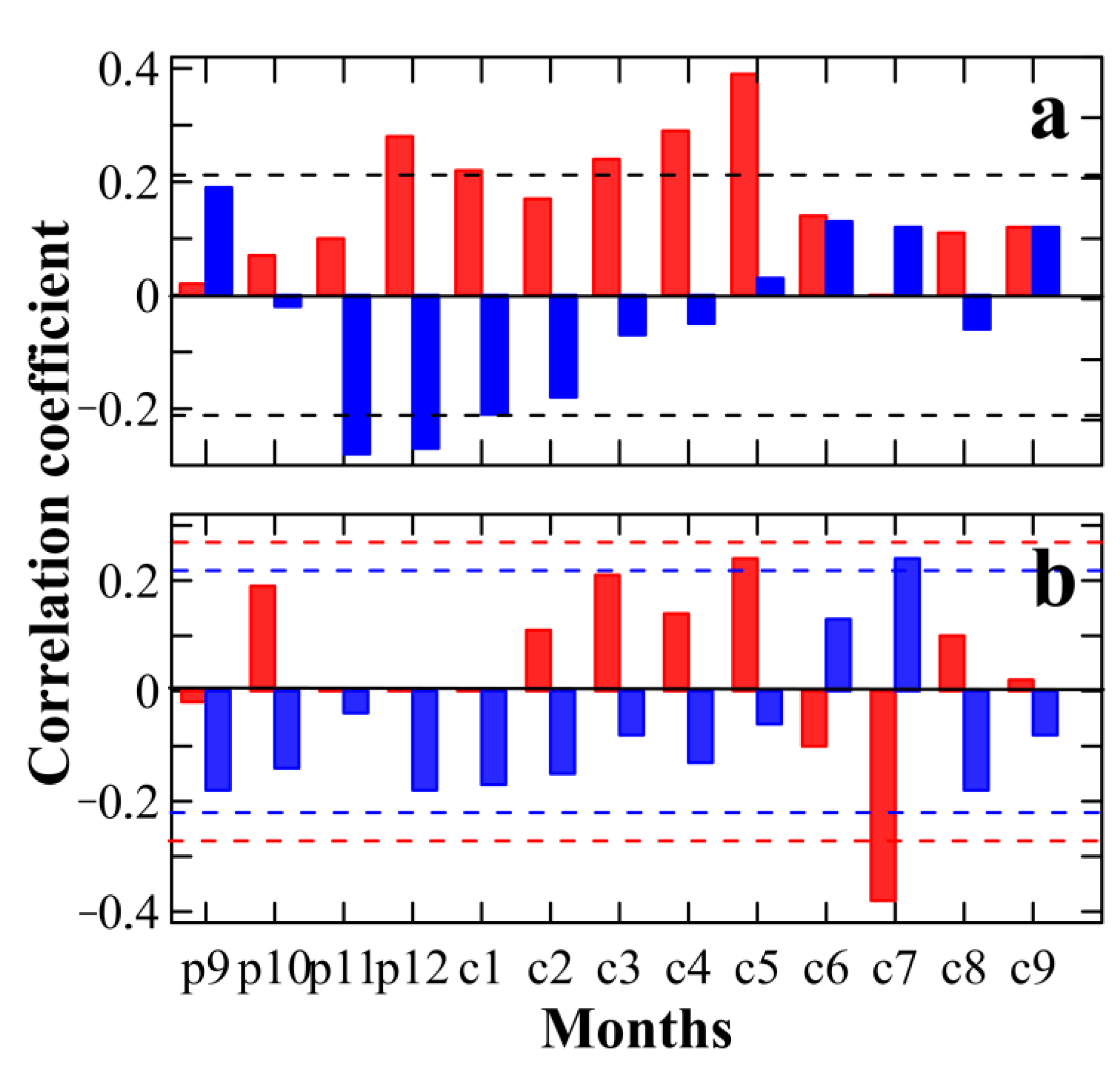
3.2. Tree Growth–Climate Relationship
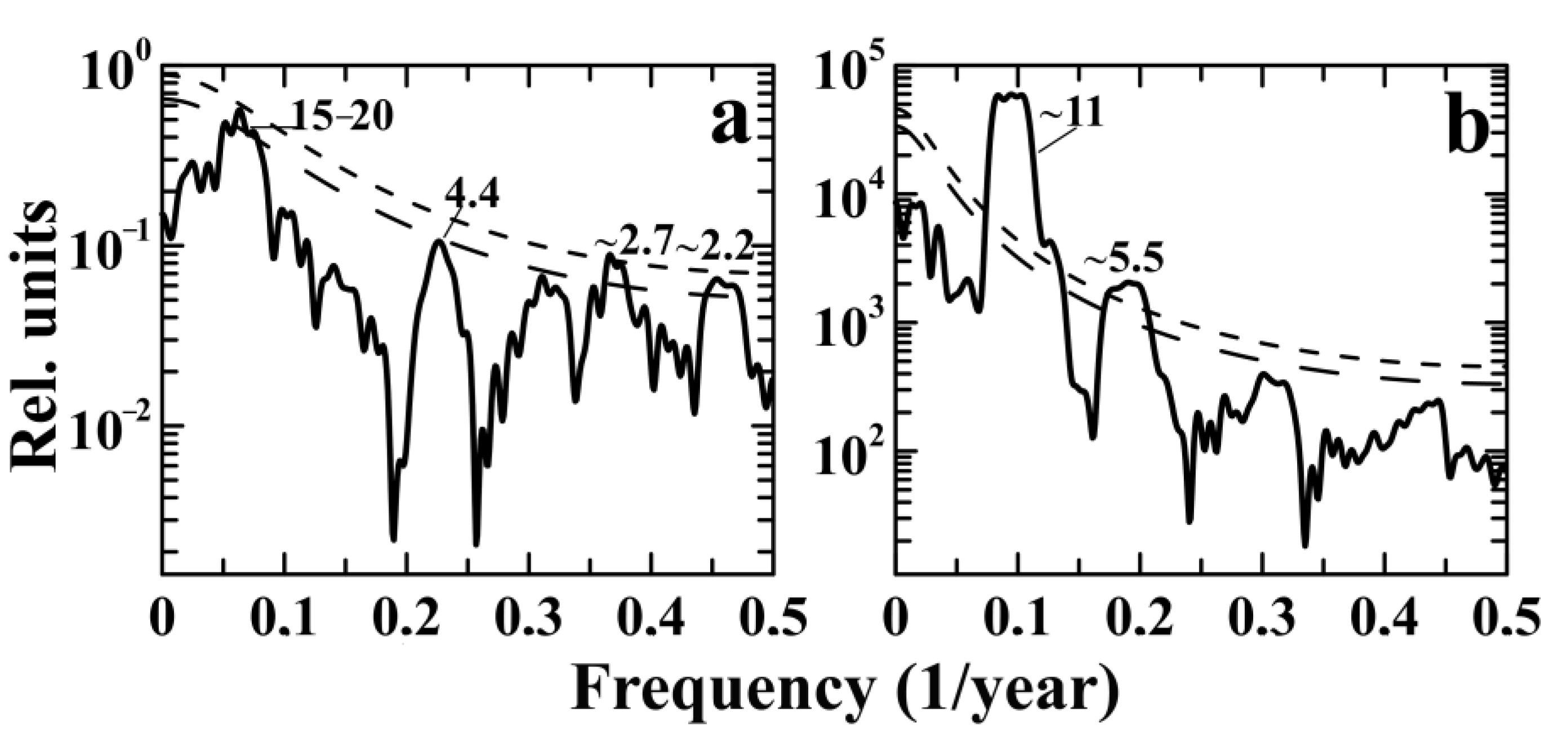
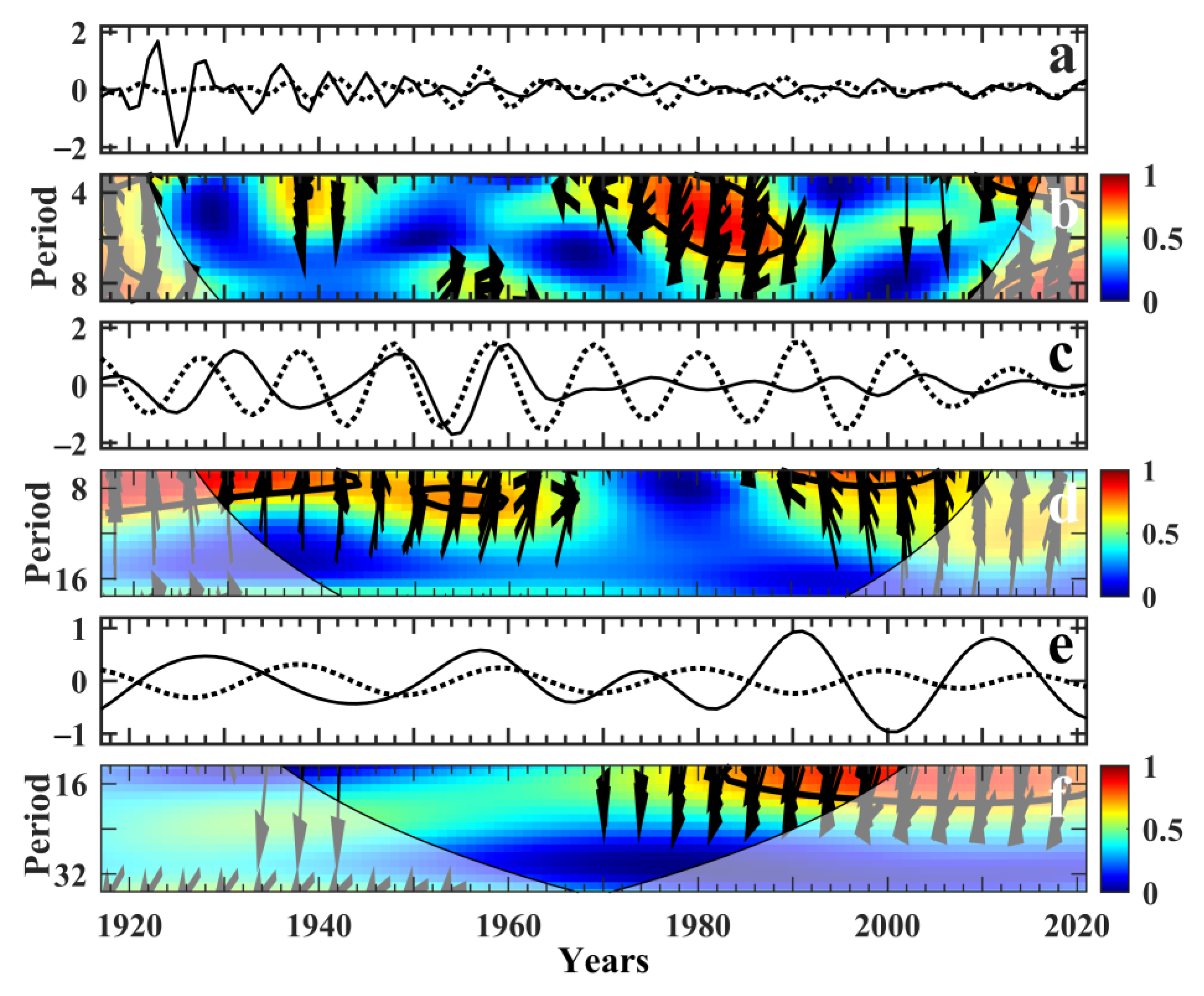
3.3. Spectral MTM and Wavelet Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fritts, H.C. Tree Rings and Climate; Academic Press: New York, NY, USA, 1976. [Google Scholar]
- Gervais, B.R.; MacDonald, G.M. A 403-year record of July temperatures and treelyne dynamics of Pinus sylvestris from the Kola Peninsula, Northwest Russia. Arct. Antarct. Alp. Res. 2000, 32, 295–302. [Google Scholar] [CrossRef]
- Miina, J. Dependence of tree-ring, earlywood and latewood indices of Scots pine and Norway spruce on climatic factors in eastern Finland. Ecol. Model. 2000, 132, 259–273. [Google Scholar] [CrossRef]
- Mäkinen, H.; Nöjd, P.; Mielikäinen, K. Climatic signal in annual growth variation of Norway spruce [Picea abies (L.) Karst.] along a transect from central Finland to the Arctic timberline. Can. J. For. Res. 2000, 30, 769–777. [Google Scholar] [CrossRef]
- Shumilov, O.I.; Kasatkina, E.A.; Lukina, N.V.; Kirtsideli, I.Y.; Kanatjev, A.G. Paleoclimatic potential of the northernmost juniper trees in Europe. Dendrochronologia 2007, 24, 123–130. [Google Scholar] [CrossRef]
- Helama, S.; Fauria, M.M.; Mielikäinen, K.; Timonen, M.; Eronen, M. Sub-Milankovich solar forcing of past climates: Mid and late Holocene perspectives. GSA Bull. 2010, 122, 11–12. [Google Scholar] [CrossRef]
- Kasatkina, E.A.; Shumilov, O.I.; Timonen, M. Solar activity imprints in tree-ring data from northwestern Russia. J. Atmo. Sol. Terr. Phys. 2019, 193, 105075. [Google Scholar] [CrossRef]
- Welling, A.; Rinne, P.; Viherä-Aarnio, A.; Kontunen-Soopela, S.; Heino, P.; Palva, E.T. Photoperiod and temperature differentially regulate the expression of two dehydrin genes during overwintering of birch (Betula pubescens Ehrh.). J. Experiment. Bot. 2004, 55, 507–516. [Google Scholar] [CrossRef]
- Takahashi, K.; Tokumitsu, Y.; Yasue, K. Climatic factors affecting the tree-ring width of Betula ermanii at the timberline on Mount Norikura, central Japan. Ecol. Res. 2005, 20, 445–451. [Google Scholar] [CrossRef]
- Velez-Ramirez, A.I.; van Ieperen, W.; Vreugdenhil, D.; Millenaar, F.F. Plants under continuous light. Trends Plant Sci. 2011, 16, 310–318. [Google Scholar] [CrossRef]
- Singh, R.K.; Svystun, T.; Al Dahmash, B.; Jönsson, A.M.; Bhalerao, R.P. Photoperiod- and temperature-mediated control of phenology in trees—A molecular perspective. New Phytol. 2017, 213, 511–524. [Google Scholar] [CrossRef]
- Brelsford, C.; Nybakken, L.; Kotilainen, T.K.; Robson, T.M. The influence of spectral composition on spring and autumn phenology in trees. Tree Physiol. 2019, 39, 925–950. [Google Scholar] [CrossRef] [PubMed]
- Kotilainen, T.; Aphalo, P.J.; Brelsford, C.C.; Böök, H.; Devraj, S.; Heikkilä, A.; Hernándes, R.; Kylling, A.; Lindfors, A.V.; Robson, T.M. Patterns in the spectral composition of sunlight and biologically meaningful spectral photon ratios as affected by atmospheric factors. Agric. For. Meteorol. 2020, 291, 108041. [Google Scholar] [CrossRef]
- Tenkanen, A.; Keinänen, M.; Oksanen, E.; Keski-Saari, S.; Kontunen-Sopela, S. Polar day syndrome: Differences in growth, photosynthetic traits and sink-size patterns between northern and southern Finnish silver birch (Betula pendula Roth) provenances in native and non-native photoperiods. Tree Physiol. 2023, 43, 16–30. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.-G.; Ma, Q.; Rossi, S.; Biondi, F.; Deslauriers, A.; Fonti, P.; Liang, E.; Mäkinen, H.; Oberhuber, W.; Rathgeber, C.B.K.; et al. Photoperiod and temperature as dominant environmental drivers triggering secondary growth resumption in Northen Hemisphere conifers. Proc. Natl. Acad. Sci. USA 2020, 117, 20645–20652. [Google Scholar] [CrossRef] [PubMed]
- Mølmann, J.A.; Junttila, O.; Johnsen, Ø.; Olsen, J.E. Effects of red, far-red and blue light in maintaining growth in latitudinal populations of Norway spruce (Picea abies). Plant Cell Environ. 2006, 29, 166–172. [Google Scholar] [CrossRef] [PubMed]
- Kremenetski, K.V.; MacDonald, G.M.; Gervais, B.R.; Borisova, O.K.; Snyder, J.A. Holocene vegetation history and climate change on the Kola Peninsula, Russia: A case study from a small tundra lake. Quatern. Int. 2004, 122, 57–68. [Google Scholar] [CrossRef]
- Shutova, E.; Weilgolaski, F.E.; Karlsen, S.R.; Makarova, O.; Berlina, N.; Filimonova, T.; Haraldsson, E.; Aspholm, P.E.; Flø, L.; Høgda, K.A. Growing seasons of Nordic mountain birch in northernmost Europe as indicated by long-term field studies and analyses of satellite images. Int. J. Biometeorol. 2006, 51, 155–166. [Google Scholar] [CrossRef]
- Aune, S.; Hofgaard, A.; Söderström, L. Contrasting climate- and land-use-driven tree encroachment patterns of subarctic tundra in northern Norway and the Kola Peninsula. Can. J. For. Res. 2011, 41, 437–449. [Google Scholar] [CrossRef]
- Young, A.B.; Cairns, D.M.; Lafon, C.W.; Moen, J.; Martin, L.E. Dendroclimatic relationships and possible implications for mountain birch and Scots pine at treeline in northern Sweden through the 21th century. Can. J. For. Res. 2011, 41, 450–459. [Google Scholar] [CrossRef]
- Krikunova, A.I.; Kostromina, N.A.; Savelieva, L.A.; Tolstobrov, D.S.; Petrov, A.Y.; Long, T.; Kobe, F.; Leipe, C.; Tarasov, P.E. Late- and postglacial vegetation and climate history of the central Kola Peninsula derived from a radiocarbon-dated pollen record of Lake Kamenistoe. Palaeogeogr. Palaeoclim. Palaeoecol. 2022, 603, 111191. [Google Scholar] [CrossRef]
- Van Bogaert, R. Recent Treeline Dynamics in Sub-Arctic Sweden: A Multi-Disciplinary Landscape Assessment. Ph.D. Thesis, Department of Geography, Ghent University, Ghent, Sweden, 2010. [Google Scholar]
- Yu, D.P.; Gu, H.Y.; Wang, Q.L. Relationships of climate change and tree ring of Betula ermanii tree line forest in Changbai Mountain. J. For. Res. 2005, 16, 187–192. [Google Scholar] [CrossRef]
- Bhattacharyya, A.; Shah, S.K.; Chaudhary, V. Would tree ring data of Betula utilis be potential for the analysis of Himalayan glacial fluctuations? Curr. Sci. 2006, 91, 754–761. Available online: http://www.jstor.org/stable/24093904 (accessed on 5 April 2023).
- Dawadi, B.; Liang, E.; Tian, L.; Devkota, L.P.; Yao, T. Pre-monsoon precipitation signal in tree rings of timberline Betula utilis in the central Himalayas. Quatern. Int. 2012, 283, 72–77. [Google Scholar] [CrossRef]
- Liang, E.; Dawadi, B.; Pederson, N.; Eckstein, D. Is the growth of birch at the upper timberline in the Himalayas limited by moisture or by temperature? Ecology 2014, 95, 2453–2465. [Google Scholar] [CrossRef]
- Gaire, N.P.; Koirala, M.; Bhuju, D.R.; Carrer, M. Site- and species-specific treeline responses to climatic variability in eastern Nepal Himalaya. Dendrochronologia 2017, 41, 44–56. [Google Scholar] [CrossRef]
- Eckstein, D.; Hoogesteger, J.; Holmes, R.L. Insect-related differences in growth of birch and pine at northern tree-line in Swedish Lapland. Ecography 1991, 14, 18–23. [Google Scholar] [CrossRef]
- Levanič, T.; Eggertsson, O. Climatic effects on birch (Betula pubescens Ehrh.) growth in Fnjoskadalur valley, northern Iceland. Dendrochronologia 2008, 25, 135–143. [Google Scholar] [CrossRef]
- Harr, L.; Esper, J.; Kirchhefer, J.A.; Zhou, W.; Hartl, C. Growth response of Betula pubescens Ehrh. to varying disturbance factors in northern Norway. Trees 2021, 35, 421–431. [Google Scholar] [CrossRef]
- Stridbeck, P.; Björklund, J.; Fuentes, M.; Gunnarson, B.E.; Jönsson, A.M.; Rayner, D.; Rocha, E.; Zhang, P.; Seftigen, K. Partly decoupled tree-ring width and leaf phenology response to 20th century temperature change in Sweden. Dendrochronologia 2022, 75, 125993. [Google Scholar] [CrossRef]
- Cahoon, S.M.P.; Sullivan, P.F.; Brownlee, A.H.; Pattison, R.R.; Andersen, H.-E.; Legner, K.; Hollingsworth, T.N. Contrasting drivers and trends of coniferous and deciduous tree growth in interior Alaska. Ecology 2018, 99, 1284–1295. [Google Scholar] [CrossRef]
- Bumann, E.; Awada, T.; Wardlow, B.; Hayes, M.; Okalebo, J.; Helzer, C.; Mazis, A.; Hiller, J.; Cherubini, P. Assessing responses of Betula papyrifera to climate variability in a remnant population along the Niobrara River Valley in Nebraska, U.S.A., through dendroecological and remote-sensing techniques. Can. J. For. Res. 2019, 49, 423–433. [Google Scholar] [CrossRef]
- Wang, X.; Zhao, X.; Gao, L. Climatic response of Betula ermanii along an altitudinal gradient in the northern slope of Changbai Mountain, China. Dendrobiology 2013, 70, 99–107. [Google Scholar] [CrossRef]
- Fonti, M.V.; Tychkov, I.I.; Shishov, V.V.; Shashkin, A.V.; Prokushkin, A.S. Plant-soil-climate interaction in observed and simulated tree-radial growth dynamics of downy birch in permafrost. Front. Plant Sci. 2022, 13, 780153. [Google Scholar] [CrossRef] [PubMed]
- Solomina, O.N.; Muravyev, Y.D.; Braeuning, A.; Kravchenko, G.N. Two new ring width chronologies of larch and birch from the Kamchatka peninsula (Russia) and their relationship to climate and volcanic activities. In Cryospheric Studies in Kamchatka II; Naruse, R., Ed.; Institute of Low Temperature Science, Hokkaido University: Sapporo, Japan, 1999; pp. 111–124. [Google Scholar]
- Cedro, A. A birch tree as a witness in a murder and cannibalism case. Forests 2023, 14, 1132. [Google Scholar] [CrossRef]
- Hartl, C.; St George, S.; Konter, O.; Harr, L.; Scholz, D.; Kirchhefer, A.; Esper, J. Warfare dendrochronology: Trees witness the deployment of the German battleship Tirpitz in Norway. Anthropocene 2019, 27, 100212. [Google Scholar] [CrossRef]
- Marshall, G.J.; Vignols, R.M.; Rees, W.G. Climate change in the Kola Peninsula, Arctic Russia, during the last 50 years from meteorological observations. J. Clim. 2016, 29, 6823–6840. [Google Scholar] [CrossRef]
- Pereverzev, V.N. Zonal features of humus formation in Al-Fe-humus podzols of the Kola Peninsula. Euras. Soil Sci. 2011, 44, 1178–1183. [Google Scholar] [CrossRef]
- Blinova, I.; Chmielewski, F.-M. Climatic warming above the Arctic Circle: Are there trends in timing and length of the thermal growing season in Murmansk Region (Russia) between 1951 and 2012? Int. J. Biometeorol. 2015, 59, 693–705. [Google Scholar] [CrossRef]
- Saltan, N.V.; Sviatkovskaya, E.A. Ecophysiological features of Larix sibirica in urbanecosystems of the Kola north in the railway influence zone. Czech Polar Rep. 2021, 11, 305–307. [Google Scholar] [CrossRef]
- Gervais, B.R.; MacDonald, G.M.; Snyder, J.A.; Kremenetski, C.V. Pinus sylvestris treeline development and movement on the Kola Peninsula of Russia: Pollen and stomate evidence. J. Ecol. 2002, 90, 627–638. [Google Scholar] [CrossRef]
- Kanatjev, A.G.; Shumilov, O.I.; Kasatkina, E.A. Software for dendrochronological measurements. Instrum. Exp. Tech. 2014, 57, 214–217. [Google Scholar] [CrossRef]
- Holmes, R.L. Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bull. 1983, 43, 69–75. Available online: http://hdl.handle.net/10150/261223 (accessed on 10 May 2023).
- Cook, E.R.; Kairiukstis, L.A. Method of Dendrochronology; Kluwer Academic Publishing: Dordrecht, Germany, 1997. [Google Scholar]
- All-Russia Research Institute of Hydrometeorological Information—World Data Center (RIHMI–WDC), Roshydromet. Available online: http://meteo.ru (accessed on 6 April 2023).
- Lean, J.R.; Beer, J.; Bradley, R. Reconstruction of solar irradiance since 1610, Implications for climate change. Geophys. Res. Lett. 1995, 22, 3195–3198. [Google Scholar] [CrossRef]
- WDC-SILSO, Royal Observatory of Belgium, Brussels. Available online: http://www.sidc.be/SILSO (accessed on 11 June 2023).
- Thomson, D.J. Spectrum estimation and harmonic analysis. Proc. IEEE 1982, 70, 1055–1067. [Google Scholar] [CrossRef]
- Ghil, M.; Allen, M.R.; Dettinger, M.D.; Ide, K.; Kondrashov, D.; Mann, M.E.; Robertson, A.W.; Saunders, A.; Tian, Y.; Varadi, F.; et al. Advanced spectral methods for climate time series. Rev. Geophys. 2002, 40, 1003. [Google Scholar] [CrossRef]
- Torrence, C.; Compo, G.P. A practical guide to wavelet analysis. Bull. Am. Meteor. Soc. 1998, 79, 61–78. [Google Scholar] [CrossRef]
- Grinsted, A.; Moore, J.C.; Jevreeva, S. Application of the cross wavelet transform and wavelet coherence to geophysical time series. Nonlin. Process. Geophys. 2004, 11, 561–566. [Google Scholar] [CrossRef]
- Farge, M. Wavelet transforms and their applications to turbulence. Annu. Rev. Fluid Mech. 1992, 24, 395–457. [Google Scholar] [CrossRef]
- Wigley, T.M.L.; Briffa, K.R.; Jones, P.D. On the average value of correlated time series, with applications in dendrochronology and hydrometeorology. J. Appl. Meteorol. Clim. 1984, 23, 201–213. [Google Scholar] [CrossRef]
- Pudas, E.; Leppälä, M.; Tolvanen, A.; Poikolainen, J.; Venäläinen, A.; Kubin, E. Trends in phenology of Betula pubescens across the boreal zone in Finland. Int. J. Biometeorol. 2008, 52, 251–259. [Google Scholar] [CrossRef]
- Marion, L.; Gričar, J.; Oven, P. Wood formation in urban Norway maple trees studied by the micro-coring method. Dendrochronologia 2007, 25, 97–102. [Google Scholar] [CrossRef]
- Ohse, B.; Jansen, F.; Wilmking, M. Do limiting factors at Alaskan treelines shift with climatic regimes? Environ. Res. Lett. 2012, 7, 015505. [Google Scholar] [CrossRef]
- Bandekar, G.; Odland, A. Ecological characterization of northernmost birch forests and treeline ecotones in Norway. Phytocoenologia 2017, 47, 111–124. [Google Scholar] [CrossRef]
- Moiseev, P.A.; Galimova, A.A.; Bubnov, M.O.; Devi, N.M.; Fomin, V.V. Tree stands and their productivity dynamics at the upper growing limit in Khibiny on the background of modern climate changes. Russ. J. Ecol. 2019, 50, 431–444. [Google Scholar] [CrossRef]
- Paulsen, J.; Korner, C. A climate-based model to predict potential treeline position around the globe. Appl. Bot. 2014, 124, 1–12. [Google Scholar] [CrossRef]
- Welling, A.; Palva, E.T. Molecular control of cold acclimation in trees. Physiol. Plantarum 2006, 127, 167–181. [Google Scholar] [CrossRef]
- Zanuzdaeva, N.V.; Karimova, M.E. Impact of climate change on phenological parameters (Lapland State Reserve, Murmansk region). Trans. Kola Sci. Centre 2021, 12, 169–174. [Google Scholar] [CrossRef]
- Vaganov, E.A.; Hughes, M.K.; Kirdyanov, A.V.; Schweingruber, F.H.; Silkin, P.P. Influence of snowfall and melt timing on tree growth in subarctic Eurasia. Nature 1999, 400, 149–151. [Google Scholar] [CrossRef]
- Linkosalo, T.; Lechowicz, M.J. Twilight far-red treatment advances leaf bud burst of silver birch (Betula pendula). Tree Physiol. 2006, 26, 1249–1256. [Google Scholar] [CrossRef]
- Kasatkina, E.A.; Shumilov, O.I.; Denisov, D.B.; Makarov, D.V. Recent shift in diatom record from Lake Rabbvatnet: Response to global warming or solar variability? Acta Bot. Brasilica 2023, 37, e20220269. [Google Scholar] [CrossRef]
- Cook, E.R.; Meko, D.M.; Stockton, C.W. A new assessment of possible solar and lunar forcing of the bidecadal drought rhythm in the Western United States. J. Clim. 1997, 10, 1343–1356. [Google Scholar] [CrossRef]
- Raspopov, O.M.; Shumilov, O.I.; Kasatkina, E.A.; Turunen, E.; Lindholm, M. 35-year climatic Bruckner cycle—Solar control of climate variability? In The Solar Cycle and Terrestrial Climate, Solar and Space Weather; Wilson, A., Ed.; ESA Publications Division: Noordwijk, The Netherlands, 2000; pp. 517–520. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Value |
|---|---|
| Chronology time span (year) | AD 1917–2021 (105) |
| Number of cores (trees) | 25 (16) |
| Standard deviation (SD) | 0.54 |
| Mean sensitivity (MS) | 0.38 |
| First-order autocorrelation | 0.50 |
| Mean inter-series correlation (RBAR) | 0.37 |
| Express population signal (EPS) | 0.9 |
| Month | MUR (70) | TER (50) | KRS (200) | LVZ (95) | KVD (250) |
|---|---|---|---|---|---|
| January | 0.18 | 0.22 | 0.14 | 0.14 | 0.12 |
| February | 0.19 | 0.17 | 0.17 | 0.11 | 0.17 |
| March | 0.27 * | 0.24 | 0.22 | 0.23 | 0.27 * |
| April | 0.3 * | 0.29 * | 0.19 | 0.25 | 0.22 |
| May | 0.34 ** | 0.39 ** | 0.30 * | 0.36 ** | 0.25 * |
| June | 0.08 | 0.14 | 0.11 | 0.08 | 0.05 |
| July | 0 | 0 | 0 | −0.12 | 0 |
| August | 0.25 | 0.11 | 0.15 | 0.18 | 0.19 |
| September | 0.09 | 0.12 | 0.07 | 0.11 | 0.14 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shumilov, O.I.; Kasatkina, E.A.; Potorochin, E.O. Solar and Climatic Factors Affecting Tree-Ring Growth of Mountain Birch (Betula pubescens) beyond the Northern Timberline on Kola Peninsula, Northwestern Russia. Forests 2024, 15, 37. https://doi.org/10.3390/f15010037
Shumilov OI, Kasatkina EA, Potorochin EO. Solar and Climatic Factors Affecting Tree-Ring Growth of Mountain Birch (Betula pubescens) beyond the Northern Timberline on Kola Peninsula, Northwestern Russia. Forests. 2024; 15(1):37. https://doi.org/10.3390/f15010037
Chicago/Turabian StyleShumilov, Oleg I., Elena A. Kasatkina, and Evgeniy O. Potorochin. 2024. "Solar and Climatic Factors Affecting Tree-Ring Growth of Mountain Birch (Betula pubescens) beyond the Northern Timberline on Kola Peninsula, Northwestern Russia" Forests 15, no. 1: 37. https://doi.org/10.3390/f15010037