
Barcoding Hymenoptera: 11 Malaise Traps in Three Thai Forests: The First 68 Trap Weeks and 15,338 Parasitoid Wasp Sequences

Abstract
:1. Introduction
2. Materials and Methods
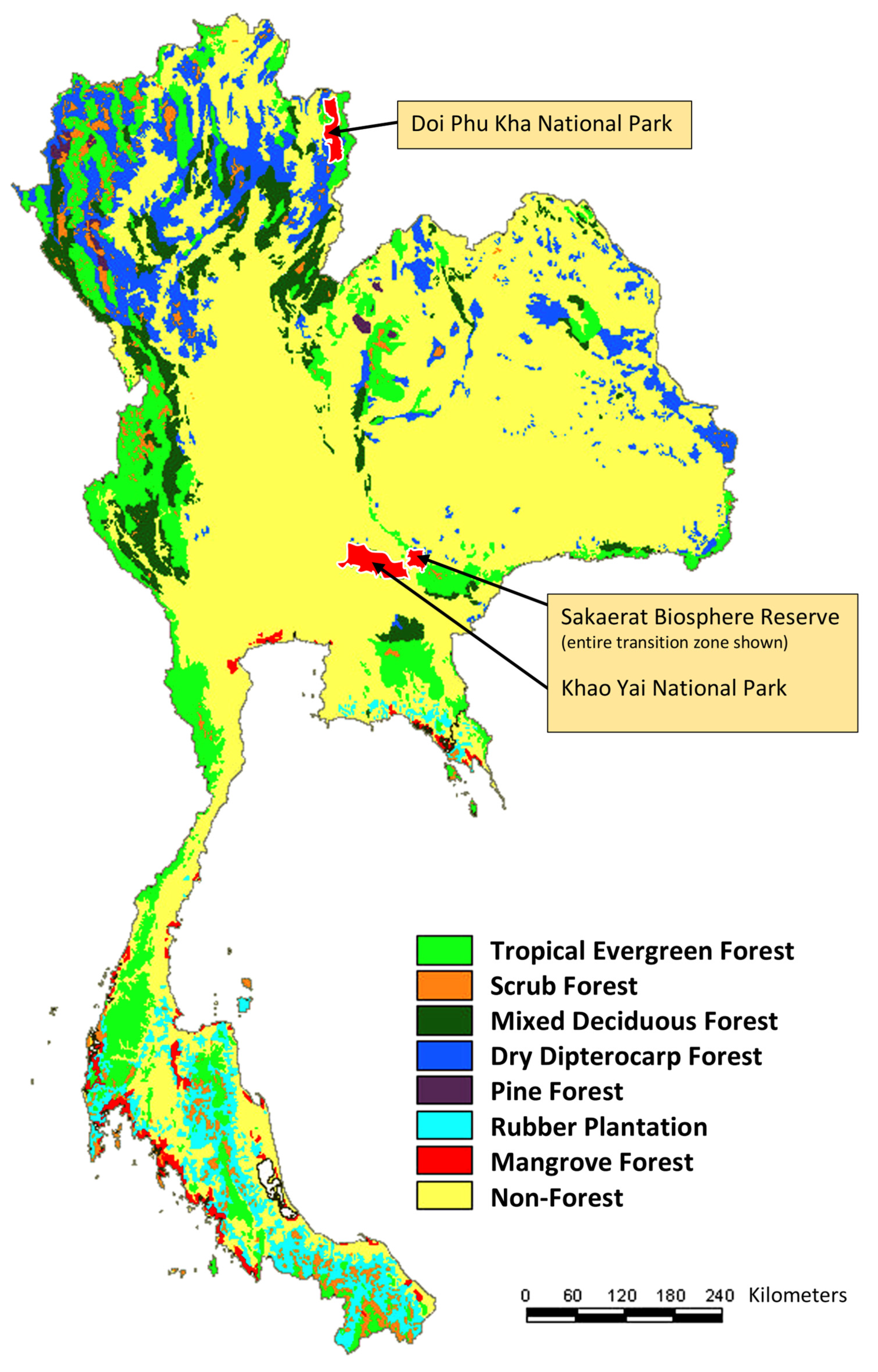
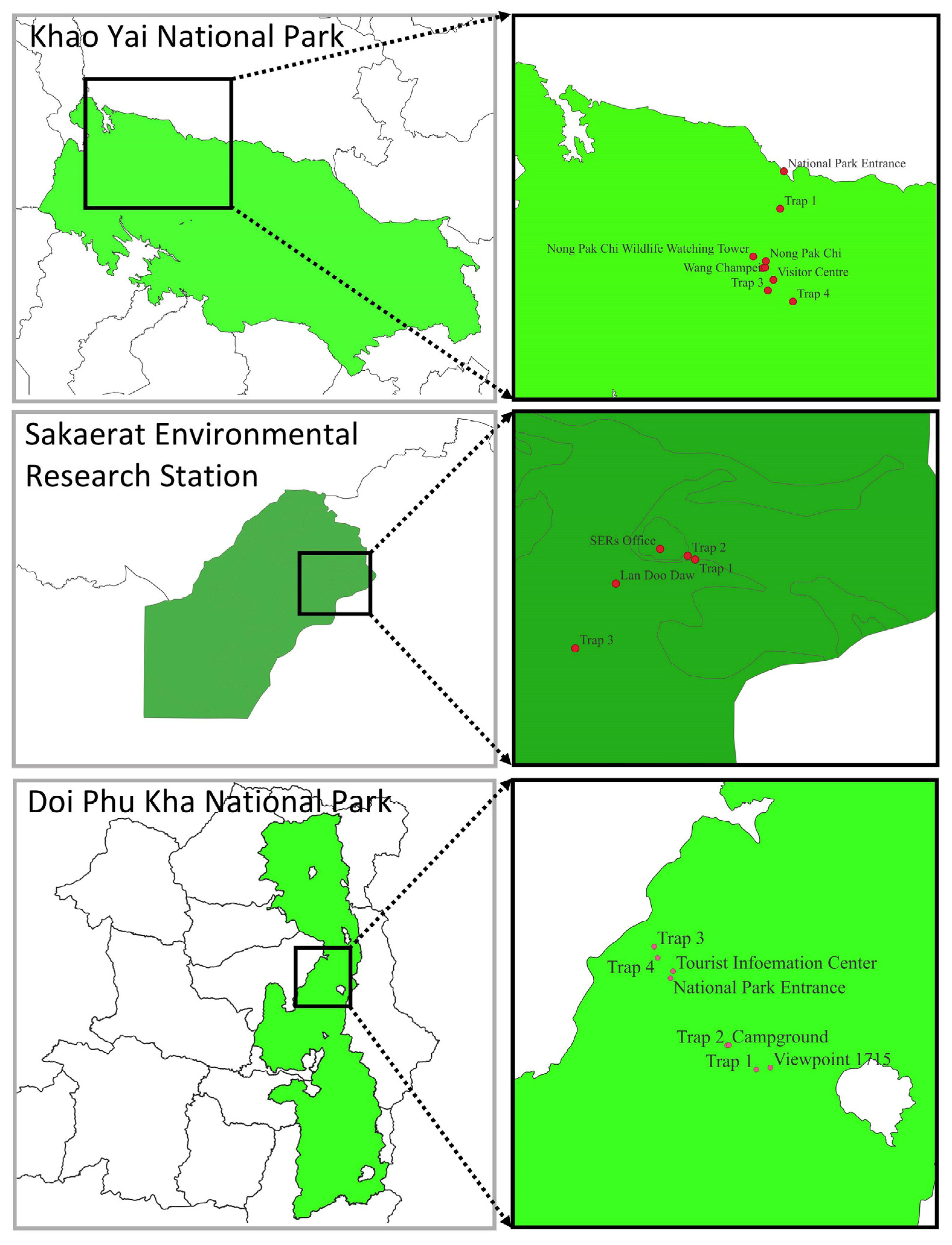
2.1. Field Sampling
2.2. Specimen Photography and Barcoding
2.3. Estimating Unknown Diversity
3. Results
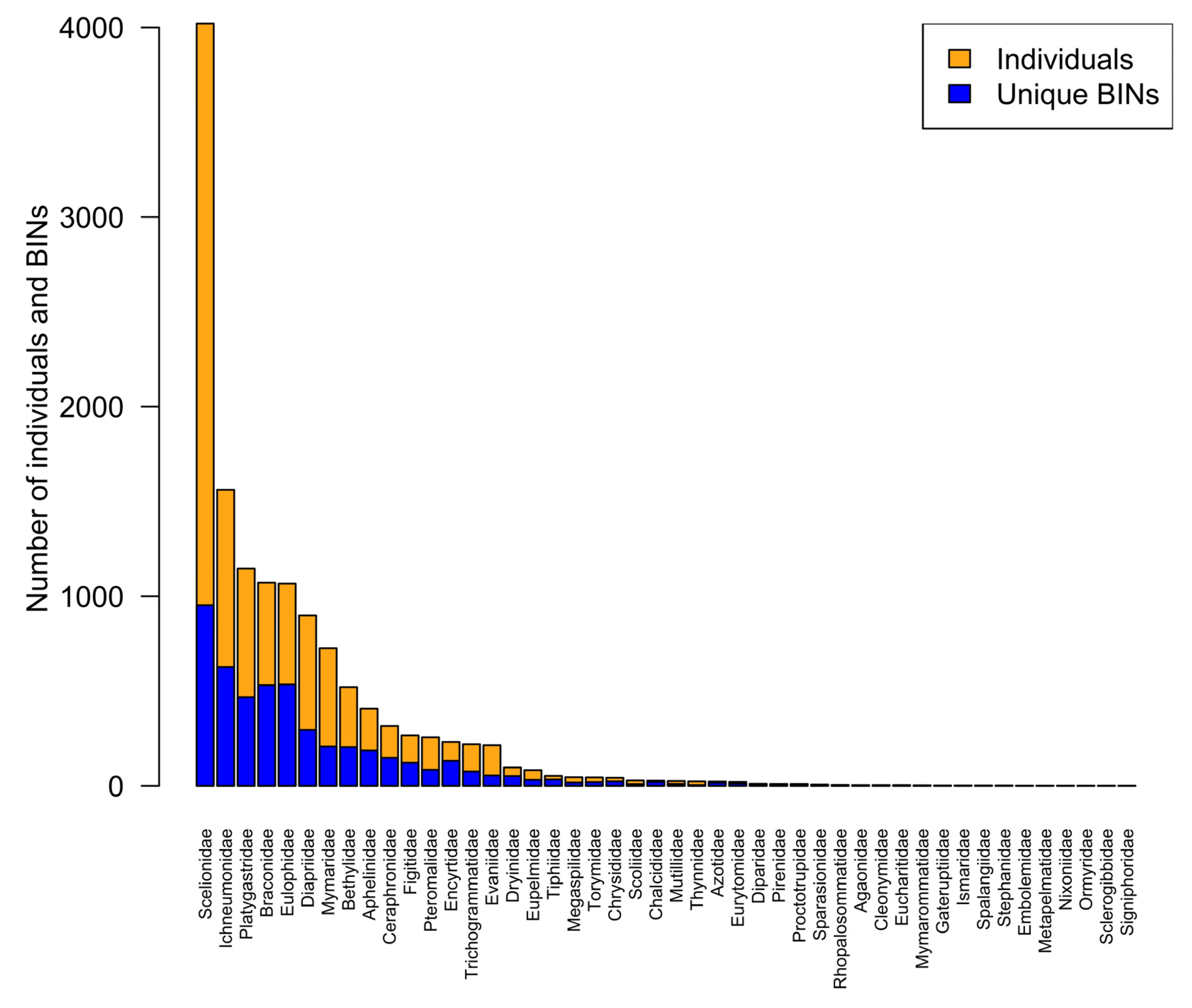
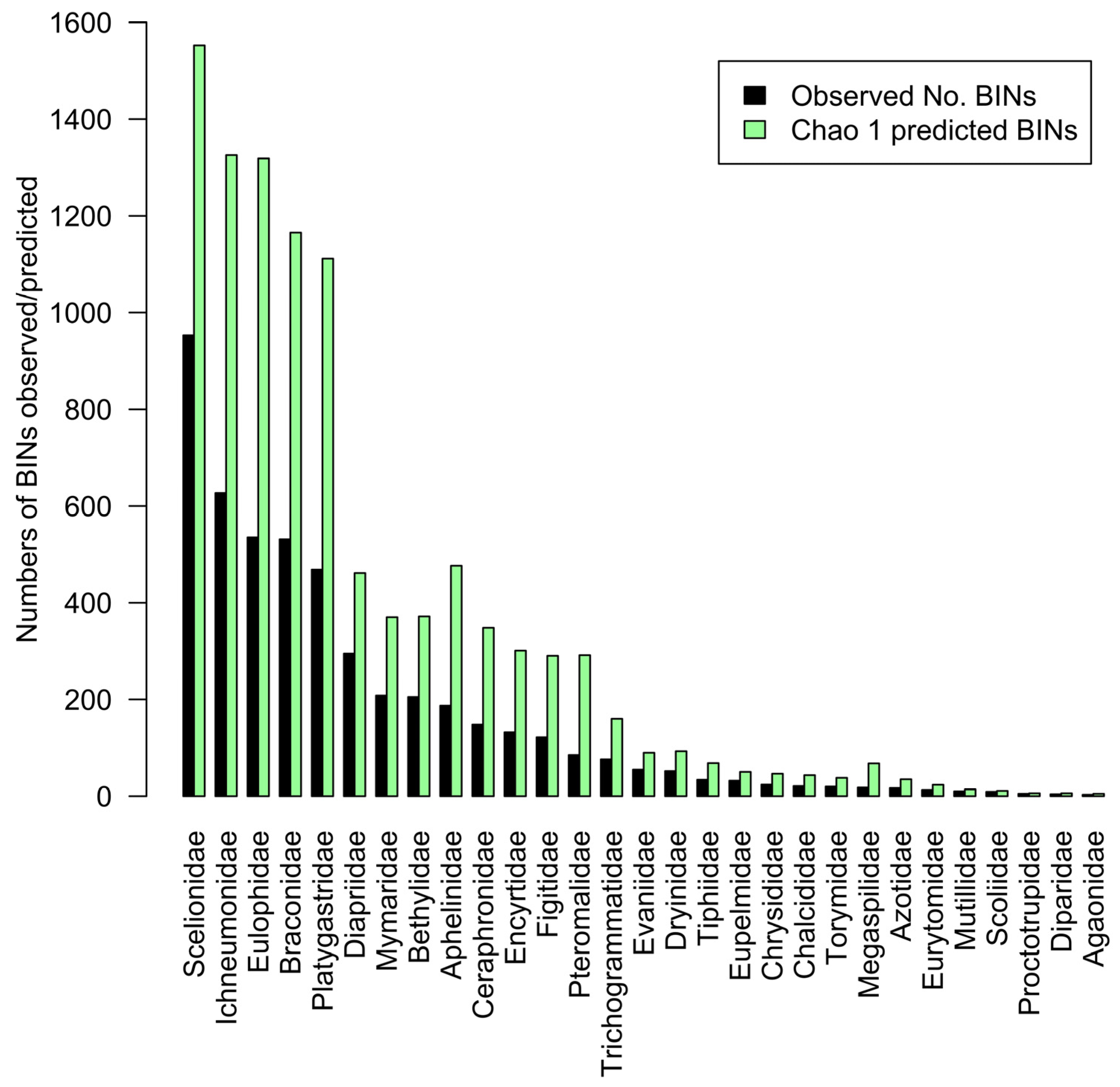
3.1. Total Parasitoid Hymenoptera BIN Representation by Family
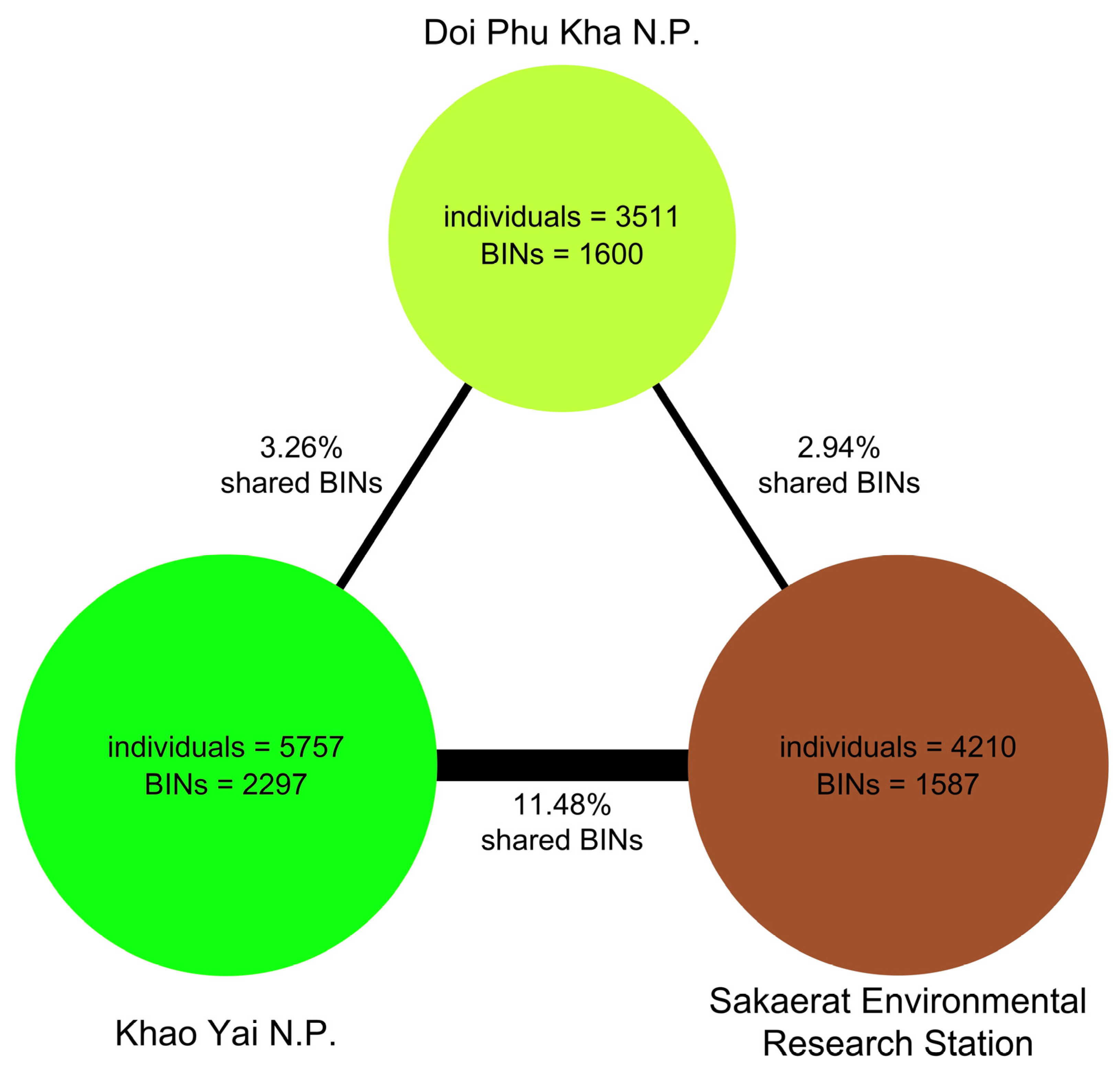
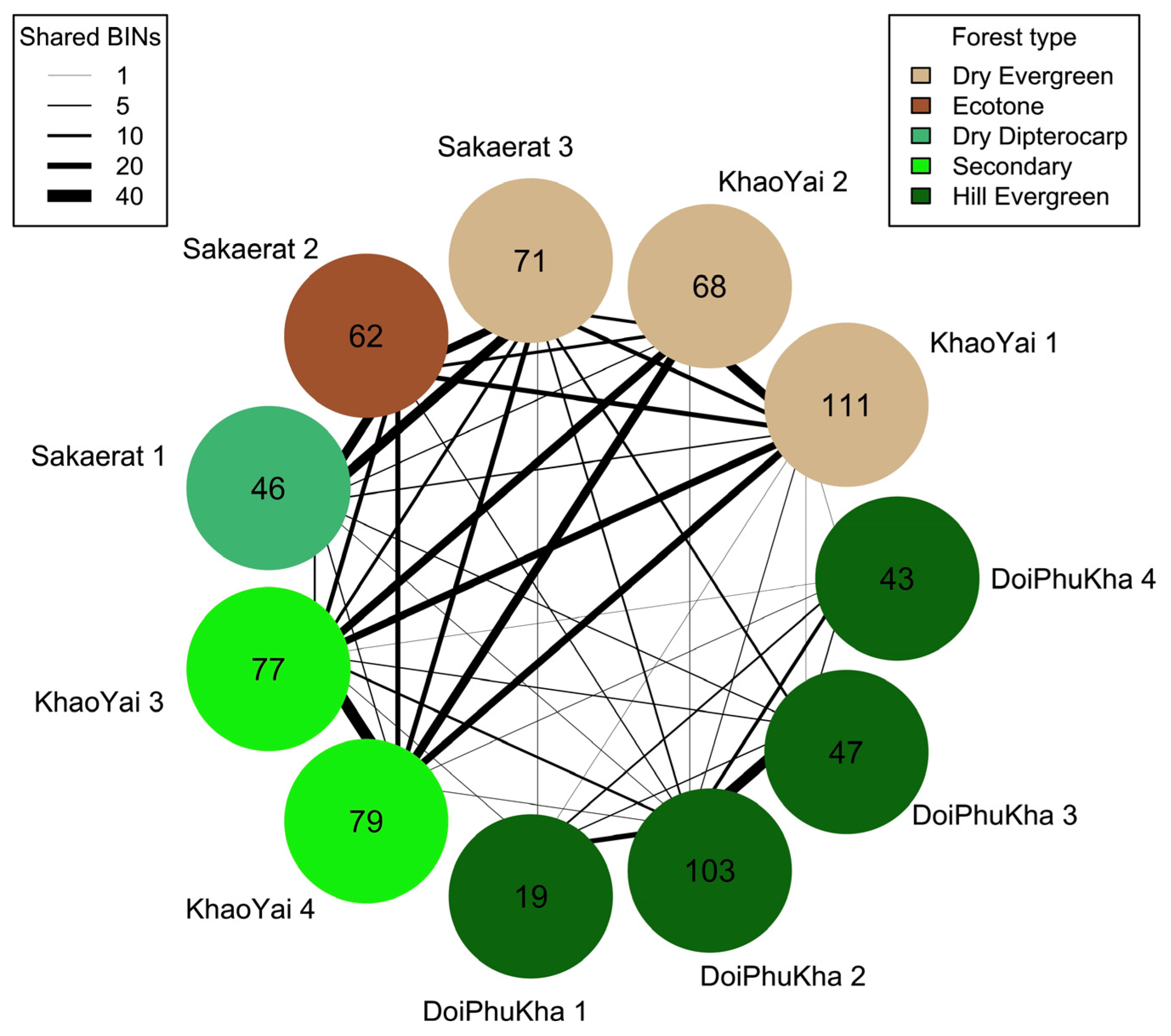
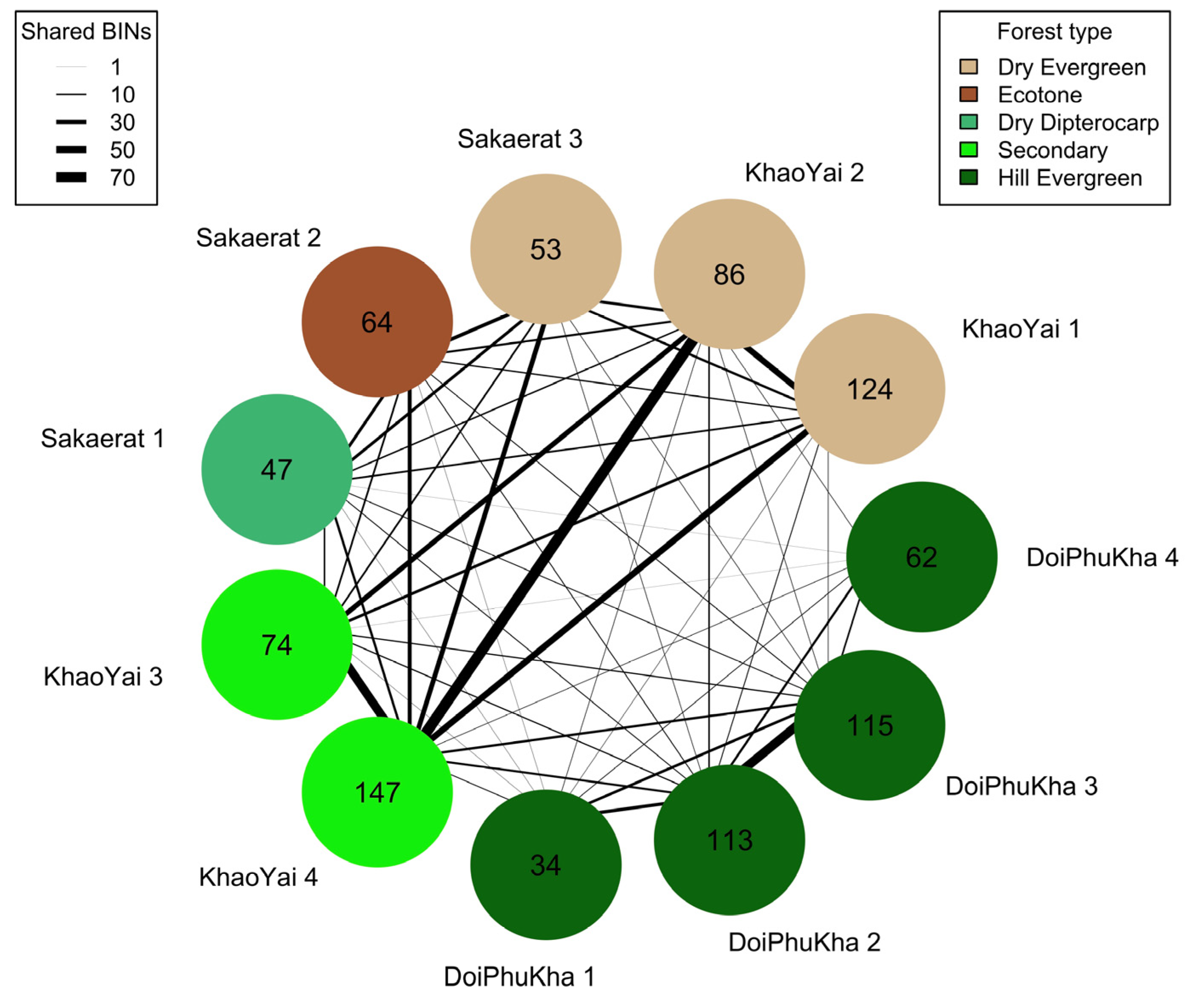
BIN Overlap between Sites and Traps
3.2. Family Representation
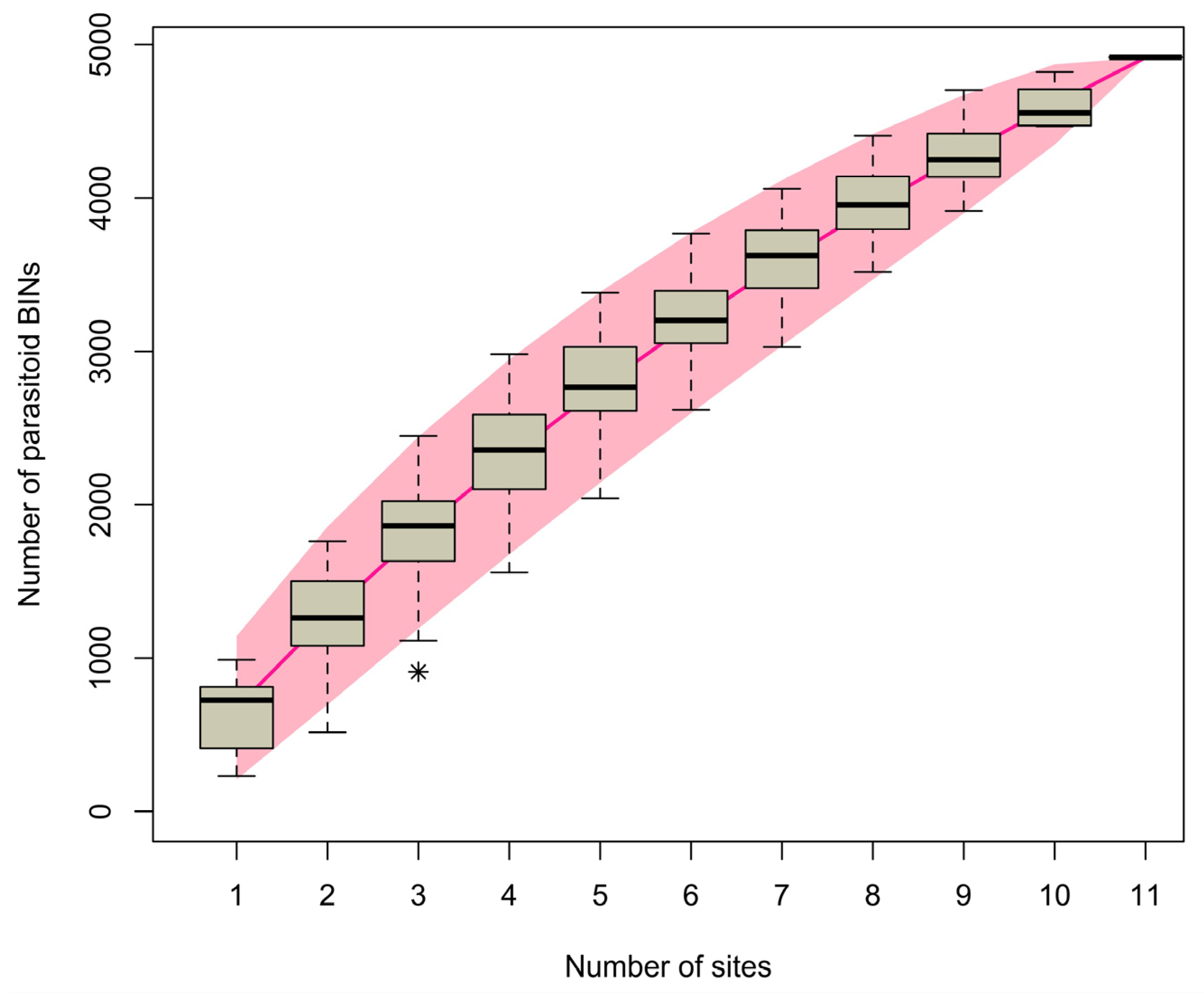
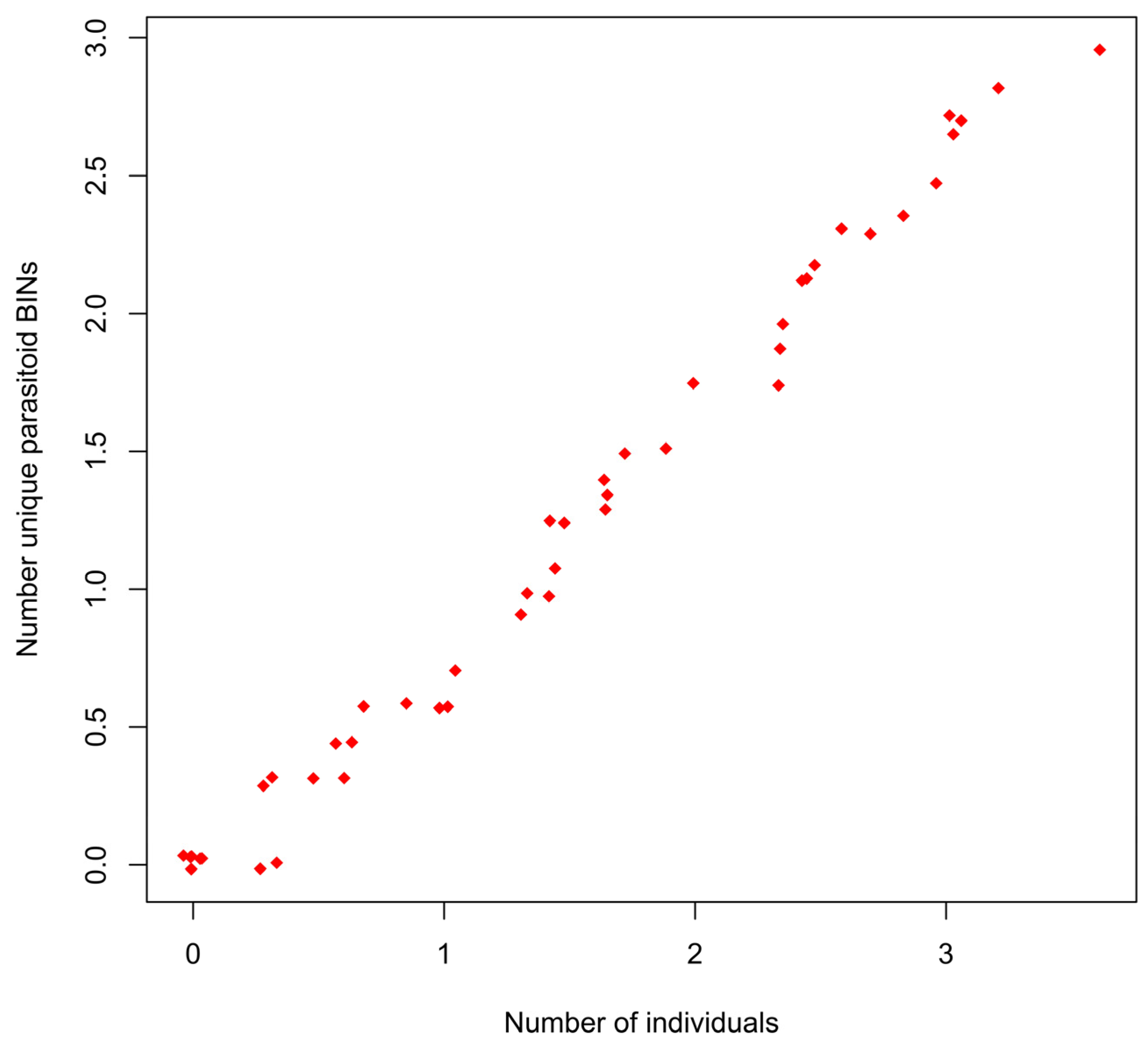
3.3. BIN Accumulation
3.4. BIN Extrapolation



3.5. Genus and Species Identification
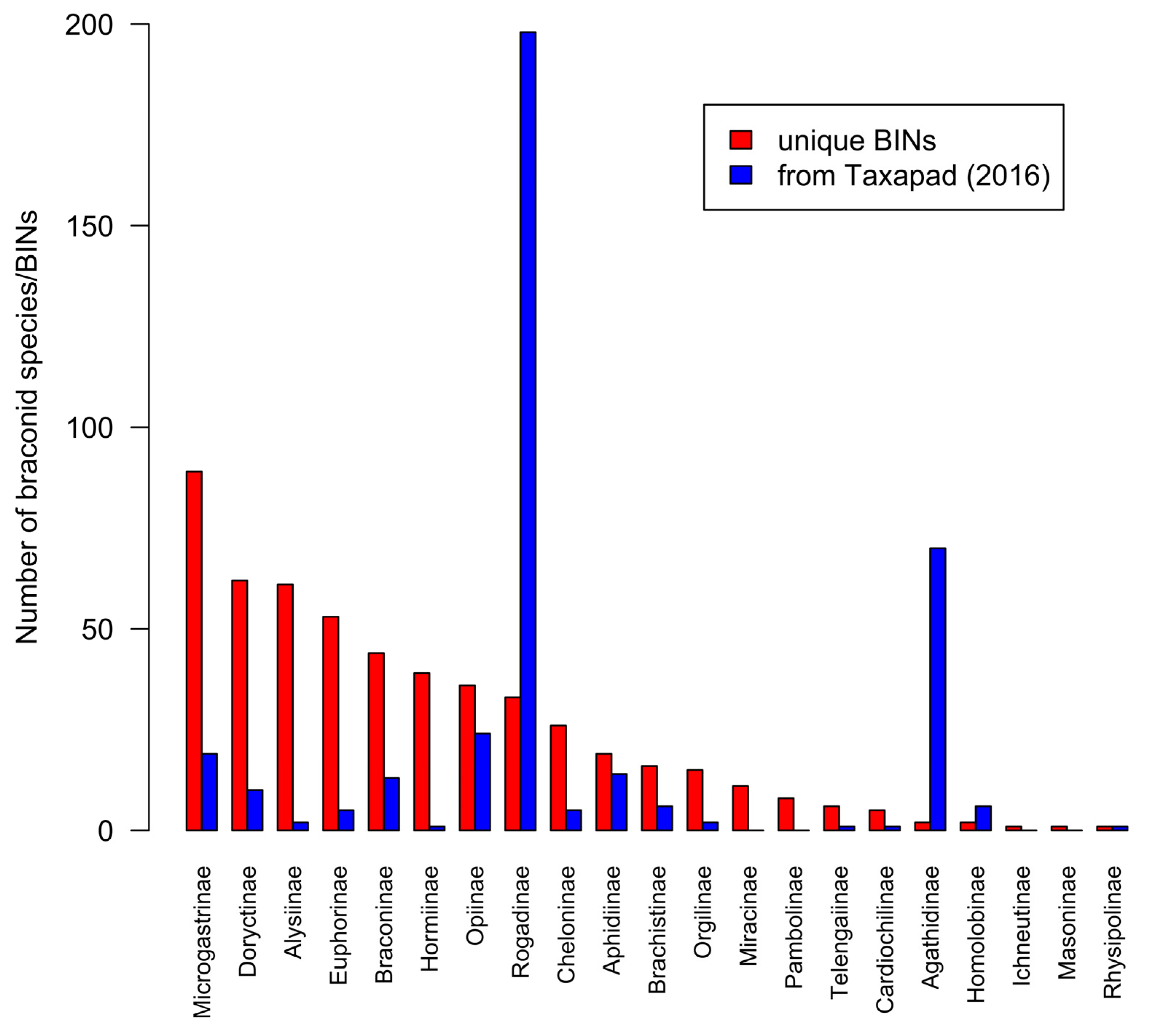
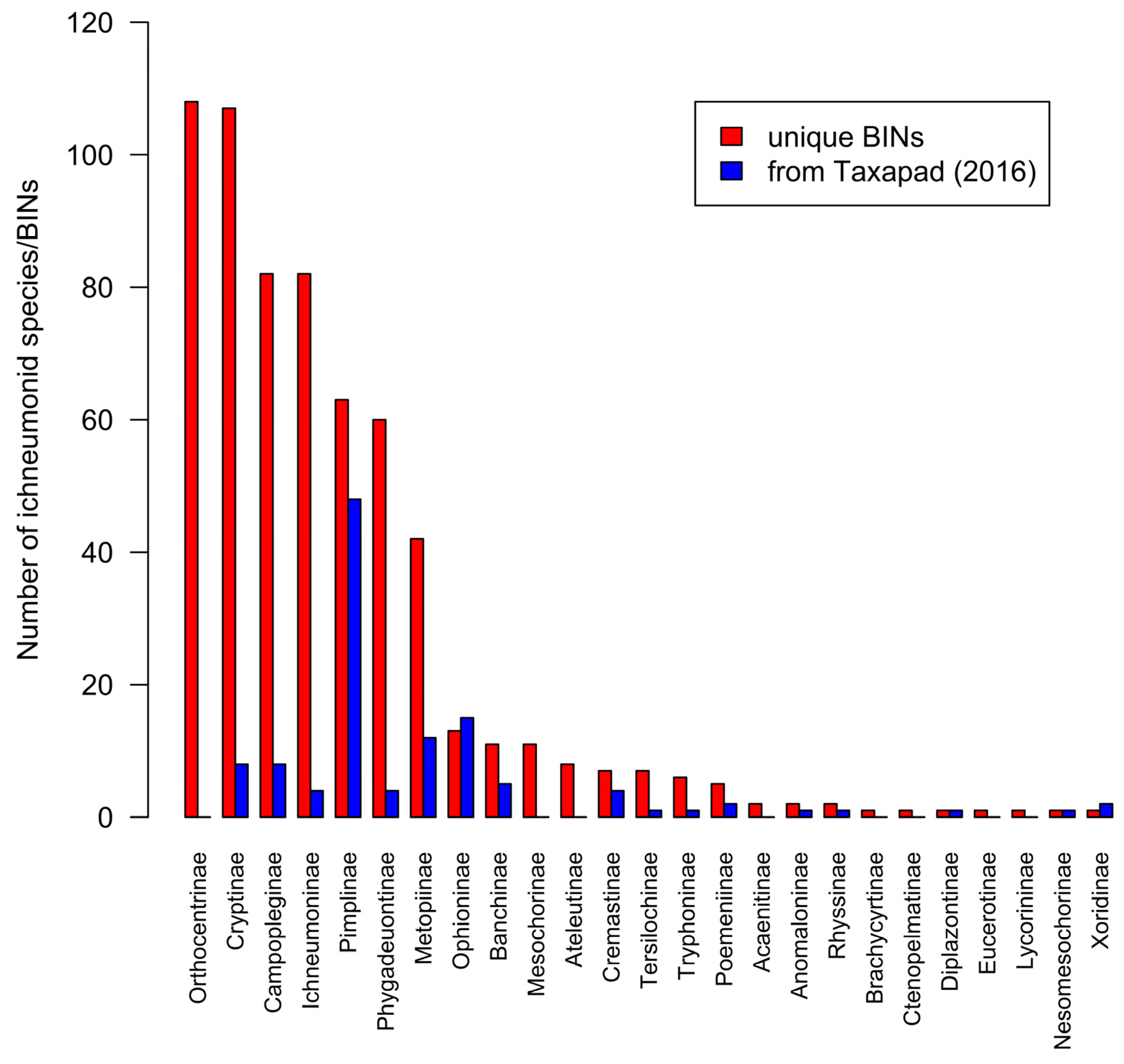
3.6. Ichneumonoidea
3.7. Life History Strategies
4. Discussion
4.1. Species Level Identifications
4.2. Anomalous Diversity of Tropical Ichneumonidae
4.3. Total Thai and Global Parasitoid Wasp Diversity
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Erwin, T.L. Tropical forests: Their richness in Coleoptera and other arthropod species. Coleopt. Bull. 1982, 36, 74–75. [Google Scholar]
- García-Robledo, C.; Kuprewicz, E.K.; Baer, C.S.; Clifton, E.; Hernández, G.G.; Wagner, D.L. The Erwin equation of biodiversity: From little steps to quantum leaps in the discovery of tropical insect diversity. Biotropica 2020, 52, 590–597. [Google Scholar] [CrossRef]
- Gullan, P.J.; Cranston, P.S. The Insects: An Outline of Entomology, 1st ed.; Chapman and Hall: London, UK, 1994. [Google Scholar]
- Gullan, P.J.; Cranston, P.S. The Insects: An Outline of Entomology, 2nd ed.; Blackwell Science: Oxford, UK, 2000. [Google Scholar]
- Stork, N.E. How many species of insects and other terrestrial arthropods are there on Earth. Annu. Rev. Entomol. 2018, 63, 31–45. [Google Scholar] [CrossRef]
- Novotny, V.; Basset, Y.; Miller, S.E.; Weiblen, G.D.; Bremer, B.; Cizek, L.; Drozd, P. Low host specificity of herbivorous insects in a tropical forest. Nature 2002, 416, 841–844. [Google Scholar] [CrossRef] [PubMed]
- Askew, R.R. Parasitic Insects; Heinemann: London, UK, 1971; pp. 1–316. [Google Scholar]
- Godfray, H.C.J. Parasitoids: Behavioral and Evolutionary Ecology; Princeton University Press: Princeton, NJ, USA, 1994; pp. 1–473. [Google Scholar]
- Quicke, D.L.J. Parasitic Wasps; Chapman & Hall: London, UK, 1997; pp. 1–470. [Google Scholar]
- Quicke, D.L.J. The Braconid and Ichneumonid Parasitic Wasps: Biology, Systematics, Evolution and Ecology; Wiley Blackwell: Oxford, UK, 2015; pp. 1–688. [Google Scholar]
- Lees, D.C.; Kawahara, A.Y.; Rougerie, R.; Ohshima, I.; Kawakita, A.; Bouteleux, O.; De Prins, J.; Lopez-Vaamonde, C. DNA barcoding reveals a largely unknown fauna of Gracillariidae leaf-mining moths in the Neotropics. Mol. Ecol. Res 2014, 4, 286–296. [Google Scholar] [CrossRef] [PubMed]
- Zaldívar-Riverón, A.; Martinez, J.J.; Ceccarelli, F.S.; De Jesús Bonilla, V.S.; Rodríguez-Pérez, A.C.; Reséndiz Flores, A.; Smith, M.A. DNA barcoding a highly diverse group of parasitoid wasps (Braconidae: Doryctinae) from a Mexican nature reserve. Mitochond. DNA 2010, 21, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Butcher, B.A.; Smith, M.A.; Sharkey, M.J.; Quicke, D.L.J. A turbo-taxonomic study of Thai Aleiodes (Aleiodes) and Aleiodes (Arcaleiodes) (Hymenoptera: Braconidae: Rogadinae) based largely on COI bar-coded specimens, with rapid descriptions of 179 new species. Zootaxa 2012, 3457, 1–232. [Google Scholar] [CrossRef]
- Kwong, S.; Srivathsan, A.; Meier, R. 2012. An update on DNA barcoding: Low species coverage and numerous unidentified sequences. Cladistics 2012, 28, 639–644. [Google Scholar] [CrossRef]
- Butcher, B.A.; Quicke, D.L.J. Parasitoid Wasps of South East Asia; CABI: Wallingford, UK, 2023; in press. [Google Scholar]
- Rodriguez, J.J.; Fernández-Triana, J.; Smith, M.A.; Janzen, D.H.; Hallwachs, W.; Erwin, T.; Whitfield, J.B. Extrapolations from field studies and known faunas converge on dramatically increased estimates of global microgastrine parasitoid wasp species richness (Hymenoptera: Braconidae). Insect Conserv. Divers. 2013, 6, 530–536. [Google Scholar] [CrossRef]
- Ratnasingham, S.; Hebert, P.D.N. A DNA-based registry for all animal species: The Barcode Index Number (BIN) system. PLoS ONE 2013, 8, e66213. [Google Scholar] [CrossRef]
- Rohland, N.; Reich, D. Cost-effective, high-throughput DNA sequencing libraries for multiplexed target capture. Genome Res. 2012, 22, 939–946. [Google Scholar] [CrossRef] [PubMed]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Hebert, P.D.N.; Penton, E.H.; Burns, J.M.; Janzen, D.H.; Hallwachs, W. Ten species in one: DNA barcoding reveals cryptic species in the neotropical skipper butterfly Astraptes fulgerator. Proc. Natl. Acad. Sci. USA 2004, 101, 14812–14817. [Google Scholar] [CrossRef] [PubMed]
- Hebert, P.D.N.; Braukmann, T.W.A.; Prosser, S.W.J.; Ratnasingham, S.; deWaard, J.R.; Ivanova, N.V.; Janzen, D.H.; Hallwachs, W.; Naik, S.; Sones, J.E.; et al. A Sequel to Sanger: Amplicon sequencing that scales. BMC Genom. 2018, 19, 219. [Google Scholar] [CrossRef]
- R Development Core Team: A Language and Environment for Statistical Computing, R Foundation for Statistical Computing, Vienna. 2016. Available online: https://cran.r-project.org (accessed on 30 July 2023).
- Chao, A.; Ma, K.H.; Hsieh, T.C.; Chiu, C.H. Online Program SpadeR (Species-richness Prediction And Diversity Estimation in R). Program and User’s Guide. 2015. Available online: http://chao.stat.nthu.edu.tw/wordpress/software_download/ (accessed on 12 August 2023).
- Chao, A.; Chiu, C. Nonparametric estimation and comparison of species richness. eLS 2016, 2016, 1–11. [Google Scholar]
- Chao, A. Non-parametric estimation of the number of classes in a population. Scand. J. Stat. 1984, 11, 265–270. [Google Scholar]
- Chao, A. Estimating population size for sparse data in capture–recapture experiments. Biometrics 1989, 45, 427–438. [Google Scholar] [CrossRef]
- Chiu, C.H.; Wang, Y.T.; Walther, B.A.; Chao, A. An improved nonparametric lower bound of species richness via a modified Good–Turing frequency formula. Biometrics 2014, 70, 671–682. [Google Scholar] [CrossRef]
- Chao, A.; Lee, S.H. Estimating the number of classes via sample coverage. J. Am. Stat. Assoc. 1992, 87, 210–217. [Google Scholar] [CrossRef]
- Lomolino, M.V. Ecology’s most general, yet protean pattern: The species-area relationship. J. Biogeogr. 2000, 27, 17–26. [Google Scholar] [CrossRef]
- Yu, D.S.; van Achterberg, C.; Horstmann, K. World Ichneumonoidea 2015: Taxonomy, Biology, Morphology, and Distribution; Nepean: Ottawa, ON, Canada, 2016. [Google Scholar]
- Strutzenberger, P.; Brehm, G.; Fiedler, K. DNA barcoding-based species delimination increases species count of Eois (Geometridae) moths in a well-studied tropical mountain forest by up to 50%. Insect Sci. 2011, 18(3), 349–362. [Google Scholar] [CrossRef]
- Hausmann, A.; Godfray, H.C.J.; Huemer, P.; Mutanen, M.; Rougerie, R.; van Nieukerken, E.J.; Ratnasingham, S.; Hebert, P.D.N. Genetic patterns in European geometrid moths revealed by the Barcode Index Number (BIN) system. PLoS ONE 2013, 8, e84518. [Google Scholar] [CrossRef]
- Hebert, P.D.N.; Ratnasingham, S.; Zakharov, E.V.; Telfer, A.C.; Levesque-Beaudin, V.; Milton, M.A.; Pedersen, S.; Jannetta, P.; DeWaard, J.R. Counting animal species with DNA barcodes: Canadian insects. Philos. Trans. R. Soc. Lond., B, Biol. Sci. 2016, 371, 20150333. [Google Scholar] [CrossRef]
- Costa, F.O.; Antunes, P.M. The contribution of the Barcode of Life initiative to the discovery and monitoring of Biodiversity. In Natural Resources, Sustainability and Humanity—A Comprehensive View; Mendonça, A., Cunha, A., Chakrabarti, R., Eds.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 37–68. [Google Scholar]
- Sharkey, M.J.; Janzen, D.H.; Hallwachs, W.; Chapman, E.G.; Smith, M.A.; Dapkey, T.; Brown, A.; Ratnasingham, S.; Naik, S.; Manjunath, R.; et al. Minimalist revision and description of 403 new species in 11 subfamilies of Costa Rican braconid parasitoid wasps, including host records for 219 species. ZooKeys 2021, 1013, 1–665. [Google Scholar]
- Zamani, A.; Vahtera, V.; Sääksjärvi, I.E.; Scherz, M.D. The omission of critical data in the pursuit of “revolutionary” methods to accelerate the description of species. Syst. Entomol. 2021, 46, 1–4. [Google Scholar] [CrossRef]
- Sharkey, M.J.; Brown, B.; Baker, A.; Mutanen, M. Response to Zamani et al. (2020): The omission of critical data in the pursuit of “revolutionary” methods to accelerate the description of species. ZooKeys 2021, 1033, 191–201. [Google Scholar] [CrossRef]
- Meier, R.; Blaimer, B.B.; Buenaventura, E.; Hartop, E.; von Rintelen, T.; Srivathsan, A.; Yeo, D. A re-analysis of the data in Sharkey et al.’s (2021) minimalist revision reveals that BINs do not deserve names, but BOLD Systems needs a stronger commitment to open science. Cladistics 2021, 38, 264–275. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Triana, J.L. Turbo taxonomy approaches: Lessons from the past and recommendations for the future based on the experience with Braconidae (Hymenoptera) parasitoid wasps. ZooKeys 2022, 1087, 199. [Google Scholar] [CrossRef]
- Thormann, B.; Ahrens, D.; Marín Armijos, D.; Peters, M.K.; Wagner, T.; Wägele, J.W. Exploring the leaf beetle fauna (Coleoptera: Chrysomelidae) of an Ecuadorian mountain forest using DNA barcoding. PLoS One 2016, 11, e0148268. [Google Scholar] [CrossRef]
- Chimeno, C.; Hausmann, A.; Schmidt, S.; Raupach, M.J.; Doczkal, D.; Baranov, V.; Hübner, J.; Höcherl, A.; Albrecht, R.; Jaschhof, M.; et al. Peering into the darkness: DNA barcoding reveals surprisingly high diversity of unknown species of Diptera (Insecta) in Germany. Insects 2022, 13, 82. [Google Scholar] [CrossRef]
- Kilian, I.C.; Espeland, M.; Mey, W.; Wowor, D.; Hadiaty, R.K.; von Rintelen, T.; Herder, F. DNA barcoding unveils a high diversity of caddisflies (Trichoptera) in the Mount Halimun Salak National Park (West Java; Indonesia). PeerJ 2022, 10, e14182. [Google Scholar] [CrossRef] [PubMed]
- Santos, B.F. Phylogeny and reclassification of Cryptini (Hymenoptera, Ichneumonidae, Cryptinae), with implications for ichneumonid higher-level classification. Syst. Entomol. 2017, 42, 650–676. [Google Scholar] [CrossRef]
- Jasso-Martinez, J.M.; Belokobylskij, S.A.; Zaldivar-Riveron, A. Molecular phylogenetics and evolution of generic diagnostic morphological features in the doryctine wasp tribe Rhaconotini (Hymenoptera: Braconidae). Zool. Anz. 2019, 279, 164–171. [Google Scholar] [CrossRef]
- Santos, B.F.; Alvarado, M.; Sääksjärvi, I.E.; van Noort, S.; Villemant, C.; Brady, S.G. Molecular phylogeny of Ateleutinae (Hymenoptera: Ichneumonidae): Systematics and biogeography of a widespread parasitoid wasp lineage. Zool. J. Linn. Soc. 2019, 185, 1057–1078. [Google Scholar] [CrossRef]
- Chen, H.; Lahey, Z.; Talamas, E.J.; Valerio, A.A.; Popovici, O.A.; Musetti, L.; Klompen, H.; Polaszek, A.; Masner, L.; Austin, A.D.; et al. An integrated phylogenetic reassessment of the parasitoid superfamily Platygastroidea (Hymenoptera: Proctotrupomorpha) results in a revised familial classification. Syst. Entomol. 2021, 46, 1088–1113. [Google Scholar] [CrossRef]
- Spasojevic, T.; Broad, G.R.; Sääksjärvi, I.E.; Schwarz, M.; Ito, M.; Korenko, S.; Klopfstein, S. Mind the outgroup and bare branches in total-evidence dating: A case study of pimpliform Darwin wasps (Hymenoptera, Ichneumonidae). Syst. Biol. 2021, 70, 322–339. [Google Scholar] [CrossRef] [PubMed]
- Quicke, D.L.J.; Fagan-Jeffries, E.P.; Jasso-Martinez, J.M.; Smith, M.A.; Hebert, P.D.N.; Hrcek, J.; Miller, S.; Sharkey, M.J.; Shaw, S.R.; Butcher, B.A. A molecular phylogeny of the parasitoid wasp subfamily Rogadinae (Ichneumonoidea: Braconidae) with descriptions of three new genera. Syst. Entomol. 2021, 46, 1019–1044. [Google Scholar] [CrossRef]
- Shimizu, S.; Broad, G.R.; Maeto, K. Integrative taxonomy and analysis of species richness patterns of nocturnal Darwin wasps of the genus Enicospilus Stephens (Hymenoptera, Ichneumonidae, Ophioninae) in Japan. Zookeys 2020, 990, 1–144. [Google Scholar] [CrossRef]
- Watanabe, K.; Matsumoto, R. Revision of the genus Xanthopimpla Saussure 1892 (Hymenoptera, Ichneumonidae, Pimplinae) from Japan. Mitteilungen Aus Dem Mus. Für Naturkunde Berlin. Dtsch. Entomol. Z. 2021, 68, 269–297. [Google Scholar] [CrossRef]
- Zhou, Q.-S.; Polaszek, A.; Qin, Y.-G.; Yu, F.; Wang, X.-B.; Wu, S.-A. Parasitoid-host associations of the genus Coccophagus (Hymenoptera: Aphelinidae) in China. Zool. J. Linn. Soc. 2018, 182, 38–49. [Google Scholar] [CrossRef]
- Dong, Z.; Liu, S.; Zhang, Z. Efficacy of using DNA barcoding to identify parasitoid wasps of the melon-cotton aphid (Aphis gossypii) in watermelon cropping system. Biocontrol 2018, 63, 677–685. [Google Scholar] [CrossRef]
- Triapitsyn, S.V.; Adachi-Hagimori, T.; Rugman-Jones, P.F.; Kado, N.; Sawamura, N.; Narai, Y. Egg parasitoids of Arboridia apicalis (Nawa 1913) (Hemiptera, Cicadellidae), a leafhopper pest of grapevines in Japan, with description of a new species of Anagrus Haliday 1833 (Hymenoptera, Mymaridae). Zookeys 2020, 945, 129–152. [Google Scholar] [CrossRef]
- Ozdemir, I.O.; Tuncer, C.; Tortorici, F.; Ozer, G. Egg parasitoids of green shield bug, Palomena prasina L. (Hemiptera: Pentatomidae) in hazelnut orchards of Turkey. Biocontrol Sci. Technol. 2023, 33, 61–75. [Google Scholar] [CrossRef]
- deWaard, J.R.; Ratnasingham, S.; Zakharov, E.V.; Borisenko, A.V.; Steinke, D.; Telfer, A.C.; Perez, K.H.J.; Sones, J.E.; Young, M.R.; Levesque-Beaudin, V.; et al. A reference library for Canadian invertebrates with 1.5 million barcodes, voucher specimens, and DNA samples. Sci. Data 2019, 6, 308. [Google Scholar] [CrossRef]
- Roslin, T.; Somervuo, P.; Pentinsaari, M.; Hebert, P.D.; Agda, J.; Ahlroth, P.; Anttonen, P.; Aspi, J.; Blagoev, G.; Blanco, S.; et al. A molecular-based identification resource for the arthropods of Finland. Mol. Ecol. Resour. 2022, 22, 803–822. [Google Scholar] [CrossRef] [PubMed]
- Awad, J.; Vasilita, C.; Wenz, S.; Alkarrat, H.; Zimmermann, O.; Zebitz, C.; Krogmann, L. New records of German Scelionidae (Hymenoptera: Platygastroidea) from the collection of the State Museum of Natural History Stuttgart. Biodivers. Data J. 2021, 9, e69856. [Google Scholar] [CrossRef] [PubMed]
- Murillo Pacheco, H.; Vanlaerhoven, S.L.; Marcos García, M.Á.; Hunt, D.W. Food web associations and effect of trophic resources and environmental factors on parasitoids expanding their host range into non-native hosts. Entomol. Exp. Et Appl. 2018, 166, 277–288. [Google Scholar] [CrossRef]
- Avalos, S.; González, E.; Mangeaud, A.; Valladares, G. Caterpillar-parasitoid food webs and biological control in two extensive crops. Biol. Control 2020, 143, 104184. [Google Scholar] [CrossRef]
- Sharkey, M.J.; Clutts, S.A. A revision of Thai Agathidinae (Hymenoptera, Braconidae), with descriptions of six new species. J. Hymenopt. Res. 2010, 22, 69–132. [Google Scholar]
- Sharkey, M.J.; Stoelb, S.A.C. Revision of Therophilus s.s. (Hymenoptera, Braconidae, Agathidinae) from Thailand. J. Hymenopt. Res. 2012, 27, 1–36. [Google Scholar]
- Sharkey, M.J.; Stoelb, S.A.C. Revision of Zelodia (Hymenoptera, Braconidae, Agathidinae) from Thailand. J. Hymenopt. Res. 2012, 26, 31–71. [Google Scholar] [CrossRef]
- Sharkey, M.J.; Stoelb, S.A.C. Revision of Agathacrista new genus (Hymenoptera, Braconidae, Agathidinae, Agathidini). J. Hymenopt. Res. 2013, 33, 99–112. [Google Scholar] [CrossRef]
- Karlsson, D.; Pape, T.; Johanson, K.A.; Liljeblad, J.; Ronquist, F. The Swedish Malaise Trap Project, or how many species of Hymenoptera and Diptera are there in Sweden? Entomol. Tidskr. 2005, 126, 43–53. [Google Scholar]
- Hendrich, L.; Morinière, J.; Haszprunar, G.; Hebert, P.D.N.; Hausmann, A.; Köhler, F.; Balke, M. A comprehensive DNA barcode database for Central European beetles with a focus on Germany: Adding more than 3500 identified species to BOLD. Mol. Ecol. Resour. 2014, 15, 795–818. [Google Scholar] [CrossRef]
- Wirta, H.; Várkonyi, G.; Rasmussen, C.; Kaartinen, R.; Schmidt, N.M.; Hebert, P.D.N.; Barták, M.; Blagoev, G.; Disney, H.; Ertl, S.; et al. Establishing a community-wide DNA barcode library as a new tool for arctic research. Mol. Ecol. Resour. 2015, 16, 809–822. [Google Scholar] [CrossRef]
- Geiger, M.; Moriniere, J.; Hausmann, A.; Haszprunar, G.; Wägele, W.; Hebert, P.; Rulik, B. Testing the Global Malaise Trap Program—How well does the current barcode reference library identify flying insects in Germany? Biodivers. Data J. 2016, 4, e10671. [Google Scholar] [CrossRef] [PubMed]
- Owen, D.F.; Owen, J. Species diversity in temperate and tropical Ichneumonidae. Nature 1974, 249, 583–584. [Google Scholar] [CrossRef]
- Janzen, D.H. Interactions of seeds and their insect predators/parasitoids in a tropical deciduous forest. In Evolutionary Strategies of Parasitic Insects and Mites; Price, P.W., Ed.; Plenum: New York, NY, USA, 1975; pp. 154–186. [Google Scholar]
- Janzen, D.H.; Pond, C.M. A comparison by sweep sampling of the arthropod fauna of secondary vegetation in Michigan England and Costa Rica. Trans. R. Ent. Soc. Lond. 1975, 127, 33–50. [Google Scholar] [CrossRef]
- Rathcke, B.J.; Price, P.W. Anomalous diversity of tropical ichneumonid parasitoids: A predation hypothesis. Am. Nat. 1976, 110, 889–893. [Google Scholar]
- Gauld, I.D.; Gaston, K.J.; Janzen, D.H. Plant allelochemicals, tritrophic interactions and the anomalous diversity of tropical parasitoids: The “nasty” host hypothesis. Oikos 1992, 65, 353–357. [Google Scholar] [CrossRef]
- Sime, K.R.; Brower, A.V.Z. Explaining the latitudinal gradient anomaly in ichneumonid species-richness: Evidence from butterflies. J. Anim. Ecol. 1998, 67, 387–399. [Google Scholar] [CrossRef]
- Quicke, D.L.J. We know too little about parasitoid wasp distributions to draw any conclusions about latitudinal trends in species richness, body size and biology. PLoS ONE 2012, 7, e32101. [Google Scholar] [CrossRef]
- Veijalainen, A.; Wahlberg, N.; Broad, G.R.; Erwin, T.L.; Longino, J.T.; Sääksjärvi, I.E. Unprecedented ichneumonid parasitoid wasp diversity in tropical forests. Proc. R. Soc. B: Biol. Sci. 2012, 279, 4694–4698. [Google Scholar] [CrossRef]
- Veijalainen, A.; Sääksjärvi, I.E.; Erwin, T.L.; Gomez, I.C.; Longino, J.T. Subfamily composition of Ichneumonidae (Hymenoptera) from western Amazonia: Insights into diversity of tropical parasitoid wasps. Insect Conserv. Divers. 2013, 6, 28–37. [Google Scholar] [CrossRef]
- Veijalainen, A.; Broad, G.R.; Sääksjärvi, I.E. Twenty seven new species of Orthocentrus (Hymenoptera: Ichneumonidae; Orthocentrinae) with a key to the Neotropical species of the genus. Zootaxa 2014, 3768, 201–252. [Google Scholar] [CrossRef] [PubMed]
- Broad, G.R.; Shaw, M.R.; Fitton, M.G. Ichneumonid wasps (Hymenoptera: Ichneumonidae): Their classification and biology. Handb. Identif. Br. Insects 2018, 7, 1–418. [Google Scholar]
- Songvorawit, N.; Quicke, D.L.J.; Butcher, B.A. Taxonomic progress and diversity of ichneumonoid wasps (Hymenoptera: Ichneumonoidea) in Southeast Asia. Trop. Nat. Hist. 2021, 21, 79–93. [Google Scholar]
- Gauld, I.D.; Mitchell, P.A. The Taxonomy, Distribution and Host Preferences of Indo-Papuan Parasitic Wasps of the Subfamily Ophioninae; CABI: Slough, UK, 1981. [Google Scholar]
- Plant, A.R.; Surin, C.; Saokhod, R.; Srisuka, W. Higher taxon diversity, abundance and community structure of Empididae, Hybotidae and Brachystomatidae (Diptera: Empidoidea) in tropical forests—results of mass-sampling in Thailand. Stud. Dipterol. 2012, 18, 121–149. [Google Scholar]
- Saunders, T.E.; Ward, D.F. Variation in the diversity and richness of parasitoid wasps based on sampling effort. PeerJ 2018, 5, e4642. [Google Scholar] [CrossRef]
- Smith, M.A.; Fernandez-Triana, J.; Roughley, R.; Hebert, P.D.N. DNA barcode accumulation curves for understudied taxa and areas. Mol. Ecol. Resour. 2009, 9, 208–216. [Google Scholar] [CrossRef]
- Wallace, A.R. The Geographical Distribution of Animals, with a Study of the Relations of Living and Extinct Faunas As Elucidating the Past Changes of the Earth’s Surface, 1st ed.; Harper & Brothers Publishers: New York, NY, USA, 1876. [Google Scholar]
- Hughes, J.B.; Round, P.D.; Woodruff, D.S. The Indochinese-Sundaic faunal transition at the Isthmus of Kra: An analysis of resident forest bird species distributions. J. Biogeogr. 2003, 30, 569–580. [Google Scholar] [CrossRef]
- Hughes, A.C.; Satasook, C.; Bates, P.J.; Bumrungsri, S.; Jones, G. Explaining the causes of the zoogeographic transition around the Isthmus of Kra: Using bats as a case study. J. Biogeogr. 2011, 38, 2362–2372. [Google Scholar] [CrossRef]
- Woodruff, D.S.; Turner, L.M. The Indochinese-Sundaic zoogeographic transition: A description and analysis of terrestrial mammal species distributions. J. Biogeogr. 2009, 36, 803–821. [Google Scholar] [CrossRef]
- Dejtaradol, A.; Renner, S.C.; Karapan, S.; Bates, P.J.J.; Moyle, R.G.; Päckert, M. Indochinese-Sundaic faunal transition and phylogeographical divides north of the Isthmus of Kra in Southeast Asian bulbuls (Aves: Pycnonotidae). J. Biogeogr. 2012, 43, 471–483. [Google Scholar] [CrossRef]
- Gostel, M.R.; Kress, W.J. The expanding role of DNA barcodes: Indispensable tools for ecology, evolution, and conservation. Diversity 2022, 14, 213. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Trap Number | Trap Code | Forest Type | Latitude | Longitude | Elevation (m) |
|---|---|---|---|---|---|---|
| Khao Yai N.P. | 1 | THAMA | Dry evergreen forest | 14°28.256′ N | 101°22.396′ E | 779 |
| Khao Yai N.P. | 2 | THAMB | Dry evergreen forest | 14°26.800′ N | 101°22.017′ E | 752 |
| Khao Yai N.P. | 3 | THAMC | Secondary forest | 14°26.016′ N | 101°22.153′ E | 732 |
| Khao Yai N.P. | 4 | THAMD | Secondary forest | 14°25.548′ N | 101°23.078′ E | 697 |
| Sakaerat | 1 | THAME | Dry dipterocarp forest | 14°30.336′ N | 101°56.147′ E | 353 |
| Sakaerat | 2 | THAMF | Dry dipterocarp + dry evergreen forest ecotone | 14°30.580′ N | 101°55.980′ E | 365 |
| Sakaerat | 3 | THAMG | Dry evergreen forest | 14°30.158′ N | 101°55.481′ E | 449 |
| Doi Phu Kha N.P. | 1 | THAMH | Hill evergreen forest | 19°12.236′ N | 101°04.667′ E | 1341 |
| Doi Phu Kha N.P. | 2 | THAMI | Hill evergreen forest | 19°12.157′ N | 101°04.388′ E | 1327 |
| Doi Phu Kha N.P. | 3 | THAMJ | Hill evergreen forest | 19°12.311′ N | 101°04.846′ E | 1356 |
| Doi Phu Kha N.P. | 4 | THAMK | Hill evergreen forest | 19°10.447′ N | 101°06.368′ E | 1698 |
| Location | Trap | 28.i | 25.ii | 25.iii | 22.iv | 20.v | 17.vi | 15–16.vii | 12–13.viii | 10.ix | 7.x |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Khao Yai N.P. | 1 | 590 | 588 | 142 | 212 | 133 | 86 | 61 | 86 | - | - |
| Khao Yai N.P. | 2 | - | - | 232 | 179 | 176 | 151 | 129 | 99 | - | - |
| Khao Yai N.P. | 3 | - | 5 | 323 | 357 | 406 | 323 | 17 | 6 | - | - |
| Khao Yai N.P. | 4 | - | 9 | 869 | 701 | 528 | 446 | 8 | 158 | - | - |
| Sakaerat | 1 | - | 134 | 471 | 103 | 429 | 142 | 386 | 457 | - | - |
| Sakaerat | 2 | - | 91 | 264 | 251 | 362 | 194 | 462 | 219 | - | - |
| Sakaerat | 3 | - | 64 | 242 | 275 | 366 | 249 | 279 | 395 | - | - |
| Doi Phu Kha N.P. | 1 | - | - | - | - | - | - | 39 | 61 | 138 | 102 |
| Doi Phu Kha N.P. | 2 | - | - | - | - | - | - | 507 | 81 | 456 | 474 |
| Doi Phu Kha N.P. | 3 | - | - | - | - | - | - | 324 | 123 | 564 | 623 |
| Doi Phu Kha N.P. | 4 | - | - | - | - | - | - | 1373 | 63 | 48 | 68 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quicke, D.L.J.; Hebert, P.D.N.; Pentinsaari, M.; Butcher, B.A. Barcoding Hymenoptera: 11 Malaise Traps in Three Thai Forests: The First 68 Trap Weeks and 15,338 Parasitoid Wasp Sequences. Forests 2023, 14, 1991. https://doi.org/10.3390/f14101991
Quicke DLJ, Hebert PDN, Pentinsaari M, Butcher BA. Barcoding Hymenoptera: 11 Malaise Traps in Three Thai Forests: The First 68 Trap Weeks and 15,338 Parasitoid Wasp Sequences. Forests. 2023; 14(10):1991. https://doi.org/10.3390/f14101991
Chicago/Turabian StyleQuicke, Donald L. J., Paul D. N. Hebert, Mikko Pentinsaari, and Buntika A. Butcher. 2023. "Barcoding Hymenoptera: 11 Malaise Traps in Three Thai Forests: The First 68 Trap Weeks and 15,338 Parasitoid Wasp Sequences" Forests 14, no. 10: 1991. https://doi.org/10.3390/f14101991